 Jingyu Wang
Jingyu Wang Wang Xing1,3
Wang Xing1,3 Dali Liu
Dali Liu- 1National Beet Medium-Term Gene Bank, Heilongjiang University, Harbin, China
- 2Key Laboratory of Beet Genetics and Breeding, College of Modern Agriculture and Ecological Environment, Heilongjiang University, Harbin, China
- 3China National Seed Group Co., Ltd., Sanya, China
- 4Heilongjiang Climate Center, Harbin, China
- 5Heilongjiang Ecological Meteorological Center, Harbin, China
Introduction: Corylus heterophylla is native to East Asia, including northern and central China, southeastern Siberia, eastern Mongolia, Korea, Japan, and other adjacent regions, and its geographical distribution is highly sensitive to climate change. Investigating shifts in its suitable habitat under 1970-2000 and future climate conditions is crucial for the conservation and sustainable utilization of its germplasm resources.
Methods: This study employed an optimized MaxEnt model, integrating species occurrence records with multiple environmental variables, to simulate and analyze potential suitable habitats and their key environmental determinants under various climate scenarios.
Results: The results demonstrated that the model configured with the feature combination (FC) of LQPH and a regularization multiplier (RM) of 3 achieved low complexity, minimal overfitting, and high predictive accuracy (AUC = 0.933). The dominant factors influencing the distribution were identified as Bio16 (Precipitation of wettest quarter, 39.5%), Bio9 (Mean temperature of driest quarter, 22.2%), Alt (Altitude, 16.2%), and Bio3 (Isothermality, 7.1%). The 1970-2000 climatically suitable area for C. heterophylla spans approximately 210.85 × 104 km2, accounting for 21.96% of China’s total land area. Projections under future climate scenarios indicate that the suitable habitat area for C. heterophylla will decrease slightly, primarily in low suitability zones, while high and medium suitability zones will expand. Its distribution pattern is expected to shift significantly northward while contracting southward, with the distribution centroid moving toward higher latitudes.
Discussion: These findings provide a scientific basis for conserving and sustainably utilizing C. heterophylla under climate change.
1 Introduction
Global climate change significantly impacts biodiversity and potential ecological responses, making this a major research focus (Trew and Maclean, 2021). These responses encompass a range of phenomena, including shifts in species distribution, alterations in phenology, and the restructuring of ecological communities, which ultimately threaten ecosystem stability (Rubenstein et al., 2023). Climate-driven alterations in species geographical distributions exacerbate biodiversity and germplasm resource loss, and accelerate extinction rates (Bellard et al., 2012). The Intergovernmental Panel on Climate Change (IPCC) Sixth Assessment Report (AR6) highlights that ecological risks from global warming may cause irreversible impacts on ecosystems and human societies (Kuang et al., 2022). Many evidence indicates climate warming reduces species’ habitat ranges (Lenoir et al., 2020), frequently driving migration towards higher altitudes and latitudes (Chen et al., 2011). Consequently, assessing the potential distribution of species under climate change is critical for biodiversity conservation and climate adaptation. Predicting future changes in suitable habitats not only offers a scientific foundation for developing proactive biodiversity strategies and enhancing ecosystem resilience, but also provides theoretical support for the preservation of germplasm resources and the design of climate-informed ecological management practices.
Corylus heterophylla Fisch. ex Trautv. (C. heterophylla), an ecologically and economically significant species within Corylus (Betulaceae), dominates northeastern, northwestern, and northern China’s mountainous forests (Cheng et al., 2019). Its extensive root system mitigates soil erosion, contributing to vegetation restoration and ecosystem rehabilitation (Wang et al., 2025). Current research on C. heterophylla emphasizes phenotypic traits (Fan et al., 2020), genetic diversity (Yang et al., 2023), and cultivation techniques (He et al., 2023), with limited focus on its ecological niche requirements. Previous research on Corylus distribution has often examined C.xheterophylla as part of broader species groups. For instance, Liu and Chen (2024) modeled the potential distribution of 18 hazel species across China under current and future climate scenarios, integrating climatic, topographic, and anthropogenic variables. In a related effort, He et al. (2023) reconstructed the historical distribution changes of the C. heterophylla complex since the Last Interglacial. While these studies offer valuable macro-scale insights, the ecological distinctiveness of C. heterophylla may not be fully resolved in multi-species or complex-level analyses due to potential niche overlap among congeners. Species-specific distribution modeling helps minimize such interspecific interference and is essential for accurately capturing a target species’ unique climatic responses and spatial patterns. It also forms the basis for developing tailored conservation and sustainable management strategies. Therefore, this study focuses exclusively on C. heterophylla to clarify its niche characteristics and response mechanisms to critical environmental factors—addressing an important gap in fine-scale, single-species distribution modeling under future climate scenarios.
Species Distribution Models (SDMs) are widely used to study species–environment relationships and predict geographical distributions (Zurell et al., 2020). Common algorithms include CLIMEX (climate explorer) (Sutherst and Maywald, 1985), MaxEnt (Phillips et al., 2006), and BIOMAPPER (biogeographical mapping and modelling) (Hirzel and Guisan, 2002). These models apply statistical methods to correlate species occurrence with environmental conditions, generating habitat suitability indices based on niche theory (Zurell et al., 2020). Among them, MaxEnt has been widely applied in species distribution modeling due to its low data requirements, high predictive stability, and strong interpretability (Brown et al., 2017). The model’s primary strength lies in its ability to construct high-performance and robust species distribution models using presence-only data (Valavi et al., 2022). The model also supports quantitative evaluation of environmental variable contributions, allows rigorous assessment of variable importance through jackknife tests, and enables reliable validation of predictive performance using receiver operating characteristic (ROC) curves and the area under the curve (AUC) (Phillips et al., 2006). Furthermore, parameter optimization approaches (Chen et al., 2025; Fianchini et al., 2025) can enhance the model by reducing overfitting and improving generalizability and reliability (Guisan et al., 2013). With the capacity to incorporate future climate scenarios, MaxEnt can systematically assess potential climate change impacts on habitat suitability (Elith and Leathwick, 2009). Given these advantages, MaxEnt is well-suited for species with restricted ranges and complex niches, and was therefore selected as the modeling framework in this study.
In this study, we employed an optimized MaxEnt model with C. heterophylla occurrence data and environmental variables (climate, soil, topography) to predict current and future suitable habitats (Lee et al., 2021). Specific objectives include: (1) identify key environmental drivers governing C. heterophylla distribution and quantify their relative contributions; (2) classify habitat suitability levels and analyze spatial patterns of the species; (3) project future changes in suitable habitat area and range shifts under multiple climate scenarios (SSP1-2.6, SSP2-4.5, SSP5-8.5), elucidating the species’ distributional response mechanisms to climate change for evidence-based conservation strategies.
2 Materials and methods
2.1 Occurrence data collection and processing
C. heterophylla occurrence records across China were compiled from authoritative sources: the Global Biodiversity Information Facility (GBIF; https://www.gbif.org/), Chinese Virtual Herbarium (CVH; https://www.cvh.ac.cn/), Plant Photo Bank of China (PPBC; https://ppbc.iplant.cn/), and published literature (He et al., 2023; Liu and Chen, 2024). Records lacking precise coordinates were georeferenced using Baidu Map Coordinate Picker (https://api.map.baidu.com/lbsapi/getpoint/index.html) based on textual descriptions, and corresponding uncertainties were estimated according to the specificity of each location description. To mitigate the effects of spatial autocorrelation and sampling bias inherent in herbarium and literature-based data (Taylor et al., 2020), the dataset was rigorously cleaned by removing duplicate and ambiguous records. Spatial thinning was then performed using the R package spThin (Allouche et al., 2006), with a filtering distance of 5 km to ensure that only one occurrence record was retained per 5 ×5 km grid cell (Wang et al., 2024). The final dataset comprised 243 spatially independent occurrence records (Figure 1), which were formatted into a CSV file for input into the MaxEnt model.

Figure 1. Occurrence points of C. heterophylla in China.
2.2 Environmental variable selection
Thirty-one environmental variables potentially influencing the distribution of C. heterophylla were initially selected, comprising 19 bioclimatic variables (under CMIP6 SSPs scenarios) (Urban et al., 2016), 9 soil parameters, and 3 topographic factors. In this study, soil parameters and topographic factors are generally considered to remain constant under future climate projections. To reveal its core climatic niche, the model excludes disturbance factors such as human activities, thereby simulating potential suitable distribution areas under future climate change (Ren et al., 2025; Shan et al., 2025). 1970–2000 climate data and future climate projections for the 2050s (2041-2060) and 2090s (2081-2100) were obtained from WorldClim (version 2.1; http://www.worldclim.org) at 2.5 arc-minute resolution. Future climate projections were derived from the BCC-CSM2-MR model developed by the China Meteorological Administration (CMA), considering three Shared Socioeconomic Pathways (SSPs): SSP1-2.6 (low radiative forcing), SSP2-4.5 (intermediate), and SSP5-8.5 (high) (Wang et al., 2024). Soil data came from the Harmonized World Soil Database (HWSD v1.2; Cold and Arid Regions Scientific Data Center), while topographic data were obtained from Geospatial Data Cloud (http://www.gscloud.cn). All topographic and soil datasets were resampled to a 2.5 arc-minute resolution using ArcGIS 10.8 to ensure consistency with the climatic data layers.
To mitigate model overfitting caused by multicollinearity among environmental variables (Wang et al., 2020), we performed a structured variable selection procedure. Initial MaxEnt runs were conducted to evaluate variable importance based on percent contribution and permutation importance. A variable was excluded only if both of these values were exactly 0% and its response curve in the jackknife test fully overlapped with the curve generated using all variables. Environmental factor correlations were analyzed using IBM SPSS Statistics (version 28.0; IBM Corp., Armonk, NY, USA). By combining the contribution rates of variables with correlation matrix analysis, factors exhibiting pairwise correlation with the coefficient |r| > 0.8 underwent selection refinement: the lower-contribution variable in each correlated pair (determined from preliminary analysis) was eliminated (Yi et al., 2016). Further consideration was given to thexbiological plausibility of each variable, evaluating its association with the ecological requirements of C. heterophylla and substantiating findings through ecological theory. For instance, the selected variables are closely linked to the species’ water requirements and temperature adaptability, exhibiting not only statistical significance but also clear biological plausibility. This process yielded nine key environmental factors for final model construction (Table 1).

Table 1. Environmental variables used in MaxEnt modeling.
2.3 Model construction
MaxEnt v3.4.4 was used to predict the suitable habitats for C. heterophylla across three time periods. The model incorporated 243 occurrence points with 25% randomly allocated for testing and 10 replicate runs. Outputs were formatted as logistic probabilities (0 to 1). Environmental variable importance was assessed through jackknife testing, with key factors identified by joint consideration of percent contribution and permutation importance. Model accuracy was evaluated using the Area under the Receiver Operating Characteristic (ROC) Curve (AUC) (Phillips et al., 2006). AUC values (0–1 scale) indicate predictive accuracy: <0.6 (failed), 0.6-0.7 (poor), 0.7-0.8 (fair), 0.8-0.9 (good), and 0.9-1.0 (excellent) (Zhang et al., 2024).
2.4 Model optimization
Default MaxEnt parameters may produce overly complex models prone to overfitting (Morales et al., 2017). In this study, the regularization multiplier (RM) and feature combination (FC) in the MaxEnt model were optimized using R’s Kuenm package (Cobos et al., 2019). From baseline settings (RM = 1, FC=LQHPT), RM was tested across 0.5-4.0 (0.5 increments) with randomized combinations of five feature classes: linear, quadratic, product, threshold, and hinge. C. heterophylla occurrence data were randomly partitioned (75% for training, 25% for testing). Model complexity was tested based on the evaluated using δAICc values and 5% missing rates, where lower δAICc indicates superior predictive performance (Giraldo and Kucuker, 2024).
2.5 Habitat suitability classification
MaxEnt output files (ASC format) were batch-converted to raster format in ArcGIS. Predicted suitability for C. heterophylla was reclassified into four categories based on mean logistic values: unsuitable (0-0.2), low suitability (0.2-0.4), medium suitability (0.4-0.6), and high suitability (0.6-1) (Gu et al., 2021; Xu et al., 2019). Distribution map for 1970-2000, 2050s and 2090s periods were generated using a China basemap. Suitable habitat areas were quantified per period using ArcGIS Spatial Analyst tools.
2.6 Analysis of spatial dynamics and centroid shifts
Based on the potential habitat distribution maps for 1970–2000 and future periods generated by the MaxEnt model, we quantitatively analyzed spatial pattern changes using the Distribution Change Analysis module in SDM toolbox (v2.0). Habitat transitions were classified into four categories (-1: range expansion; 0: species absence; 1: stable habitat; 2: range contraction). Spatial change maps were generated in ArcGIS 10.8. Centroid positions for 1970–2000 and future suitable habitats were calculated using the Centroid Changes (Liners) tool in SDM toolbox (v2.0), with migration distances indicating directional shifts in habitat distribution.
3 Results
3.1 Model performance and validation
The MaxEnt model simulated C. heterophylla potential habitats using 243 occurrence records and nine environmental factors. Default parameters (RM = 1, FC=LQPTH) yielded δAICc=88.48, whereas optimized parameters (RM = 3, FC=LQPH) achieved δAICc=0, indicating superior model performance (Table 2). Ten cross-validations with optimized settings produced a mean AUC of 0.933, confirming high predictive accuracy for habitat suitability (Figure 2).

Table 2. Performance evaluation of MaxEnt models under different parameter settings.

Figure 2. Receiver operating characteristic (ROC) curve for C. heterophylla habitat suitability modeling using MaxEnt.
3.2 Environmental factors affecting the distribution of C. heterophylla
Analysis of regularization training gain, permutation importance, and jackknife tests from the MaxEnt model simulation identified key environmental factors influencing C. heterophylla distribution. Bio16 (Precipitation of wettest quarter) showed highest contribution (39.5%), followed by Bio9 (Mean temperature of driest quarter, 22.2%), Alt (Altitude, 16.2%), and Bio3 (Isothermality, 7.1%), collectively accounting for 85% of total contribution. Permutation importance highlighted Bio16 (41.1%) and Alt (27.6%) as most significant, totaling 68.7% (Table 3; Figure 3). In contrast, the remaining variables—REF Depth (4.0%), Bio19 (3.9%), T_bs (3.5%), Slope (2.3%), and Bio4 (1.2%)made markedly lower contributions, collectively accounting for only 14.9%. Results indicate that precipitation during Bio16 (Precipitation of wettest quarter) constitutes the dominant factor in the model. Alt (Altitude) is identified as a secondary yet critical determinant, while temperature-related variables (Bio9, Bio3) further refine habitat suitability. Together, these form the core ecological drivers for C. heterophylla.

Table 3. Contribution rates and permutation importance of environmental factors governing C. heterophylla distribution.

Figure 3. Jackknife test of regularized training gain for environmental variables.
C. heterophylla occurrence probability followed unimodal responses to environmental gradients (Figure 4). Using probability > 0.5 as threshold (Araújo et al., 2019) the optimal range for Bio16 (Precipitation of the Wettest Quarter) was found to be between 300 mm and 450 mm, with the highest probability (approximately probability = 0.6) occurring at 350 mm. The suitable range for Bio9 (The mean temperature of the driest quarter) was -12°C to 2°C, peaking at -2°C (approximately probability = 0.56). The optimal Alt (Altitude) range was 100 to 700 m, with a peak at 300 m (approximately probability = 0.63). For Bio3 (Isothermality), the suitable range was between 24 and 30, peaking at 28 (approximately probability = 0.57).
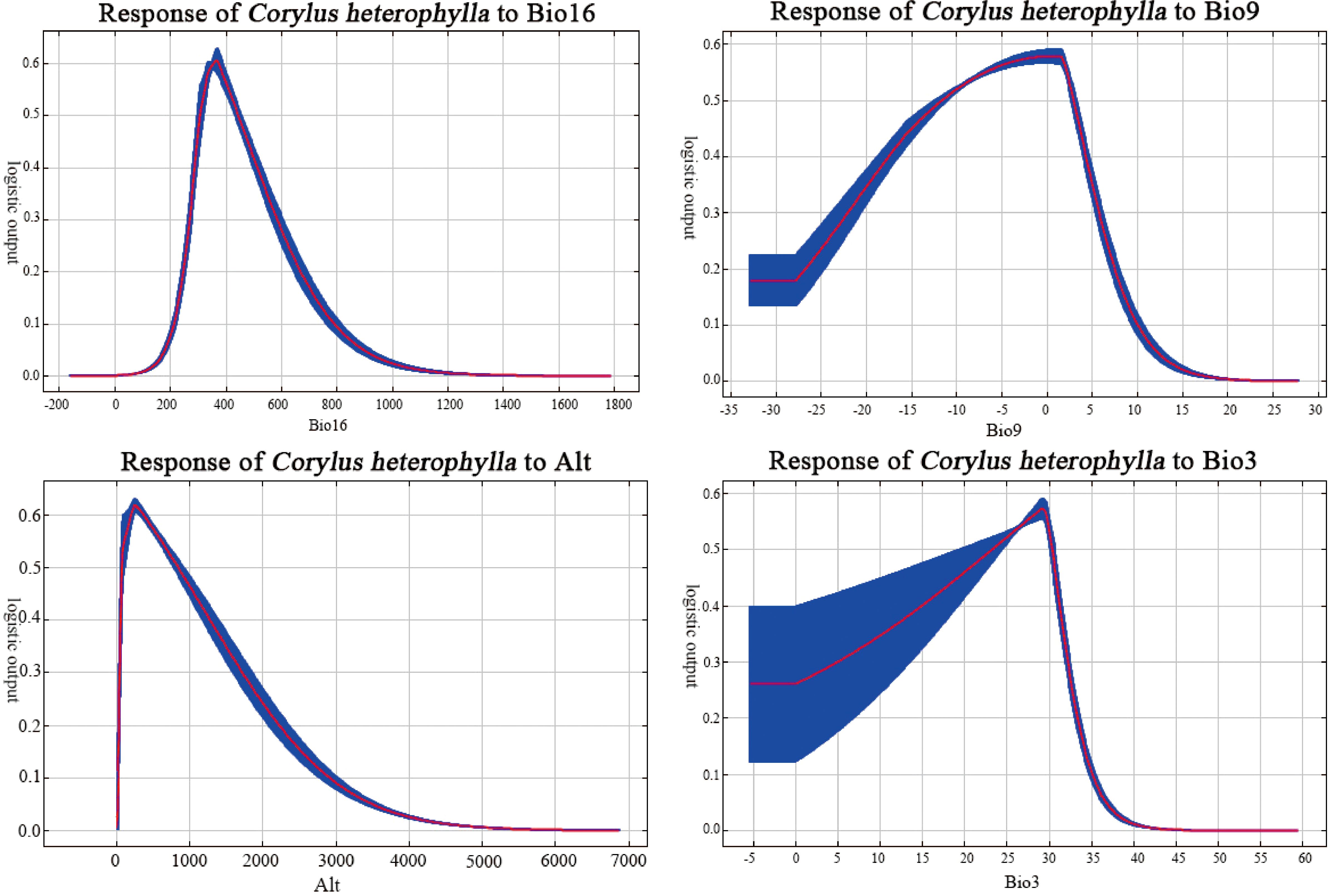
Figure 4. Response curves of C. heterophylla occurrence probability to key environmental factors.
3.3 Projections of C. heterophylla distribution across temporal scales
3.3.1 1970–2000 habitat suitability
Under 1970–2000 conditions (Figure 5), C. heterophylla primarily inhabits northeastern and northern China (Heilongjiang to Hubei; 21.96% of land area). Total suitable habitat spans 210.85 × 104 km2, comprising low-suitability zones (108.37 × 104 km2, 11.29%) concentrated in Henan, Sichuan, Chongqing; medium-suitability zones (63.39 × 104 km2, 6.6%) predominantly in Heilongjiang, Inner Mongolia, Henan, and Gansu; and high-suitability zones (39.09 × 104 km2, 4.07%) focused in Jilin, Liaoning, Hebei, Shanxi (Table 4). A comparison between the observed distribution points of C. heterophylla (Figure 1) and its predicted potential distribution (Figure 5) reveals that the actual occurrences predominantly reside within medium-high suitability areas.

Figure 5. 1970–2000 distribution of C. heterophylla in China.

Table 4. Potential suitable habitat area (×104 km2) for C. heterophylla under 1970–2000 and future climate scenarios.
3.3.2 Future distribution projections
Projections indicate that under all scenarios, areas with low suitability will continue to shrink, while areas with moderate and high suitability will expand (Figure 6; Table 4). By the 2050s: Under SSP1-2.6, low suitability areas decrease by 40.38×104 km², high suitability areas increase by 12.99×104 km², and medium suitability areas increase by 19.79×104 km²; Under SSP2-4.5, low-suitability areas decrease by 44.23×104 km², medium-suitability areas expand by 26.2×104 km², and high-suitability areas expand by 9.91×104 km²; The SSP5-8.5 scenario shows a decrease of 46.95×104 km² in low suitability areas, an increase of 24.74×104 km² in moderate suitability areas, and an expansion of 18.08×104 km² in high suitability areas. By the 2090s, SSP1-2.6 shows a 38.55×104 km² reduction in low suitability zones, a 17.1×104 km² expansion in moderate suitability zones, and an 11.98×104 km² expansion in high suitability zones; SSP2-4.5 shows a 48.06×104 km² loss in low-intensity areas, a 24.54×104 km² expansion in moderate-intensity areas, and a 13.59×104 km² gain in high-intensity areas; SSP5-8.5 shows a low-level reduction of 63.66×104 km², but is accompanied by a significant expansion of 32.9×104 km² in the moderately suitable zone and 35.16×104 km² in the highly suitable zone. Among these scenarios, SSP5-8.5 exhibits the highest sensitivity to climate change due to its substantial increase in moderately and highly suitable habitats.

Figure 6. Distribution of suitable areas for C. heterophylla in China in ssp126 (A, B), ssp245 (C, D), ssp585 (E, F) scenarios in the 2050s (A, C, E) and 2090s (B, D, F).
3.4 Changes in spatial patterns of C. heterophylla under different climatic scenarios
Future habitat suitability for C. heterophylla shows substantial overlap (69.20%~77.56% retention) with 1970–2000 suitable areas across all scenarios, mainly concentrated in Heilongjiang, Jilin, Liaoning, Beijing, Tianjin, Hebei, Shanxi, Shandong. Over all, the rate of habitat expansion ranges from 18.83% to 37.78%, with newly suitable areas mainly emerging in northern and high-altitude marginal regions. Conversely, range contraction occurs predominantly along the southern margins of the historical distribution. The highest increase is observed under the 2090s SSP5-8.5 scenario, while the smallest increase is observed under the 2050s SSP1-2.6 scenario. Suitable areas are expected to expand mainly in the northern regions of Heilongjiang, Inner Mongolia, northern Qinghai, Gansu, and northern Ningxia. The rate of loss is 22.44% ~ 30.70%, with the highest loss under the 2090s SSP2-4.5 scenario, and the lowest loss under the 2050s SSP1-2.6 scenario. The suitable area for C. heterophylla is anticipated to contract mainly in southern Hubei, southern Hunan, southern Henan, eastern Sichuan, and southern Guizhou (Table 5; Figure 7).

Table 5. Changes in habitat area of C. heterophylla across future climate scenarios.
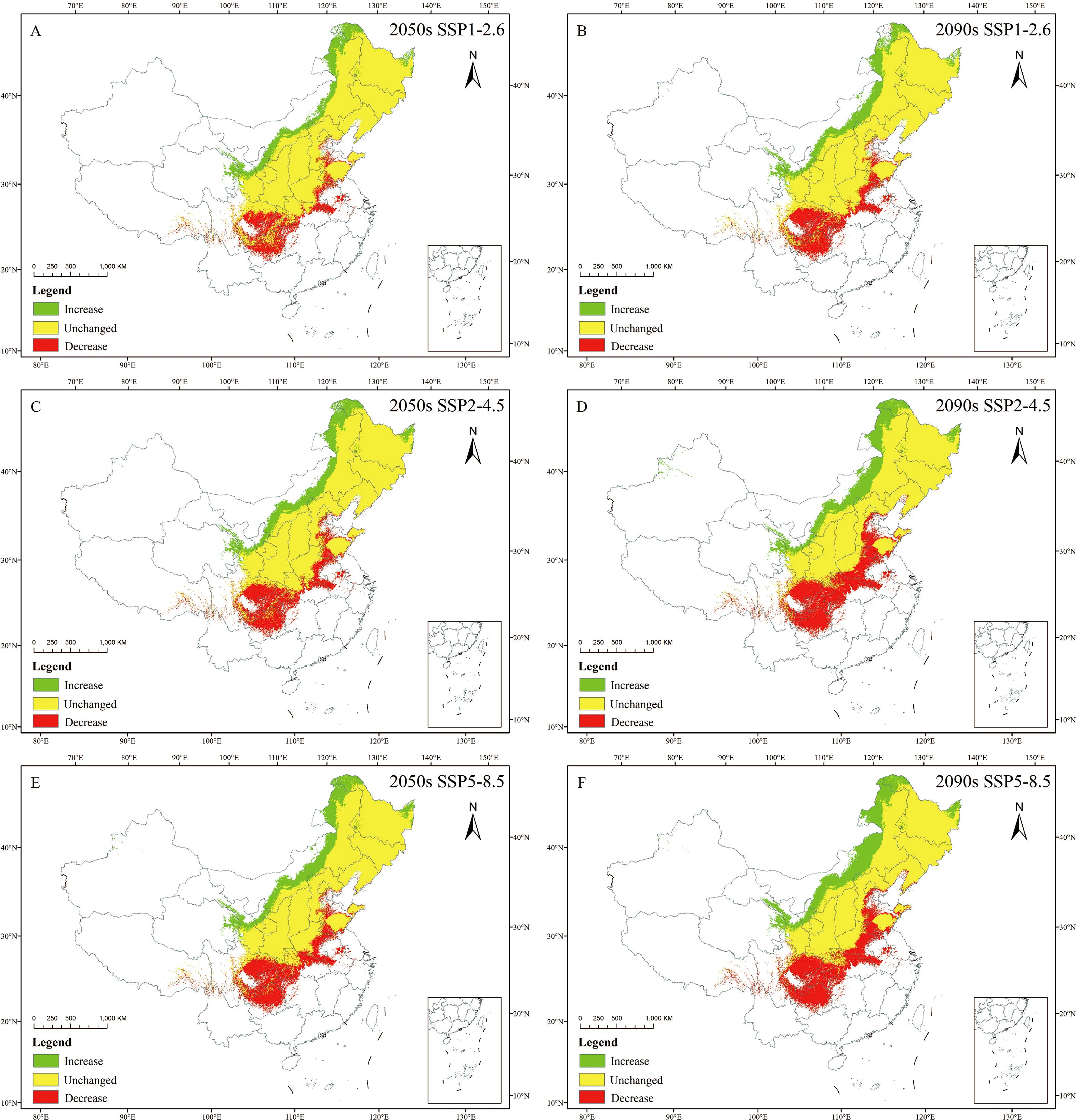
Figure 7. Spatial changes in C. heterophylla in China in ssp126 (A, B), ssp245 (C, D), ssp585 (E, F) scenarios in the 2050s (A, C, E) and 2090s (B,D, F).
Northern Heilongjiang, Inner Mongolia, northern Qinghai, Gansu, northern Ningxia, southern Hubei, southern Hunan, southern Henan, eastern Sichuan, and southern Guizhou represent core change zones for future habitat suitability shifts in China.
3.5 Centroid migration trends of C. heterophylla under different climatic scenarios
C. heterophylla distribution centroids exhibited consistent northward shifts across all scenarios. During the 1970–2000 period, the centroid was located near Dazhangjia Village, Longyao County, Xingtai City, Hebei Province (114.849°E, 39.409°N). Under the SSP1-2.6 scenario, the centroid is projected to shift to the vicinity of Zhoukoudian, Fangshan District, Beijing (115.94°E, 39.66°N) by the 2050s, and further toward Fengtai District, Beijing (116.402°E, 39.857°N) by the 2090s. Under the SSP2-4.5 scenario, it is expected to migrate to the area around Miyun District, Beijing (116.911°E, 40.598°N) by the 2050s, and eventually reach Huangtukang Township, Weichang Manchu and Mongolian Autonomous County, Chengde City, Hebei Province (117.697°E, 41.764°N) by the 2090s. Under the SSP5-8.5 scenario, the centroid will move to Datou Mountain Township, Weichang Manchu and Mongolian Autonomous County (117.269°E, 41.858°N) by the 2050s, and finally arrive at Tianqiao Town, Fengning Manchu Autonomous County, Chengde City (117.185°E, 41.014°N) by the 2090s.
4 Discussion
4.1 Key environmental determinants of habitat suitability of C. heterophylla
The results from the environmental variables replacement contribution analysis and jackknife test indicate that Bio16 (Precipitation of wettest quarter, 39.5%), Bio9 (Mean temperature of driest quarter, 22.2%), Alt (Altitude, 16.2%), and Bio3 (Isothermality, 7.1%) represent primary environmental factors governing the distribution of C. heterophylla (Table 3; Figure 3). However, discrepancies exist regarding the identification of primary environmental factors across different studies. Liu and Chen (2024) reported that the Bio6 (Minimum temperature of coldest month) and human footprint were the main drivers for hazelnut distribution. This divergence may arise from their analysis of multiple C. heterophylla species collectively rather than species-specific responses. The ecological adaptability of different Corylus species varies significantly, and pooled analyses may fail to capture their species-specific environmental requirements. Similarly, He et al. (2023) identified Bio4 (Temperature seasonality), Bio16 (Precipitation of wettest quarter), and Bio15 (Precipitation seasonality) as dominant factors, potentially because their study did not account for topographic influences. Topographic parameters, including altitude, slope, and aspect, significantly influence plant distribution patterns, particularly in regions with complex geomorphology, by mediating habitat suitability through their effects on localized climatic conditions and edaphic properties (Huang et al., 2024). Consequently, the integration of multidimensional environmental variables is critical for robust identification of the mechanistic drivers governing phytogeographic distributions.
Bio16 showed an optimal range of 300–450 mm, with the highest probability of occurrence peaking at 350 mm (Figure 4), reflects the species’ sensitivity to water supply during the critical summer fruit development stage. In many temperate regions, peak precipitation coincides with summer (Fick and Hijmans, 2017), providing essential soil moisture for nut filling and oil synthesis. Moderate rainfall ensures high yields and quality, while excessive rainfall leading to waterlogging can cause root asphyxiation and increase disease incidence (Marsal et al., 1997). Bio9 exhibited a suitable range of -12°C to 2°C, indicating strong adaptation to cold and dry winters. This physiological trait enables C. heterophylla to survive extreme winter temperatures as low as -30°C to -40°C (Erdogan and Mehlenbacher, 2000). Regarding Alt (Altitude), this study identified a constrained distribution range of 100–700 m, peaking at 300 m. Wang et al. (2020) reported a broader suitable altitude range of 0–3000 m for C. mandshurica. This divergence suggests a clear altitudinal niche partitioning between the two species, which may reduce direct competition and facilitate their coexistence in sympatric regions. Huo et al. (2016) demonstrated that the northern populations of C. heterophylla and C. mandshurica exhibit high climatic similarity, though C. mandshurica generally occurs at higher elevations than C. heterophylla in sympatric regions. This finding is in line with the results of this study. Bio3 has an optimal range of 24 to 30. As the ratio of Bio2 (Mean diurnal temperature range) to Bio7 (Temperature annual range), this variable ecologically reflects the relative magnitude of diurnal versus seasonal temperature variation. The identified optimum indicates that the species thrives in regions with pronounced annual temperature seasonality. This climatic preference aligns with the physiological traits of the C. heterophylla as a temperate deciduous shrub, particularly its dependence on cold winters (Fadón et al., 2020). Sufficient low-temperature accumulation is crucial for meeting the “chilling requirement” necessary to break dormancy, thereby ensuring synchronized spring bud break and normal inflorescence development (González De Andrés et al., 2023). Conversely, in regions with Bio3 values significantly exceeding 30 (e.g., maritime climates), elevated winter temperatures may fail to meet cold requirements, triggering dormancy disorders that impair normal growth and development (Luedeling et al., 2011). Thus, the distribution and adaptation of C. heterophylla depend not only on growing-season temperature conditions but critically on temperature patterns exhibiting pronounced seasonal rhythms to synchronize its phenological processes with the external environment.
4.2 Potential impacts of climate change on suitable habitats for C. heterophylla
The analysis of environmental variables and their impact on the potential distribution of C. heterophylla under 1970–2000 and future climate scenarios provides significant insights into the species’ ecological adaptability and future prospects. Using the optimized MaxEnt model, we predict the species’ geographic distribution under different time periods. Under 1970–2000 climatic conditions, C. heterophylla is primarily distributed in Northeast China (Changbai Mountains and Xiao Xing’anling ranges), North China (Yanshan Mountains), and Northwest China (northern slopes of the Qinling Mountains) (Figure 5). The total suitable area covers 210.85×104 km² (21.96% of China’s land area), with the highly suitable area accounting for 39.09×104 km² and being concentrated in Jilin, Liaoning, Hebei, Shanxi provinces (Table 4). Future climate scenario simulations consistently project a contraction in the total suitable habitat range of C. heterophylla. This trend aligns with earlier studies reporting accelerated range loss in its southern low-elevation populations (Yang et al., 2022). Model results further suggest that although low-suitability areas will gradually decrease, medium- and high-suitability zones are expected to expand in a spatially heterogeneous manner across climate scenarios, particularly in regions such as northern Heilongjiang, northern Inner Mongolia, and northwestern provinces including Gansu and Ningxia (Figure 7). This latitudinal and altitudinal expansion is driven by combined effects of climate warming and projected shifts in precipitation patterns. Altered thermal and moisture conditions collectively enable previously cold-limited northern and high-elevation areas to gradually meet the eco-physiological requirements of C. heterophylla (Álvarez-Álvarez et al., 2025). These newly expanded areas represent potential future habitats for the C. heterophylla, holding critical conservation value for maintaining its population persistence and mitigating biodiversity loss caused by the species’ southern distribution decline.
4.3 Spatial change patterns and centroid shift analysis
Under future climate change scenarios, the suitable distribution range of C. heterophylla exhibits a bipolar migration pattern characterized by “northward expansion and southward contraction” (Figure 7). Newly suitable habitats are predominantly distributed in northern Heilongjiang, northern Inner Mongolia, Gansu, Qinghai, and other northwestern regions, whereas substantial habitat loss is observed in southern China, including Hubei, southern Henan, and Sichuan. Compared to the SSP1-2.6 and SSP2-4.5, SSP5-8.5 scenario shows the most significant changes in suitable areas. This can be attributed to the substantial increases in both precipitation and temperature across northern China under this high-emission pathway (Zhou et al., 2020), which collectively create thermal hygric conditions that more closely match the species’ requirements during critical growth phases (Fick and Hijmans, 2017). In contrast, under the SSP2-4.5 and SSP1-2.6 scenarios, the more moderate warming and limited precipitation increases are insufficient to generate equivalent habitat gains in the north. Consequently, the net change in these scenarios is dominated by habitat losses in the southern parts of the range, resulting in a greater overall reduction in total suitable area. CMIP6-based projections of future global temperatures reveal persistent warming trends across all emission scenarios, albeit with significant variations in magnitude. Zhao et al. (2021) further estimate a global temperature increase ranging from 0.6 to 7.8 °C, which may exacerbate the frequency and intensity of extreme climate events. Additionally, CMIP6 models project increased precipitation across most regions of China, with the most substantial enhancement occurring under the high-emission scenario (SSP5-8.5) (Álvarez-Álvarez et al., 2025). The northward migration of suitable habitats can be attributed to synergistic improvements in hydrothermal conditions in northern regions, which optimize the growth environment for C. heterophylla. Conversely, in the southern parts of the current distribution range, future projections indicate that precipitation may exceed the species identified optimal upper threshold of 450 mm (Figure 4; Xu et al., 2022). Such excessive rainfall could result in periodic soil waterlogging, elevating the risk of root hypoxia—a condition known to be detrimental to C. heterophylla (Bailey-Serres and Voesenek, 2008).
The distribution centroid of C. heterophylla demonstrates a consistent northward migration trend under all future climate scenarios relative to its historical position (1970–2000) in Hebei Province (Figure 8). This migratory trend is most pronounced under the higher-emission scenarios (SSP2-4.5 and SSP5-8.5) and is strongly linked to projected changes in precipitation patterns. Climate models indicate a general increase in growing-season precipitation across northern China (Zhou et al., 2020). This enhanced water supply may transform previously water-limited areas in the north into new suitable habitats for C. heterophylla. Under high-emission scenarios, while traditional distribution areas like North China may experience intensified seasonal droughts and precipitation uncertainty, some more northern regions could benefit from modest precipitation increase or greater climatic stability (Zhang et al., 2021). This shift in precipitation patterns favors a contraction of suitable habitat toward these more favorable high-latitude zones. Conversely, in the southern parts of the current range, increased precipitation is projected to frequently exceed the species’ optimal upper threshold (450 mm; Figure 4), leading to waterlogging stress and habitat degradation. Supporting this, Wang et al. (2019) documented upward shifts in suitable ranges for most plant species in Changbai Mountain in response to rising temperatures and altered precipitation patterns, with significantly greater migration rates and distances under high-emission scenarios compared to low-emission ones. These findings collectively suggest that enhanced warming under high-emission scenarios likely drives more substantial displacements of C. heterophylla’s distribution center.

Figure 8. The geographical distribution change of the centroid of C. heterophylla suitable area under different climate scenarios [(B) is the expansion of part (A)].
5 Conclusions
This study employed an optimized MaxEnt model to predict the potential distribution shifts of C. heterophylla under current and future climate change scenarios. Bio16 (Precipitation of wettest quarter), Bio9 (Mean temperature of driest quarter, 15.5%), Alt (Altitude, 15.3%), and Bio3 (Isothermality, 7.1%) were identified as core environmental factors constraining C. heterophylla’s distribution, revealing the species’ strong dependence on seasonal hydrothermal patterns. Although the total suitable habitat area is projected to decrease under future climates, the extent of moderately to highly suitable habitats is expected to increase, accompanied by a pronounced northward shift in the distribution centroid. These findings provide practical insights for the conservation and utilization of C. heterophylla germplasm resources. It is recommended to proactively establish climate-resilient conservation bases in northern expansion zones to ensure sustainable use and enhance production potential under future climates. In southern contraction zones, a “defensive conservation” strategy should be adopted, prioritizing the establishment of germplasm nurseries and continuous population monitoring to safeguard genetic diversity.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author contributions
JW: Formal Analysis, Writing – original draft, Data curation, Investigation. WX: Validation, Writing – review & editing, Funding acquisition, Supervision. PS: Validation, Investigation, Writing – review & editing. DL: Funding acquisition, Validation, Investigation, Writing – review & editing. CC: Writing – review & editing, Funding acquisition, Investigation, Methodology.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Heilongjiang Provincial Natural Science Foundation of China (LH2023C090), and the Earmarked Fund (CARS-17), the Innovation and Development Project of China Meteorological Administration (CXFZ2025J048), the Precision Identification Project of Germplasm Resources (22250677), and the Hainan Province Science and Technology Project “Innovation and Breeding Techniques of Sugar Beet Germplasm Resources” (B24CQ008P).
Acknowledgments
We appreciate the support from the Tianchi Program and Sugar Beet Breeding Research Project.
Conflict of interest
Author WX was employed by the company China National Seed Group Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1668828/full#supplementary-material
References
Allouche, O., Tsoar, A., and Kadmon, R. (2006). Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 43, 1223–1232. doi: 10.1111/j.1365-2664.2006.01214.x
Álvarez-Álvarez, P., Aviñoa-Arias, A., Díaz-Varela, E., López-Bao, J. V., and Pérez-Girón, J. C. (2025). Impact of climate change over distribution and potential range of chestnut in the Iberian Peninsula. Front. For. Glob. Change 8. doi: 10.3389/ffgc.2025.1561027
Araújo, M. B., Anderson, R. P., Barbosa, A. M., Beale, C. M., Dormann, C. F., Early, R., et al. (2019). Standards for distribution models in biodiversity assessments. Sci. Adv. 5, eaat4858. doi: 10.1126/sciadv.aat4858
Bailey-Serres, J. and Voesenek, L. A. C. J. (2008). Flooding stress: acclimations and genetic diversity. Annu. Rev. Plant Biol. 59, 313–339. doi: 10.1146/annurev.arplant.59.032607.092752
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W., and Courchamp, F. (2012). Impacts of climate change on the future of biodiversity. Ecol. Lett. 15, 365–377. doi: 10.1111/j.1461-0248.2011.01736.x
Brown, J. L., Bennett, J. R., and French, C. M. (2017). SDMtoolbox 2.0: the next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 5, e4095. doi: 10.7717/peerj.4095
Chen, I. C., Hill, J. K., Ohlemüller, R., Roy, D. B., and Thomas, C. D. (2011). Rapid range shifts of species associated with high levels of climate warming. Science 333, 1024–1026. doi: 10.1126/science.1206432
Chen, S., Jiang, Z., Song, J., Xie, T., Xue, Y., and Yang, Q. (2025). Prediction of potential habitat of Verbena officinalis in China under climate change based on optimized MaxEnt model. Front. Plant Sci. 16. doi: 10.3389/fpls.2025.1563070
Cheng, Y., Jiang, S., Zhang, X., Li, H., Wang, K., and Sun, H. (2019). Whole-genome re-sequencing of Corylus heterophylla blank-nut mutants reveals sequence variations in genes associated with embryo abortion. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01465
Cobos, M. E., Peterson, A. T., Barve, N., and Osorio-Olvera, L. (2019). kuenm: an R package for detailed development of ecological niche models using MaxEnt. PeerJ 7, e6281. doi: 10.7717/peerj.6281
Elith, J. and Leathwick, J. R. (2009). Species distribution models: ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 40, 677–697. doi: 10.1146/annurev.ecolsys.110308.120159
Erdogan, V. and Mehlenbacher, S. A. (2000). Interspecific hybridization in hazelnut (Corylus). J. Amer. Soc Hortic. Sci. 125, 489–497. doi: 10.21273/JASHS.125.4.489
Fadón, E., Herrero, M., and Rodrigo, J. (2020). Chilling and heat requirements of temperate stone fruit trees (Prunus sp.). Agronomy 10, 409. doi: 10.3390/agronomy10030409
Fan, L., Ren, J., Yang, Y., and Zhang, L. (2020). Comparative analysis on essential nutrient compositions of 23 wild hazelnuts (Corylus heterophylla) grown in Northeast China. J. Food Qual. 2020, 9475961. doi: 10.1155/2020/9475961
Fianchini, M., Solidoro, C., and Melaku Canu, D. (2025). Improving MaxEnt reliability with multi-criteria analysis and site weighting: a case study on Caulerpa cylindracea. Ecol. Solut. Evid. 6, e70074. doi: 10.1002/2688-8319.70074
Fick, S. E. and Hijmans, R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Giraldo, D. C. and Kucuker, D. M. (2024). Ecological niche modeling of Lactarius deliciosus using kuenm R package: insights into habitat preferences. Fungal Biol. 128, 2022–2031. doi: 10.1016/j.funbio.2024.07.010
González De Andrés, E., Colangelo, M., Luelmo-Lautenschlaeger, R., López-Sáez, J. A., and Camarero, J. J. (2023). Sensitivity of Eurasian rear-edge birch populations to regional climate and local hydrological conditions. Forests 14, 1360. doi: 10.3390/f14071360
Gu, C., Tang, Y., Liu, L., Wang, B., Zhang, Y., Yu, H., et al. (2021). Predicting the potential global distribution of Ageratina adenophora under current and future climate change scenarios. Ecol. Evol. 11, 12092–12113. doi: 10.1002/ece3.7974
Guisan, A., Tingley, R., Baumgartner, J. B., Naujokaitis-Lewis, I., Sutcliffe, P. R., Tulloch, A. I., et al. (2013). Predicting species distributions for conservation decisions. Ecol. Lett. 16, 1424–1435. doi: 10.1111/ele.12189
He, J., Chao, L., Guo, J., Yun, P., Wang, Q., and Han, F. (2023). Cultivation techniques and harvesting utilization of hazelnuts. Inner Mong. For. Invest. Des. 46, 74–76. doi: 10.13387/j.cnki.nmld.2023.06.016
He, X., Ma, W., Zhao, T., Yang, X., Ma, Q., Liang, L., et al. (2023). Ecological differentiation and historical distribution changes of Corylus heterophylla species complex since the last interglacial period. J. Beijing For. Univ. 45, 11–23. doi: 10.12171/j.1000-1522.20210350
Hirzel, A. and Guisan, A. (2002). Which is the optimal sampling strategy for habitat suitability modelling. Ecol. Model. 157, 331–341. doi: 10.1016/s0304-3800(02)00203-x
Huang, E., Chen, Y., and Yu, S. (2024). Climate factors drive plant distributions at higher taxonomic scales and larger spatial scales. Front. Ecol. Evol. 11. doi: 10.3389/fevo.2023.1233936
Huo, H., Ma, Q., Li, J., Zhao, T., and Wang, G. (2016). Distribution patterns and suitable habitat climate evaluation of Corylus germplasm resources in China. J. Plant Genet. Resour. 17, 801–808. doi: 10.13430/j.cnki.jpgr.2016.05.002
Kuang, S., Zhou, Z., Liang, M., Gao, X., and Cao, Y. (2022). Interpretation of the IPCC sixth assessment report working group II report. Environ. Prot. 50, 71–75. doi: 10.14026/j.cnki.0253-9705.2022.09.001
Lee, S., Jung, H., and Choi, J. (2021). Projecting the impact of climate change on the spatial distribution of six subalpine tree species in South Korea using a multi-model ensemble approach. Forests 12, 37. doi: 10.3390/f12010037
Lenoir, J., Bertrand, R., Comte, L., Bourgeaud, L., Hattab, T., Murienne, J., et al. (2020). Species better track climate warming in the oceans than on land. Nat. Ecol. Evol. 4, 1044–1059. doi: 10.1038/s41559-020-1198-2
Liu, Y. and Chen, L. (2024). Predicting the impact of climate change on Corylus species distribution in China: integrating climatic, topographic, and anthropogenic factors. Ecol. Evol. 14, e70528. doi: 10.1002/ece3.70528
Luedeling, E., Girvetz, E. H., Semenov, M. A., and Brown, P. H. (2011). Climate change affects winter chill for temperate fruit and nut trees. PLoS One 6, e20155. doi: 10.1371/journal.pone.0020155
Marsal, J., Girona, J., and Mata, M. (1997). Leaf water relation parameters in almond compared to hazelnut trees during a deficit irrigation period. J. Amer. Soc Hortic. Sci. 122, 582–587. doi: 10.21273/jashs.122.4.582
Morales, N. S., Fernández, I. C., and Baca-González, V. (2017). MaxEnt’s parameter configuration and small samples: are we paying attention to recommendations? A systematic review. PeerJ 5, e3093. doi: 10.7717/peerj.3093
Phillips, S. J., Anderson, R. P., and Schapire, R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
Ren, J., Li, S., Zhang, Y., Yang, Q., Liu, J., Fan, J., et al. (2025). MaxEnt-based evaluation of climate change effects on the habitat suitability of Magnolia officinalis in China. Front. Plant Sci. 16. doi: 10.3389/fpls.2025.1601585
Rubenstein, M. A., Weiskopf, S. R., Bertrand, R., Carter, S. L., Comte, L., Eaton, M. J., et al. (2023). Climate change and the global redistribution of biodiversity: substantial variation in empirical support for expected range shifts. Environ. Evid. 12, 7. doi: 10.1186/s13750-023-00296-0
Shan, Y., Shen, H., Huang, L., Hamezah, H. S., Han, R., Ren, X., et al. (2025). Optimized MaxEnt analysis revealing the change of potential distribution area of Lygodium japonicum in China driven by global warming. Front. Plant Sci. 16. doi: 10.3389/fpls.2025.1601956
Sutherst, R. W. and Maywald, G. F. (1985). A computerised system for matching climates in ecology. Agric. Ecosyst. Environ. 13, 281–299. doi: 10.1016/0167-8809(85)90016-7
Taylor, A. T., Hafen, T., Holley, C. T., González, A., and Long, J. M. (2020). Spatial sampling bias and model complexity in stream-based species distribution models: a case study of Paddlefish (Polyodon spathula) in the Arkansas River basin, USA. Ecol. Evol. 10, 705–717. doi: 10.1002/ece3.5905
Trew, B. T. and Maclean, I. M. (2021). Vulnerability of global biodiversity hotspots to climate change. Glob. Ecol. Biogeogr. 30, 768–783. doi: 10.1111/geb.13272
Urban, M. C., Bocedi, G., Hendry, A. P., Mihoub, J. B., Pe’er, G., Singer, A., et al. (2016). Improving the forecast for biodiversity under climate change. Science 353, aad8466. doi: 10.1126/science.aad8466
Valavi, R., Guillera-Arroita, G., Lahoz-Monfort, J. J., and Elith, J. (2022). Predictive performance of presence-only species distribution models: a benchmark study with reproducible code. Ecol. Monogr. 92, e01486. doi: 10.1002/ecm.1486
Wang, L., Cui, L., and Zhang, Y. (2025). Preliminary report on artificial promotion restoration experiment of wild Corylus heterophylla. For. Sci. Technol. 50, 21–23 + 58. doi: 10.19750/j.cnki.1001-9499.2025.01.006
Wang, Q., Fan, B., and Zhao, G. (2020). Prediction of potential suitable habitats for Corylus mandshurica in China under climate change. Chin. J. Ecol. 39, 3774–3784. doi: 10.13292/j.1000-4890.202011.014
Wang, L., Wang, W., Wu, Z., Du, H., Zong, S., and Ma, S. (2019). Potential distribution shifts of plant species under climate change in Changbai Mountains, China. Forests 10, 498. doi: 10.3390/f10060498
Wang, Y., Zhao, Y., Miao, G., Zhou, X., Yu, C., and Cao, Y. (2024). Predicting the potential distribution of Dendrolimus punctatus and its host Pinus massoniana in China under climate change conditions. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1362020
Xu, H., Chen, H., and Wang, H. (2022). Future changes in precipitation extremes across China based on CMIP6 models. Int. J. Climatol. 42, 635–651. doi: 10.1002/joc.7264
Xu, D., Zhuo, Z., Wang, R., Ye, M., and Pu, B. (2019). Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling. Glob. Ecol. Conserv. 19, e00691. doi: 10.1016/j.gecco.2019.e00691
Yang, Z., Ma, W., He, X., Zhao, T., Yang, X., Wang, L., et al. (2022). Species divergence and phylogeography of Corylus heterophylla Fisch. complex (Betulaceae): inferred from molecular, climatic and morphological data. Mol. Phylogenet. Evol. 168, 107413. doi: 10.1016/j.ympev.2022.107413
Yang, Z., Zhao, T., Liang, L., Jiang, L., Wang, L., Hou, S., et al. (2023). Genetic analysis of the cultivars of Ping’ou hybrid hazelnut (C. heterophylla Fisch. × C. avellana L.) in China based on SSR markers. Forests 14, 1405. doi: 10.3390/f14071405
Yi, Y., Cheng, X., Yang, Z., and Zhang, S. (2016). MaxEnt modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour.) in Yunnan, China. Ecol. Eng. 92, 260–269. doi: 10.1016/j.ecoleng.2016.04.010
Zhang, W., Furtado, K., Wu, P., Zhou, T., Chadwick, R., Marzin, C., et al. (2021). Increasing precipitation variability on daily-to-multiyear time scales in a warmer world. Sci. Adv. 7, eabf8021. doi: 10.1126/sciadv.abf8021
Zhang, F., Liang, F., Wu, K., Xie, L., Zhao, G., and Wang, Y. (2024). The potential habitat of Angelica dahurica in China under climate change scenario predicted by MaxEnt model. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1388099
Zhao, Z., Luo, Y., and Huang, J. (2021). Global warming and abrupt climate change. Adv. Clim. Change Res. 17, 114–120. doi: 10.12006/j.issn.1673-1719.2020.180
Zhou, T., Zhang, W., Zhang, L., Zhang, X., Qian, Y., Peng, D., et al. (2020). The dynamic and thermodynamic processes dominating the reduction of global land monsoon precipitation driven by anthropogenic aerosols emission. Sci. China Earth Sci. 63, 919–933. doi: 10.1007/s11430-019-9553-5
Keywords: Corylus heterophylla, suitable area, climate change, MAXENT model, CMIP6
Citation: Wang J, Xing W, Sun P, Liu D and Cheng C (2025) Predicting the potential distribution of Corylus heterophylla in China under future climate change using an optimized MaxEnt model. Front. Plant Sci. 16:1668828. doi: 10.3389/fpls.2025.1668828
Received: 18 July 2025; Accepted: 24 October 2025;
Published: 17 November 2025.
Edited by:
Shuguang Wang, Southwest Forestry University, ChinaReviewed by:
Fen Yu, Jiangxi Agricultural University, ChinaJong Bin Jung, Mokpo National University, Republic of Korea
Copyright © 2025 Wang, Xing, Sun, Liu and Cheng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dali Liu, ZGFsaWxpdV9obGp1QDE2My5jb20=; Chunxiang Cheng, NTM3NDM0MTlAcXEuY29t