 Xiaomeng Shi1†
Xiaomeng Shi1† Changping Tian
Changping Tian Aidi Zhang
Aidi Zhang Jianzhao Li
Jianzhao Li- 1School of Horticulture, Ludong University, Yantai, China
- 2Yantai Academy of Agricultural Sciences, Yantai, China
- 3School of Food Engineering, Ludong University, Yantai, China
- 4Yantai Technology Center of Characteristic Plant Gene Editing and Germplasm Innovation, Ludong University, Yantai, China
The NAC gene family is an important transcription factor unique to plants, widely involved in plant growth, development, and response to different stress. The physicochemical properties, phylogenetics, chromosome localization, gene structure and motif, collinearity, and gene expression under stress of the PavNAC transcription factor family in sweet cherry were analyzed. The results showed that the whole genome sequence of sweet cherry contains 132 PavNAC transcription factor genes, which are unevenly distributed on 8 chromosomes and divided into 10 subfamilies. The protein length of PavNAC ranged from 152 to 755 amino acids, and the CDS length ranged from 459 to 2268. The gene structure showed that most PavNAC members contain 3 exons and 10 motifs. Collinearity analysis result showed that 54 PavNAC genes in sweet cherry have homologous genes in Arabidopsis. Transcriptome data of different development stages of ‘Hong Deng’ and ‘Black Pearl’ showed that PavNAC42 and PavNAC101 were specifically expressed in both varieties. Finally, expression of 15 PavNAC genes under different abiotic stress were analyzed by real-time PCR. The results showed that PavNAC70, PavNAC9, and PavNAC10 were induced by low temperature. PavNAC70 and PavNAC81 were both induced by salt and PEG stresses. The study laid the foundation for subsequent molecular breeding and further exploration of the function of PavNAC genes in sweet cherry.
Introduction
Transcription factors are proteins that control the transcription rate of genetic information from DNA to messenger RNA (Latchman, 1997). They play a role by combining with cis acting promoter elements, and participate in the response of plants to abiotic stress such as drought, high salt, and low temperature (Liu et al., 2014; Zhang et al., 2016). According to its DNA domain, transcription factors in plants can be divided into several gene families, such as WRKY, bZIP, MYB, DREB, AP2/ERF, C2H2, NAC, etc (Nogueira et al., 2005; Singh et al., 2021). These genes typically function collectively as members of large families, and individual members within a family can participate in different stress responses.
The NAC transcription factor gene family is one of the largest transcription factor families unique to plants (Gao et al., 2020; Lin et al., 2023). The NAC protein structure consists of a highly conserved DNA binding domain located at the N-terminus and a highly variable transcriptional regulatory region located at the C-terminus (Nie et al., 2020). Among them, the N-terminal is further divided into five sub domains: A, B, C, D, and E, which contain nearly 160 amino acid residues (Zhang et al., 2016). Among them, the sub domains A, C, and D are highly conserved, while the B and E domains vary among different plants (Ooka et al., 2003; Olsen et al., 2005). At the same time, the C-terminal exhibits high diversity in amino acid composition and function. With the continuous progress and improvement of bioinformatics technology, more and more plant NAC families have been identified and studied. For example, researchers have found 117 NAC proteins in Arabidopsis, 151 NAC proteins in rice (Nuruzzaman et al., 2010), 102 NAC proteins in cocoa beans (Shen et al., 2020), and 104 NAC proteins in medium grain tomato (Su et al., 2014).
The NAC gene plays multiple functions in plants (Puranik et al., 2012), including regulating the development of flowers and leaves (Vroemen et al., 2003; Aida et al., 1997), thickening of secondary cell walls (Zhong et al., 2006), protein and lipid metabolism pathways (Chi et al., 2017), leaf senescence and fruit development (Ma et al., 2018), lateral root formation (Xie et al., 2000), seed germination (Kim et al., 2008), and playing a role in plant senescence (Balazadeh et al., 2011). Researches have also found that NAC transcription factors play an important role in the response of plants to different stress. The NAC transcription factor region is rich in stress response elements, such as low temperature response elements, water deficiency response elements, damage response elements (Nakashima et al., 2012). The OsNAC2 gene in rice can interact with ABA, enhancing its ability to resist drought and high salt stress (Jiang et al., 2019). PtrNAC72 in goji berries can respond to drought stress by regulating proline (Wu et al., 2016). StNAC053 in potatoes can regulate related genes to increase the activity of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), thereby enhancing their drought tolerance (Wan F. X. et al., 2021). Under low temperature stress, banana MaNAC1 can interact with ICE-CBF, thereby improving the cold resistance (Shan et al., 2014). The SsNAC23 gene in sugarcane can also participate in response to low temperature stress (Nogueira et al., 2005). Under high salt stress, TsNAC1 in salt mustard binds to a proton transporter protein to regulate plant salt tolerance (Liu et al., 2017).
Sweet cherry (Prunus avium L.) is a deciduous tree of the Rosaceae family, native to Europe and western Asia. It is widely cultivated in the mountainous areas of eastern China and also in temperate regions around the world. Sweet cherry is also one of the famous cultivated cherry varieties, with large fruit shape, beautiful flavor, rich nutrition, and containing various nutrients and bioactive components, such as glucose, vitamin C, and anthocyanins (Gao and Mazza, 1995). In agricultural production, the normal development of sweet cherry is a key factor in enhancing yield and influencing cultivation expansion. However, sweet cherry is highly vulnerable to low-temperature frost damage during the flowering and early fruit stages, leading to reduced fruit yield and diminished quality. With a relatively shallow root system, sweet cherry is also susceptible to significant fruit drop when water supply is insufficient during development, such as under drought conditions, resulting in economic losses. Additionally, sweet cherry is extremely sensitive to salt stress, and soil salinization further restricts its cultivation. Therefore, studying their growth and development mechanisms is particularly important.
However, little is known about the NAC gene family in sweet cherry, particularly its roles in stress resistance. In order to better understand the molecular regulation mechanism of sweet cherry during development, it is necessary to study the NAC gene family in sweet cherry. The NAC family members were identified within the entire genome of sweet cherry, and their physicochemical properties, phylogenetic relationships, gene structure, conserved motifs, and chromosomal localization were analyzed. Furthermore, we analyzed the expression levels of selected PavNAC genes at different developmental stages in two sweet cherry cultivars, ‘Black Pearl’ and ‘Hong Deng’, using transcriptome data, as well as the expression of PavNAC genes under abiotic stress conditions, including low temperature, drought, and high salt. Finally, expression of 15 PavNAC genes under different abiotic stress were analyzed by q-PCR.
Materials and methods
Plant materials
The experimental materials used in this experiment were 3-month-old sweet cherry tissue cultured seedlings from the sweet cherry germplasm resource bank of Yantai academy of agricultural sciences. Transcriptome analysis was performed on two sweet cherry cultivars, ‘Hong Deng’ and ‘Black Pearl’. 200 mM NaCl was added to the MS medium for NaCl treatment. 20% PEG6000 (w/v) was added to MS medium for PEG treatment. MS medium was used as the control group. Sweet cherry seedlings were transplanted and propagated in vitro and cultured under normal conditions for 1, 2, 3, 4, and 5 days. Select 15 healthy seedlings with normal growth status and place them into a low-temperature (LT, 4°C) incubator for 1, 2, 3, 4, and 5 days as low-temperature treatment. The control group seedlings were grown in a room temperature (RT, 24 ± 1°C) growth chamber. The experiment was performed with three independent biological replicates for each of the five treatment groups. Finally, these seedlings were frozen in liquid nitrogen and stored at - 80°C for RNA extraction and gene expression analysis in subsequent experiments.
Identification of PavNAC genes family
The protein, CDS and genome sequence of sweet cherry were obtained from NCBI database (http://www.ncbi.nlm.nih.gov). The PF02365 of the NAC domain was download from Pfam database (http://pfam.xfam.org). Then, HMMER was used to find the possible NAC transcription factor family from sweet cherry genome. In order to verify whether the existing family members are accurate, the online software SMART tool (http://smart.embl-heidelberg.de) was used to confirm the specific number of NAC gene family members in sweet cherry by analyzing its domain.
Phylogenetic analysis of the PavNAC genes
In order to further study and analyze the evolutionary relationship between NAC families. The “One Step Build a ML Tree” function in TBtools software was used to build an evolutionary tree containing 132 PavNAC and 105 Arabidopsis NAC full-length protein sequences. According to the known classification information of Arabidopsis NAC gene family and phylogenetic tree analysis, sweet cherry NAC family was classified into different subgroups.
Chromosomal distribution of PavNAC genes
The starting and ending positions of candidate genes are obtained through the general feature format (GFF) annotation file extracted from the sweet cherry genome, and they are renamed according to their positions on the chromosome. The “Gene Location from GTF/GFF” function in TBtools software was used to make the location map of genes on the chromosome, where Chr width ratio was set to 0.03, and the color and font were modified and adjusted automatically. The settings of other default parameters remain unchanged.
Gene structures and motif analyses of PavNAC genes
In order to further understand the gene structure of PavNACs, the intron and exon composition of all gene sequences were analyzed by comparing with genome sequences. The “Visualize Gene Structure” function in TBtools software was used to analyze gene structure, drag the sweet cherry genome annotation file (GFF) and the gene ID of the PavNAC family to draw a domain analysis diagram of the PavNAC gene family. The online tool MEME (http://meme-suite.org/tools/meme) was used to find the conserved motif. The motif discovery function is used to predict and analyze the conserved motifs of 132 NACs. The number of software parameter motifs is set to 10, and default values are used for other parameters. In addition, TBtools visualization can also be used to combine evolutionary tree files with gene structure data to form composite graphs.
Collinearity analysis of PavNAC genes
To further explore the common origin of the genome, we used TBtools software to analyze the linear relationship between sweet cherry and Arabidopsis species. The “one-step MCScanX” function of TBtools was used, the genome sequence files and gene structure annotation files of sweet cherry and Arabidopsis were dragged to the corresponding boxes to start, and the process was completed when TBtools displayed “completion”. Then the “DuaI Systeny Plot” function to pull the Ctl, GFF, and collinearity files obtained in the previous step into the corresponding boxes based on the file type, and enter the gene ID of the PavNAC gene family to start the plot.
RNA extraction and quantitative real-time PCR
RNAprep pure plant plus kit (Tiangen, Beijing, China) was used to isolate total RNA for qPCR analysis. The integrity of RNA was detected by agarose gel electrophoresis, and the concentration of RNA was detected by spectrophotometer. Hiscript III RT supermax (Vazyme, Nanjing, China) was used to eliminate genomic DNA contamination and synthesize the first strand cDNA. RT-qPCR primers were designed using Primer3.0 software, with PavActin used as the reference gene, and their specificity was assessed using the NCBI BLAST program (Supplementary Table S1). ChamQ Universal SYBR qPCR Master Mix Kit (Vazyme, Nanjing, China) and bio-rad CFX connected real-time system were used to complete Q-PCR.
Statistical analysis
In statistical analysis, LSD analysis after one-way ANOVA was used. The graphics were generated with GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA).
Results
Identification of PavNAC genes family
BLAST alignment was performed on the identified sweet cherry NAC genes, then duplicate sequences were removed to ultimately identify 132 PavNAC genes, which were named PavNAC001-PavNAC132 based on their positions on chromosomes (Supplementary Table S2). According to the analysis of various physicochemical properties of the PavNAC gene family, the number of amino acids in the PavNAC family members ranges from 152 (PavNAC122) to 755 (PavNAC86), with an average CDS length of around 1107 bp. The longest CDS length is 2268 bp, and the shortest is only 459 bp.
Phylogenetic analysis of the PavNAC genes
To further investigate the evolutionary relationship of PavNAC, 132 sweet cherry PavNAC transcription factor protein sequences and 105 AtNAC transcription factor protein sequences from Arabidopsis were used to construct a phylogenetic tree. Based on the known family member classification information in Arabidopsis NAC transcription factors, the sweet cherry NAC gene family was classified into ten subgroups (Figure 1). The X subgroup had the most branches and members. The VII and VI subgroups contain the most members of the PavNAC gene family, with 32 and 28, respectively. The I, II, and V subgroups contain the fewest members of the PavNAC gene family, with only 2-3. All subgroups contain both sweet cherry and Arabidopsis NAC proteins, indicating a certain degree of evolutionary similarity between the NAC families of the two species. The difference in the number of branches and branching genes among different subgroups indicated that there is still a difference in each subfamily even when the homologous domain is the same. These differences may lay the structural foundation for the functional diversity of the PavNAC gene family.
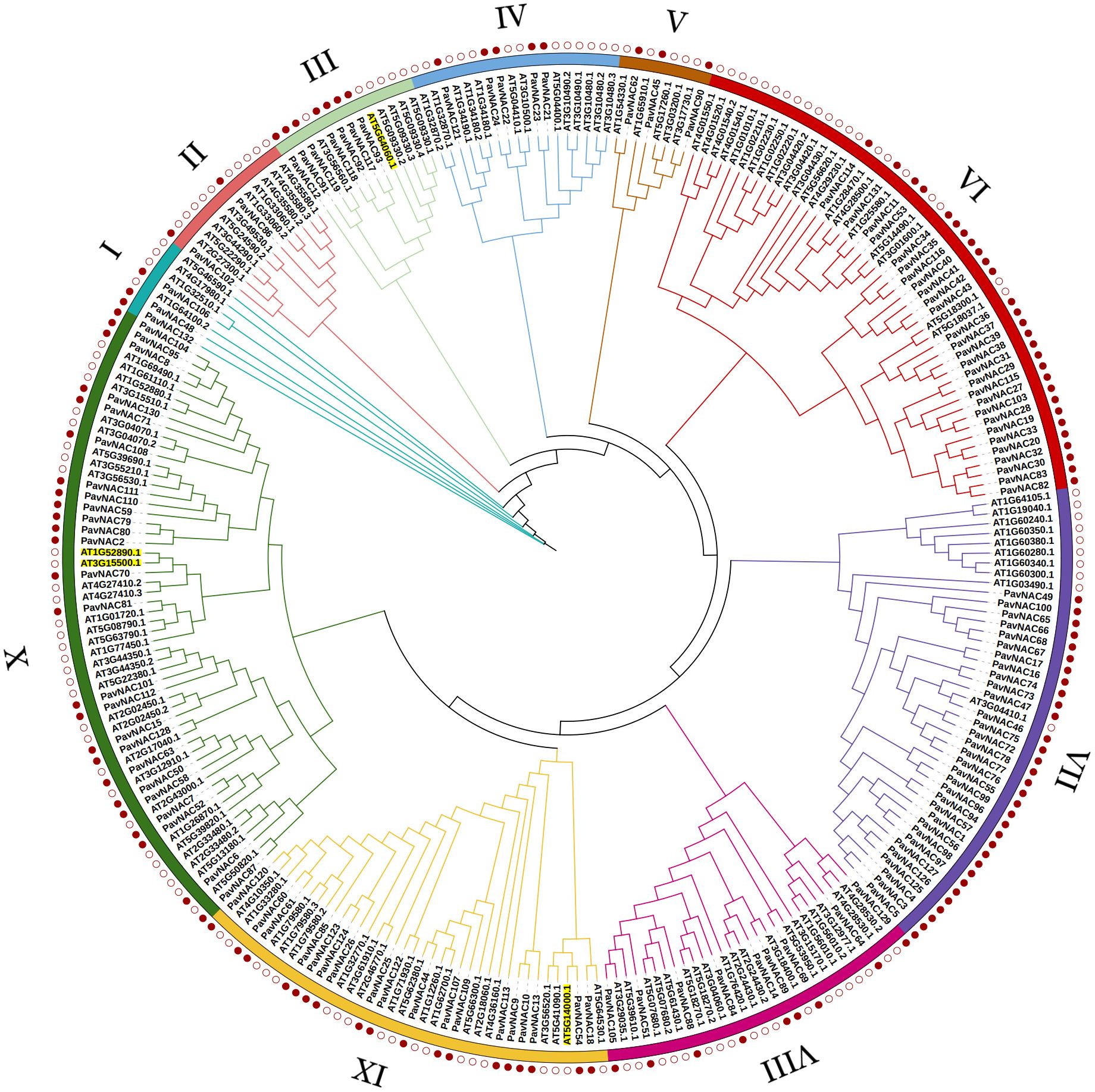
Figure 1. Phylogenetic analysis of NAC proteins in sweet cherry and Arabidopsis. 132 PavNAC transcription factor proteins from sweet cherry and 105 AtNAC proteins from Arabidopsis were selected to construct a rooted phylogenetic tree. This NAC evolutionary tree is divided into 10 different subgroups.
Chromosome distribution of PavNAC genes
132 PavNAC gene family members were distributed on all 8 sweet cherry chromosomes, but the number of distributions on each chromosome were different (Figure 2). Among them, the most members of the PavNAC gene family were distributed on chromosome 2, with a total of 28, while the least members of the PavNAC gene family were distributed on chromosome 5, with only 9. Eighteen members of the PavNAC gene family were distributed on chromosome 1. Twenty-seven members of the PavNAC gene family were distributed on chromosome 4, accounting for approximately 4.89% of the total genome. In addition, genes on the unified chromosome mostly belong to different subgroups in the evolutionary tree.

Figure 2. Localization of PavNAC genes on sweet cherry chromosomes. The left scale bar represents the length (Mb) of each chromosome. The green color represents 8 different chromosomes, and the black short line represents the distribution of the PavNAC genes on each chromosome.
Gene structures and motif analyses of PavNAC genes
The introns and exons are crossed and embedded in a complete gene sequence, and each gene sequence contains an unequal number of introns and exons. Through the structural analysis of PavNAC family genes, it was found that the number of exons and introns in different subgroups were different, and the number in the same group was similar (Figure 3). According to the PavNAC structural analysis chart, the number of introns varies greatly, ranging from 0 to 10, and the number of exons ranges from 1 to 11. PavNAC38 contains the most introns (10) and exons (11). Most PavNAC members contain three exons, which may be due to the relatively conservative domain of the NAC gene family (Figure 3). In order to further understand the structural characteristics of PavNAC gene family proteins, MEME software was used to analyze the conserved motifs of sweet cherry NAC proteins and obtained 10 motif information. It can be seen from the figure that motif 1, motif 2, motif 3, motif 4, motif 5 and motif 6 are present in more than 70% of sweet cherry PavNAC family members, and the family members containing motif 1, motif 3 and motif 6 are up to 90%. The motif sequence of most PavNAC members is: motif 1, motif 5, motif 4, motif 3, motif 2 and motif 6, indicating that PavNAC protein has similar motif composition. However, there are also a few PavNAC proteins with different motif composition.

Figure 3. Phylogenetic relationship, motif analysis, and gene structure of PavNAC genes in sweet cherry. The left image is the phylogenetic tree of PavNAC proteins. The middle image represents the exon intron structure of the PavNAC genes. The black long line represents the length of the introns, and the yellow boxes represent the number of exons. The right image shows the conserved motifs in the PavNAC protein identified by MEME. The gray lines represent non conservative sequences, and the 10 motifs are represented by different colored boxes.
Collinearity analysis of PavNAC genes
In order to further explore the evolution of PavNAC gene and the evolution relationship of NAC gene between different species, a collinearity analysis was carried out on Arabidopsis and sweet cherry (Figure 4). The result showed that there are 54 PavNAC genes with collinearity relationship between sweet cherry and Arabidopsis. In addition, the fifth chromosome in sweet cherry had the most homologous gene with Arabidopsis.

Figure 4. Homology analysis of NAC genes between sweet cherry and Arabidopsis. The red line represents the collinear relationship between sweet cherry and Arabidopsis. The orange bars at the top represent the five chromosomes of Arabidopsis, where Pt denotes the chloroplast genome and Mt denotes the mitochondrial genome of Arabidopsis. The green bars at the bottom represent the eight chromosomes of sweet cherry.
HeatMap of PavNAC genes
In order to study the expression characteristics of PavNAC genes during the growth and development of different sweet cherry cultivars, we conducted a study on the expression characteristics of 132 PavNAC gene family members in different tissues and organs by using the transcriptome data of ‘Black Pearl’ and ‘Hong Deng’ (Figure 5). During the fruit development of ‘Black Pearl’, the expression of PavNAC71/6/48/22/24/8/70 showed a higher expression level than other detected genes (Figure 5A). The expression of PavNAC71/22/24/70 showed a higher expression level than other detected genes in peel and flesh of ‘Hong Deng’ (Figure 5B).

Figure 5. Heat map of PavNAC genes in different sweet cherry varieties. (A) The expression heat map of PavNAC genes in ‘Black Pearl’, from left to right, shows the different development stages at 10, 25, 33, and 40 DAFB. (B) Heat map of PavNAC genes in skin and flesh of ‘Hong Deng’. PHD1 stands for skin of 33 DAFB, FHD1 stands for flesh of 33 DAFB, PHD2 stands for skin of 48 DAFB, and FHD2 stands for flesh of 48 DAFB. The icon on the far right represents the relative expression value, with red indicating induction and blue indicating inhibition.
Expression of PavNAC genes under different abiotic stresses
To investigate the regulatory roles of NAC family genes in the abiotic stress response of sweet cherry, we identified PavNAC genes that were either orthologs of known stress-responsive Arabidopsis NAC genes or clustered within the same phylogenetic clades. Based on this analysis, 15 PavNAC genes were selected for further study. Finally, 15 genes including PavNAC9, PavNAC10,, PavNAC12, PavNAC13, PavNAC18, PavNAC54, PavNAC70, PavNAC81, PavNAC92, PavNAC93, PavNAC107, PavNAC113, PavNAC117, PavNAC118 and PavNAC119 were quantitatively analyzed through qRT-PCR. The results showed that low temperature stress promoted the expression of most PavNAC genes (Figure 6). The expression levels of PavNAC9, PavNAC10, PavNAC70 and PavNAC81 under low temperature stress were higher than those under normal temperature, and they all showed a trend of first increasing and then decreasing, reaching the peak on the fourth day of stress. Among them, PavNAC70 exhibited a markedly high expression level of 50, which corresponds to an approximately 50-fold upregulation relative to the control, under low-temperature stress at the fourth day. To further investigate the regulatory role of the PavNAC genes in response to high salt and PEG, quantitative analysis was conducted on these 15 genes. The results showed that most PavNAC genes were upregulated under high salt stress, with the peak expression of PavNAC9, PavNAC13, PavNAC18, PavNAC70, PavNAC92, PavNAC93 and PavNAC118 on the fifth day (Figure 7). PavNAC93 showed the most significant expression response, with an expression level of up to 4000. High salt stress caused PavNAC93 to show an expression value of approximately 4000, which is 4000-fold higher than the control. Under PEG treatment, most PavNAC genes showed a downward expression trend, with PavNAC12, PavNAC13, PavNAC18, PavNAC54, PavNAC81, PavNAC107, PavNAC113 and PavNAC119 being significantly expressed on the first day. Among these, PavNAC18 and PavNAC107 displayed the highest expression levels, reaching values of up to 100. It can be seen that different PavNAC genes in sweet cherry can exert physiological regulatory functions in different abiotic stresses.

Figure 6. The relative expression level of PavNAC genes under low-temperature treatment. The black lines represent seedlings in the room temperature growth chamber as control. The red lines represent seedlings treated with low temperature. Data are presented as mean ± SD (n = 3). One-way ANOVA followed by Fisher’s LSD post-hoc test revealed statistically significant differences among the groups (p < 0.05).

Figure 7. The relative expression levels of PavNAC genes under drought and salt stress. The black lines represent MS medium as the control group. The red and blue lines represent the culture medium treated with salt and PEG, respectively. Data are presented as mean ± SD (n = 3). One-way ANOVA followed by Fisher’s LSD post-hoc test revealed statistically significant differences among the groups (p < 0.05).
Discussion
There are various transcription factors in higher plants, many of which are related to their stress resistance (Gong et al., 2014). NAC transcription factors, as the largest class of plant specific transcription factors, can also enhance plant adaptability to stress (Li F. et al., 2019; Lin et al., 2023). NAC transcription factors are highly conserved at the N-terminus and highly differentiated at the C-terminus, which may enable the NAC gene family to control protein functions and affect plant growth and development (Olsen et al., 2005; Puranik et al., 2012). Although it has been identified in various plants such as Arabidopsis (Ooka et al., 2003), tomato (Su et al., 2014), celery (Duan et al., 2020), and rice (Meng et al., 2009), there is still a lack of research in sweet cherry.
This study conducted a genome-wide identification and analysis of the PavNAC transcription factor family in sweet cherry. The reliability of this research process primarily depended on high-quality genome assembly and precise genome annotation. As highlighted by Yang et al., the advent of high-quality long-read sequencing technologies and advanced assembly algorithms has propelled genome assembly into the telomere-to-telomere (T2T) era, establishing a robust foundation for the accurate identification of various gene family members (Yang et al., 2025). Building upon this resource, we successfully identified 132 PavNAC genes, effectively circumventing gene omission resulting from assembly gaps. Upon obtaining the complete gene set, precise gene annotation was crucial for ensuring the accuracy of all subsequent analyses. Lan et al. emphasized that annotation quality directly determines the accuracy of gene boundaries and intron-exon structures (Lan et al., 2025). In this study, both the phylogenetic analysis of PavNAC genes and the identification of conserved motifs relied on this accurate initial annotation. Furthermore, the selection of 15 key PavNAC candidate genes in sweet cherry for qRT-PCR validation also depended on the well-established genome annotations of Arabidopsis thaliana. In future research, integrating additional experimental evidence, such as long-read transcriptome data, will enable further refinement and expansion of the PavNAC gene models, thereby providing a solid foundation for investigating the regulatory functions of NAC transcription factors in abiotic stress resistance in sweet cherry.
Using the genome data of sweet cherry, this study identified 132 PavNAC genes, which were lower than that of rice (151 NAC genes) (Nuruzzaman et al., 2010), soybean (152 NAC genes) (Le et al., 2011) and tobacco (152 NAC genes) (Rushton et al., 2008), but higher than that of eggplant (49 NAC genes) (Wang et al., 2021) and Arabidopsis (117 NAC genes) (Nuruzzaman et al., 2010). The number of NAC gene family member in different species is different, which indicates that higher plants have experienced different types of gene replication in the process of evolution, and the number of similar gene families also indicates that they may have similar physiological functions. The NAC transcription factors of sweet cherry and Arabidopsis build a phylogenetic tree with 10 subgroups (Figure 1). The NAC transcription factors of Arabidopsis and sweet cherry are distributed in each subgroup, but there are differences in the number of NAC transcription factors of Arabidopsis and sweet cherry in each subgroup, indicating that the NAC family genes have the phenomenon of gene replication or loss in the long-term evolution process. Genes can be amplified in a variety of ways, such as genome replication, tandem replication, fragment replication and reverse locus replication (Wang et al., 2007). The results of chromosome mapping showed that PavNAC was widely distributed in sweet cherry genome, and there was a tandem repeat relationship, indicating that tandem repeat was an important amplification method of PavNAC gene in sweet cherry (Figure 2). In addition, the gene structure analysis showed that most PavNAC members contained two introns, and the conserved domains were mostly at the N-terminal, which was similar to the NAC gene structure in other plants (Figure 3). In general, the frequency of intron loss is higher than that of intron acquisition in the process of gene evolution. PavNAC97 and PavNAC38 contain more introns, and they may be the early members of PavNAC. PavNAC members belonging to the same subgroup have similar exon intron structures, which may be related to their similar functions. The results of domain analysis showed that in the same subgroup, PavNAC members contained the same number and order of motifs, and different subgroups had different motifs or specific motifs, which might play different roles. Motif 1, motif 3 and motif 6 were the most abundant among all PavNAC transcription factors. The results of collinearity analysis showed that the fifth chromosome was the most collinear chromosome with Arabidopsis thaliana, indicating that the NAC genes on this chromosome may retain orthologous relationships with those in Arabidopsis, making them strong candidates for functional studies based on known Arabidopsis gene functions (Figure 4).
The NAC gene family also plays an important role in promoting plant growth and development, such as leaf senescence, ovule development, seed coat formation, fruit ripening (Dong et al., 2019). The expression levels of the PavNAC gene family in two different varieties of sweet cherry at different flowering stages of ‘Black Pearl’ and in the skin and flesh of ‘Hong Deng’ (Figure 5). Heat map analysis shows that both PavNAC42 and PavNAC101 are highly expressed at 25 DAFB in ‘Black Pearl’. It is speculated that PavNAC42 and PavNAC110 are involved in the regulation of plant growth and development in the late flowering stage. PavNAC101 is significantly expressed in PHD compared to FHD in ‘Hong Deng’. The Arabidopsis NAC gene AtNAC2, which is homologous to PavNAC101, is involved in regulating the growth of thin-walled metastatic cell walls. Therefore, it is speculated that PavNAC101 has a similar function (He et al., 2005; Nigarish et al., 2020).
In recent years, numerous transcription factor genes involved in plant responses to low temperature, drought, and high salinity have been investigated (Gong and Liu, 2013). To systematically examine the regulation of PavNAC gene expression under different abiotic stresses, this study also analyzed the effects of high salt, low temperature, and drought stress on the expression patterns of PavNAC genes. The control mechanisms of abiotic stress rely on the activation and regulation of a multitude of stress-related genes. Salt stress adversely affects plant growth and development by disrupting cellular osmotic balance and facilitating the accumulation of harmful substances (Zelm et al., 2020). Studies on the function of the NAC gene family in Arabidopsis have revealed that the AtNAC019 protein can regulate the expression of cold stress-related genes by recognizing and binding to core DNA sequences (Jensen et al., 2010). In our research, we found that PavNAC70, which is homologous to AtNAC019, was highly expressed under low temperature stress, indicating that PavNAC70 is also involved in the response to cold stress (Figure 6). Moreover, the sharp decline in the expression of most genes by the fourth day may suggest that low temperature leads to disruptions in physiological and metabolic activities. Transcriptional regulation constitutes a critical component of the plant response to low temperature stress (Li Y. Y. et al., 2019). In recent years, it has been progressively established that NAC transcription factors play significant roles in salt stress resistance. Research demonstrated that ONAC022 functions as a stress-responsive NAC with transcriptional activator activity, playing a positive role in drought and salt stress tolerance by modulating ABA-mediated pathways (Hong et al., 2016). AtNAC019, AtNAC055, and AtNAC072 are responsive to various abiotic stresses in Arabidopsis, including drought, low temperature, and high salinity (Tran et al., 2004). In our study, real-time PCR analysis revealed a particularly high expression level of PavNAC93 in the high salt treatment group after four days (Figure 7). Furthermore, it was observed that different genes reached distinct peak expression levels at different time points, suggesting that the response of PavNAC genes to abiotic stress may also depend on the duration and intensity of the stress. The NAC transcription factor NTL4 (AtNAC053) promotes reactive oxygen species production during salt stress-induced leaf senescence in Arabidopsis, thereby mitigating the negative effects of high salinity. Given that AtNAC053 is homologous to PavNAC93 in the phylogenetic tree, it is hypothesized that PavNAC93 may perform a similar function (Lee et al., 2012). The role of this gene in the stress response of sweet cherry warrants further investigation. Under drought stress, the expression of most genes exhibited a trend opposite to that under high salt stress, with high expression observed as early as the first day of PEG-simulated drought treatment. This indicates that the impact of high salt stress becomes more pronounced during later stages, underscoring the need for greater attention to the potential damage caused by salt stress in practical agricultural production. Three key NAC genes in Arabidopsis—AtNAC019, AtNAC055, and AtNAC072—have been confirmed to play important roles in drought stress response. PavNAC81 in sweet cherry is homologous to these three Arabidopsis genes, suggesting that PavNAC81 may perform a similar function in the drought stress response of sweet cherry. In addition to the 15 PavNAC genes investigated in this study, it is likely that numerous other genes are also involved in the stress response. Our findings further support the roles of PavNAC genes in responding to low temperature, drought, and high salinity stresses. Furthermore, by analyzing the phylogenetic relationship between sweet cherry PavNAC genes and NAC genes from other species, this study has laid the groundwork for further investigation into NAC gene functions.
Conclusions
In this study, 132 PavNAC transcription factors in sweet cherry genome was identified. In addition, the expression patterns of PavNAC gene in tissues of different varieties were analyzed, and it was found that PavNAC42 and PavNAC101 were specifically expressed in ‘Black Pearl’ and ‘Hong Deng’ varieties, respectively. To verify the resistance of sweet cherry to abiotic stress, partial PavNAC genes were selected for qRT-PCR analysis. Research has found that PavNAC70 is highly expressed under low temperature stress, while PavNAC93 is highly expressed under high salt and PEG stress. These data will provide scientific basis for future research work, as well as excellent genetic resources for improving the stress resistance of sweet cherry through genetic engineering methods.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author contributions
XS: Investigation, Visualization, Writing – original draft. XA: Formal Analysis, Investigation, Visualization, Writing – original draft. CT: Writing – original draft. AZ: Writing – review & editing. JL: Conceptualization, Supervision, Writing – review & editing. SS: Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, and/or publication of this article. This research was funded by the Scientific and Technological Innovation Development Plan of Yantai, grant number 2023JCYJ089.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1712680/full#supplementary-material
References
Aida, M., Ishida, T., Fukaki, H., Fujisawa, H., and Tasaka, M. (1997). Genes involved in organ separation in arabidopsis: an analysis of the cup-shaped cotyledon mutant. Plant Cell 9, 841–857. doi: 10.1105/tpc.9.6.841
Balazadeh, B., Kwasniewski, M., Caldana, C., Mehrnia, M., Zanor, M. I., Xue, G. P., et al. (2011). ORS1, an H2O2-responsive NAC transcription factor, controls senescence in Arabidopsis thaliana. Mol. Plant 4, 346–360. doi: 10.1093/mp/ssq080
Chi, Y. H., Melencion, S. M. B., Alinapon, C. V., Kim, M. J., Lee, E. S., Paeng, S. K., et al. (2017). The membrane-tethered NAC transcription factor, AtNTL7, contributes to ER-stress resistance in Arabidopsis. Biochem. Biophys. Res. Commun. 488, 641–647. doi: 10.1016/j.bbrc.2017.01.047
Dong, X. S., Jiang, Y., Yang, Y. N., Xiao, Z. W., Bai, X. H., Gao, J., et al. (2019). Identification and expression analysis of the NAC gene family in coffea canephora. Agronomy 9, 670. doi: 10.3390/agronomy9110670
Duan, A. Q., Yang, X. L., Feng, K., Liu, J. X., Xu, Z. S., and Xiong, A. S. (2020). Genome-wide analysis of NAC transcription factors and their response to abiotic stress in celery (Apium graveolens L.). Comput. Biol. Chem. 84, 107186. doi: 10.1016/j.compbiolchem.2019.107186
Gao, L. and Mazza, G. (1995). Characterization, quantitation, and distribution of anthocyanins and colorless phenolics in Sweet Cherries. Food Chem. 43, 305–316. doi: 10.1021/jf00050a015
Gao, Y. M., Liu, H. L., Wu, L., Shi, Y. N., and Xiang, Y. (2020). Systematic identification and analysis of NAC gene family in moso bamboo (Phyllostachys edulis). BMC Genomics 12, 4064. doi: 10.21203/rs.3.rs-18155/v1
Gong, X. Q., Hu, J. B., and Liu, J. H. (2014). Cloning and characterization of FcWRKY40,A WRKY transcription factor from Fortunella crassifolia linked to oxidative stress tolerance. Plant Cell 119, 197–210. doi: 10.1007/s11240-014-0526-0
Gong, X. Q. and Liu, J. H. (2013). Genetic transformation and genes for resistance to abiotic and biotic stresses in Citrus and its related genera. Plant Cell 113, 137–147. doi: 10.1007/s11240-012-0267-x
He, X. J., Mu, R. L., Cao, W. H., Zhang, Z. G., Zhang, J. S., and Chen, S. Y. (2005). AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 44, 903–916. doi: 10.1111/j.1365-313X.2005.02575.x
Hong, Y. B., Zhang, H. J., Huang, L., Li, D. Y., and Song, F. M. (2016). Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 22 7, 4. doi: 10.3389/fpls.2016.00004
Jensen, M. K., Kjaersgaard, T., Petersen, K., and Skriver, K. (2010). NAC genes: time-specific regulators of hormonal signaling in Arabidopsis. Plant Signal Behav. 5, 907–910. doi: 10.4161/psb.5.7.12099
Jiang, D. G., Zhou, L. Y., Chen, W. T., Ye, N. H., Xia, J. X., and Zhuang, C. X. (2019). Overexpression of a microRNA-targeted NAC transcription factor improves drought and salt tolerance in Rice via ABA-mediated pathways. Rice 12, 76. doi: 10.1186/s12284-019-0334-6
Kim, S. G., Lee, A. K., Yoon, H. K., and Park, C. M. (2008). A membrane-bound NAC transcription factor NTL8 regulates gibberellic acid-mediated salt signaling in Arabidopsis seed germination. Plant J. 55, 77–88. doi: 10.1111/j.1365-313X.2008.03493.x
Lan, L., Hu, H. F., Jia, Y., Zhang, X. N., Jia, M. L., Li, C. D., et al. (2025). Tips for improving genome annotation quality. gComm 2, e005. doi: 10.48130/gcomm-0025-0006
Latchman, D. S. (1997). Transcription factors: an overview. Int. J. Biochem. Cell Biol. 29, 1305–1312. doi: 10.1016/S1357-2725(97)00085-X
Le, D. T., Nishiyama, R., Watanabe, Y., Mochida, K., Yamaguchi-Shinozak, K., Shinozaki, K., et al. (2011). Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res. 18, 263–276. doi: 10.1093/dnares/dsr015
Lee, S., Seo, P. J., Lee, H. J., and Park, C. M. (2012). A NAC transcription factor NTL4 promotes reactive oxygen species production during drought-induced leaf senescence in Arabidopsis. Plant J. 70, 831–844. doi: 10.1111/j.1365-313X.2012.04932.x
Li, F., Guo, X. H., Liu, J. X., Zhou, F., Liu, W. Y., Wu, J., et al. (2019). Genome-wide identification, characterization, and expression analysis of the NAC transcription factor in chenopodium quinoa. Genes 10, 500. doi: 10.3390/genes10070500
Li, Y. Y., Wang, X. W., Ban, Q. Y., Zhu, X. X., Jiang, C. J., Wei, C. L., et al. (2019). Comparative transcriptomic analysis reveals gene expression associated with cold adaptation in the tea plant Camellia sinensis. BMC Genomics 20, 624. doi: 10.1186/s12864-019-5988-3
Lin, H. B., Luan, X. Y., Chen, C. H., Gong, X., Li, X. Q., Li, H. H., et al. (2023). A systematic genome-wide analysis and screening of the NAC family genes related to wood formation in Cinnamomum camphora. Genomics 115, 110631. doi: 10.1016/j.ygeno.2023.110631
Liu, J. H., Peng, T., and Dai, W. S. (2014). Critical cis-acting elements and Interacting transcription factors: key players associated with abiotic stress responses in plants. Plant Mol. Biol. Rep. 32, 303–317. doi: 10.1007/s11105-013-0667-z
Liu, C., Wang, B., Li, Z. X., Peng, Z. H., and Zhang, J. R. (2017). TsNAC1 is a key transcription factor in abiotic stress resistance and growth. Plant Physiol. 176, 742–756. doi: 10.1104/pp.17.01089
Ma, X. M., Zhang, Y. J., Turečková, V., Xue, G. P., Fernie, A. R., Mueller-Roeber, B., et al. (2018). The NAC transcription factor SlNAC2 regulates leaf senescence and fruit yield in tomato. Plant Physiol. 177, 1286–1302. doi: 10.1104/pp.18.00292
Meng, Y. J., Huang, F. L., Shi, Q. Y., Cao, J. J., Chen, D. J., Zhang, J. W., et al. (2009). Genome-wide survey of rice microRNAs and microRNA–target pairs in the root of a novel auxin-resistant mutant. Planta 230, 883–898. doi: 10.1007/s00425-009-0994-3
Nakashima, K., Takasaki, H., Mizoi, J., Shinozaki, K., and Yamaguchi-Shinozaki, K. (2012). NAC transcription factors in plant abiotic stress respanses. Biochim. Biophys. Acta 1819, 97–103. doi: 10.1016/j.bbagrm.2011.10.005
Nie, G., Yang, X. Y., Yang, Z. F., Zhong, M. Y., Zhu, Y. Q., Zhou, J., et al. (2020). Genome-wide investigation of the NAC transcript factor family in perennial ryegrass (Lolium perenne L.) and expression analysis under various abiotic stressor. Genomics 112, 4224–4231. doi: 10.1016/j.ygeno.2020.06.033
Nigarish, M., Chen, Y. K., Chen, X. H., Muhammad, A. N., Junaid, I., Hafiz, M. R., et al. (2020). Genome-wide identification and comprehensive analyses of NAC transcription factor gene family and expression patterns during somatic embryogenesis in Dimocarpus longan lour. Plant Physiol. Biochem. 157, 169–184. doi: 10.1016/j.plaphy.2020.10.009
Nogueira, F. T. S., Schlogl, P. S., Camargo, S. R., Fernandez, J. H., De Rosa, V. E., Jr, Pompermayer, P., et al. (2005). SsNAC23, a member of the NAC domain protein family, is associated with cold, herbivory and water stress in sugarcane. Plant Sci. 169, 93–106. doi: 10.1016/j.plantsci.2005.03.008
Nuruzzaman, M., Manimekalai, R., Sharoni, A. M., Satoh, K., Kondoh, H., Ooka, H., et al. (2010). Genome-wide analysis of NAC transcription factor family in rice. Gene 465, 30–44. doi: 10.1016/j.gene.2010.06.008
Olsen, A. N., Ernst, H. A., Leggio, L. L., and Skriver, K. (2005). NAC transcription factors: structurally distinct, functionally diverse. Trends Plant Sci. 10, 79–87. doi: 10.1016/j.tplants.2004.12.010
Ooka, H., Satoh, K., Doi, K., Nagata, T., Otomo, Y., Murakami, K., et al. (2003). Comprehensive analysis of nac family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 10, 239–247. doi: 10.1093/dnares/10.6.239
Puranik, S., Sahu, P. P., Srivastava, P. S., and Prasad, M. (2012). Nac proteins: regulation and role in stress tolerance. Trends Plant Sci. 17, 369–381. doi: 10.1016/j.tplants.2012.02.004
Rushton, P. J., Bokowiec, M. T., Han, S. C., Zhang, H. B., Brannock, J. F., Chen, X. F., et al. (2008). Tobacco transcription factors: novel insights into transcriptional regulation in the Solanaceae. Plant Physiol. 147, 280–295. doi: 10.1104/pp.107.114041
Shan, W., Kuang, J. F., Lu, W. J., and Chen, J. Y. (2014). Banana fruit NAC transcription factor MaNAC1 is a direct target of MaICE1 and involved in cold stress through interacting with MaCBF1. Plant Cell Environ. 37, 2116–2127. doi: 10.1111/pce.12303
Shen, S. Y., Zhang, Q. R., Shi, Y., Sun, Z. M., Zhang, Q. Q., Hou, S. J., et al. (2020). Genome-wide analysis of the NAC domain transcription factor gene family in theobroma cacao. Genes 11, 35. doi: 10.3390/genes11010035
Singh, S., Koyama, H., Bhati, K. K., and Alok, A. (2021). The biotechnological importance of the plant-specifc NAC transcription factor family in crop improvement. J. Plant Res. 134, 475–495. doi: 10.1007/s10265-021-01270-y
Su, H. Y., Zhang, S. Z., Yin, Y. L., Zhu, D. Z., and Han, L. Y. (2014). Genome-wide analysis of NAM-ATAF1,2-CUC2 transcription factor family in Solanum lycopersicum. J. Plant Biochem. Biotechnol. 71, 11–21. doi: 10.1007/s13562-014-0255-9
Tran, L. S. P., Nakashima, K., Sakuma, Y., Simpson, S. D., Fujita, Y., Maruyama, K., et al. (2004). Isolation and functional analysis of arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 16, 2481–2498. doi: 10.1105/tpc.104.022699
Vroemen, C. M., Mordhorst, A. P., Albrecht, C., Kwaaitaal, M. A. C. J., and de Vries, S. C. (2003). The cup-shaped cotyledon3 gene is required for boundary and shoot meristem formation in arabidopsis. Plant Cell 15, 1563–1577. doi: 10.1105/tpc.012203
Wan, F. X., Gao, J., Wang, G. L., Niu, Y., Wang, L. Z., Zhang, X. G., et al. (2021). Genome-wide identification of NAC transcription factor family and expression analysis of ATAF subfamily members under abiotic stress in eggplant. Sientia Hortic., 110424. doi: 10.1016/j.scienta.2021.110424
Wang, Q., Guo, C., Li, Z. Y., Sun, J. H., Deng, Z. C., Wen, L. C., et al. (2021). Potato NAC transcription factor StNAC053 enhances salt and drought tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 22, 2568. doi: 10.3390/ijms22052568
Wang, D. K., Pei, K. M., Fu, Y. P., Sun, Z. X., Li, S. J., Liu, H. Q., et al. (2007). Genome-wide analysis of the auxin response factors (ARF) gene family in rice (Oryza sativa). Genes 394, 13–24. doi: 10.1016/j.gene.2007.01.006
Wu, H., Fu, B., Sun, P. P., Xiao, C., and Liu, J. H. (2016). A NAC transcription factor represses putrescine biosynthesis and affects drought tolerance. Plant Physiol. 172, 1532–1547. doi: 10.1104/pp.16.01096
Xie, Q., Frugis, G., Colgan, D., and Chua, N. H. (2000). Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 14, 3024–3036. doi: 10.1101/gad.852200
Yang, Y. X., Du, W. J., Li, Y. C., Lei, J. W., and Pan, W. H. (2025). Recent advances and challenges in de novo genome assembly. gComm 2, e014. doi: 10.48130/gcomm-0025-0015
Zelm, E. V., Zhang, Y. X., and Testerink, C. (2020). Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 71, 403–433. doi: 10.1146/annurev-arplant-050718-100005
Zhang, L. N., Zhang, L. C., Xia, C., Zhao, G. Y., Jia, J. Z., and Kong, X. Y. (2016). The novel wheat transcription factor tanac47 enhances multiple abiotic stress tolerances in transgenic plants. Front. Plant Sci. 6, 109415. doi: 10.3389/fpls.2015.01174
Keywords: sweet cherry, NAC transcription factors, expression analysis, abiotic stress, genome wide analysis
Citation: Shi X, Ai X, Tian C, Zhang A, Li J and Sun S (2025) Genome-wide investigation and expression analysis of sweet cherry PavNAC gene family under different abiotic stress. Front. Plant Sci. 16:1712680. doi: 10.3389/fpls.2025.1712680
Received: 25 September 2025; Accepted: 05 November 2025; Revised: 05 November 2025;
Published: 24 November 2025.
Edited by:
Zhiqiang Wu, Chinese Academy of Agricultural Sciences, ChinaReviewed by:
Guang-Long Wang, Huaiyin Institute of Technology, ChinaQuanjuan Fu, Shandong Institute of Pomology, China
Copyright © 2025 Shi, Ai, Tian, Zhang, Li and Sun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jianzhao Li, emhhb2ppYW5saTk1QDE2My5jb20=; Shuyang Sun, c3lzdW44MUBsZHUuZWR1LmNu
†These authors have contributed equally to this work