 Aurimas Sabeckis
Aurimas Sabeckis Roma Semaškienė
Roma Semaškienė Karolina Lavrukaitė
Karolina Lavrukaitė- Department of Plant Pathology and Protection, Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, Akademija, Kėdainiai, Lithuania
Root rots, seedling blights and snow mold diseases caused by Microdochium nivale and M. majus threaten winter wheat production in temperate regions. This study investigated the occurrence and dynamics of both pathogens in three-year field trials using quantitative PCR analysis of winter wheat cultivars sown at different times and treated with various seed treatment fungicides. Both species were consistently detected, but their prevalence varied by year: M. nivale was dominant under prolonged snow cover in 2023, whereas M. majus reached highest levels in 2022, when snow cover was less persistent. Later sowing generally reduced M. nivale infection in years with moderate disease pressure, although this effect was diminished under epidemic conditions. Seed treatment fungicides containing fludioxonil or succinate dehydrogenase inhibitor compounds (fluxapyroxad and sedaxane) consistently suppressed pathogen DNA levels, whereas triazole-dominant treatments showed more variable results. Cultivar-related differences were also evident, with ‘Ada’ and ‘KWS Emil’ frequently exhibiting higher infection levels than ‘Skagen’ or ‘Patras.’ These findings highlight that Microdochium spp. incidence is shaped by the interaction of environmental conditions, agronomic practices, and host genotype. By integrating these factors, this study advances our understanding of pathogen ecology and contributes to sustainable management strategies for winter wheat under changing climatic conditions.
1 Introduction
Winter wheat (Triticum aestivum L.) is an essential crop in temperate regions such as Northern and Eastern Europe. The vast growing scale increases the susceptibility of this vital cereal to a range of pathogenic fungi (Ristaino et al., 2021), particularly those within the Microdochium genus, which pose a significant threat to global food security (Gogoleva et al., 2024).
Microdochium species and their taxonomy have been a topic of interest for researchers for a long time. Initially, pathogens were suggested to be classified as varieties of species of the genus Fusarium (Fusarium nivale var. nivale and majus) (Wollenweber, 1930) with the later classification of F. nivale to Microdochium nivale by Samuels and Hallett (1983) to Microdochium genus. In 2005, Glynn et al. demonstrated distinct genetic heterogeneity between isolates of Microdochium majus and Microdochium nivale within a functional gene. This study provided the first sequence-based phylogenetic evidence supporting the recognition of M. majus and M. nivale as separate species (Glynn et al., 2005). Recognizing M. nivale and M. majus as distinct species is therefore important not only for taxonomy, but also for understanding their epidemiology and developing targeted management strategies in winter cereals.
Both pathogens are considered principal causative agents of pink snow mold and fusarium head blight (Tronsmo et al., 2001; Jørgensen et al., 2011; Nielsen et al., 2013), leading to considerable yield losses and grain quality reduction. They have a broad host range and can damage all winter and spring cereals affecting plants throughout basically all growth stages. M. nivale and M. majus infections can result in reduced germination, as well as pre- and post-emergence seedling death after winter, root and stem base diseases, and leaf blight (Ponomareva et al., 2021). Therefore, beyond their role in the most prominent snow mold damage, Microdochium spp. can contribute to significant crop losses during the early growth stages and are important causal agents of root rots and seedling blights (Jørgensen et al., 2011).
Microdochium spp. are soil- and seed-borne fungi that infect the plant base, particularly under conditions of prolonged soil wetness, low temperatures, and snow cover (Gorshkov et al., 2020). Infected plants often exhibit pale or necrotic leaf sheaths, reduced root systems, and poor survival during winter. Yield losses caused by Microdochium-related diseases can vary significantly depending on the weather conditions, cultivar susceptibility, and agronomic practices (Váňová et al., 2011).
While seed treatment fungicides are still considered as the most effective control measurement against Microdochium spp. it is important to use them with considered caution. Fungicide performance can vary between years and environments, therefore no single treatment consistently provides complete protection. In addition, extensive chemical control can lead to pathogen resistance for certain active ingredients, emphasizing the need for integrated strategies that combine seed treatment with appropriate sowing time and resistant cultivars (Ren et al., 2016). Active ingredients such as fludioxonil, triazoles (e.g., tebuconazole, prothioconazole, triticonazole), and SDHIs (succinate dehydrogenase inhibitors) (e.g., sedaxane, fluxapyroxad) (Walker et al., 2009) are widely applied and have shown efficacy in reducing seed and soil-borne inoculum, thereby lowering root rot and snow mold incidences (Nielsen et al., 2013; Ren et al., 2016). Although the development and use of resistant cultivars is considered the most sustainable and economical method for the control of some Microdochium caused diseases (Ren et al., 2016), the resistance of the most important cereal cultivars to these pathogens is currently low (Ponomareva et al., 2021). Therefore, snow mold and related diseases are an important problem in the breeding and management of winter cereals (Temirbekova et al., 2022).
Temperate climates are experiencing shifts, with longer, wetter autumns and milder winters, which create a favorable environment for Microdochium survival and proliferation (Juroszek and von Tiedemann, 2011). These conditions extend the period of environmental suitability and host susceptibility. These shifts could lead to increased disease pressure and altered species distributions, necessitating a deeper understanding of the factors influencing Microdochium dynamics in wheat protection (Gagkaeva et al., 2020). In addition, Microdochium pathogens gradually adapt to a warmer climate, spreading to territories with less snow and causing diverse types of plant diseases throughout the growing period (Tsers et al., 2023).
The intricate interplay between these climatic factors and agricultural practices significantly influences the incidence and severity of diseases such as pink snow mold, seedling blight, and Fusarium head blight caused by these pathogens (Gawronska and Gołębiowska-Pikania, 2016; Gagkaeva et al., 2020). Therefore, understanding the prevalence and factors influencing the incidence of Microdochium species is important for developing effective disease-management strategies. Although the morphological differences between M. nivale and M. majus can assist in differentiating these species in laboratory settings, molecular analysis is a more reliable tool for species identification. Notably, M. majus typically produces larger macroconidia that are primarily three-septate, comprising over 60% of its spores, whereas M. nivale tends toward fewer septa and smaller spores (Gagkaeva et al., 2020). Despite the broader average dimensions of macroconidia in M. majus, the conidial size ranges overlap, making morphological evaluation a useful but not fully reliable diagnostic trait (Jewell and Hsiang, 2013; Nielsen et al., 2013).
This study aimed to elucidate the impact of seed treatment fungicides, sowing time, and cultivar selection on the incidence of Microdochium nivale and M. majus in winter wheat using real-time quantitative polymerase chain reaction (qPCR) for the precise identification and quantification of fungal DNA.
2 Materials and methods
2.1 Experimental design
A three-year field study was conducted from 2021 to 2023 to investigate the occurrence of Microdochium spp. under various agronomic conditions. The experiments were conducted with three main factors: wheat cultivar, seed treatment, and sowing time. Four winter wheat cultivars (“Ada,” “KWS Emil,” “Skagen,” and “Patras”) were used for this experiment, each treated with four different commercial seed treatment fungicide formulations and an untreated control (Table 1). To evaluate the effect of sowing time, experiments were sown on two different dates: the second to third decade of September and the first decade of October, in accordance with the optimal and late sowing times of local agronomic practices. Field trials were conducted in Central Lithuania on loamy, drained soil classified as Cambisols with Endocalcaric and Endogleyic properties (Kochiieru et al., 2024). During the trial period winter, wheat was sown after winter oilseed rape using conventional tillage practices. Maintenance with fertilizers and herbicides was performed according to agronomical recommendations. Sowing was conducted using a Wintersteiger Kubota plot drill (Wintersteiger, Austria) in a randomized complete block design with four replicates. The seeding rate was adjusted annually to achieve a target of 450 viable seeds per square meter based on each seed lot’s germination percentage and thousand-kernel weight. Each experimental plot measured 15 m².

Table 1. Active ingredients and doses of seed treatment fungicides used in the field trial.
2.2 Meteorological conditions
Meteorological data from the local Dotnuva meteorological station (Figure 1) revealed notable variations in autumn and winter conditions across the years of the trial. The 2021–2022 season experienced a cooler, drier autumn followed by a relatively mild winter, with a high amount of precipitation in November, mostly snowfall. In contrast, the 2022–2023 season was marked by near-average temperatures and relatively high snowfall from December to June, whereas the 2023–2024 season was characterized by fluctuating temperatures and inconsistent precipitation.

Figure 1. Average monthly air temperatures and total monthly precipitation in trial site, 2021–2023, compared with the SCN – Standard Climate Normal 1990–2020 (Lithuanian Hydrometeorological Service).
Snow cover also varied considerably; in 2022–2023, snow persisted for up to 77 consecutive days, with temperatures fluctuating around 0°C in January, creating stable, humid conditions at the soil surface. In comparison, snow cover lasted only 46 days in 2021–2022 and declined further to 36 days in 2023–2024 (Sabeckis et al., 2025).
These patterns are highly relevant to the development of M. nivale and M. majus, both of which are known to thrive under prolonged snow cover owing to their tolerance of low temperatures, as both species remain active under low temperature conditions. Moreover, they exhibit pathogenicity during extended cool periods, particularly when snow insulates the soil and weakens plant defenses.
2.3 Sampling and DNA extraction
In early spring of each year, prior to stem elongation, 30 (three plants from 10 different spots) randomly selected plants were collected from every plot. Collected winter wheat samples were cut into 1 cm long stem base segments and rinsed thoroughly under running tap water to remove soil and debris. The air-dried samples were homogenized in liquid nitrogen. A 100 mg subsample of the homogenized material was used for DNA extraction using The E.Z.N.A.® SP Plant DNA Kit (Omega Biotek, Inc., United States) according to the manufacturer’s instructions.
2.4 Standard curves and quantitative real-time polymerase chain reaction analysis
The quantification of DNA in the sample using qPCR was based on the standard curve method. To prepare Microdochium standard curves, genomic DNA was extracted from pure cultures at known concentrations. Pure cultures of M. nivale and M. majus obtained from the Leibniz Institute DSMZ (Germany) were used as standards. The extracted DNA from these cultures was used to prepare standard curves. DNA concentration and purity were assessed using a biophotometer (Eppendorf, Germany). Plant standard curves were generated using DNA isolated from healthy, young leaves of the target cereal species at known concentrations, measured using a Biophotometer (Eppendorf, Germany). To construct standard curves, six 10-fold serial dilutions of DNA (1:10–1:106 ) were used in the qPCR reactions (Suproniene et al., 2010; Nielsen et al., 2013; Jonavičienė, 2016).
Extracted DNA from the test samples was diluted at ratios of 1:10, 1:20, 1:50, and 1:100 to evaluate the qPCR reaction quality against the standard curves. Based on amplification performance, the dilution that produced the most consistent and reliable results was selected, and all sample DNA was subsequently diluted 1:10 with double-distilled water (ddH2 O) for further analysis.
Quantitative real-time PCR (qPCR) was conducted following the protocol described by Nielsen et al. (2013), with modifications from Jonavičienė (2016). Each reaction was performed in a final volume of 15 µL, comprising 7.5 µL of Power SYBR® Green PCR Master Mix (Applied Biosystems, USA), 300 nM of each primer, 0.5 µg µL−1 of bovine serum albumin (BSA; Thermo Fisher Scientific, Lithuania), and 2.5 µL of template DNA. The species-specific primers used in this analysis are listed in Table 2.

Table 2. Species-specific primers and their sequences used for qPCR analysis.
All qPCR reactions were conducted in duplicate using a 7900HT Fast Real-Time PCR System (Applied Biosystems, USA), applying the thermal cycling protocol described by Nielsen et al. (2013).
2.5 Statistical analysis
The qPCR values are expressed as nanograms (ng) of fungal DNA per microgram (μg) of plant DNA. Results are presented as means, and the standard deviation of the values was calculated. A three-way ANOVA (Type III sums of squares) was performed to evaluate the main and interaction effects of the sowing date, cultivar, and seed treatment fungicide. Statistical analyses were conducted using JASP (version 0.19.3; JASP Team, Amsterdam, The Netherlands). Statistical significance was set at p <0.05. The results are presented as mean ± standard deviation (SD). Figures and tables were generated using Microsoft Excel (Microsoft Corp., Redmond, WA, USA).
3 Results
Root rot was observed in winter wheat during the entire research period. Despite consistent disease pressure, most of the applied seed treatment fungicides were effective against root rot pathogens (Sabeckis et al., 2025). Based on these findings, a molecular study was conducted. Field samples were analyzed using quantitative PCR (qPCR) analysis. Data confirmed the presence of M. nivale and M. majus in all the analyzed winter wheat cultivars. The quantity of fungal DNA varied depending on the cultivar, sowing time, and year of sampling. The highest concentrations of M. nivale and M. majus DNA were recorded in 2023, whereas the lowest were detected in 2024. These fluctuations likely reflect the influence of meteorological conditions that are favorable for Microdochium spp. development.
3.1 M. nivale DNA quantitative assay
Quantitative PCR analysis confirmed the presence of M. nivale in all tested winter wheat cultivars, although there was substantial variation in pathogen DNA quantity in different years (Table 3). The highest values of M. nivale DNA in all tested cultivars were detected in 2023, coinciding with the presence of snow mold in the trial and/or prolonged snow cover (Sabeckis et al., 2025), whereas lower levels occurred in 2024, reflecting less favorable conditions for pathogen development. For example, in the cultivar “Ada” (optimal sowing, untreated), M. nivale DNA reached 723.77 ng μg−1 in 2023 but declined to 7.99 ng μg−1 in 2024. Similarly, “Skagen” (optimal sowing, untreated) exhibited 241.35 in 2023 and maintained comparatively high levels in 2024 (70.04 ng μg−1), despite the overall reduction in disease pressure.

Table 3. Mean ( ± SD) quantity of Microdochium nivale DNA (ng pathogen DNA per μg plant DNA) in winter wheat cultivars sown at optimal and late dates under different seed treatment fungicide applications, 2022–2024.
The data suggested that sowing time influenced infection levels, with later sowing generally resulting in reduced M. nivale DNA compared to optimal sowing. In most cases, sowing time was most evident in low to moderate pressure years (2022 and 2024), whereas in spring 2023, under more favorable environmental conditions for the pathogen, higher DNA quantities were detected regardless of sowing time, thereby reducing the impact of this factor. Seed treatment with fungicides typically reduced pathogen DNA levels compared with those in untreated controls. Notably, Maxim 025 FS (fludioxonil) and Bariton Super (protioconazole + tebuconazole + fludioxonil) often provided substantial reductions in DNA of M. nivale in winter wheat samples under high disease pressure. Kinto Plus (fluxapyroxad + triticonazole + fludioxonil), as well as Vibrance Star (sedaxane + fludioxonil + triticonazole) also demonstrated pathogen suppression, though with variable results throughout the research years.
However, in the context of effective reduction of pathogen DNA, the efficacy of individual seed treatment fungicides varied, and no single treatment consistently outperformed the others. The results also indicated differences between cultivars in certain years, with more sensitive cultivars “Ada” and “Skagen” often exhibiting higher infection levels than “KWS Emil” and “Patras,” although cultivar-related distinctions diminished during epidemic conditions when disease pressure was universally high.
In general, M. nivale DNA was present in different amounts in all wheat cultivars, with the highest DNA amounts in a year with snow mold and/or prolonged snow cover. In fact, later sowing times led to reduced M. nivale DNA compared to optimal sowing times. Fungicide seed treatments reduced the DNA amount of the pathogen, but the efficacy was inconsistent between years and cultivars.
3.2 M. majus DNA quantitative assay
The data from the quantitative PCR analysis for M. majus are presented in Table 4. Analysis showed the presence of the pathogen in winter wheat across all three years, with even more pronounced variation between seasons and treatments that are observed for M. nivale. The highest DNA quantities were generally observed in 2022, with exceptionally high values in some cultivar and seed treatment combinations, particularly in untreated cv. “Ada” (optimal sowing, 1,027.21 ng μg−1) and cv. “KWS Emil” (574.30 ng μg−1). In contrast, the pathogen DNA levels were much lower in 2023. Similar to the M. nivale data, the lowest quantities occurred in 2024, when DNA quantities in most treatments and cultivars were near zero, indicating unfavorable conditions for M. majus development.
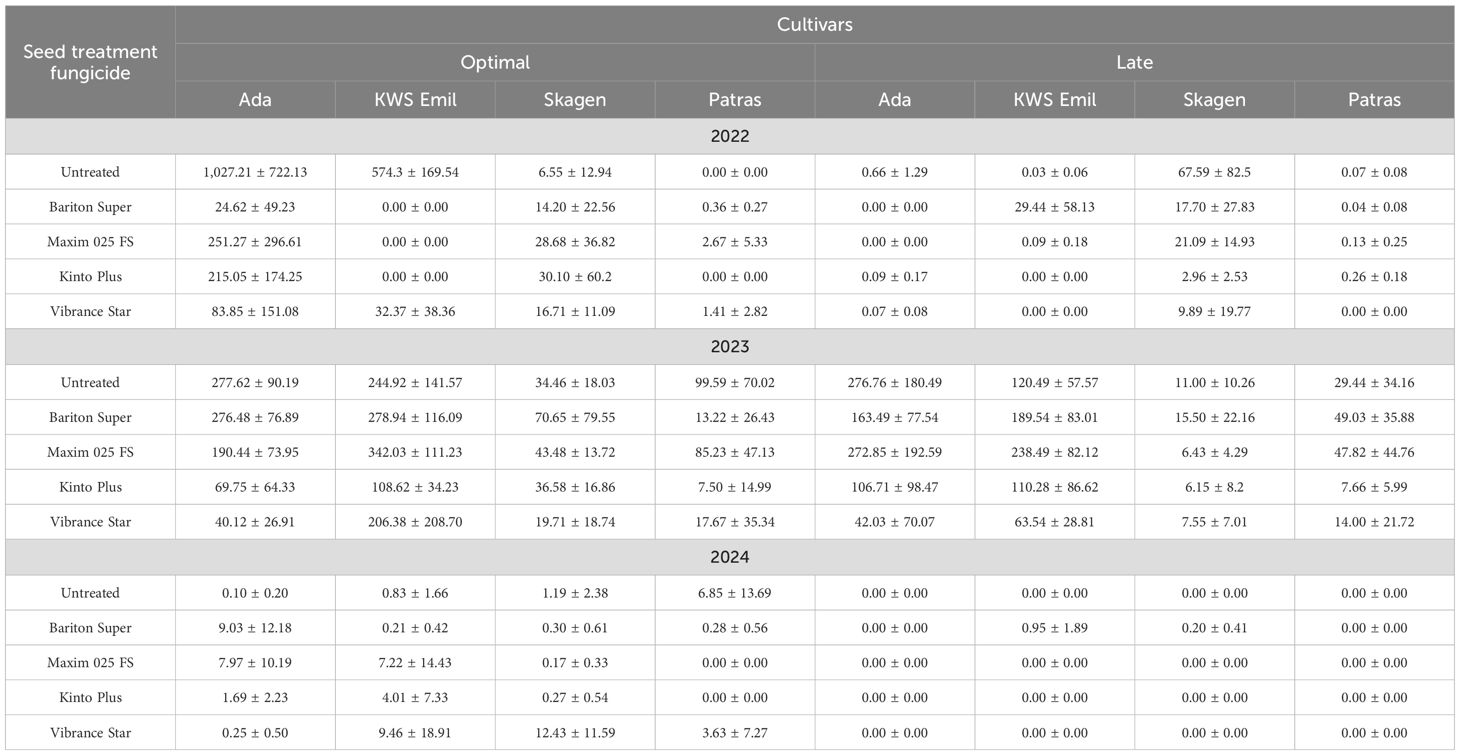
Table 4. Mean ( ± SD) quantity of Microdochium majus DNA (ng pathogen DNA per μg plant DNA) in winter wheat cultivars sown at optimal and late dates under different seed treatment fungicide applications, 2022–2024.
The time of sowing also affected the number of DNA and, therefore, the infection patterns in certain years. In 2022, later-sown winter wheat contained greatly reduced DNA quantities in most cultivars; for example, the values in cv. “Ada” samples dropped from 1027.21 (optimal) to 0.66 (late), and cv. “KWS Emil” from 574.30 to 0.03 ng μg−1. However, in 2023, the effect of sowing time was less consistent, and in some cases, late sowing still resulted in high levels of pathogen DNA, as seen in cv. “Ada,” where late-sown wheat samples contained values (276.76 ng μg−1) comparable to those of optimally sown wheat (277.62 ng μg−1).
M. majus DNA levels were generally lower in treated seed winter wheat compared to untreated controls, particularly in years when quantities were exceptionally high. In 2022, at the optimal sowing time, winter wheat cv. “Ada” treated with prothioconazole + tebuconazole + fludioxonil (Bariton Super) contained 24.62 ng μg−1 compared to 1027.21 ng μg−1 in the untreated control, while cv. “KWS Emil” samples did not contain pathogen DNA compared to 574.30 ng μg−1 in the untreated control with the same fungicide. Treatments with Maxim 025 FS (fludioxonil), Kinto Plus (fluxapyroxad + triticonazole + fludioxonil), and Vibrance Star (sedaxane + fludioxonil + triticonazole) also showed strong reductions in some cases, although their relative efficacy varied in 2023. For example, seed treatment with Maxim 025 FS in cv. “KWS Emil” winter wheat sowed optimal contained unexpectedly high values of pathogen DNA—342.03 ng μg−1. In 2024, the fungicide impact was less revealing because of the very low overall infection. For cultivar comparison, differences were more pronounced in 2022, with “Ada” and “KWS Emil” showing much higher DNA quantities than “Skagen” and “Patras,” especially under optimal sowing without treatment. In 2023, the results were less distinctive, with DNA quantities resulting in moderate levels, although winter wheat samples of cultivars “Ada” and “KWS Emil” tended to rank higher than the others. By 2024, cultivar differences were minimal because of uniformly low infection levels.
Overall, the results indicate that M. majus pressure was greatest in 2022, less intense in 2023, and negligible in 2024, with clear indications of cultivar and sowing time effects apparent only under high to moderate disease pressure with high general DNA quantity. Fungicide treatments were generally effective in reducing values, although their relative performance varied between years and cultivars.
3.3 Interaction of different factors on M. nivale and M. majus DNA Quantity
An interaction analysis was performed to evaluate whether the effects of sowing time, cultivar, and seed treatment fungicides on Microdochium DNA levels were influenced by each other, providing insight into how these factors jointly affect pathogen development. Data for M. nivale presented in Table 5 showed clear two- and three-factor interactions between sowing time, cultivar, and seed treatment fungicides, which were the most evident in 2023, reflecting complex factor relationships under high disease pressure. Whereas M. majus interactions (Table 6) interaction effects were strongest in 2022, indicating that factor combinations had the greatest influence during years of exceptionally high pathogen DNA levels.
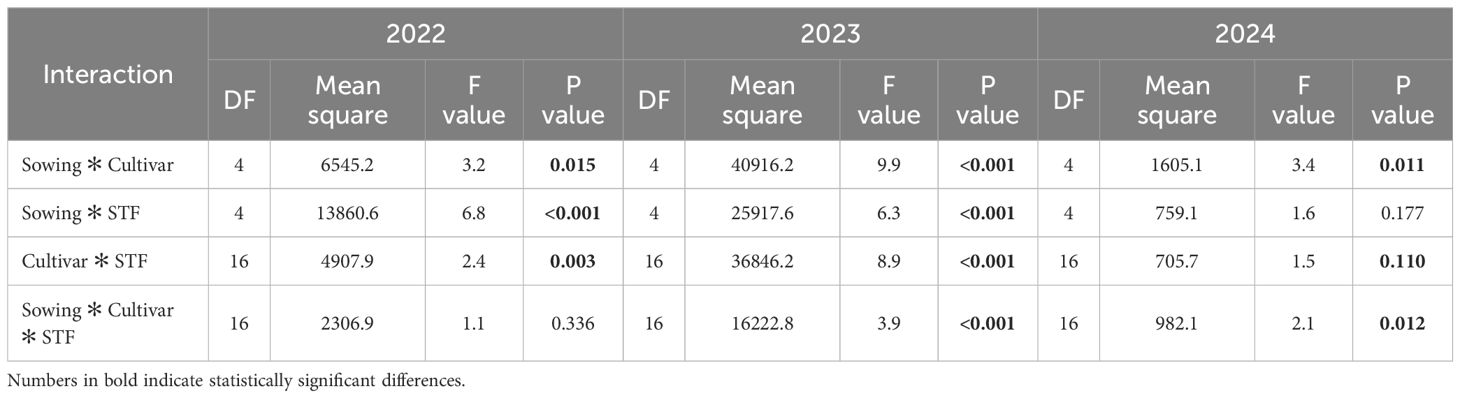
Table 5. Analysis of variance for interactions between sowing time, cultivar, and seed treatment fungicides (STF) on M. nivale DNA quantity in winter wheat, 2022–2024.
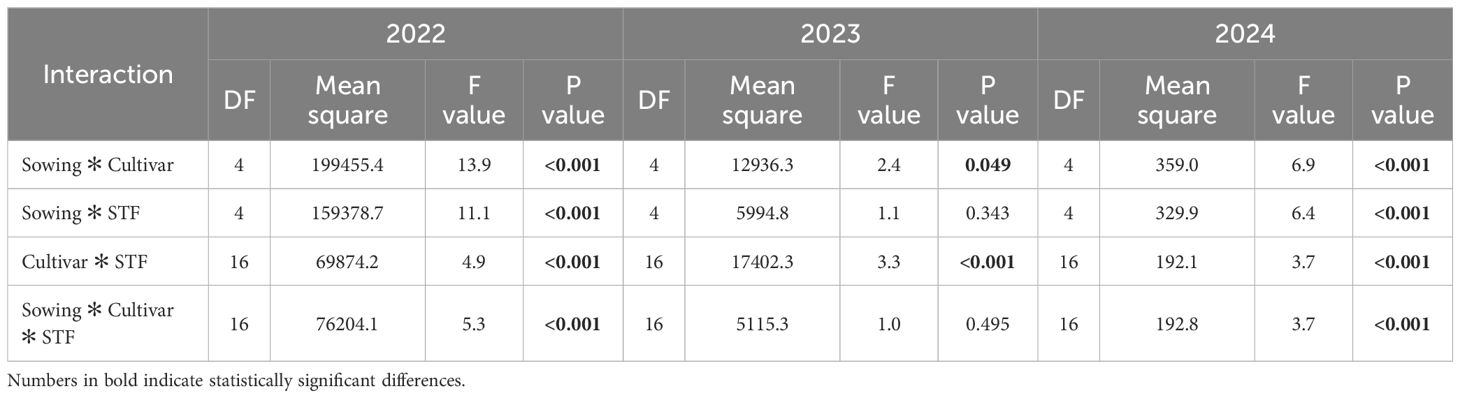
Table 6. Analysis of variance for interactions between sowing time, cultivar, and seed treatment fungicides (STF) on M. majus DNA quantity in winter wheat, 2022–2024.
Data for M. nivale showed that interaction patterns varied across years. The sowing time and cultivar interaction was significant in all three years, with the strongest in 2023 (F = 9.9, sig. <0.001), indicating that cultivar responses to sowing time were most evident under high disease pressure. The sowing time × seed treatment fungicide interaction was significant in 2022 and 2023 (sig. <0.001), suggesting that fungicide performance depended on the sowing date in these years, but not in 2024. Cultivar × fungicide interactions were significant in 2022 (p = 0.003) and especially in 2023 (F = 8.9, sig <0.001), showing strong cultivar-specific differences in fungicide responses under epidemic conditions. The three-way interaction was significant in 2023 (p <0.001) and 2024 (p = 0.012), indicating complex, environment-dependent factor combinations.
Conversely, for M. majus, the sowing time × cultivar interaction was significant only in 2022 (F = 15.6, sig. <0.001), indicating that cultivar adaptation to sowing time was most critical in years with extremely high pathogen presence. Furthermore, the interactions tended to be stronger in 2022 than in subsequent years. The sowing time × cultivar interaction was highly significant in 2022 (F = 13.9, p <0.001) and 2024 (F = 6.9, p <0.001), but was weaker in 2023 (p = 0.049). The sowing time × fungicide interaction was significant in 2022 and 2024 (p <0.001 for both), but not in 2023, suggesting a reduced role of sowing date in moderating fungicide effects during that year. Cultivar × fungicide interactions were significant in all years, with the strongest effect in 2022 (F = 4.9, p <0.001), indicating consistent cultivar-specific differences in the fungicide response. The three-way interaction was significant in 2022 and 2024 (p <0.001 for both) but absent in 2023, suggesting that under certain environmental conditions, the combined influence of sowing time, cultivar, and fungicide was reduced.
Overall, M. majus showed more pronounced and consistent interaction effects in high-pressure years like 2022, whereas M. nivale exhibited complex multi-factor interactions under epidemic conditions in 2023.
4 Discussion
The results of this three-year field study provide valuable insights into the complex dynamics of M. nivale and M. majus in winter wheat, highlighting the influence of environmental conditions, particularly prolonged snow cover, sowing time, seed treatment fungicides, and cultivar susceptibility to pathogen populations.
Our study revealed notable fluctuations in Microdochium DNA levels, with the highest concentrations of M. nivale and M. majus DNA recorded in 2023 and 2022, respectively, and the lowest levels in 2024. Specifically, the high DNA quantities of M. nivale in 2023 coincided with prolonged snow cover (up to 77 consecutive days) and temperatures fluctuating around 0°C in June (Sabeckis et al., 2025). This strongly aligns with the existing literature that emphasizes Microdochium nivale and M. majus as psychrotolerant fungi that thrive under prolonged snow cover (Gorshkov et al., 2020), which provides insulation, darkness, and humidity, thereby creating highly favorable conditions for snow mold development (Gorshkov et al., 2020; Tsers et al., 2023).
Experimental and field studies have demonstrated that M. nivale and M. majus differ in their host associations and competitive behaviors. In controlled inoculation experiments, M. majus isolates exhibited a weak but consistent selective advantage in wheat and oat seedlings, whereas M. nivale showed a strong competitive advantage in rye (Simpson et al., 2000). These observations are supported by large-scale field surveys of historical samples in Denmark, where M. majus was more frequently detected in wheat, barley, oats, and triticale, whereas M. nivale predominated in rye, suggesting that M. majus has been the dominant pathogen in cereals for the last 50 years (Nielsen et al., 2013). Conversely, a comprehensive study in Northeast Europe, which genetically characterized 46 Microdochium isolates from cereal (grains and plant tissues), found that 59% corresponded to M. nivale, while only 28% matched M. majus (Gagkaeva et al., 2020). This difference between regional studies may be linked to climatic differences, particularly snow cover: the continental interior of northeastern Europe, the presence of consistent and prolonged snow cover could favor the proliferation of M. nivale, whereas in the North Sea–Baltic maritime climate, snow cover is less constant and sporadic, likely creating conditions more favorable for M. majus growth. Our results align with this reasoning, as high M. nivale DNA levels were detected in 2023 under prolonged snow cover, whereas M. majus was more prominent in 2022, a season characterized by different meteorological conditions. Notably, DNA quantities in 2021 were relatively high despite moderate visual root rot pressure in field experiments designed to evaluate disease damage (Sabeckis et al., 2025), suggesting that pathogen DNA levels in stem base diseases do not always correspond directly with symptom expression, a pattern also observed for Oculimacula spp. in winter rye (Ramanauskienė et al., 2014). A study on Microdochium isolates showed that M. nivale and M. majus were the most pathogenic to wheat compared to other species (Ünal, 2024). Other researchers have found that Microdochium nivale might be more aggressive as a causal agent of root rot and also has a significantly stronger reducing effect on plant stem and root length compared to M. majus (Ren et al., 2015). Furthermore, these two species exhibit different capacities to cause damage depending on the temperature (Imathiu et al., 2010).
Our findings indicate distinct differences in cultivar susceptibility, with “Ada” consistently exhibiting higher levels of M. nivale DNA than other cultivars, suggesting its increased susceptibility. Similarly, for M. majus in 2022, “Ada” and “KWS Emil” showed significantly higher DNA quantities than “Skagen” and “Patras,” particularly regarding optimal sowing without treatment. Specific cultivar resistance mechanisms can interact with variability in pathogenicity among Microdochium populations, as demonstrated in other studies (Jewell and Hsiang, 2013; Abdelhalim et al., 2020; Gagkaeva et al., 2020). The impact of environmental conditions on winter survival among wheat lines can sometimes be more significant than inoculation itself, making it difficult to draw definitive conclusions on specific snow mold resistance challenges without considering environmental interactions (Ergon et al., 2003). Moreover, the importance of environmental context in winter is further highlighted by Yarosh et al. (2024), who addressed that snow cover and ecological plasticity strongly influenced cultivar resistance.
Seed treatment fungicides generally reduced Microdochium DNA quantity, supporting their significant role in disease control, although their effectiveness varied considerably. For instance, Bariton Super notably reduced M. majus in “Ada” at optimal sowing time from 1,027.21 to 24.62 ng μg−1 in 2022, while in Maxim 025 FS and Kinto Plus treated seed samples, the quantity averaged 251.27 and 215.05 ng μg−1, respectively. Fungicide treatments are recognized for improving plant vigor and enhancing winter survival in wheat and have been shown to be effective against seed- and soil-borne diseases such as smuts, kernel bunt, and root rots, with various active ingredients demonstrating efficacy in multiple studies (Bugingo et al., 2025). In some instances, however, M. majus DNA quantities were unexpectedly high (e.g., Maxim 025 FS in “KWS Emil” optimal sowing time in 2023), suggesting that fungicide performance can vary under different environmental conditions and disease pressure. Fludioxonil-based and SDHI (fluxapyroxad or sedaxane in Kinto Plus and Vibrance Star, respectively) containing seed treatments provided the most consistent suppression, whereas triazole-dominant mixtures showed effective yet more variable performance. This is in line with the results obtained by other researchers (Müllenborn et al., 2008; Brown et al., 2021).
The influence of sowing time on Microdochium infection levels was most evident for M. nivale, as in 2022, later sowing times significantly reduced DNA quantities in most tested cultivars. In addition, DNA quantities were lower in later-sown winter wheat in most years for M. nivale. Results regarding root rot suggested that later sowing reduced disease levels in years with low to moderate pressure, although this effect was less pronounced under high disease pressure, such as in 2023 (Sabeckis et al., 2025). Comparable results were obtained in Latvian field trials, suggesting that later sowing consistently lowers snow mold incidence in winter wheat, despite earlier sown plants accumulating more carbohydrates linked with cold tolerance (Ruza et al., 2011). Similarly, a study in Finland reported that postponing sowing reduced pink snow mold, leaf rust, and winter damage in winter rye, reinforcing the protective role of delayed sowing in northern climates (Serenius et al., 2005).
5 Conclusions
This study provides specific insights into the epidemiology of Microdochium nivale and M. majus in winter wheat using qPCR-based quantification of samples from 3-year field trials. Both pathogens were detected across all cultivars, but their prevalence varied with environmental conditions: M. nivale dominated under prolonged snow cover, whereas M. majus prevailed when snow cover was less persistent. Sowing time was more influential for M. nivale infection in years with moderate disease pressure, with later sowing generally reducing the pathogen DNA quantity. Although this effect was slightly less visible under conditions of higher pathogen pressure. Seed treatment fungicides, particularly those containing fludioxonil and SDHI compounds (fluxapyroxad or sedaxane), consistently suppressed pathogen DNA levels, although no treatment provided complete or uniform control across the years. Cultivar-related differences were also evident, with ‘Ada’ and ‘KWS Emil’ often exhibiting higher infection levels, suggesting that the genetic background interacts with pathogen species and environmental conditions to shape pathogen prevalence. By evaluating the influences and interactions of multiple environmental, agronomic, and genetic factors, this study contributes to advancing the sustainable management of Microdochium-related diseases and safeguarding winter wheat yields under changing climatic conditions.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
AS: Formal Analysis, Investigation, Methodology, Writing – original draft. RS: Conceptualization, Supervision, Writing – review & editing. AJ: Conceptualization, Methodology, Writing – original draft, Writing – review & editing. KL: Writing – original draft. EV: Formal Analysis, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Part of this research was supported by the long-term research program “Harmful organisms in agro and forest ecosystems” implemented by the Lithuanian Research Centre for Agriculture and Forestry.
Acknowledgments
The authors would like to thank the technical team of the Department of Plant Pathology and Protection of the Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry for their contribution to this research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abdelhalim, M., Brurberg, M. B., Hofgaard, I. S., Rognli, O. A., and Tronsmo, A. M. (2020). Pathogenicity, host specificity and genetic diversity in Norwegian isolates of Microdochium nivale and Microdochium majus. Eur. J. Plant Pathol. 156, 885–895. doi: 10.1007/s10658-020-01939-5
Brown, M., Jayaweera, D. P., Hunt, A., Woodhall, J. W., and Ray, R. V. (2021). Yield losses and control by sedaxane and fludioxonil of soilborne Rhizoctonia, Microdochium, and Fusarium species in winter wheat. Plant Dis. 105, 2521–2530. doi: 10.1094/PDIS-11-20-2401-RE
Bugingo, C., Ali, S., Yabwalo, D., and Byamukama, E. (2025). Optimizing fungicide seed treatments for early foliar disease management in wheat under northern great plains conditions. Agronomy 15, 291. doi: 10.3390/agronomy15020291
Ergon, Å., Skinnes, H., and Tronsmo, A. M. (2003). Testing snow mould resistance of winter wheat: inoculation experiments with microdochium nivale in the field. Acta Agric. Scand. B Soil Plant Sci. 53, 110–117. doi: 10.1080/09064710310009055
Gagkaeva, T., Orina, A. S., Gavrilova, O. P., and Gogina, N. N. (2020). Evidence of microdochium fungi associated with cereal grains in Russia. Microorganisms 8, 340. doi: 10.3390/microorganisms8030340
Gawronska, K. and Gołębiowska-Pikania, G. (2016). The effects of cold-hardening and Microdochium nivale infection on oxidative stress and antioxidative protection of the two contrasting genotypes of winter triticale. Eur. Food Res. Technol. 242, 1267–1276. doi: 10.1007/s00217-015-2630-8
Glynn, N. C., Hare, M. C., Parry, D. W., and Edwards, S. G. (2005). Phylogenetic analysis of EF-1 alpha gene sequences from isolates of Microdochium nivale leads to elevation of varieties majus and nivale to species status. Mycol Res. 109, 872–880. doi: 10.1017/S0953756205003370
Gogoleva, O. A., Ryazanov, E. A., Murzagulova, G. S., Ponomarev, S. N., Chastukhina, I. B., Sakhabutdinov, I. T., et al. (2024). Intra- and interpopulation diversity of the phytopathogenic fungi of the microdochium nivale species. J. Fungi 10, 841. doi: 10.3390/jof10120841
Gorshkov, V., Osipova, E., Ponomareva, M., Ponomarev, S., Gogoleva, N., Petrova, O., et al. (2020). Rye snow mold-associated microdochium nivale strains inhabiting a common area: variability in genetics, morphotype, extracellular enzymatic activities, and virulence. J. Fungi 6, 335. doi: 10.3390/jof6040335
Imathiu, S. M., Hare, M. C., Ray, R. V., Back, M., and Edwards, S. G. (2010). Evaluation of pathogenicity and aggressiveness of F. langsethiae on oat and wheat seedlings relative to known seedling blight pathogens. Eur. J. Plant Pathol. 126, 203–216. doi: 10.1007/s10658-009-9533-0
Jewell, L. E. and Hsiang, T. (2013). Multigene differences between Microdochium nivale and Microdochium majus. Botany 91, 99–106. doi: 10.1139/cjb-2012-0178
Jonavičienė, A. (2016). Beicų įtaka žieminių kviečių pašaknio puviniams ir pavasariniam pelėsiui. Žemės ūkio mokslai 23, 149–158. doi: 10.6001/zemesukiomokslai.v23i4.3402
Jørgensen, L. N., Nielsen, L. K., and Nielsen, B. J. (2011). Control of seedling blight in winter wheat by seed treatments – impact on emergence, crop stand, yield and deoxynivalenol. Acta Agric. Scand. B Soil Plant Sci. 62, 1–10. doi: 10.1080/09064710.2011.641028
Juroszek, P. and von Tiedemann, A. (2011). Potential strategies and future requirements for plant disease management under a changing climate. Plant Pathol. 60, 100–112. doi: 10.1111/j.1365-3059.2010.02410.x
Kochiieru, M., Veršulienė, A., Feiza, V., Feizienė, D., Shatkovska, K., and Deveikytė, I. (2024). The action of environmental factors on carbon dioxide efflux per growing season and non-growing season. Sustainability 16, 4391. doi: 10.3390/su16114391
Müllenborn, C., Steiner, U., Ludwig, M., and Oerke, E.-C. (2008). Effect of fungicides on the complex of Fusarium species and saprophytic fungi colonizing wheat kernels. Eur. J. Plant Pathol. 120, 157–166. doi: 10.1007/s10658-007-9204-y
Nicolaisen, M., Supronienė, S., Nielsen, L. K., Lazzaro, I., Spliid, N. H., and Justesen, A. F. (2009). Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods 76, 234–240. doi: 10.1016/j.mimet.2008.10.016
Nielsen, L. K., Justesen, A. F., Jensen, J. D., and Jørgensen, L. N. (2013). Microdochium nivale and Microdochium majus in seed samples of Danish small grain cereals. Crop Prot. 43, 192–200. doi: 10.1016/j.cropro.2012.09.002
Ponomareva, M. L., Gorshkov, V., Yu, Ponomarev, S. N., Korzun, V., and Miedaner, T. (2021). Snow mold of winter cereals: a complex disease and a challenge for resistance breeding. Theor. Appl. Genet. 134, 419–433. doi: 10.1007/s00122-020-03725-7
Ramanauskienė, J., Gaurilčikienė, I., Supronienė, S., Ronis, A., and Česnulevičienė, R. (2014). Evaluation of eyespot incidence and structure of Oculimacula spp. population in winter rye in Lithuania. Zemdirbyste 101, 425–430. doi: 10.13080/z-a.2014.101.054
Ren, R., Foulkes, J., Mayes, S., Yang, X., and Ray, R. V. (2016). Identification of novel quantitative trait loci for resistance to Fusarium seedling blight caused by Microdochium majus and M. nivale in wheat. Field Crops Res. 191, 1–12. doi: 10.1016/j.fcr.2016.03.011
Ren, R., Yang, X., and Ray, R. V. (2015). Comparative aggressiveness of Microdochium nivale and M. majus and evaluation of screening methods for Fusarium seedling blight resistance in wheat cultivars. Eur. J. Plant Pathol. 141, 281–294. doi: 10.1007/s10658-014-0541-3
Ristaino, J. B., Anderson, P. K., Bebber, D. P., Brauman, K. A., Cunniffe, N. J., Fedoroff, N. V., et al. (2021). The persistent threat of emerging plant disease pandemics to global food security. Proc. Natl. Acad. Sci. 118, e2022239118. doi: 10.1073/pnas.2022239118
Ruza, A., Bankina, B., and Strikauska, S. (2011). The impact of sowing time on sugar content and snow mould development in winter wheat. Acta Biologica Universitatis Daugavpiliensis 11, 88–95.
Sabeckis, A., Semaškienė, R., Jonavičienė, A., Venslovas, E., Lavrukaitė, K., and Almogdad, M. (2025). Effect of seed treatment and sowing time on microdochium spp. Caused root rot in winter wheat cultivars. Agronomy 15, 330. doi: 10.3390/agronomy15020330
Samuels, G. J. and Hallett, I. C. (1983). Microdochium stoveri and Monographella stoveri, new combinations for Fusarium stoveri and Micronectriella stoveri. Trans. Br. Mycological Soc. 81, 473–483. doi: 10.1016/S0007-1536(83)80115-6
Serenius, M., Huusela-Veistola, E., Hanna, A., Pahkala, K., and Laine, A. (2005). Effects of sowing time on pink snow mould, leaf rust and winter damage in winter rye varieties in Finland. Agric. Food Sci. 14, 362–376. doi: 10.2137/145960605775897696
Simpson, D. R., Rezanoor, H. N., Parry, D. W., and Nicholson, P. (2000). Evidence for differential host preference in Microdochium nivale var. majus and Microdochium nivale var. nivale. Plant Pathol. 49, 261–268. doi: 10.1046/j.1365-3059.2000.00453.x
Suproniene, S., Justesen, A. F., Nicolaisen, M., and Mankevičienė, A. (2010).Distribution of trichothecene and zearalenone producing Fusarium species in grain of different cereal species and cultivars grown under organic farming conditions in Lithuania. Available online at: https://www.researchgate.net/publication/45503936 (Accessed August 24, 2025).
Temirbekova, S. K., Kulikov, I. M., Ashirbekov, M. Z., Afanasyeva, Y. V., Beloshapkina, O. O., Tyryshkin, L. G., et al. (2022). Evaluation of Wheat Resistance to Snow Mold Caused by Microdochium nivale (Fr) Samuels and I.C. Hallett under Abiotic Stress Influence in the Central Non-Black Earth Region of Russia. Plants 11, 699. doi: 10.3390/plants11050699
Tronsmo, A. M., Hsiang, T., Okuyama, H., Nakajima, T., Iriki, N., Gaudet, D. A., et al. (2001). Low temperature plant microbe interactions under snow. Hokkaido Natl. Agric. Experiment Station Japan 7, 75–85.
Tsers, I., Marenina, E., Meshcherov, A., Petrova, O., Gogoleva, O., Tkachenko, A., et al. (2023). First genome-scale insights into the virulence of the snow mold causal fungus Microdochium nivale. IMA Fungus 14, 2. doi: 10.1186/s43008-022-00107-0
Ünal, F. (2024). Phylogenetic analysis of Microdochium spp. associated with turfgrass and their pathogenicity in cereals. PeerJ 12, e16837. doi: 10.7717/peerj.16837
Váňová, M., Matušinsky, P., Javůrek, M., and Vach, M. (2011). Effect of soil tillage practices on severity of selected diseases in winter wheat. Plant Soil Environ. 57, 245–250. doi: 10.17221/334/2010-PSE
Walker, A., Auclair, C., Gredt, M., and Leroux, P. (2009). First occurrence of resistance to strobilurin fungicides in Microdochium nivale and Microdochium majus from French naturally infected wheat grains. Pest Manag Sci. 65, 906–915. doi: 10.1002/ps.1772
Wollenweber, H. W. (1930). Fusaria Autographice Delineate. 2nd edn. Ed. Wollenweber, H. W. (Berlin).
Keywords: Microdochium nivale, seed treatment fungicides, sowing time, epidemiology, cultivar susceptibility
Citation: Sabeckis A, Semaškienė R, Jonavičienė A, Lavrukaitė K and Venslovas E (2025) Evaluation of sowing time and seed treatment fungicides on Microdochium spp. DNA quantity in winter wheat cultivars. Front. Plant Sci. 16:1694784. doi: 10.3389/fpls.2025.1694784
Received: 28 August 2025; Accepted: 06 November 2025; Revised: 28 October 2025;
Published: 02 December 2025.
Edited by:
Prem Lal Kashyap, ICAR-Indian Institute of Wheat and Barley Research, IndiaReviewed by:
Sanjay Kumar Goswami, Indian Institute of Sugarcane Research (ICAR), IndiaFathi Abdellatif Belhouadjeb, Centre de Recherche en Agropastoralisme, Algeria
Copyright © 2025 Sabeckis, Semaškienė, Jonavičienė, Lavrukaitė and Venslovas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aurimas Sabeckis, YXVyaW1hcy5zYWJlY2tpc0BsYW1tYy5sdA==