 Wenxin Xu1,2,3†Tangxiu Li1†
Wenxin Xu1,2,3†Tangxiu Li1† Jing Wu4You Wang2,3
Jing Wu4You Wang2,3 Junting Feng5Aiping Luan2,3Okandze Poho Pérol Carzorel6Shuqiang He2,3Junhu He2,3*
Junting Feng5Aiping Luan2,3Okandze Poho Pérol Carzorel6Shuqiang He2,3Junhu He2,3* Chengjie Chen2,3*
Chengjie Chen2,3* Wuqiang Ma1*
Wuqiang Ma1*- 1Sanya Institute of Breeding and Multiplication & Key Laboratory of Quality Regulation of Tropical Horticultural Crop in Hainan Province, School of Tropical Agriculture and Forestry, Hainan University, Sanya, Hainan, China
- 2State Key Laboratory of Tropical Crop Breeding, Key Laboratory of Crop Gene Resources and Germplasm Enhancement in South China, Ministry of Agriculture and Rural Affairs, Haikou, Hainan, China
- 3Key Laboratory of Tropical Crops Germplasm Resources Genetic Improvement and Innovation of Hainan Province, Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan, China
- 4Haikou Experimental Station Chinese Academy of Tropical Agricultural Sciences, Chinese Academy of Tropical Agricultural Sciences, Haikou, Hainan, China
- 5State Key Laboratory of Tropical Crop Breeding, Sanya Research Institute, Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences, Sanya, Hainan, China
- 6National Centre for Crop Disease Control, Ministry of Agriculture, Animal Husbandry and Fisheries, Brazzaville, Republic of Congo
Carboxylesterases (CXEs) are ester hydrolyzing enzymes closely associated with the degradation of volatile esters and aroma release in fruit. Pineapple possesses a strong, ester-rich aroma, yet its CXE gene family has not been characterized. Here, we performed a genome-wide identification and analysis of the pineapple CXE family, detected 20 members, and pinpointed AcCXE4 and AcCXE7 as key negative regulators of aroma accumulation and prime candidates for aroma-oriented breeding. Phylogenetic comparison with CXEs from Arabidopsis, tomato, strawberry, Nanguo pear, and peach resolved five clades, in which AcCXE4 grouped with PuCXE15, a reported aroma-related gene in Nanguo pear, while AcCXE7 clustered with tomato SlCXE1 and AcCXE3 with apple MdCXE1, suggesting these members participate in ester metabolism in pineapple and are potential contributors to fruit-aroma formation. Besides, although AcCXE13 and AcCXE20 show conserved gene structure and sequence, their exon numbers and motif architectures differ from other AcCXEs, implying functional specialization. Comparative genomics indicated that family expansion in pineapple was driven primarily by tandem duplication and large segmental duplication. Integrating cis-regulatory element profiling, transcriptome analyses, and experimental validation, we found that most AcCXEs are likely responsive to light and hormone signaling (including the jasmonate pathway) and to abiotic stress cues. Several AcCXE genes exhibited decreasing expression across cultivars, tissues, and developmental stages, showing negative correlations with aroma accumulation, with AcCXE4 and AcCXE7 displaying the strongest association with pineapple aroma formation. Collectively, this work systematically defines the pineapple CXE family and highlights priority targets to inform molecular improvement of fruit aroma.
Introduction
Pineapple (Ananas comosus (L.) Merr.) is one of the three major tropical fruits worldwide and is renowned for its distinctive, attractive aroma (Ali et al., 2020). In production, cultivar differences, fruit maturity, climate variability, cultivation practices (Coelho et al., 2024), and biotic stresses (Larrea-Sarmiento et al., 2022) directly influence the intensity and composition of pineapple fruit aroma, thereby affecting market quality. Over the past decade, studies have shown that during ripening, a substantial accumulation of esters, driven by accelerated biosynthesis and reduced hydrolysis, underpins pineapple’s characteristic flavor profile (Montero‐Calderón et al., 2010; Steingass et al., 2015; George et al., 2023, 2024).
The accumulation of ester aroma compounds in fruit is governed by two opposing processes, biosynthesis and degradation, with the latter largely mediated by carboxylesterases (CXEs). CXEs belong to the α/β-hydrolase superfamily, are widespread in plants, animals, and microbes, and are characterized by a conserved catalytic triad and a GXSXG motif (Kim et al., 1997), features that confer hydrolytic activity toward diverse ester substrates (Martínez-Rivas et al., 2022). In plants, the CXE family typically comprises on the order of a dozen to several dozen members, with copy number shaped by species ploidy and lineage-specific diversification. For example, 20 CXEs have been identified in Arabidopsis (Marshall et al., 2003), 33 in peach (Cao et al., 2019a), and 35 in grapevine (Zhang et al., 2022). Studies in Nanguo pear, apple, and peach have demonstrated that CXEs influence fruit-aroma formation by promoting ester degradation, exhibiting functional diversification and substrate specificity across taxa (Souleyre et al., 2011; Cao et al., 2019b; Qi et al., 2023). Recent advances in other economically important fruits have further expanded the understanding of CXEs-related aroma metabolism. In mango, revealed that rapid ripening involves dynamic transformations of aldehydes and esters driven by the catabolism of linoleic and linolenic acids, highlighting the crucial role of fatty acid-derived substrates in volatile formation (Wang et al., 2025). Similarly, studies in citrus have shown that CXEs-like and other hydrolase genes are associated with the modulation of terpenoid composition during fruit maturation, suggesting that esterases may also participate indirectly in the remodeling of terpene-derived volatiles (Hu et al., 2024). These findings underscore that CXE-mediated volatile metabolism represents a conserved yet functionally diversified mechanism among fruit species, providing a valuable reference for elucidating the aroma formation process in pineapple.
Despite esters being the predominant constituents of pineapple aroma, the CXE family underlying ester metabolism in pineapple has not been systematically characterized. Using the latest pineapple reference genome and annotations (Feng et al., 2024), we conducted a comprehensive analysis of the CXE gene family, including genome-wide identification, phylogenetic and duplication-origin analyses, sequence characterization, and expression profiling across germplasm with contrasting aroma intensity/composition and across fruit ripening stages, complemented by preliminary experimental validation. These results provide mechanistic insight into the molecular basis of pineapple aroma and establish foundational resources for its genetic improvement.
Materials and methods
Materials
Fruits of ‘Hongmi’ (HM), ‘Xiangshui’ (XS), ‘Mangguo’ (MG), ‘Weiduoliya’ (WD), and ‘HongXiangshui’ (HXS) were harvested from the pineapple germplasm orchard of the Tropical Crops Genetic Resources Research Institute, Chinese Academy of Tropical Agricultural Sciences, located at Baodao Xincun, Danzhou, Hainan, China (19°29′17″N, 109°29′4″E; elevation 130 m; mean annual temperature 23 °C; relative humidity 85%; annual precipitation ~1,500 mm). Fruits were sampled at three aroma development stages: the non-aromatic stage (approximately 56 days after flowering, with no detectable aroma by sensory evaluation and a total soluble solids (TSS) content of about 12–13°Brix), the initial aroma stage (approximately 63 days after flowering, with a faint fruity aroma detected and a TSS content of about 14–15°Brix), and the strong aroma stage (approximately 68 days after flowering, characterized by a pronounced sweet aroma and a TSS content of about 15–18°Brix). The stage classification was based on sensory evaluation in combination with days after flowering and total soluble solids content. After harvest, fruits were held in the laboratory for 24 h to equilibrate volatiles and minimize field temperature/humidity effects. Samples were then immediately frozen in liquid nitrogen and stored at -80 °C until analysis.
Physicochemical characteristics of AcCXE family
The pineapple reference genome sequences and gene structural annotation were downloaded from the pineapple genome database (https://ananas.watchbio.cn). CXE protein sequences from Arabidopsis retrieved from UniProt were used as queries for BLASTP (v2.16.0) searches against the pineapple proteome (E-value < 1 × 10-5). Candidate AcCXEs were further screened by homology against the UniProtKB/Swiss-Prot database to remove redundant entries. Conserved domains were predicted with InterPro (https://www.ebi.ac.uk/interpro/, v107.0), and proteins containing the α/β-hydrolase fold (Pfam: PF07859) were retained as CXE candidates. All gene structures of AcCXEs were further curated by GSAman (https://tbtools.cowtransfer.com/s/a11146181df14f, v0.9.53). Physicochemical properties of AcCXE proteins were computed using TBtools-II (v2.363) (Chen et al., 2023), and subcellular localizations were predicted with WoLF PSORT (https://wolfpsort.hgc.jp/).
Phylogenetic analysis of the AcCXE family
CXE protein sequences from Arabidopsis, peach, Nanguo pear, tomato, and apple were retrieved from NCBI, GDR (https://www.rosaceae.org/), and TAIR. Homologs were identified using two approaches hmmsearch and BLASTP (v2.16.0), and redundant entries were removed. The filtered CXE sets from these species were combined with pineapple AcCXEs to infer a maximum-likelihood phylogeny using the “One Step Build a ML Tree” tool in TBtools-II (v2.363). The resulting tree was formatted and annotated in Evolview (https://www.evolgenius.info/evolview/#/treeview).
Chromosomal distribution and synteny analysis of the AcCXE family
AcCXE loci and their annotations were processed in TBtools-II (v2.363) (“One Step MCScanX—Super Fast”) to generate chromosome-level gene-distribution files and map the physical positions of AcCXE genes. Intra-genomic duplication relationships (tandem and segmental) among CXEs were then identified with MCScanX (v1.0.0). The results were visualized using the “Advanced Circos” module in TBtools-II (v2.363).
Gene structure and conserved domain analysis of the AcCXE family
For gene structure and conserved-domain analyses, conserved domains of AcCXE proteins were predicted using NCBI CDD (Batch) (Marchler-Bauer and Bryant, 2004) and Pfam with default parameters. Conserved motifs were identified across the 20 AcCXE proteins using MEME Suite (https://meme-suite.org/meme/, v5.5.8) (Bailey et al., 2009), with the maximum number of motifs set to 10 to capture motif types and counts across subfamilies. Gene structure, conserved motifs, conserved domains, and sequence identifiers were then integrated and visualized in TBtools-II (v2.363).
Promoter cis-acting element analysis of the AcCXE gene family in pineapple
Genomic coordinates of CXE loci were obtained from the GP genome GFF using TBtools-II (v2.363), and the 2,000-bp sequences upstream of the translation start codon (ATG) were extracted as putative promoter regions. These sequences were submitted to PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) for prediction of cis-acting regulatory elements (Lescot et al., 2002). Detected element types were then enumerated, classified, and summarized.
Expression patterns of the AcCXE gene family across cultivars and fruit developmental stages in pineapple
Transcriptome datasets for root, stem, leaf, petal, ovule, and fruit core were obtained from NCBI BioProject PRJNA483249 (Mao et al., 2018). RNA-seq data for different cultivars and developmental stages were generated by our laboratory; three biological replicates per cultivar were sequenced, and expression values were averaged. All reads were quantified with Kallisto (v0.51.1) and normalized as TPM (Bray et al., 2016). Fruits of ‘Hongmi’ (HM), ‘Xiangshui’ (XS), and ‘HongXiangshui’ (HXS) were sampled at three aroma stages (aroma-absent, aroma-onset, aroma-intense). Total RNA was extracted and reverse-transcribed for RT–qPCR. Primer specificity was assessed using Primer5, and primers were synthesized by Wuhan Zhuandao Biotechnology Co., Ltd. Actin served as the reference gene (Yi et al., 2023; Zhang et al., 2024b). Relative expression was calculated with the 2−ΔΔCt method (Rao et al., 2013), with three technical replicates per sample. Statistical analyses were performed in IBM SPSS Statistics 27 using two-tailed t-tests, and figures were prepared in GraphPad Prism 8.0.
Results
Characteristics of AcCXE family members in pineapple
From the pineapple genome, we identified 20 CXE family members and named them AcCXE1–AcCXE20 in ascending order of chromosomal position. Protein physicochemical analysis showed lengths of 169–463 amino acids and predicted isoelectric points (pI) of 4.73–8.93 (Table 1). Subcellular localization prediction indicated 12 proteins in the cytosol and 6 in chloroplasts; AcCXE10 was predicted to localize to the endoplasmic reticulum, and AcCXE9 to the nucleus.

Table 1. Members of the AcCXE gene family in pineapple (Ananas comosus) and their physicochemical properties.
Phylogenetic and evolutionary analysis of the AcCXE family
To elucidate the evolution of the pineapple CXE family, we constructed a maximum-likelihood (ML) phylogeny comprising 147 CXE proteins from six species: 20 from pineapple, 20 from Arabidopsis thaliana, 35 from Nanguo pear (Pyrus ussuriensis), 33 from peach (Prunus persica), 23 from tomato (Solanum lycopersicum), and 16 from apple (Malus domestica), and grouped them accordingly (Figure 1A). The tree resolved five major clades (Group I–V), with Group I and Group II containing the largest numbers of members (Figure 1D), suggesting these clades dominated family expansion. Pineapple CXEs were distributed across all five clades. Notably, AcCXE4 clustered with PuCXE15, a gene implicated in ester degradation in Nanguo pear (Qi et al., 2023); AcCXE7 clustered with tomato SlCXE1 (Goulet et al., 2012), and AcCXE3 with apple MdCXE1 (Souleyre et al., 2011), indicating that AcCXE4, AcCXE7, and AcCXE3 may participate in ester hydrolysis in pineapple. In terms of family size, Nanguo pear and peach harbored the most CXEs (35 and 33, respectively), whereas pineapple possessed 20. Overall, the phylogeny and gene counts indicate clade-level conservation with lineage-specific diversification.

Figure 1. Maximum-likelihood phylogeny of CXE proteins from six species. (A) Phylogenetic relationships of CXE gene families across six species, resolving five clades. (B) Species tree of the six taxa from TimeTree. (C–D) Proportional composition of CXE clades within each species. Ananas comosus (red star), Arabidopsis thaliana (yellow triangle), Pyrus ussuriensis (red square), Prunus persica (blue circle), Solanum lycopersicum (blue triangle), Malus domestica (red circle).
Chromosome distribution and synteny analysis of the AcCXE family
To assess the chromosome distribution of AcCXEs, we visualized their loci with a Circos plot (Figure 2A). The genes are dispersed across 12 chromosomes. Single-copy loci occur on contig04, contig06, contig10, contig16, contig18, and contig23, indicating an overall scattered pattern. Several genes co-localize on the same chromosome, for example, AcCXE1 and AcCXE2 on contig03, and AcCXE17 and AcCXE19 on contig21. Inspection of collinearity links revealed both tandem and segmental duplications that likely contributed to family expansion: AcCXE7-AcCXE8-AcCXE4-AcCXE5 form a tandem array, whereas AcCXE6 and AcCXE16 represent a segmental duplicate pair.

Figure 2. Comparative genomic analysis between pineapple and other species, focusing on CXE gene family. (A) Intragenomic synteny among AcCXE loci. Circos tracks (from inner to outer): GC skew density, gene density, and GC content. (B) Inter-species synteny between pineapple and banana/Solanum lycopersicum (tomato). (C) Inter-species synteny between pineapple and Oryza sativa (rice)/ Arabidopsis thaliana.
Comparative synteny showed conserved collinearity between pineapple CXE loci and those in banana, tomato, rice, and Arabidopsis, albeit with different counts. We detected 14 and 6 syntenic pairs between pineapple and banana or tomato, respectively (Figure 2B), and 15 and 2 pairs between pineapple and rice or Arabidopsis, respectively (Figure 2C). The higher numbers for banana and rice relative to tomato and Arabidopsis suggest substantial divergence in the chromosomal neighborhoods harboring CXE genes between monocots and dicots. Given that banana and tomato are well-studied for fruit aroma, these syntenies provide a useful reference for inferring the roles of pineapple CXEs in volatile ester metabolism and aroma formation.
Conserved motifs, domains and gene structure analysis of AcCXEs
To further explore conserved features of pineapple CXEs, we analyzed their protein domains and motifs. Ten conserved motifs were identified across AcCXE sequences (Figure 3B). Motifs 1, 2, 3, 6, and 10 are highly conserved and present in all members. Motif number and order are broadly consistent within subclades, whereas several genes carry fewer motifs-e.g., AcCXE15-suggesting functional specialization. Gene-structure analysis (Figure 3D) showed that most AcCXEs contain 3–4 exons; members within the same phylogenetic branch share similar architectures (Figure 3A), indicating lineage-specific structural diversification. AcCXE13 and AcCXE20 are conserved at the gene-structure and sequence levels, yet their exon counts and motif compositions differ from most other AcCXEs, implying possible neofunctionalization. All AcCXEs harbor the conserved α/β-hydrolase superfamily domain, and most also contain the Abhydrolase_3 domain.

Figure 3. Conserved features analysis of the AcCXE gene family. From left to right, phylogenetic relationships, conserved motifs, functional domains, and gene structure of AcCXEs, respectively.
Promoter cis-acting element analysis of the AcCXE gene family in pineapple
To explore potential regulatory events of AcCXE genes in abiotic stress and development, we surveyed cis-acting elements within the 2,000-bp upstream promoter regions (Figure 4). After excluding ubiquitous core elements such as the CAAT-box and TATA-box, a total of 452 cis-elements were identified and classified into four major categories: light-responsive, stress-responsive (abiotic), development-related, and hormone-responsive elements.
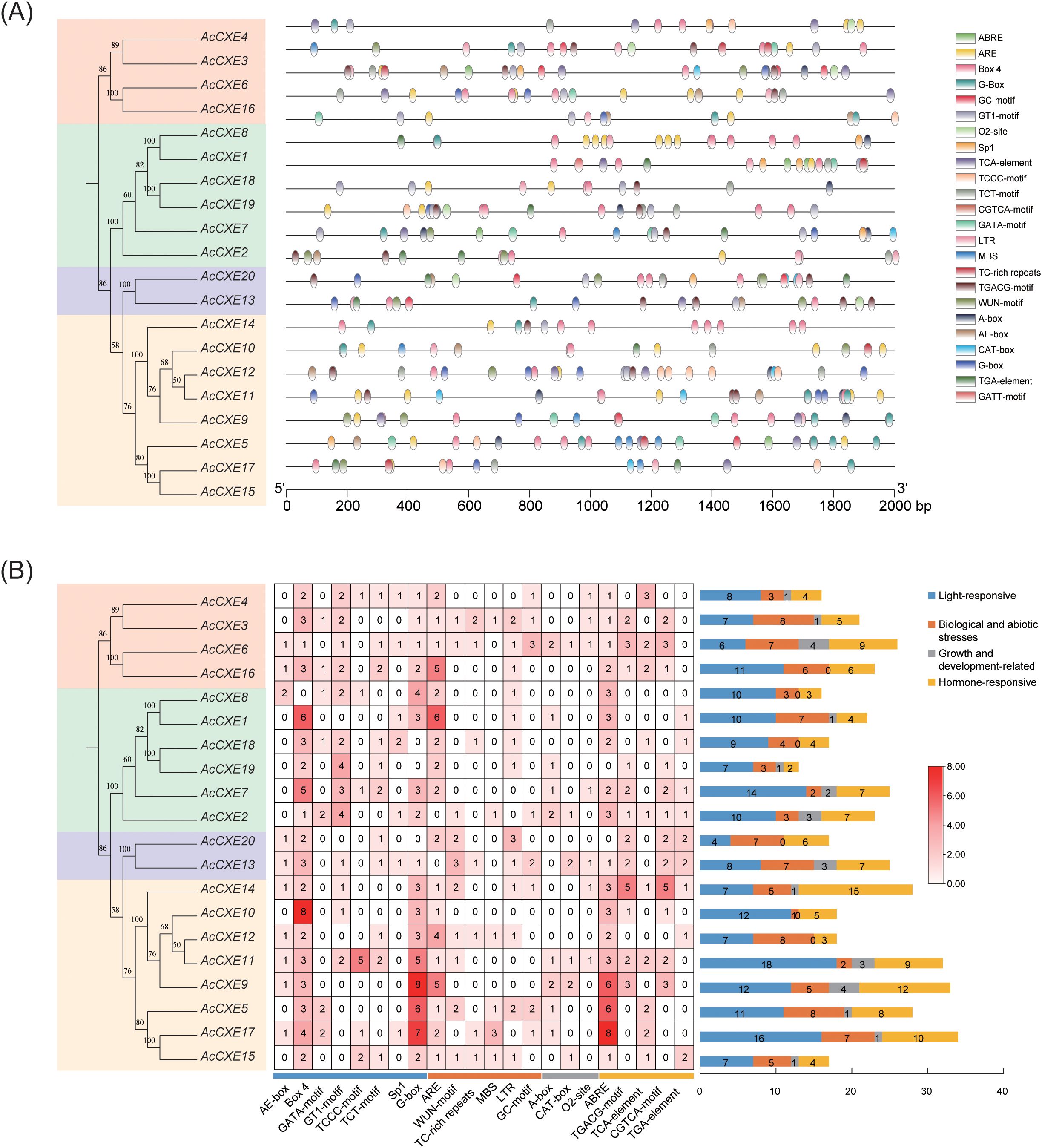
Figure 4. Cis-element analysis of AcCXEs’ promoters. (A) Chromosome distributions of different classes of cis-elements in pineapple CXE gene promoter sequences; (B) Statistics analysis of cis-elements of AcCXEs’ promoter sequences.
Among them, light-responsive cis-elements were abundant—including G-box, Box4, GT1-motif, and TCT-motif. Box4 occurred most frequently in AcCXE10, whereas G-box was most frequent in AcCXE9, suggesting important roles for these genes in light signal transduction. Hormone-responsive elements were enriched for ABRE (ABA-responsive), TGACG-motif (MeJA-responsive), TGA-element (auxin-responsive), and TCA-element (salicylic acid–responsive); the prevalence of MeJA-related motifs indicates extensive involvement of the family in jasmonate-mediated regulation. Stress-associated elements (ARE, MBS, LTR, TC-rich repeats) were common, implicating AcCXEs in responses to low temperature, drought, and anaerobic stress. Development-related elements—including CAT-box, A-box, and O2-site—were also frequent, consistent with regulation of tissue-specific expression and developmental processes.
Expression pattern analysis of the AcCXE gene family across cultivars and developmental stages in pineapple
To characterize the expression profile of AcCXEs across cultivars, tissues, and fruit development, we analyzed RNA-seq datasets. By cultivar (Figure 5A), AcCXE5 and AcCXE7 were highly expressed in the light-aroma cultivar ‘HongXiangshui’ (HXS), consistent with a putative negative role in aroma formation. By tissue (Figure 5B), AcCXE4, AcCXE5, and AcCXE9 showed elevated expression in the fruit core, which exhibits weak aroma, again aligning with negative regulation. Across developmental stages (Figure 5C), AcCXE3, AcCXE4, and AcCXE13 were down-regulated as fruit aroma intensified, suggesting repression of ester accumulation. Collectively, these patterns nominate six candidates, AcCXE3, AcCXE4, AcCXE5, AcCXE7, AcCXE9, and AcCXE13, as key genes associated with pineapple aroma metabolism.
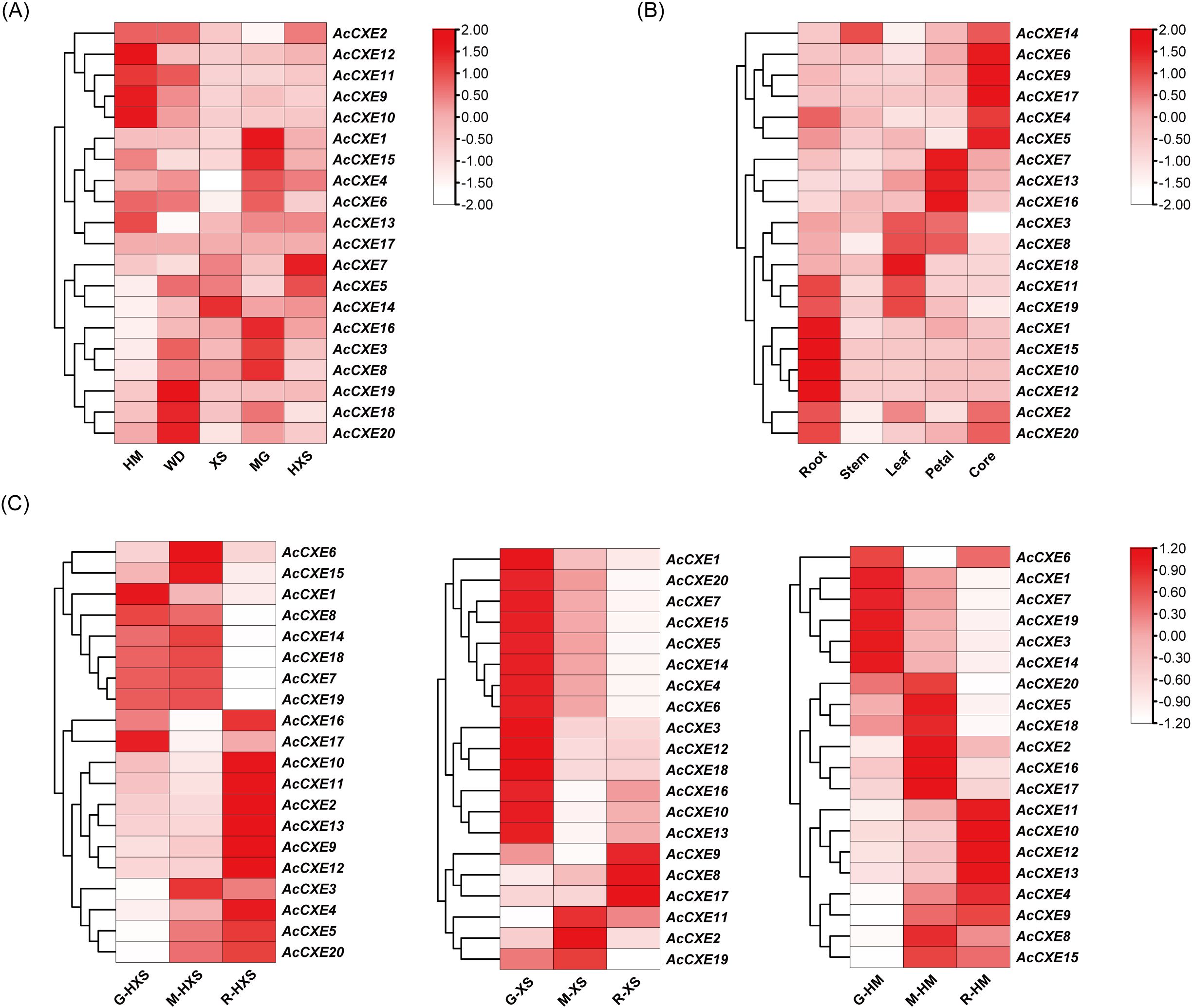
Figure 5. Expression profiles of AcCXEs across different varieties, tissues and fruit development stages. (A) Transcript abundance of AcCXE genes across cultivars. (B) Transcript abundance across tissues. (C) Transcript abundance across aroma-development stages in three aroma-type cultivars (HM, XS, HXS). All expression data are estimated using TPM method, generated from RNA-seq experiments, averaged over three biological replicates.
RT–qPCR validation of AcCXE gene expression in pineapple
Six candidates, AcCXE3, AcCXE4, AcCXE5, AcCXE7, AcCXE9, and AcCXE13, were selected for RT-qPCR validation (Figure 6). AcCXE4 and AcCXE7 showed higher expression in the light-aroma cultivar ‘HongXiangshui’ (HXS) than in the sweet/fruit-aroma cultivars ‘Hongmi’ (HM) and ‘Xiangshui’ (XS). Notably, AcCXE4 expression decreased with ripening in XS but increased in HXS, consistent with a role for CXEs as negative regulators of ester accumulation. These results implicate AcCXE4 and AcCXE7 as key genes closely associated with pineapple aroma formation.

Figure 6. Relative qRT-PCR expression of six AcCXE genes across ripening stages in three pineapple cultivars. Expression was normalized to Actin and calculated by the 2−ΔΔCt method; values are means ± SD (n = 3). HXS, ‘Hong Xiangshui’; XS, ‘Xiangshui’; WD, ‘Victoria’; MG, ‘Mango’; HM, ‘Hongmi’. Ripening stages: Green (immature, green peel), Middle (partially ripe), Ripe (fully ripe). Different lowercase letters indicate significant differences among groups (P < 0.05 or P < 0.01).
Discussion
We comprehensively characterized the pineapple AcCXE family by assessing copy number and physicochemical properties, phylogeny, gene structure, conserved motifs, promoter cis-elements, and expression patterns, and we conducted RT-qPCR for preliminary validation. The 20 AcCXE members identified in the genome differ in basic features, with protein lengths of 170–460 amino acids and predicted isoelectric points of 4.7-8.9, indicating potential structural and functional divergence. Subcellular localization predictions placed most CXEs in the cytosol, with a minority in chloroplasts, the endoplasmic reticulum, and the nucleus, suggesting a predominant role in cytosolic ester hydrolysis.
Phylogenetically, pineapple CXEs clustered with CXEs from model plants (for example, Arabidopsis thaliana, Solanum lycopersicum, and strawberry) and fruit trees (for example, Nanguo pear) into five clades, supporting broad evolutionary conservation together with lineage-specific expansions in certain groups (for example, Group IV) (Goulet et al., 2012; Rui et al., 2022; Zhang et al., 2024a). Several AcCXEs grouped with genes previously implicated in fruit-aroma regulation: AcCXE4 with PuCXE15, AcCXE7 with tomato SlCXE1, and AcCXE3 with apple MdCXE1. Given that PuCXE15 promotes ester degradation in Nanguo pear (Qi et al., 2023) and that MdCXE1 and SlCXE1 mediate ester catabolism in apple and tomato (Souleyre et al., 2011; Goulet et al., 2012), the corresponding pineapple genes likely participate in volatile ester turnover during pineapple aroma formation.
In gene structure and motif analyses, AcCXEs showed an overall conserved organization, which helps resolve relationships among family members (Babenko et al., 2004; Roy and Penny, 2007). Exon number and length varied among genes. Such shifts, often coupled with exon count and length, are characteristic of gene structural evolution (Zhu et al., 2009). Domain analysis indicated that most AcCXEs contain the conserved carboxylesterase domain. AcCXE13 lacks Motif 4 and Motif 8, both of which are conserved in most AcCXE members. Motif 4 is generally located near the catalytic serine residue and is considered critical for forming the catalytic pocket and recognizing ester substrates, whereas Motif 8 contributes to maintaining the stability of the α/β-hydrolase fold. The absence of these motifs in AcCXE13 may therefore lead to structural alteration or loss of catalytic efficiency, implying potential functional divergence from other AcCXE proteins. Motif variation may underlie functional divergence, and structural diversity likely confers substrate specificity and regulatory flexibility within the AcCXE family (Su et al., 2020).
AcCXEs were distributed across all chromosomes, with clustered arrays in several regions. Tandem and segmental duplications are likely the principal drivers of family expansion (Die et al., 2018). Comparative synteny with tomato, rice, Arabidopsis, and banana revealed 14 and 15 syntenic pairs with the monocots, banana and rice, and 6 and 2 pairs with the dicots tomato and Arabidopsis. This pattern indicates stronger conservation of CXE loci in monocots and suggests functional diversification after the monocot-dicot divergence. Building on established findings for fruit aroma in banana and tomato, these collinear relationships support roles for pineapple CXEs in ester metabolism and aroma formation.
Prediction of cis-acting elements indicated that AcCXEs may respond to light, hormones, abiotic stress, and developmental cues. Light-responsive elements were most abundant. Box4 was most frequent in AcCXE10 and G-box was most frequent in AcCXE9, suggesting roles in light signaling and possible involvement in ester metabolism (Ain-Ali et al., 2021). Among hormone-related elements, jasmonate-responsive motifs were most enriched, indicating participation of the AcCXE family in the JA pathway, which is important for fruit ripening and defense responses (Wasternack and Hause, 2013). Stress-associated elements such as ARE, MBS, and LTR were enriched, suggesting that some AcCXEs respond to low temperature, drought, and anaerobic stress. Furthermore, the distribution of hormone-related elements may help explain the differential expression of certain AcCXEs genes among pineapple varieties. For instance, AcCXE4 and AcCXE7 showed higher expression in the low-aroma variety ‘HongXiangshui’, which may be associated with the abundance of ABA-responsive elements (ABRE) in their promoter regions. These elements might be activated during the late maturation stage of this variety, thereby enhancing gene expression and accelerating ester degradation, ultimately contributing to its weaker aroma intensity.
Integrating RNA-seq and qPCR, we systematically screened AcCXE genes potentially associated with aroma intensity. At the transcript level, AcCXE3, AcCXE4, AcCXE5, AcCXE7, AcCXE9, and AcCXE13 showed expression patterns consistent with the negative regulation of ester accumulation by CXEs. qPCR across cultivars showed that AcCXE4 and AcCXE7 were expressed at significantly higher levels at ripening in the light-aroma cultivar ‘HongXiangshui’ than in the sweet-aroma cultivars. Their expression decreased with ripening in ‘Xiangshui’ but increased in ‘HongXiangshui’, which may underlie cultivar differences in aroma content. The expression of PuCXE15, which clustered together with AcCXE4, is highly negatively correlated with the contents of key esters, and its overexpression or silencing, respectively, leads to a decrease or increase in ester content, suggesting that this gene plays a direct role in aroma regulation. We therefore infer that AcCXE4 and AcCXE7 modulate ester metabolism to influence volatile-aroma production, with cultivar- and stage-dependent effects.
In summary, the pineapple AcCXE family constitutes a conserved, multifunctional regulatory network shaped by long-term gene duplication and functional divergence. AcCXEs likely participate in ester turnover, modulation of fruit aroma, regulation of stress responses, and control of development and growth. These findings provide a foundation for functional studies and molecular improvement. Future work can resolve AcCXE-mediated regulation of aroma biosynthesis and stress responses at the molecular and metabolomic levels.
Conclusion
We systematically identified and analyzed 20 CXE genes in pineapple. Comprehensive assessments of physicochemical properties, phylogeny, gene structure, conserved motifs, chromosomal distribution, promoter cis-elements, expression profiles, and RT-qPCR highlighted AcCXE4 and AcCXE7, whose expression patterns were negatively correlated with aroma formation, consistent with CXE-mediated ester catabolism. We infer that these genes likely mediate degradation of volatile esters and thereby shape pineapple aroma, making them priority targets for future aroma research and breeding.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Author contributions
JF: Writing – review & editing. WX: Writing – review & editing, Writing – original draft, Visualization, Data curation, Methodology. TL: Writing – original draft, Data curation, Writing – review & editing. JW: Writing – review & editing, Validation. YW: Writing – review & editing. JH: Writing – review & editing, Funding acquisition, Resources, Project administration. AL: Funding acquisition, Resources, Writing – review & editing, Supervision. CC: Funding acquisition, Supervision, Writing – review & editing, Conceptualization, Writing – original draft. WM: Supervision, Data curation, Visualization, Conceptualization, Writing – review & editing. OC: Writing – review & editing. SH: Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by the National Key Research and Development Program of China (No. 2024YFE0214700), the Project of State Key Laboratory of Tropical Crop Breeding (SKLTCBZRJJ202502) and the National Natural Science Foundation of China (32572983 and 32502660). CC and AL were supported by the Central Public-interest Scientific Institution Basal Research Fund for the Chinese Academy of Tropical Agricultural Sciences (1630032024026, 1630032025001 and 1630012025707). WM were supported by 2025 Science and Technology Project “Hainan Qiongshan Litchi Science and Technology Backyards” supported by Hainan State Farms Investment Holding Group Co., Ltd.
The funder was not involved in the study design, collection, analysis, or interpretation of data, the writing of this article, or the decision to submit it for publication.
Acknowledgments
We thank Ning Liao, Guohui Lv, Runan Wang and Guanglai Zhu for their general help in sample collection. We extend our appreciation to Guiyang Watchbio Co., Ltd.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ain-Ali, Q. U., Mushtaq, N., Amir, R., Gul, A., Tahir, M., and Munir, F. (2021). Genome-wide promoter analysis, homology modeling and protein interaction network of Dehydration Responsive Element Binding (DREB) gene family in Solanum tuberosum. PloS One 16, e0261215. doi: 10.1371/journal.pone.0261215
Ali, M. M., Hashim, N., Abd Aziz, S., and Lasekan, O. (2020). Pineapple (Ananas comosus): a comprehensive review of nutritional values, volatile compounds, health benefits, and potential food products. Food Res. Int. 137, 109675. doi: 10.1016/j.foodres.2020.109675
Babenko, V. N., Rogozin, I. B., Mekhedov, S. L., and Koonin, E. V. (2004). Prevalence of intron gain over intron loss in the evolution of paralogous gene families. Nucleic Acids Res. 32, 3724–3733. doi: 10.1093/nar/gkh686
Bailey, T. L., Boden, M., Buske, F. A., Frith, M., Grant, C. E., Clementi, L., et al. (2009). MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 37 , W202–W208. doi: 10.1093/nar/gkp335
Bray, N. L., Pimentel, H., Melsted, P., and Pachter, L. (2016). Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 34, 525–527. doi: 10.1038/nbt.3519
Cao, X., Duan, W., Wei, C., Chen, K., Grierson, D., and Zhang, B. (2019a). Genome-wide identification and functional analysis of carboxylesterase and methylesterase gene families in Peach (Prunus persica L. Batsch). Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01511
Cao, X., Xie, K., Duan, W., Zhu, Y., Liu, M., Chen, K., et al. (2019b). Peach carboxylesterase PpCXE1 is associated with catabolism of volatile esters. J. Agric. Food Chem. 67, 5189–5196. doi: 10.1021/acs.jafc.9b01166
Chen, C., Wu, Y., Li, J., Wang, X., Zeng, Z., Xu, J., et al. (2023). TBtools-II: A “one for all, all for one. Bioinf. platform Biol. big-data Min. Mol. Plant 16, 1733–1742. doi: 10.1016/j.molp.2023.09.010
Coelho, E. F., Lima, L. W. F., Stringam, B., de Matos, A. P., Santos, D. L., Reinhardt, D. H., et al. (2024). Water productivity in pineapple (Ananas comosus) cultivation using plastic film to reduce evaporation and percolation. Agric. Water Manage. 296, 108785. doi: 10.1016/j.agwat.2024.108785
Die, J. V., Castro, P., Millán, T., and Gil, J. (2018). Segmental and tandem duplications driving the recent NBS-LRR gene expansion in the asparagus genome. Genes 9, 568. doi: 10.3390/genes9120568
Feng, J., Zhang, W., Chen, C., Liang, Y., Li, T., Wu, Y., et al. (2024). The pineapple reference genome: Telomere-to-telomere assembly, manually curated annotation, and comparative analysis. J. Integr. Plant Biol. 66, 2208–2225. doi: 10.1111/jipb.13748
George, J., Nguyen, T., Sanewski, G., Hardner, C., and Smyth, H. E. (2024). Stable isotope dilution assay and HS-SPME-GCMS quantification of key aroma volatiles of Australian pineapple (Ananas comosus) cultivars. Food Chem. 455, 139956. doi: 10.1016/j.foodchem.2024.139956
George, J., Nguyen, T., Williams, D., Hardner, C., Sanewski, G., and Smyth, H. E. (2023). Review of the aroma chemistry of pineapple (Ananas comosus). J. Agric. Food Chem. 71, 4069–4082. doi: 10.1021/acs.jafc.2c08546
Goulet, C., Mageroy, M. H., Lam, N. B., Floystad, A., Tieman, D. M., and Klee, H. J. (2012). Role of an esterase in flavor volatile variation within the tomato clade. Proc. Natl. Acad. Sci. U.S.A. 109, 19009–19014. doi: 10.1073/pnas.1216515109
Hu, Z., Chen, M., Zhu, K., Liu, Y., Wen, H., Kong, J., et al. (2024). Multiomics integrated with sensory evaluations to identify characteristic aromas and key genes in a novel brown navel orange (Citrus sinensis). Food Chem. 444, 138613. doi: 10.1016/j.foodchem.2024.138613
Kim, K. K., Song, H. K., Shin, D. H., Hwang, K. Y., Choe, S., Yoo, O. J., et al. (1997). Crystal structure of carboxylesterase from Pseudomonas fluorescens, an alpha/beta hydrolase with broad substrate specificity. Structure 5, 1571–1584. doi: 10.1016/s0969-2126(97)00306-7
Larrea-Sarmiento, A. E., Olmedo-Velarde, A., Wang, X., Borth, W., Domingo, R., Matsumoto, T. K., et al. (2022). Genetic diversity of viral populations associated with Ananas germplasm and improvement of virus diagnostic protocols. Pathogens 11, 1470. doi: 10.3390/pathogens11121470
Lescot, M., Déhais, P., Thijs, G., Marchal, K., Moreau, Y., Van de Peer, Y., et al. (2002). PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 30, 325–327. doi: 10.1093/nar/30.1.325
Mao, Q., Chen, C., Xie, T., Luan, A., Liu, C., and He, Y. (2018). Comprehensive tissue-specific transcriptome profiling of pineapple (Ananas comosus) and building an eFP-browser for further study. PeerJ 6, e6028. doi: 10.7717/peerj.6028
Marchler-Bauer, A. and Bryant, S. H. (2004). CD-Search: protein domain annotations on the fly. Nucleic Acids Res. 32, W327–W331. doi: 10.1093/nar/gkh454
Marshall, S. D., Putterill, J. J., Plummer, K. M., and Newcomb, R. D. (2003). The carboxylesterase gene family from Arabidopsis thaliana. J. Mol. Evol. 57, 487–500. doi: 10.1007/s00239-003-2492-8
Martínez-Rivas, F. J., Blanco-Portales, R., Moyano, E., Alseekh, S., Caballero, J. L., Schwab, W., et al. (2022). Strawberry fruit FanCXE1 carboxylesterase is involved in the catabolism of volatile esters during the ripening process. Hortic. Res. 9, uhac095. doi: 10.1093/hr/uhac095
Montero-Calderón, M., Rojas-Graü, M. A., and Martín-Belloso, O. (2010). Aroma profile and volatiles odor activity along gold cultivar pineapple flesh. J. Food Sci. 75, S506–S512. doi: 10.1111/j.1750-3841.2010.01831.x
Qi, L., Li, X., Zang, N., Zhang, Z., Yang, Y., Du, Y., et al. (2023). Genome-wide identification of CXE and PuCXE15 functions in the catabolism of volatile ester in ‘Nanguo’ pear fruit. Plant Physiol. Biochem.PPB 203, 107996. doi: 10.1016/j.plaphy.2023.107996
Rao, X., Huang, X., Zhou, Z., and Lin, X. (2013). An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinforma. Biomath. 3, 71–85.
Roy, S. W. and Penny, D. (2007). Patterns of intron loss and gain in plants: intron loss-dominated evolution and genome-wide comparison of O. sativa and A. thaliana. Mol. Biol. Evol. 24, 171–181. doi: 10.1093/molbev/msl159
Rui, C., Peng, F., Fan, Y., Zhang, Y., Zhang, Z., Xu, N., et al. (2022). Genome-wide expression analysis of carboxylesterase (CXE) gene family implies GBCXE49 functional responding to alkaline stress in cotton. BMC Plant Biol. 22, 194. doi: 10.1186/s12870-022-03579-9
Souleyre, E. J., Marshall, S. D., Oakeshott, J. G., Russell, R. J., Plummer, K. M., and Newcomb, R. D. (2011). Biochemical characterisation of MdCXE1, a carboxylesterase from apple that is expressed during fruit ripening. Phytochemistry 72, 564–571. doi: 10.1016/j.phytochem.2011.01.020
Steingass, C. B., Langen, J., Carle, R., and Schmarr, H. G. (2015). Authentication of pineapple (Ananas comosus [L.] Merr.) fruit maturity stages by quantitative analysis of γ- and δ-lactones using headspace solid-phase microextraction and chirospecific gas chromatography-selected ion monitoring mass spectrometry (HS-SPME-GC-SIM-MS). Food Chem. 168, 496–503. doi: 10.1016/j.foodchem.2014.07.071
Su, H. G., Zhang, X. H., Wang, T. T., Wei, W. L., Wang, Y. X., Chen, J., et al. (2020). Genome-wide identification, evolution, and expression of GDSL-Type esterase/lipase gene family in soybean. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00726
Wang, Y., Zhou, Z., Liu, Y., Pang, R., Zhao, X., Chen, Q., et al. (2025). Investigating the aroma transition during rapid mango ripening driven by dynamic catabolism of fatty acids. Food Chem. 491, 145326. doi: 10.1016/j.foodchem.2025.145326
Wasternack, C. and Hause, B. (2013). Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. Ann. Bot. 111, 1021–1058. doi: 10.1093/aob/mct067
Yi, W., Luan, A., Liu, C., Wu, J., Zhang, W., Zhong, Z., et al. (2023). Genome-wide identification, phylogeny, and expression analysis of GRF transcription factors in pineapple (Ananas comosus). Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1159223
Zhang, J., Wang, X., Dou, G., Meng, D., Tang, C., Lv, J., et al. (2024a). Genome-wide identification of rice CXE gene family and mining of alleles for potential application in rice improvement. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1435420
Zhang, L., Zhou, K., Wang, M., Li, R., Dai, X., Liu, Y., et al. (2022). The functional characterization of carboxylesterases involved in the degradation of volatile esters produced in strawberry fruits. Int. J. Mol. Sci. 24, 383. doi: 10.3390/ijms24010383
Zhang, W., Wu, J., He, J., Liu, C., Yi, W., Xie, J., et al. (2024b). AcMYB266, a key regulator of the red coloration in pineapple peel: a case of subfunctionalization in tandem duplicated genes. Hortic. Res. 11, uhae116. doi: 10.1093/hr/uhae116
Keywords: pineapple, CXE gene family, carboxylesterase, esters, aroma
Citation: Xu W, Li T, Wu J, Wang Y, Feng J, Luan A, Carzorel OPP, He S, He J, Chen C and Ma W (2025) Characterization of CXE genes in pineapple and their aroma-related expression during fruit ripening. Front. Plant Sci. 16:1733743. doi: 10.3389/fpls.2025.1733743
Received: 28 October 2025; Accepted: 17 November 2025; Revised: 10 November 2025;
Published: 08 December 2025.
Edited by:
Xitong Fei, Northwest A and F University, ChinaReviewed by:
Yunpeng Cao, Chinese Academy of Sciences (CAS), ChinaKangyu Wang, Jilin Agriculture University, China
Copyright © 2025 Xu, Li, Wu, Wang, Feng, Luan, Carzorel, He, He, Chen and Ma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wuqiang Ma, d3VxaWFuZ21hQGhhaW5hbnUuZWR1LmNu; Chengjie Chen, Y2NqQGNhdGFzLmNu; Junhu He, aGVqdW5odUBjYXRhcy5jbg==
†These authors have contributed equally to this work