

- Département des Sciences Fondamentales, Université du Québec à Chicoutimi, Chicoutimi, QC, Canada
In the short term, trees rely on the internal storage of water because it affects their ability to sustain photosynthesis and growth. However, water is not rapidly available for transpiration from all the compartments of the plant and the living tissues of the stem act as a buffer to preclude low water potentials during peaks of transpiration. In this paper, electronic dendrometers were used from mid-June to mid-September 2008 to compare the radius variations in stem and roots of black spruce [Picea mariana (Mill.) B.S.P.] in two sites of the boreal forest of Quebec, Canada, with different soil characteristics and water retention. The duration of the daily cycles was similar between sites and measurement heights but greater amplitudes of contraction and expansion were observed on the stem and in the site with the shallowest soil organic layer. The expansion phase had higher amplitudes and lasted longer than contraction. On average, the contraction phase occurred between 07:00 and 16:30 (legal time), while expansion lasted 14.5 h. The roots in the site with the deepest organic layer showed a wider variation in the onset of contraction, which could be as late as 13:00. The probability of observing the contraction phase depended on precipitation. With a precipitation <0.5 mm h−1, the bivariate posterior probabilities estimated >60% probability of observing contraction between 05:00 and 21:00, decreasing to 20% with precipitation >1.1 mm h−1. These findings demonstrated that the depth of the organic layer plays an important role in maintaining the internal water reserve of trees. The dynamics of water depletion and replenishment can modify the water potential of xylem and cell turgor during the enlargement phase, thus affecting radial growth. Changes in temperature and precipitation regime could influence the dynamics of internal water storage in trees growing on shallower and drier soils.
Introduction
In the short term, trees rely on internal storage to constantly fulfill their water needs because there may be a significant delay between water loss from leaves and water uptake by roots (Cermak et al., 2007). Transpiration, mainly during warm and sunny days, induces sudden and wide changes in water potential that propagate within the tree, initiating the water flow and a wave of water depletion at increasing distances from the crown (Chaves et al., 2003). Consequently, the water storage capacity in trees is of considerable biological interest because it influences the ability to sustain photosynthesis and growth, not only during periods of drought (Cruiziat et al., 2002; Cermak et al., 2007; Giovannelli et al., 2007; Rossi et al., 2009), but whenever water transport occurs within the xylem conduits.
Trees can store water in three different compartments: (i) in sapwood, (ii) in the cell walls or inactive vessels, and (iii) within the living cells of leaves, bark, and wood rays (Zimmermann, 1983). However, water is not necessarily directly available for transpiration from all these compartments. If water is rapidly withdrawn from the dead cells of xylem, cavitation may occur because of the rigidity of this tissue (Jarvis, 1975). Instead, the water stored in the living cells of bark can more efficiently contribute to the daily transpiration stream because of the elasticity of the tissue and their high connectivity with xylem (Herzog et al., 1995; Steppe et al., 2005). The water released from the bark thus acts as a buffer to preclude low water potentials in xylem as a result of transpiration peaks. Therefore, there is evidence that the living cells of the bark parenchyma, phloem, and cambium are among the most important compartments for water storage in trees (Lassoie, 1973; Waring et al., 1979; Steppe et al., 2005).
When dehydrating and rehydrating, living cells shrink and swell. Because of these elastic properties and following the transpiration from leaves and water uptake from roots, the daily rhythms of depletion and replenishment of water storage in the outermost tissues of the stem cause quantifiable variations in its size, which can be measured with automatic dendrometers (Deslauriers et al., 2003; Giovannelli et al., 2007). As a general rule, stem and roots contract in the daytime, when the transpiration and photosynthesis processes are vigorous, and expand at night, when water reserves are gradually replenished. Most variations in size occur in the elastic bark tissues outside the cambium, whereas xylem undergoes only small daily variations (Irvine and Grace, 1997). Moreover, under the natural ranges of water potential currently reached in the stem, changes in radius are directly proportional to the water content in bark (Hergoz et al., 1995; Zweifel and Häsler, 2001; Zweifel et al., 2001; Cermak et al., 2007). Automatic measurements of radius variations in the stem can therefore provide an effective and sensitive proxy for the water status of trees.
Historical climate analyses and current projections indicate that the climate is changing at a rate never previously experienced. In the past century, mean annual temperature over southern regions of Canada has increased by an average of 0.9°C (Zhang et al., 2000). This temperature increase affects the hydrological cycle by increasing evapotranspiration of plants and leads to a modification of the amount and distribution of precipitation. Consistent with the global trend observed at high latitudes (Easterling et al., 2000), Canada as a whole is experiencing increased annual precipitation, but mainly in the form of snowfall (Zhang et al., 2001; Motha and Baier, 2005), while extreme conditions, such as dry periods, are occurring more frequently during summer (Zhang et al., 2000). There is general agreement that the increase in precipitation will be unable to balance the increase in evaporation induced by the higher air temperatures, causing declines in soil moisture as well as periods of intense drought (Easterling et al., 2000; Motha and Baier, 2005). The cold temperatures occurring in the boreal forest keep evaporation to a minimum, and the soils are often wet. These cold environments with high ground water levels are optimal conditions for the growth of black spruce [Picea mariana (Mill.) B.S.P.]. However, because of its shallow root system (Polomski and Kuhn, 1998), black spruce may be dramatically dependent on precipitations, so modifications to the temperature and rainfall distribution could significantly affect growth and survival of the individuals growing on shallower soils with reduced water availability.
In conifers, the daily rhythms of water depletion and replenishment are determined by several factors such as soil water content, atmospheric vapor pressure deficit (VPD), and rainfall (Zweifel et al., 2005; Deslauriers et al., 2007). During the growing season, precipitation events <1 mm can be quickly followed by a marked stem swelling (Braekke and Kozlowski, 1975; Zweifel et al., 2005). Therefore, even minor variations in the precipitation regime may have an important impact on water storage replenishment in trees. In the boreal forest, the consequences of a warming on the dynamics of growth have recently been estimated (Rossi et al., 2011), but the influence of a changing precipitation regime on the water reserves of trees is unknown because the relationships between plants and water in the boreal forest have been less studied and a number of factors play a role in the responses of trees to water stress. Developing a clear understanding of the way black spruce reacts to specific climatic and weather conditions could provide information for predicting the responses of the boreal forest to future global changes. We therefore monitored radius variation in black spruce in continuum using stem and root dendrometers. Specifically, the aims of the study were (1) to investigate the amplitude and duration of the daily cycle of radius variation (2) to compare the responses to precipitation events in two sites with different soil characteristics in the boreal forest of Quebec, Canada.
Materials and Methods
Study Sites
The study was carried out in two black spruce stands in the boreal forest of Quebec, Canada. The first site, BER, is located at 611 masl in the Monts Valins (48°51′N, 70°20′W). The second site, SIM, is in the Parc des Laurentides at 350 masl (48°12′N, 71°14′W). Both sites are typical, even-aged and mature stands established after a forest fire, with black spruce being the dominant species. The climate is continental, with long cold winters and short warm summers. BER is colder, with a higher soil water content and snow depth than in SIM, although the amount of rainfall is similar in the two sites (Table 1). Soils of both stands are classified as podzols formed on post-glacial till deposits, but the depth of the organic layer in BER ranges between 20 and 40 cm, while that in SIM is only 10–20 cm. The maximum rooting depth in SIM is thus restricted by the bedrock and adventitious roots appear to be disposed almost horizontally on the top of the soil. By contrast, rooting is deeper in BER, with the adventitious roots plunging into the soil at an angle of c.a. 60°.
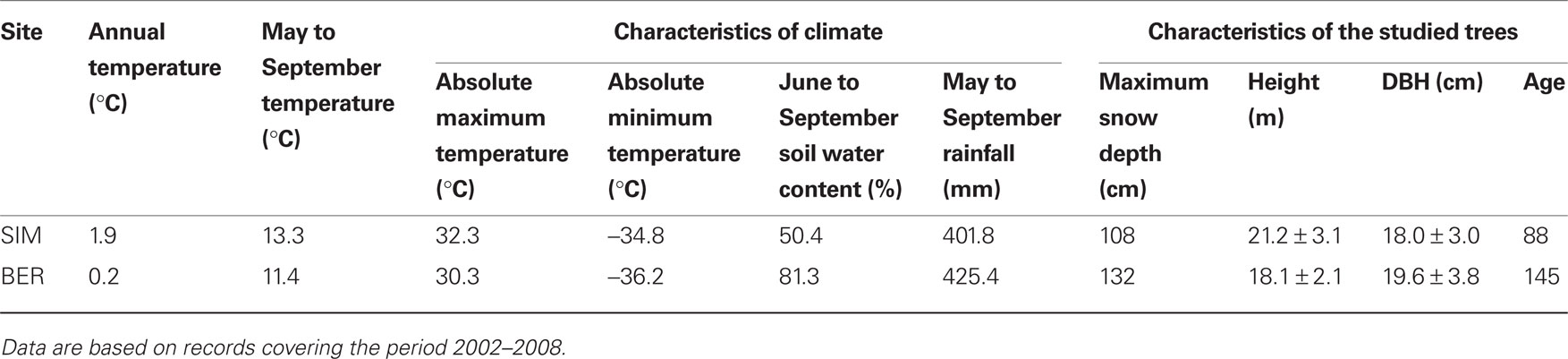
Table 1. Characteristics of climate and studied trees [height, diameter at breast height (DBH), and age] of two black spruce stands in the boreal forest of Quebec, Canada.
Data Collection
Six trees were sampled at each study site among those with upright stem and growth patterns similar to the average of the stand (Table 1). The homogeneity in growth rates was assessed during a preliminary investigation by extracting wood cores and counting the number of tracheids in three previous tree rings (Lupi et al., 2010). The study was conducted from mid-June to mid-September 2008. This period corresponded approximately to the period of vigorous growth observed in this species and in the same sites (Rossi et al., 2011).
Electronic point dendrometers (Agricultural Electronics Corp., Tucson, AZ, USA) were used to measure tree radius variations in continuum (Figure 1). All trees were equipped with a point dendrometer installed at breast height. A further nine dendrometers, six in SIM and three in BER, were installed at 20 cm from the stump along the adventitious roots. More root dendrometers were available at SIM because some trees were being monitored for another ongoing study. The sensors for the point dendrometers were linear variable differential transducers (LVDT) enclosed in an aluminum housing and fixed to the tree with stainless steel rods having a thermal linear expansion coefficient of 17 μm m−1 C−1 implanted 6–7 cm deep in the xylem. With this equipment, the percentage of metal expansion was less than 1% of stem variation. A sensing rod held against the surface of the bark measured the radius variations, which in our monitoring represented the overall variation in size of xylem and phloem together. The dendrometer sensitivity to temperature and humidity was negligible due to the use of dimensionally stable compounds in their manufacture. The dead bark was partially removed to minimize error due to hygroscopic thickness variations. Data were automatically collected every 15 min and hourly means were calculated and stored in a CR1000 datalogger (Campbell Scientific, Inc.). Data were recorded in legal time.
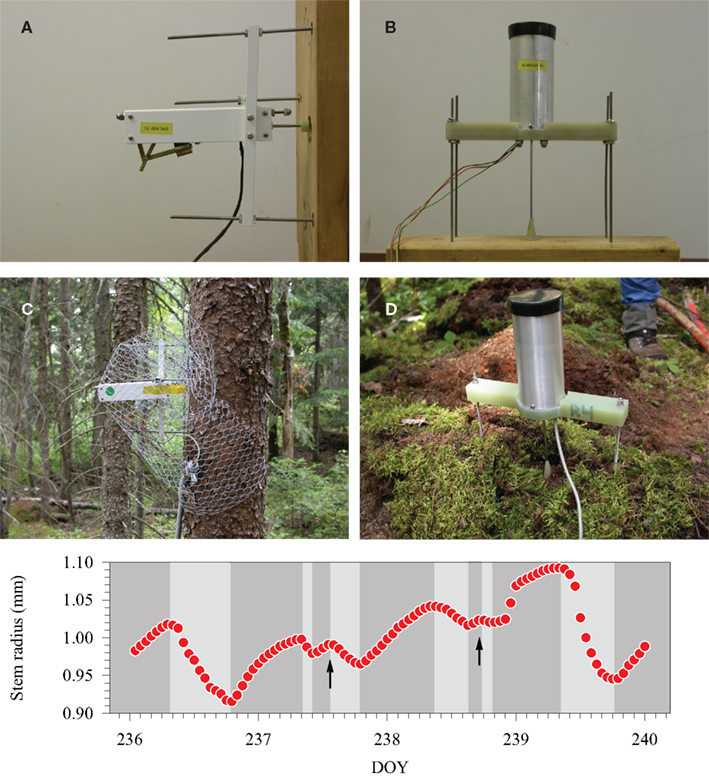
Figure 1. Stem and root dendrometers during assemblage (A,B) and installed on the tree (C,D). Cycles of contraction (light-gray background) and expansion (dark-gray background) of radius variation detected by a high-resolution, automatic dendrometer in black spruce. Arrows indicate examples of variations deviating from the theoretical diurnal pattern. Modified from Deslauriers et al. (2010).
In each site, a weather station was installed in a forest gap close to the studied trees to measure environmental parameters. Air and soil temperature, relative humidity, volumetric soil water content, and rainfall were recorded every 5 min and hourly means or sums were computed and stored in the datalogger. The probe for soil temperature and the time domain reflectometer for soil water content were installed in the middle part of the organic layer.
Extracting Radius Variation
The time-series of root and stem radial variation measured with dendrometers were processed to detect the phases of contraction and expansion according to the stem cycle approach (Downes et al., 1999; Deslauriers et al., 2003) by using the SAS routines provided by Deslauriers et al. (2011). The contraction phase was considered as the period between a maximum and a minimum radius. The expansion phase was the period between a minimum and the next maximum radius (Figure 1). The amplitudes of contraction and expansion were calculated as the difference between maximum and minimum radius, regardless of the time of the day when they occurred. The timings of onset of contraction and expansion were also recorded. Because of the variable weather conditions and rainfall events, deviations from the theoretical circadian pattern described by Hergoz et al. (1995) were observed in all trees within the stand. These small variations were maintained in the dataset to accurately extract all possible information from the time-series (Figure 1).
Statistical Analyses
The mean duration and amplitude were compared between sites and measurement heights (stem and root) and the interaction between the two factors was tested with analysis of variance (ANOVA) using the GLM procedure in the SAS statistical package (SAS Institute, Cary, NC, USA). Statistical comparisons between the two factors were performed by repetitively and randomly resampling the original dataset to calculate the average F-statistic and related standard deviations of the distribution (Efron, 1979). A 1000-times-bootstrapping was performed in order to improve the robustness of results. The tests were considered non-significant (P > 0.05) when the error bars crossed 0 (Chernick, 2008).
Discriminant analysis was performed to calculate the probability of observing the contraction according to the hour of the day and rainfall. To calculate this probability, contraction was coded in a binary response as absent (value 0) or present (value 1) for a given hour of the day. The resulting phase-specific densities were used to calculate bivariate posterior probability distributions for each hour of the day (Hora and Wilcox, 1982). The classification criterion adopted parametric methods based on multivariate normal distributions within each class to derive discriminant quadratic functions, and assumed unequal variances between classes using the observed within-group covariance matrices (DISCRIM procedure in SAS; SAS Institute, Cary, NC, USA). Estimates were calculated only for values within the ranges of variation of the measurements.
Results
Weather Patterns
The daily pattern of the air temperature was similar in the two sites, with the coldest temperatures observed at 05:00. On average, air temperature reached 17–18°C between 12:00 and 16:00. In SIM, soil temperature closely followed air temperature, as a consequence of the shallow organic layer. In contrast, soil temperature in BER was constant, with a maximum range measured during the growing period of less than 2°C. During the day, relative humidity was similar in the two sites, although lower nighttime values were observed in SIM. As a consequence, VPD was always lower in BER, especially at night. Soil water content ranged between 60 and 80% until mid-July, then gradually decreased, with the more abrupt reductions observed in SIM that reached 10% at the beginning of September (Figure 2). Similar rainfall amounts of up to 54.1 mm were detected in the two sites (Figure 3). Rainfall events occurred frequently, with 69 and 67 rainy days observed during the study period in BER and SIM, respectively.
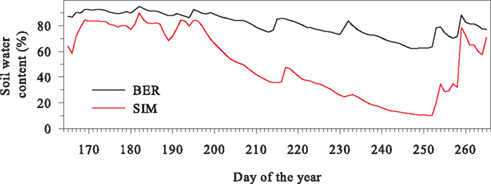
Figure 2. Soil water content measured in the two black spruce stands of the boreal forest of Quebec, Canada.
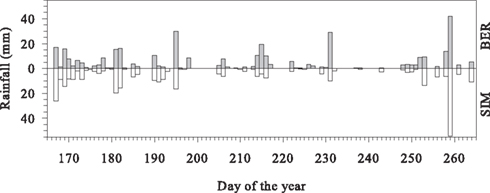
Figure 3. Daily rainfall measured in the two black spruce stands of the boreal forest of Quebec, Canada.
Comparisons between Sites and Measurement Heights
Duration and amplitude of contraction and expansion showed frequency distributions positively skewed or distinctly exponential, with most data concentrated in the lower classes except for the duration of expansion (Figure 4). The duration of contraction was similar between sites and between stem and roots, with values between 2 and 10 h being the most frequent. In contrast, ANOVA detected significant differences between sites and measurement heights (stem vs. roots) in the amplitude of contraction (ANOVA, bootstrapped F-values, P > 0.05). Greater amplitudes were observed in SIM and on the stem. The duration of expansion commonly ranged between 12 and 18 h and did not differ significantly either between sites or between measurement heights. Significant differences were observed in the amplitude of expansion, with the higher values calculated in SIM and on the stem (ANOVA, bootstrapped F-statistic, P < 0.05). Compared to contraction, the expansion phase had slightly higher amplitudes and lasted substantially longer (Figure 4). The interactions between sites and measurement heights for duration and amplitude were not statistically significant (bootstrapped F-statistic, P > 0.05), which means that the patterns of duration and amplitude were similar between sites and measurement heights when data from stem and roots or from SIM and BER were pooled.
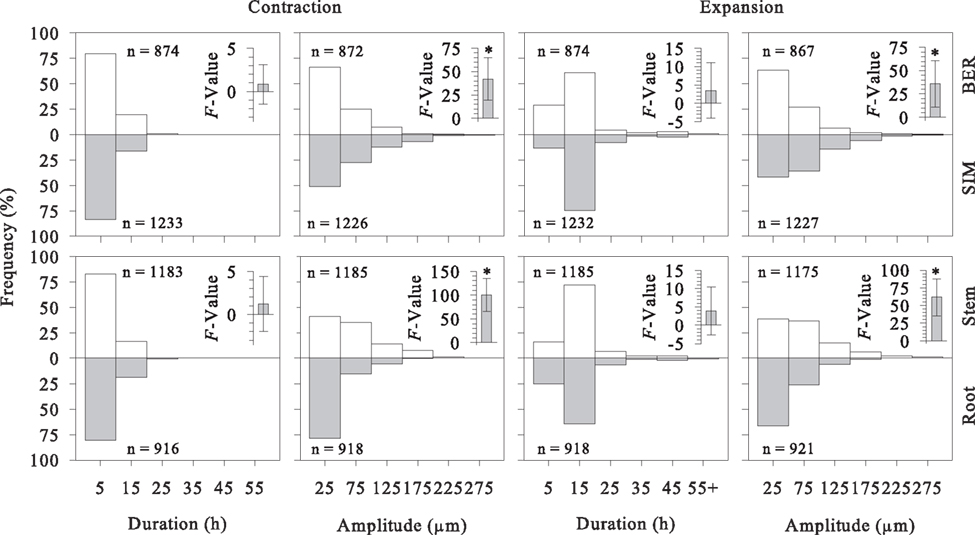
Figure 4. Frequency distribution of duration and amplitude of contraction and expansion in stem and roots of black spruce in two stands of the boreal forest of Quebec, Canada. The enclosed vertical bars represent mean and twice standard deviation of the bootstrapped F-statistic calculated by 1000 replications. Asterisks indicate significant differences (P < 0.05), while differences are considered non-significant (P > 0.05) when the error bars crossed 0.
Timings of Contraction and Expansion
In BER, the onset of contraction and expansion presented a unimodal distribution in the stem, while the distribution showed an evident bimodal pattern in roots (Figure 5). The contraction clearly started at 07:00 in the stem, while large variations were observed in roots, with data concentrated at 09:00 and between 12:00 and 14:00. In SIM, the frequency distributions calculated for the onset of contraction was unimodal for both stem and root with the maximum of occurrence at 07:00. Therefore, only the onset of contraction for roots in BER differs from the other measurement height and site. The expansion phase began in late afternoon in both stem and roots and in both sites. In roots, the expansion phase started at 16:00 in BER. A wider range of variation was observed in SIM, with the onset of expansion occurring between 15:00 and 17:00. The onset of stem expansion occurred between 16:00 and 18:00 in BER and between 15:00 and 17:00 in SIM. As a result, the start of the expansion phase was observed 1 h earlier at SIM, for both stem and roots.
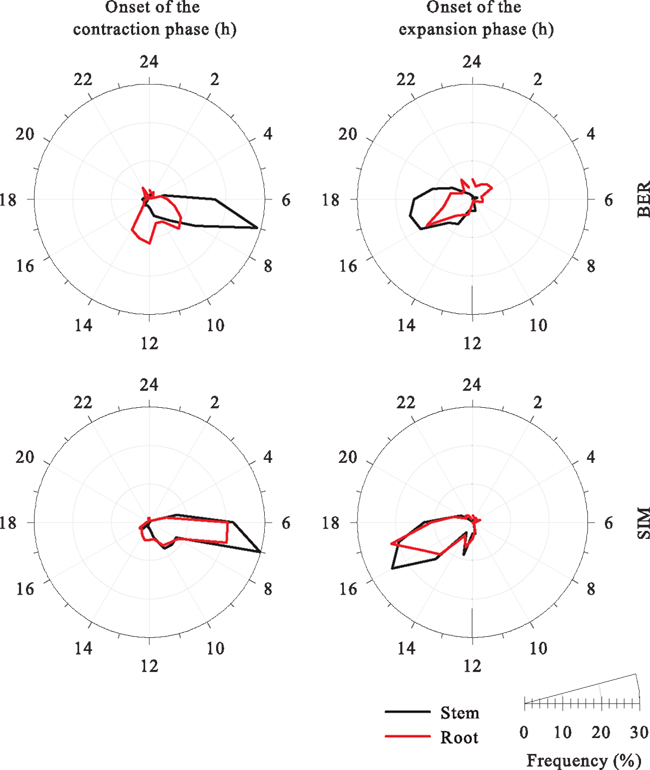
Figure 5. Circular frequency distribution of onset of the contraction and expansion phases in stem and roots of black spruce in two stands of the boreal forest of Quebec, Canada. Hours are reported as legal time.
Posterior Probabilities of Contraction
Although most contractions started between 07:00 and 14:00 (Figure 5), duration of contraction (calculated as the time between onset of contraction and onset of expansion) lasted much longer. The probability of observing contraction at any time depended on the occurrence of precipitation events (Figure 6). According to the bivariate posterior probability, contractions had 60–80% probability of being observed between 05:00 and 21:00, with a precipitation <0.5 mm h−1. Stem contraction had 80–100% probability of occurring between 11:00 and 15:00 in BER, with a precipitation <0.2 mm h−1. The probability of observing root contraction in BER depended less on the hour of the day or amount of precipitation, since the distribution pattern was more dispersed. By contrast, root contraction in SIM occurred during specific hours and was affected by precipitation, since 60–80% probability was located between 04:00 and 22:00 with a precipitation <0.4 mm h−1 (Figure 6). Also, stem contractions in the same site had >60% probability of being observed between 05:00 and 20:00 with the same amount of precipitation. Overall, the probability of observing radius contraction decreased at increasing precipitation and <20% probability was estimated with rainfall >1.1 mm h−1 (Figure 6).
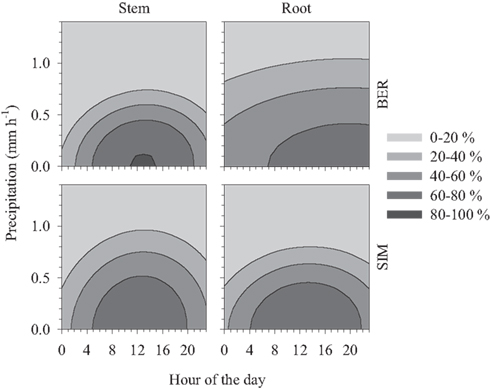
Figure 6. Bivariate posterior probabilities for classifying a radius variation as contraction estimated by discriminant analyses performed between the hour of the day (legal time) and the amount of precipitation in two black spruce stands of the boreal forest of Quebec, Canada.
Discussion
The higher daily variations in radius observed in stems than in roots revealed the internal water use of black spruce. During transpiration, the first sources of water were the compartments closest to water loss (Whitehead and Jarvis, 1981), thus water reserves should be required according to a polar gradient from needles toward the roots. As a consequence, the upper part of the stem is subjected to greater tensions than the belowground part of tree (Cruiziat et al., 2002; Cermak et al., 2007). Similar results were also observed by Zweifel and Häsler (2001) and Cermak et al. (2007). However, there is evidence that the thickness of the tissues is proportional to the amplitudes of the diurnal stem radius variations (Zweifel and Häsler, 2001), and the thicker bark on stems could partially explain the differences in amplitude observed between roots and stem during the study (Figure 4).
The lack of interaction in the amplitude of contraction and expansion between sites and measurement heights suggested that water reserves in roots and stems were used in similar proportions. However, the use of water reserves differed between the two sites. The duration of the daily phases was identical, but the radial variations in both stem and roots had greater amplitudes in SIM, where the higher VPD suggested the occurrence of higher transpiration rates, although the lack of data concerning sap flow prevented a precise quantification of the differences between sites (Deslauriers et al., 2007). During the day, sap flow closely follows the same pattern of variation as VPD, with an abrupt increase occurring early in the morning, followed by a plateau and a reduction in the afternoon (Anfodillo et al., 1998). Deslauriers et al. (2007) demonstrated that both factors have a direct effect on duration and amplitude of the contraction phase. The water deficit experienced during the day and exhibited by greater contraction, was subsequently balanced during the night by one or several equivalent expansion phases. In both sites, the replenishment of water storage in stem and roots was slower than its depletion. Nevertheless, water availability was lower in SIM, especially from mid-July, when soil water content began to diverge substantially from that in BER (Figure 2). The larger radius variations indicated that the water reserves in SIM were more solicited, also demonstrating a less favorable water status of the trees in that site. The use of the water stored in the outermost tissues of the stem makes trees less dependent on the current water availability in the soil. However, although efficient in the short term, this strategy cannot be sustained for long because it could result in severe declines in the water resources of trees.
In both sites, the onset of water replenishment (i.e. the expansion phase) was more dispersed in time compared to the onset of water depletion (i.e. the contraction phase; Figure 5). This reflects the key role of transpiration in driving water use. The high time resolution of our analyses enabled the detection of small radius variations that did not follow the theoretical diurnal pattern of radial variation. Brief rainfall events can initiate a refilling of the water storage anytime during the day (Figure 6). In SIM, the warmer site and with the shallower organic layer, radial variations of stem and roots were synchronous, but this was not the case in BER, where the onset of contraction in roots was observed to occur later than in the stem. In BER, trees are assumed to experience lower transpiration rates associated with better water supply and lower temperatures throughout growing season in respect to SIM. The climatic conditions are key factors in the process of degradation, cycling, and accumulation of organic matter, which led to a greater depth of the organic layer in the soil of BER. A hypothesis is that the requirement for water from roots was markedly lower or even lacking during some days in BER, so the observed radius variations could be mostly related to water replenishment in other tree compartments.
The importance of precipitation events for the water status in black spruce depended on water availability in the soil, which in our experimental design was related to the depth of the organic layer. The time window with higher probability of observing the contraction phase was comparable in the two sites (12 h in BER vs. 11 h in SIM). However, a higher amount of precipitation was needed in SIM to stop contraction and allow tissues to expand. Moreover, during rainy periods, clouds reduce radiation and VPD and so decrease transpiration and the demand for water from the leaves. Consequently, other environmental factors could affect the observed relationships between contraction and precipitation and play an important role in the dynamics of depletion and replenishment of water within trees.
In the studied stands, contraction generally occurred between 06:00 and 17:00, legal time, which is the period between the onset of contraction and that of expansion (Figure 5). In similar environments located in the same region as our sites, balsam fir showed the contraction phases occurring between 08:00 and 17:00, legal time, when the long cycles associated with precipitations were excluded (Deslauriers et al., 2003). In subalpine Norway spruce, Zweifel and Häsler (2001) reported the onset of contraction in the stem at 09:00 during sunny July days, while at the Italian timberline, trees exhibited shorter periods of contraction, commonly between 10:30 and 18:30, solar time (Deslauriers et al., 2007). Compared to these results, the onset of contraction at our latitudes occurred earlier. It is well known that the alternation between tissue contraction and expansion in both stem and roots is largely determined by the course of transpiration, availability of water and saturation of internal storage tissues (Zweifel et al., 2001). However, to observe a marked increase in radius, the final balance between the water exchanges in the different compartments (transpiration, water uptake, and the cyclic depletion and replenishment of water in the outermost tissues of the stem) must result in higher amounts of water stored than released. In stem and roots, this is especially important in the first stages of growth during the year, when water sustains the turgor of the enlarging cells in the developing earlywood (Rossi et al., 2009). Our results demonstrated that, in the boreal forest, precipitation is an important factor in the replenishment of the internal water reserves in trees growing on the shallowest and driest organic layers.
Conclusion
This paper demonstrates that the climatic conditions and the depth of the soil organic layer can play an important role in maintaining the internal water reserves in black spruce. Because of its shallow root system, black spruce relies on water supplied by precipitations to maintain its water status. Future changes in air temperature and precipitation regime modifying water availability could substantially influence the dynamics of water depletion and replenishment occurring in individuals of this species growing in the stands with shallower or drier soils. However, this study was not able to disentangle the effects of the correlated weather variables on stem contraction and expansion, and a more accurate understanding of the role of climate on growth is still required to definitively assess the dynamics of water storage in trees.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was funded by Natural Sciences and Engineering Research Council of Canada (NSERC), Consortium de Recherche sur la Forêt Boréale Commerciale, Consortium OURANOS, and the Ministère des Ressources Naturelles et de la Faune (MRNF). The authors thank F. Gionest, D. Laprise, C. Lupi, G. Savard, and M. Thibeault-Martel for technical support in the field and A. Garside for checking the English text.
References
Anfodillo, T., Rento, S., Carraro, V., Furlanetto, L., Urbinati, C., and Carrer, M. (1998). Tree water relations and climatic variations at the alpine timberline: seasonal changes of sap flux and xylem water potential in Larix decidua Miller, Picea abies (L.) Karst. and Pinus cembra L. Ann. For. Sci. 55, 159–172.
Braekke, F. H., and Kozlowski, T. T. (1975). Shrinkage and swelling of stems of Pinus resinosa and Betula papyrifera in northern Wisconsin. Plant Soil 43, 387–410.
Cermak, J., Kucera, J., Bauerle, W. L., Phillips, N., and Hinckley, T. M. (2007). Tree water storage and its diurnal dynamics related to sap flow and changes in stem volume in old-growth Douglas-fir trees. Tree Physiol. 27, 181–198.
Chaves, M. M., Maroco, J. P., and Pereira, J. S. (2003). Understanding plant responses to drought – from genes to the whole plant. Funct. Plant Biol. 30, 239–264.
Chernick, M. R. (2008). Bootstrap Methods: A Guide for Practitioners and Researchers. Hoboken: Wiley-Interscience.
Cruiziat, P., Cochard, H., and Ameglio, T. (2002). Hydraulic architecture of trees: main concepts and results. Ann. For. Sci. 59, 723–752.
Deslauriers, A., Anfodillo, T., Rossi, S., and Carraro, V. (2007). Using simple causal modeling to understand how water and temperature affect daily stem radial variation in trees. Tree Physiol. 27, 1125–1136.
Deslauriers, A., Morin, H., Urbinati, C., and Carrer, M. (2003). Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada). Trees 17, 477–484.
Deslauriers, A., Rossi, S., Morin, H., and Krause, C. (2010). “Analyse du développement intraannuel des cernes de croissance,” in La dendroécologie: principes, méthodes et applications, eds S. Payette, and L. Filion (Québec: Presses de l’Université Laval), 61–92.
Deslauriers, A., Rossi, S., Turcotte, A., Morin, H., and Krause, C. (2011). A three steps procedure to analyse the time-series of automatic dendrometer data by using SAS program. Dendrochronologia. doi: 10.1016/j.dendro.2011.01.008
Downes, J., Beadle, C., and Worledge, D. (1999). Daily stem growth patterns in irrigated Eucalyptus globulus and E. nitens in relation to climate. Trees 14, 102–111.
Easterling, D. R., Meehl, G. A., Parmesan, C., Changnon, S. A., Karl, T. R., and Mearns, L. O. (2000). Climate extremes: observations, modeling, and impacts. Science 289, 2068–2074.
Efron, B. (1979). 1977 Rietz lecture – bootstrap methods – another look at the jackknife. Ann. Stat. 7, 1–26.
Giovannelli, A., Deslauriers, A., Fragnelli, G., Scaletti, L., Castro, G., Rossi, S., and Crivellaro, A. (2007). Evaluation of drought response of two poplar clones (Populus × canadensis Mönch ‘I-214’ and P. deltoides Marsh. ‘Dvina’) through high resolution analysis of stem growth. J. Exp. Bot. 58, 2673–2683.
Hergoz, K. M., Häsler, R., and Thum, R. (1995). Diurnal changes in the radius of a subalpine Norway spruce stem: their relation to the sap flow and their use to estimate transpiration. Trees 10, 94–101.
Hora, S. C., and Wilcox, J. B. (1982). Estimation of error rates in several population discriminant-analysis. J. Marketing Res. 19, 57–61.
Irvine, J., and Grace, J. (1997). Continuous measurements of water tensions in the xylem of trees based on the elastic properties of wood. Planta 202, 455–461.
Jarvis, P. G. (1975). “Water transfer in plants,” in Heat and Mass Transfer in the Biosphere. Part 1: Transfer Processes in the Plant Environments, eds. D. A. De Vries and N. H. Afgan (New York: John Wiley), 369–394.
Lassoie, J. P. (1973). Diurnal dimensional fluctuations in a Douglas-fir stem in response to tree water status. For. Sci. 19, 251–255.
Lupi, C., Morin, H., Deslauriers, A., and Rossi, S. (2010). Xylem phenology and wood production: resolving the chicken-or-egg dilemma. Plant Cell Environ. 33, 1721–1730.
Motha, R. P., and Baier, W. (2005). Impacts of present and future climate variability on agriculture in the temperate regions: North America. Clim. Change 70, 137–164.
Polomski, J., and Kuhn, N. (1998). Wurzelsysteme. Birmensdorf: Eidgenössische Forschungsanstalt für Wald, Schnee und Landschaft.
Rossi, S., Morin, H., Deslauriers, A., and Plourde, P.-Y. (2011). Predicting xylem phenology in black spruce under climate warming. Global Change Biol. 17, 614–625.
Rossi, S., Simard, S., Rathgeber, C. B. K., Deslauriers, A., and De Zan, C. (2009). Effects of a 20-day-long dry period on cambial and apical meristem growth in Abies balsamea seedlings. Trees 23, 85–93.
Steppe, K., DePauw, D. J. W., Lemeur, R., and Vanrolleghem, P. A. (2005). A mathematical model linking tree sap flow dynamic to daily stem diameter fluctuations and radial stem growth. Tree Physiol. 26, 257–273.
Waring, R. H., Whitehead, D., and Jarvis, P. G. (1979). The contribution of stored water to transpiration in Scots pine. Plant Cell Environ. 2, 309–317.
Whitehead, D., and Jarvis, P. G. (1981). “Coniferous forests and plantations,” in Water Deficits and Plant Growth, ed. T. T. Kozlowski (New York: Academic Press), 49–52.
Zhang, X., Hogg, W. D., and Mekis, E. (2001). Spatial and temporal characteristics of heavy precipitation events over Canada. J. Clim. 14, 1923–1936.
Zhang, X., Vincent, L. A., Hogg, W. D., and Niitsoo, A. (2000). Temperature and precipitation trends in Canada during the 20th century. Atmos. Ocean 38, 395–429.
Zweifel, R., and Häsler, R. (2001). Dynamics of water storage in mature subalpine Picea abies: temporal and spatial patterns of change in stem radius. Tree Physiol. 21, 561–569.
Zweifel, R., Item, H., and Häsler, R. (2001). Link between diurnal stem radius changes and tree water relations. Tree Physiol. 21, 869–877.
Keywords: boreal forest, climate change, stem cycle approach, plant–water relationships, soil depth, water reserves
Citation: Turcotte A, Rossi S, Deslauriers A, Krause C and Morin H (2011) Dynamics of depletion and replenishment of water storage in stem and roots of black spruce measured by dendrometers. Front. Plant Sci. 2:21. doi: 10.3389/fpls.2011.00021
Received: 03 March 2011;
Paper pending published: 15 March 2011;
Accepted: 03 June 2011;
Published online: 17 June 2011.
Edited by:
Karina Vera Rosa Schafer, Rutgers University Newark, USAReviewed by:
Heidi Renninger, Boston University, USADirk Vanderklein, Montclair State University, USA
Javier Vázquez, University of Huelva, Spain
Copyright: © 2011 Turcotte, Rossi, Deslauriers, Krause and Morin. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: Sergio Rossi, Département des Sciences Fondamentales, Université du Québec à Chicoutimi, 555 Boulevard de l’Université, Chicoutimi, QC, Canada G7H 2B1. e-mail:c2VyZ2lvLnJvc3NpQHVxYWMuY2E=