 Jay Ram Lamichhane
Jay Ram Lamichhane Vittorio Venturi
Vittorio Venturi- 1INRA, UAR 1240 Eco-Innov, Thiverval-Grignon, France
- 2International Centre for Genetic Engineering and Biotechnology, Trieste, Italy
Plant diseases are often thought to be caused by one species or even by a specific strain. Microbes in nature, however, mostly occur as part of complex communities and this has been noted since the time of van Leeuwenhoek. Interestingly, most laboratory studies focus on single microbial strains grown in pure culture; we were therefore unaware of possible interspecies and/or inter-kingdom interactions of pathogenic microbes in the wild. In human and animal infections, it is now being recognized that many diseases are the result of multispecies synergistic interactions. This increases the complexity of the disease and has to be taken into consideration in the development of more effective control measures. On the other hand, there are only a few reports of synergistic pathogen–pathogen interactions in plant diseases and the mechanisms of interactions are currently unknown. Here we review some of these reports of synergism between different plant pathogens and their possible implications in crop health. Finally, we briefly highlight the recent technological advances in diagnostics as these are beginning to provide important insights into the microbial communities associated with complex plant diseases. These examples of synergistic interactions of plant pathogens that lead to disease complexes might prove to be more common than expected and understanding the underlying mechanisms might have important implications in plant disease epidemiology and management.
Introduction
A widely accepted current concept is that a pathogen colonizes a host and responds to the host environment resulting in the manipulation of expression of its resistance genes. Many studies have demonstrated this type of host–pathogen interaction in monospecies infections. In contrast to this, in the wild, microbes have been known to be part of complex multispecies consortia/communities since the time of van Leeuwenhoek during 1600s (Gest, 2004). The earliest reports regarding microbial communities as causal agents of a disease are attributed to Pasteur in the 1800s who observed that a disease can also be due to synergistic interactions of different microorganisms. Although microbial plant diseases of definite etiology are still mainly considered as being caused by single microbial cultures, evidence is now beginning to mount that there can be synergisms between different pathogens in complex plant diseases.
In human pathology, there is now a growing awareness that infectious agents frequently do not operate alone and their virulence can be affected by their interaction with other commensals or pathogens (Singer, 2010). Consequently, the study of multispecies synergistic interactions is emerging as a new important subject for better understanding of microbial diseases (Short et al., 2014). Several examples of synergistic commensal–pathogen and pathogen–pathogen interactions that lead to increased human disease severity have been recently reported (Singer, 2010; Peters et al., 2012; Bosch et al., 2013). Similar examples of pathogen–pathogen synergistic interactions that result in increased disease severity in animals have also been reported (Harms et al., 2001; Ellis et al., 2004). For example, polyparasitism as a decisive factor has been recently demonstrated in a protozoal disease in marine mammals; more precisely, co-infections of Toxoplasma gondii and Sarcocystis neurona were more recurrently related with mortality and protozoal encephalitis when compared to solitary infections (Gibson et al., 2011).
In contrast to mammalian pathology, the concept of monospecies/monostrain infections is more deeply rooted in plant pathology. In such cases, plant disease epidemics have almost exclusively been associated to a single pathogen belonging to a clonal group. Examples are bacterial canker of kiwifruit (Mazzaglia et al., 2012), bleeding canker of horse chestnut (Green et al., 2010), diseases of fruit and nut trees (Hajri et al., 2012), Bayoud disease of date palm (Tantaoui et al., 1996) and potato late blight (Goodwin et al., 1995). None of these studies have focused on the possible role of other microbial populations associated with infected plants in disease outcome. This is not surprising as new methodologies to analyze total microbial populations from diseased plant tissues are only beginning to be available on a routine basis. The few studies, using traditional approaches, reveal that many plant species can frequently be infected at the same time by more than one pathogenic species (Fitt et al., 2006); in many cases, a single microbe infection may not result in severe disease symptoms while the co-infection with another microbial species may lead to severe disease development due to synergistic interactions.
The objective of this review is to highlight, describe, and discuss known examples of plant diseases which involve pathogen–pathogen interactions. There are two main possibilities in which different plant pathogens interact: (i) a disease complex where disease is the result of the interaction of several plant pathogens belonging to the same species or phylum, (ii) a disease complex due to different plant pathogens belonging to different phyla. This review does not cover plant diseases involving commensal–pathogen, nematode–microbe, or insect–microbe interactions. The universality of these synergistic interactions between plant pathogens in the living world is an alarm bell and microbiologists and plant pathologists need therefore to better consider in the future that one pathogenic agent can team up with others rather than acting alone.
Synergistic Pathogen–Pathogen Interactions
Plant diseases where more than one pathogen is involved in the infection process are commonly termed as “complex” since their diagnosis and subsequent control are more complicated. Such diseases occur as a result of a network that involves a wide range of microbial interactions. Monoculture inoculations are commonly performed to evaluate the pathogenicity behavior of a given pathogen. Consequently, our knowledge of their possible synergism that leads to increased disease severity is poor. It is likely that synergism among different pathogens leading to more severe disease symptoms occur more often than expected (Begon et al., 2006). Such synergistic interactions in plants may be of crucial importance for the understanding of microbial pathogenesis and evolution and consequent development of effective disease control strategies.
A non-exhaustive list of synergistic pathogen–pathogen infections in plants that often lead to increased disease severity is reported in Table 1. This overall picture is beginning to facilitate the understanding of epidemiology and control of numerous complex diseases. There are now several recent reports on the diagnosis of complex diseases and their successful management (Gleason et al., 2011; Clark et al., 2012; Martin et al., 2013; Freeman et al., 2014).
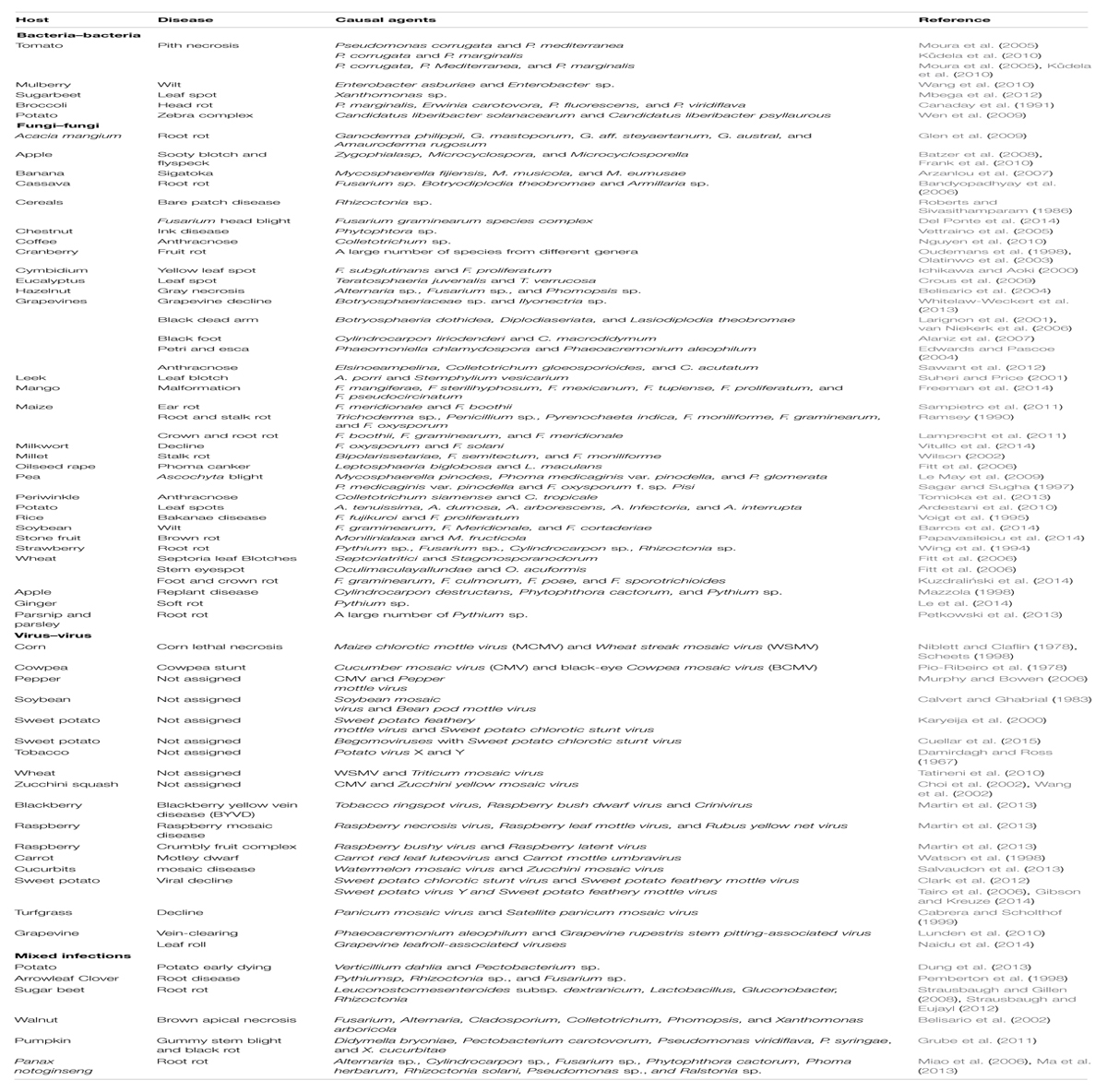
TABLE 1. Pathogen–pathogen synergistic interactions that lead to plant disease occurrence and increased disease severity.
Bacteria–Bacteria Interactions
Tomato pith necrosis is thus far a leading example of co-infection due to synergistic interactions among several bacterial pathogens. Overall, eight bacterial species namely Pseudomonas cichorii (Wilkie and Dye, 1974), P. corrugata (Scarlett et al., 1978), P. viridiflava (Goumas and Chatzaki, 1998), P. mediterranea (Saygili et al., 2008), P. fluorescens (Saygili et al., 2008), Pectobacterium atrosepticum (Malathrakis and Goumas, 1987), Pectobacterium carotovorum (Dhanvanthari and Dirks, 1987; formerly Erwinia), and Dickeya chrysanthemi (Formerly Erwinia; Alivizatos, 1985) can cause tomato pith necrosis alone or in association with the other bacterial species. The severity of the disease is greatly enhanced when co-infection of one or more bacterial species occurs. In particular, co-infection of P. corrugata–P. marginalis, or P. corrugata–P. mediterranea have been reported to cause severe infection in tomato (Moura et al., 2005; Saygili et al., 2008; Kůdela et al., 2010). Similarly, bacterial soft or head rot of broccoli is another complex disease caused by numerous plant pathogenic bacteria. Overall, Pectobacterium carotovorum, P. marginalis, P. fluorescens, and P. viridiflava have been reported to cause broccoli head rot (Canaday et al., 1991). Bacterial strains belonging to these species are also capable of causing soft rot on unwounded broccoli when co-inoculated. The mechanism(s) for this cooperativity among different bacterial species is currently unknown.
Fungi–Fungi Interactions
Synergistic interactions between different fungal pathogens have been studied intensively. For example, the young grapevine decline disease is present across many regions worldwide, and is caused by the following fungal pathogens when present alone: Ilyonectris sp., Phaeomoniella chlamydospora, Togninia sp., and Botryosphaeriaceae sp. (Mugnai et al., 1999; Gramaje and Armengol, 2011). The fungal causal agent(s) vary considerably between the grapevine producing regions. A recent study demonstrated that co-infection of several fungal species belonging to Botryosphaeriaceae sp. and Ilyonectria sp. results in very severe decline of young grafted grapevines in the field (Whitelaw-Weckert et al., 2013). Similarly, laboratory experiments further confirmed that co-inoculation of Ilyonectria and Botryosphaeriaceae isolates led to an increased disease severity compared to monoculture inoculations of Ilyonectria isolates (Whitelaw-Weckert et al., 2013). The different pathogens isolated from grapevine decline symptoms throughout cultivated areas are likely to have co-evolved due to their close association. Prior to these studies, only one pathogen was thought to be the causal agent of grapevine decline depending on the region due to a marked dominance of one species over all the other ones for each infection.
Numerous examples of co-existence of fungal pathogens on arable crops has been described in the UK (Fitt et al., 2006). Such examples include a disease complex of wheat leaves known as septoria leaf blotches caused by Septoria tritici and Stagonospora nodorum; wheat stem affected by Oculimacula yallundae and O. acuformis and phoma stem canker on oilseed rape caused by Leptosphaeria biglobosa and L. maculans. Another complex disease of wheat caused by a group of Fusarium species is foot and crown rot. Overall, four species of the pathogen (Fusarium graminearum, F. culmorum, F. poae, and F. sporotrichioides) are associated with the disease although their prevalence differs from one geographic region to another (Kuzdraliński et al., 2014). It has also been reported that the majority of fields in eastern Poland are subjected to the attack of at least one or two of Fusarium species. The presence of F. graminearum was found to foster the occurrence of F. culmorum and this result was observable also for F. poae and F. sporotrichioides. An additional disease complex of major economic significance worldwide is Fusarium head blight, especially due to the grain contamination with harmful mycotoxins produced by the fungus during pathogenesis (McMullen et al., 2012). Over 16 known species of the F. graminearum species complex have been reported as the causal agent of Fusarium head blight (O’Donnell et al., 2008; Yli-Mattila et al., 2009; Sarver et al., 2011). Studies carried out in Brazil show that the prevalence of the species in Fusarium head blight varies from one geographic region to another (Del Ponte et al., 2014). Our current knowledge is very poor concerning mechanisms that explain the geographic variation and prevalence of specific pathogens in plants affected by a particular complex disease and it is possible that such variations are related to the ecological preference of these pathogens. Moreover, abiotic factors and cultural practices might also influence this variation in pathogen prevalence.
Black spot disease complex of pea was previously known to be caused by three fungal pathogens (Le May et al., 2009); however, four additional pathogens have been recently found in association with this disease. These new pathogens include Phoma koolunga (Davidson et al., 2009), Phoma herbarum (Li et al., 2011), Boerema exigua var. exigua (Li et al., 2012), and Phoma glomerata (Tran et al., 2014). All these fungi are necrotrophic, generalist and polyphagous species and these characteristics favor colonization of new environments. It is possible that these pathogens use synergism as a strategy to infect a large variety of plants which might also explain why some pathogens occur more easily than others in a given environment or plant host.
Leaf spot of eucalyptus is a disease complex caused by numerous species of fungi of the genus Teratosphaeria (Crous et al., 2009). Diseased eucalyptus plants sampled from South Africa yielded two fungal species (i.e., Teratosphaeria juvenalis and T. verrucosa) that co-occur in the same leaves and even in the same spots. In Australia, T. gauchensis and T. zuluensis, (which predominantly cause eucalyptus stem cankers) have been reported to occur in leaf spots either alone or in association with some of the other species belonging to Teratosphaeria (Crous et al., 2009). On mango, a complex disease known as mango malformation is caused by F. mangiferae, F. mexicanum, F. proliferatum, F. pseudocircinatum, F. sterilihyphosum, and F. tupiense where individual species of this fungus prevail in association with the symptomatic tissues (Freeman et al., 2014).
The importance of the temporal order of host infection by different pathogens is another important factor to be considered. Several reports have been described in the literature in this regard. Le May et al. (2009) demonstrated that the simultaneous inoculation of two plant pathogenic fungi (Mycosphaerella pinodes and Phoma medicaginis var. pinodella) associated to the Ascochyta blight disease complex limits disease development and their reproduction. However, when plants pre-inoculated with one pathogen were then inoculated with another there was a marked increase in severity of the disease. In contrast to this report, Sagar and Sugha (1997) reported a decrease in necrotic symptoms of pea caused by Phoma medicaginis var. pinodella previously inoculated with the vascular root pathogen F. oxysporum f. sp. pisi. The order/succession of host infection by each pathogen in a complex disease and its trophic level might affect the nature of interaction that these pathogens eventually develop during disease occurrence. Such temporal effects could also be related to the notion of ecological niche.
There are also reports of synergistic interactions between more or less aggressive strains of numerous pathogens. Kaur et al. (2011) for example demonstrated that the severity of white rust symptoms (caused by the pathogen Albugo candida) on mustard increases and the symptoms appear earlier when a host highly susceptible to A. candida but resistant to Hyaloperonospora parasitica (the causal agent of downy mildew), was first inoculated on day 1 with a less aggressive strain of H. parasitica followed by a 10-days-post inoculation with an aggressive strain of A. candida. Taken together, the outcomes due to co-occurrence of the same pathogens on the same host may result in antagonism and/or synergism which are likely influenced by the order of their association with the infected plant. In-depth future studies are needed to consider the temporal aspect of infection by different pathogens and their interactions, both among themselves and with the plant, to uncover the possible underlying mechanisms.
Virus–Virus Interactions
Disease synergisms among two or more plant pathogenic viruses increasing the severity of symptoms has been reported on a variety of crop species (Table 1). For example, Maize chlorotic mottle virus (MCMV) and Wheat streak mosaic virus (WSMV) cause corn lethal necrosis. The interactions between these two viruses results in a significant increase (up to 10-fold) of the MCMV concentration in plants (Scheets, 1998). In addition, WSMV infection is considerably enhanced by the presence of MCMV both in terms of frequency and intensity. Likewise, a strong synergistic interaction was found between Cucumber mosaic virus (CMV) and black-eye Cowpea mosaic virus (BCMV) in severely stunted cowpea in fields (Pio-Ribeiro et al., 1978). In an experimental inoculation study, each virus caused relatively mild disease when inoculated singly and plants showed significantly reduced stunting. In contrast, disease severity and the extent of stunting increased when these two viruses are co-inoculated under the same conditions. Another example is blackberry yellow vein disease (BYVD) complex which is caused by the cooperation between different viral species (Martin et al., 2013). More specifically, Tobacco ringspot virus, Raspberry bush dwarf virus, and a new virus which belongs to the genus Crinivirus are involved. The BYVD disease severity was stringently related with the number of viruses infecting plants. No disease symptom was caused by the incoming viruses in single infections while symptoms clearly become visible in mixed infections. Hence, in some cases the absolute number of viruses infecting plants is likely to be more important than the type of viruses involved in the infection. However, the role of vectors in transmitting complex viral diseases is not clear in the above described examples. For example, are the viruses commonly transmitted by the same vector? If this is the case, are there likely to be differences in the acquisition time or latency of the organisms that could affect whether they are co-transmitted? Answers to these questions are likely to be important in disease control; however, information available to date is not sufficient and further studies are required.
In addition to synergisms between plant viruses, several studies reported mixed virus infections leading to mutual exclusion (antagonism); this aspect has been recently reviewed by Syller (2012).
Mixed Interactions
There are a few reports in the literature of plant disease complexes involving association of more than one pathogenic microbial phyla (Table 1). An example is brown apical necrosis of walnut fruit where numerous plant pathogenic fungi (Fusarium, Alternaria, Cladosporium, Colletotrichum, and Phomopsis) and a bacterium (Xanthomonas arboricola) are involved (Belisario et al., 2002). Another example is root rot disease complex of Panax notoginseng where a large number of plant pathogenic fungi (Alternaria panax, Alternaria tenuis, Cylindrocarpon destructans, Cylindrocarpon didynum, F. solani, F. oxysporum, Phytophthora cactorum, Phoma herbarum, and Rhizoctonia solani) and bacteria (Pseudomonas sp. and Ralstonia sp.) have been found (Miao et al., 2006; Ma et al., 2013). The mechanisms of interaction that result in communication and synergism of pathogens in these complex diseases are currently unknown.
New Approaches are Needed for Studies of Complex Plant Diseases
An initial thorough analysis that correctly identifies the disease causing agent(s) is the primary step of managing a plant disease (Adams et al., 2013). Suitable disease management tools can then be applied such as the use of an anti-microbial compounds which can be administered depending on the plant type and part affected by the disease. Although the application of a chemical substance can be of importance, a more sustainable disease management can be achieved only through the development of more long-term strategies. To this aim, a better knowledge of pathogen–pathogen synergism in causing complex plant diseases is of paramount importance. For example, many anti-microbial strategies currently used in agriculture are specific to control a given microbial pathogen. Targeted chemical control strategies become limiting when more than one pathogenic agent contributes to the disease as the application of the specific substance may not necessarily result in successful disease management. It is therefore important to study plant disease complexes, the synergisms in pathogen–pathogen interactions as well as the underlying mechanisms to identify important links that may be manipulated to ensure crop health. This could be a difficult task since disease complexes are related to environmental conditions, cultural practices, and geography (Willocquet et al., 2002). It is therefore important to design the experimental approach leading to identification of pathogen cortege in relation to the crop production system.
The diagnosis and management of complex diseases can be lengthy resulting in significant yield losses. Often the use of classical isolation techniques on selected or semi-selected media may not yield any of the causal agents or sometimes only their partial isolation. Because of the complexity of polymicrobial diseases, the study in this regard was somewhat overlooked in the past. In most of the studies reported in Table 1, the authors performed the isolation of pathogen on culture growth media. In addition, other more specific (e.g., immunofluorescence or PCR) or generic (e.g., morphological identification) assays were used. However, currently we have new knowledge and techniques which may facilitate the understanding of the total microbial species involved in plant diseases as well as the underlying mechanisms. Hence, studies of complex diseases now need to benefit from culture-independent analyses (high-throughput sequencing for example). This approach does not have the limitations of the classical culture-based approach, which is often lengthy and costly (Nikolaki and Tsiamis, 2013). In the modern era of biodiversity surveillance, techniques such as next-generation sequencing (NGS) have enabled high-throughput analyses of complex microbial populations (reviewed by van Dijk et al., 2014). This has transformed microbiology and has revealed that microbial diversity is vastly underestimated based on classical cultivation-based techniques (Gilbert and Dupont, 2011). In the last 10 years, metagenomic projects have been combined with NGS technologies boosting studies in microbial ecology at a very fast pace (Venter et al., 2004; Tringe and Hugenholtz, 2008).
Although plant pathology in general and plant disease complex in particular could stand to gain from exploiting NGS and metagenomic approaches, the current literature reveals only a limited number of applications of this technology. Only a few studies have applied such methods with a regard to the diagnosis of new pathogens (Adams et al., 2009, 2013). For example, sequences of an entire viral genome (determined via a single step of high-throughput parallel sequencing) highlighted the presence of three novel viruses in sweet potato plants which were infected with known pathogenic viruses occurring at extremely low titers (Kreuze et al., 2009). These novel technologies are thus a powerful tool to understand the implication of two or more microbes and their contribution in plant disease occurrence.
There are some limitations in using these novel OMICs methods with respect to studying the role of microbial consortia and plant disease. Although these will help to better characterize complex diseases they will not necessarily allow to determine which microbe is the dominant factor in the disease occurring process. Identification of the infection-site derived nucleic acid sequence is not unequivocal evidence that the microorganism in question is the causal agent of disease. For example, Adams et al. (2009) determined the complete virus genome sequence of a pathogenic virus via metagenomic anaylsis. This data is convincing proof that a transmissible infection with virus-like symptoms was linked with the occurrence of the new Cucumovirus full genome present in the infection site despite the fact that Koch’s postulates were not fulfilled. Importantly, viral particles were not observed nor the disease was re-established in the original host highlighting that only the virus presence can be determined via metagenomic sequence analysis. Metagenomics will therefore pose a new challenge for taxonomy and role of phytopathogens in disease (Studholme et al., 2011). Another aspect when considering mixed infections will be that of distinguishing pathogens from saprotrophic microorganisms.
Another important feature in studying complex diseases concerns growth models and the pathogen cortege. Examples are RICEPEST (Willocquet et al., 2002) and WHEATPEST (Willocquet et al., 2008) models which have been developed to simulate yield losses due to several pests under different production situations. These models are the first to include the impact of several diseases on yield losses. A limit to these models is that although impact of the different diseases is considered, they do not take into account the potential interaction between the pathogens. Future studies need to take into account such limits in order to develop models that simulate the possible synergism between plant pathogens.
Recently, Elena et al. (2014) proposed that evolutionary game theory provides an adequate theoretical framework to analyze mixed viral infections and to predict the long-term evolution of the mixed populations. Here we recommend that the same approach can be used to analyze mixed fungal and bacterial infections which need to be considered in the future studies.
What Mechanisms are Known?
Synergistic interactions between pathogens in humans have been reported to occur through several mechanisms such as chemical signaling influencing gene expression or via metabolic exchange/complementarity in order to avoid competition for nutrients and improve metabolic ability of the consortium (Frey-Klett et al., 2011). Pathogen–pathogen interactions are also known to result in viral induced bacterial adhesion, interference with the host immune system, production of viral products, direct bacterial effectors and viral-derived disruption of the epithelium (Singer, 2010; Peters et al., 2012; Bosch et al., 2013). On the other hand, studies involving mixed infections in plant diseases are still in their infancy and the underlying mechanisms of possible synergistic interactions are currently unknown. However, it has been recently reported that a plant pathogen undergoes interspecies signaling via quorum sensing signals with residential commensal microbiota indicating the occurrence of intimate multispecies interactions in planta (Hosni et al., 2011). Signaling among different bacterial species is also likely to play an important role in the synchronization of behaviors as well as expression of virulence factors in mixed populations. The example of Hosni et al. (2011) could pave the way for the discovery of other interspecies interactions among microbes living in close association with plants.
Synergistic interactions among bacterial pathogens could also be indirect as for example the ability of certain harmless or beneficial plant-associated bacteria and pathogens to suppress host immunity or alter the plant micro-environment that can promote colonization by other pathogens. Several studies have demonstrated that plant defenses are induced following the infection of an avirulent bacterial strain resulting in its own growth restriction as well as that of a co-inoculated virulent strain (Klement and Lovrekovich, 1961; Averre and Kelman, 1964; Omer and Wood, 1969). The growth of non-pathogenic bacterial strains could reach higher cell numbers when co-inoculated with pathogenic bacteria (Young, 1974).
On the other hand, growth differences of virulent bacterial species could occur when they are co-inoculated owing to variations in fitness rather than virulence (Llama-Palacios et al., 2002). Similarly, growth interference in a mixed infections between different strains of P. syringae has been reported to be influenced by the initial population size (Macho et al., 2007). The lowest range of initial population necessary to circumvent interference has been reported to be dependent both on the type of virulence factor that differentiates the co-inoculated strains as well as the pathogen aggressiveness. In another study, growth interference in a mixed infection between different strains of P. syringae is strain-dependent and the populations of strains in mixed inoculations were lower than those in independent inoculations (Bartoli et al., 2015). The mechanisms supporting these synergistic and/or antagonistic interactions need to be explored taking into account the importance of immune suppression or modulation of phytohormone-based signaling mechanisms.
The mechanism of synergistic interaction among the Sweet potato feathery mottle virus and Sweet potato chlorotic stunt virus has been investigated and shown to be mediated by the SPCSV encoded RNase3 protein (Cuellar et al., 2009). The authors suggest that RNase3 may synergize Sweet potato feathery mottle virus and other viruses by targeting a specific host component via interference with small-RNA biogenesis; the precise mechanism of this interaction is currently unknown.
A previous study (García-Marcos et al., 2009) on synergistic interactions between Potato virus X and Potato virus Y, which led to an increased systemic infection in Nicotiana benthamiana, reported transcriptional changes and oxidative stress associated with the synergistic infection. This stress correlated with the misregulation of antioxidative genes in a microarray experiment. Expression of genes encoding oxylipin biosynthesis were upregulated by the synergistic infection caused by the two viruses and were not by single infection with Potato virus X or Potato virus Y. Interestingly, oxylipin biosynthesis genes were recently shown to positively regulate programmed cell death during compatible infections with the synergistic pair Potato virus X-Potato virus Y and Tomato spotted wilt virus (García-Marcos et al., 2013).
Polymicrobial Diseases and Koch’s Postulates
Of the four criteria postulated by Koch [they are (i) the microbe must be isolated from an infected host and obtained in pure culture; (ii) it must cause infection when inoculated into a healthy host; (iii) it must be re-isolated from the inoculated, symptomatic organism; and (iv) it must be shown to be the same as the originally cultured microorganism], some are not valid for polymicrobial infections as for example re-inoculation of the pathogen not necessarily causes disease if synergism is lacking. Another postulate raised by Koch was that the pathogen should be isolated from diseased and not from healthy organisms. This is also challenged for an increasing number of human diseases as well as plant diseases. Pierce’s disease of grapevine is an example, where the bacterial pathogen Xylella fastidiosa can colonize xylem vessels of asymptomatic plants for a long period of time (Purcell, 2013). Another issue related to these postulates are the non-culturable pathogens (Oliver, 2010) for which none of the Koch’s criteria can be fulfilled. The identification of polymicrobial diseases and other issues, some raised here, suggests that Koch’s postulates need to be revised. For example, pathosystems may be divided into two groups; simple diseases in which only one organism is involved, and complex diseases in which there is an interplay of more than one organism (e. g., insect, commensal, pathogen, non-culturable organism etc.). For the complex diseases, an additional criterion (fifth) taking into account the interactions between microrganisms involved in disease occurrence that are positively correlated with disease occurrence and/or severity might be necessary.
Concluding Remarks
The pathogenic microorganisms associated with plants now need to be isolated/studied with the view that possible antagonistic, mutualistic, or synergistic interactions are taking place. A careful assessment of the roles of all the microorganisms isolated from the infection sites needs to be evaluated as multispecies interactions and consortia can be involved in establishment and aggravation of the disease. This will need to involve the interdisciplinary research collaboration between bacteriologists, mycologists and virologists. Understanding the biology and molecular interactions of these inter-microbial processes may be important in defining new targets and strategies for disease control.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Dr. Claudia Bartoli and Dr. Sujatha Subramoni for the critical reading of the paper.
References
Adams, I. P., Glover, R. H., Monger, W. A., Mumford, R., Jackeviciene, E., Navalinskiene, M., et al. (2009). Next-generation sequencing and metagenomic analysis: a universal diagnostic tool in plant virology. Mol. Plant Pathol. 10, 537–545. doi: 10.1111/j.1364-3703.2009.00545.x
Adams, I. P., Miano, D. W., Kinyua, Z. M., Wangai, A., Kimani, E., Phiri, N., et al. (2013). Use of next-generation sequencing for the identification and characterization of Maize chlorotic mottle virus and Sugarcane mosaic virus causing maize lethal necrosis in kenya. Plant Pathol. 62, 741–749. doi: 10.1111/j.1365-3059.2012.02690.x
Alaniz, S., León, M., Vicent, A., García-Jiménez, J., Abad-Campos, P., and Armengol, J. (2007). Characterization of Cylindrocarpon species associated with black foot disease of grapevine in Spain. Plant Dis. 91, 1187–1193. doi: 10.1094/PDIS-91-9-1187
Alivizatos, A. S. (1985). Bacterial wilt of tomato in Greece caused by Erwinia chrysanthemi. Plant Pathol. 34, 638–639. doi: 10.1111/j.1365-3059.1985.tb01419.x
Ardestani, S. T., Sharifnabi, B., Zare, R., and Moghadam, A. A. (2010). New Alternaria species associated with potato leaf spot in various potato growing regions of Iran. Iran. J. Plant Pathol. 45, 83–86.
Arzanlou, M., Abeln, E. C. A., Kema, G. H. J., Waalwijk, C., Carlier, J., de Vries, I., et al. (2007). Molecular diagnostics for the sigatoka disease complex of banana. Phytopathology 97, 1112–1118. doi: 10.1094/PHYTO-97-9-1112
Averre, C. W., and Kelman, A. (1964). Severity of bacterial wilt as influenced by ratio of virulent to avirulent cells of Pseudomonas solanacearum in inoculum. Phytopathology 54, 779–783.
Bandyopadhyay, R., Mwangi, M., Aigbe, S. O., and Leslie, J. F. (2006). Fusarium species from the cassava root rot complex in west Africa. Phytopathology 96, 673–676. doi: 10.1094/PHYTO-96-0673
Barros, G. G., Zanon, M. S. A., Chiotta, M. L., Reynoso, M. M., Scandiani, M. M., and Chulze, S. N. (2014). Pathogenicity of phylogenetic species in the Fusarium graminearum complex on soybean seedlings in Argentina. Eur. J. Plant Pathol. 138, 215–222. doi: 10.1007/s10658-013-0332-2
Bartoli, C., Lamichhane, J. R., Berge, O., Guilbaud, C., Varvaro, L., Balestra, G. M., et al. (2015). A framework to gauge the epidemic potential of plant pathogens in environmental reservoirs: the example of kiwifruit canker. Mol. Plant Pathol. 16, 137–149. doi: 10.1111/mpp.12167
Batzer, J. C., Arias, M. M. D., Harrington, T. C., Gleason, M. L., Groenewald, J. Z., and Crous, P. W. (2008). Four species of Zygophiala (Schizo-thyriaceae, Capnodiales) are associated with the sooty blotch and flyspeckcomplex on apple. Mycologia 100, 246–258. doi: 10.3852/mycologia.100.2.246
Begon, M., Towsend, C. R., and Harper, J. L. (2006). Ecology, Individuals, Populations and Communities, 4th Edn. London: Blackwell Science.
Belisario, A., Maccaroni, M., Coramusi, A., Corazza, L., Pryor, B. M., and Figuli, P. (2004). First report of Alternaria species groups involved in disease complexes of hazelnut and walnut fruit. Plant Dis. 88, 426–426. doi: 10.1094/PDIS.2004.88.4.426A
Belisario, A., Maccaroni, M., Corazza, L., Balmas, V., and Valier, A. (2002). Occurrence and etiology of brown apical necrosis on Persian (English) walnut fruit. Plant Dis. 86, 599–602. doi: 10.1094/PDIS.2002.86.6.599
Bosch, A. A., Biesbroek, G., Trzcinski, K., Sanders, E. A. M., and Bogaert, D. (2013). Viral and bacterial interactions in the upper respiratory tract. PLoS Pathog. 9:e1003057. doi: 10.1371/journal.ppat.1003057
Cabrera, O., and Scholthof, K.-B. G. (1999). The complex viral etiology of St. Augustine Decline. Plant Dis. 83, 902–904. doi: 10.1371/journal.ppat.1003057
Calvert, L. A., and Ghabrial, S. A. (1983). Enhancement by Soybean mosaic virus of bean pod mottle virus titer in doubly infected soybean. Phytopathology 73, 992–997. doi: 10.1094/PDIS.1999.83.10.902
Canaday, C. H., Wyatt, J. E., and Mullins, J. A. (1991). Resistance to broccoli to bacterial soft rot caused by Pseudomonas marginalis and fluorescent Pseudomonas species. Plant Dis. 75, 715–720. doi: 10.1094/Phyto-73-992
Choi, S. K., Yoon, J. Y., Ryu, K. H., Choi, J. K., Palukaitis, P., and Park, W. M. (2002). Systemic movement of a movement-deficient strain of Cucumber mosaic virus in zucchini squash is facilitated by a cucurbit infecting Potyvirus. J. Gen. Virol. 83, 3173–3178. doi: 10.1094/PD-75-0715
Clark, C. A., Davis, J. A., Abad, J. A., Cuellar, W. J., Fuentes, S., Kreuze, J. F., et al. (2012). Sweet potato viruses: 15 years on understanding and managing complex diseases. Plant Dis. 96, 168–185. doi: 10.1094/PDIS-07-11-0550
Crous, P. W., Groenewald, J. Z., Summerell, B. A., Wingfield, B. D., and Wingfield, M. J. (2009). Co-occurring species of Teratosphaeria on Eucalyptus. Persoonia 22, 38–48. doi: 10.3767/003158509X424333
Cuellar, W. J., Galvez, M., Fuentes, S., Tugume, J., and Kreuze, J. (2015). Synergistic interactions of Begomoviruses with Sweet potato chlorotic stunt virus (genus Crinivirus) in sweetpotato (Ipomoea batatas L.). Mol. Plant Pathol. 16, 459–471. doi: 10.1111/mpp.12200
Cuellar, W. J., Kreuze, J. F., Rajamäki, M.-L., Cruzado, K. R., Untiveros, M., and Valkonen, J. P. (2009). Elimination of antiviral defense by viral RNase III. Proc. Natl. Acad. Sci. U.S.A. 106, 10354–10358. doi: 10.1073/pnas.0806042106
Damirdagh, I. S., and Ross, A. F. (1967). A masked synergistic interaction of Potato virus X and Y in inoculated leaves of tobacco. Virology 31, 296–307. doi: 10.1016/0042-6822(67)90174-2
Davidson, J. A., Hartley, D., Priest, M., Herdina, M. K., McKay, A., and Scott, E. S. (2009). A new species of Phoma causes ascochyta blight symptoms on field peas (Pisum sativum) in South Australia. Mycologia 101, 120–128. doi: 10.3852/07-199
Del Ponte, E. M., Spolti, P., Ward, T. J., Gomes, L. B., Nicolli, C. P., Kuhnem, P. R., et al. (2014). Regional and field-specific factors affect the composition of Fusarium head blight pathogens in subtropical no-till wheat agroecosystem of Brazil. Phytopathology 105, 246–254. doi: 10.1094/PHYTO-04-14-0102-R
Dhanvanthari, B. N., and Dirks, V. A. (1987). Bacterial stem rot of greenhouse tomato: etiology, spatial distribution, and the effect of high humidity. Phytopathology 77, 1457–1463. doi: 10.1094/Phyto-77-1457
Dung, J. K. S., Johnson, D. A., and Schroeder, B. K. (2013). Role of co-infection by Pectobacterium and Verticillium dahliae in the development of early dying and aerial stem rot of Russet Burbank potato. Plant Pathol. 63, 299–307. doi: 10.1111/ppa.12086
Edwards, J., and Pascoe, I. G. (2004). Occurrence of Phaeomoniella chlamydospora and Phaeoacremonium aleophilum associated with Petri disease and esca in Australian grapevines. Australas. Plant Pathol. 33, 273–279. doi: 10.1071/AP04016
Elena, S. F., Bernet, G. P., and Carrasco, J. L. (2014). The games plant viruses play. Curr. Opin. Virol. 8, 62–67. doi: 10.1016/j.coviro.2014.07.003
Ellis, J., Clark, E., Haines, D., West, K., Krakowka, S., Kennedy, S., et al. (2004). Porcine circovirus-2 and concurrent infections in the field. Vet. Microbiol. 98, 159–163. doi: 10.1016/j.vetmic.2003.10.008
Fitt, B. D. L., Huang, Y. J., Bosch, F. V. D., and West, J. S. (2006). Coexistence of related pathogen species on arable crops in space and time. Annu. Rev. Phytopathol. 44, 163–182. doi: 10.1146/annurev.phyto.44.070505.143417
Frank, J., Crous, P. W., Groenewald, J. Z., Oertel, B., Hyde, K. D., Phengsintham, P., et al. (2010). Microcyclospora and Microcyclosporella: novel genera accommodating epiphytic fungi causing sooty blotch on apple. Persoonia 24, 93–105. doi: 10.3767/003158510X510560
Freeman, S., Shtienberg, D., Maymon, M., Levin, A. G., and Ploetz, R. C. (2014). New insights into mango malformation disease epidemiology lead to a new integrated management strategy for subtropical environments. Plant Dis. 98, 1456–1466. doi: 10.1094/PDIS-07-14-0679-FE
Frey-Klett, P., Burlinson, P., Deveau, A., Barret, M., Tarkka, M., and Sarniguet, A. (2011). Bacterial-fungal interactions: hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. Rev. 75, 583–609. doi: 10.1128/MMBR.00020-11
García-Marcos, A., Pacheco, R., Manzano, A., Aguilar, E., and Tenllado, F. (2013). Oxylipin biosynthesis genes positively regulate programmed cell death during compatible infections with the synergistic pair Potato virus X-Potato virus Y and Tomato spotted wilt virus. J. Virol. 87, 5769–5783. doi: 10.1128/JVI.03573-12
García-Marcos, A., Pacheco, R., Martiáñez, J., González-Jara, P., Díaz-Ruíz, J. R., and Tenllado, F. (2009). Transcriptional changes and oxidative stress associated with the synergistic interaction between Potato virus X and Potato virus Y and their relationship with symptom expression. Mol. Plant Microbe Interact. 22, 1431–1444. doi: 10.1094/MPMI-22-11-1431
Gest, H. (2004). The discovery of microorganisms by Robert Hooke and Antoni van Leeuwenhoek, fellows of the Royal Society. Notes Rec. R. Soc. Lond. 58, 187–201. doi: 10.1098/rsnr.2004.0055
Gibson, A. K., Raverty, S., Lambourn, D. M., Huggins, J., Magargal, S. L., and Grigg, M. E. (2011). Polyparasitism is associated with increased disease severity in Toxoplasma gondii-Infected marine sentinel Species. PLoS Negl. Trop. Dis. 5:e1142. doi: 10.1371/journal.pntd.0001142
Gibson, R. W., and Kreuze, F. (2014). Degeneration in sweetpotato due to viruses, virus-cleaned planting material and reversion: a review. Plant Pathol. 64, 1–15. doi: 10.1111/ppa.12273
Gilbert, J. A., and Dupont, C. L. (2011). Microbial metagenomics: beyond the genome. Annu. Rev. Mar. Sci. 3, 347–371. doi: 10.1146/annurev-marine-120709-142811
Gleason, M. L., Batzer, J. C., Sun, G. Y., Zhang, R., Arias, M. M. D., Crous, P. W., et al. (2011). A new view of sooty blotch and flyspeck. Plant Dis. 95, 368–383. doi: 10.1094/PDIS-08-10-0590
Glen, M., Bougher, N. L., Francis, A. A., Nigg, N. G., Lee, S. S., Irianto, R., et al. (2009). Ganoderma and Amauroderma species associated with root-rot disease of Acacia mangium plantation trees in Indonesia and Malaysia. Australas. Plant Pathol. 38, 345–356. doi: 10.1071/AP09008
Goodwin, S. B., Sujkowski, L. S., and Fry, W. E. (1995). Rapid evolution of pathogenicity within clonal lineages of the potato late blight disease fungus. Phytopathology 85, 669–676. doi: 10.1094/Phyto-85-669
Goumas, D. E., and Chatzaki, A. K. (1998). Characterization and host range evaluation of Pseudomonas viridiflava from melon, blite, tomato, chrysanthemum and eggplant. Eur. J. Plant Pathol. 104, 181–188. doi: 10.1023/A:1008675704834
Gramaje, D., and Armengol, J. (2011). Fungal trunk pathogens in the grapevine propagation process: potential inoculum sources, detection, identification, and management strategies. Plant Dis. 95, 1040–1055. doi: 10.1094/PDIS-01-11-0025
Green, S., Studholme, D. J., Laue, B. E., Dorati, F., Lovell, H., Arnold, D., et al. (2010). Comparative genome analysis provides insights into the evolution and adaptation of Pseudomonas syringae pv. aesculi on Aesculus hippocastanum. PLoS ONE 5:e10224. doi: 10.1371/journal.pone.0010224
Grube, M., Fürnkranz, M., Zitzenbacher, S., Huss, H., and Berg, G. (2011). Emerging multi-pathogen disease caused by Didymella bryoniae and pathogenic bacteria on Styrian oil pumpkin. Eur. J. Plant Pathol. 131, 539–548. doi: 10.1007/s10658-011-9829-8
Hajri, A., Pothier, J. F., Fischer-Le Saux, M., Bonneau, S., Poussier, S., Boureau, T., et al. (2012). Type three effector gene distribution and sequence analysis provide new insights into the pathogenicity of plant-pathogenic Xanthomonas arboricola. Appl. Environ. Microbiol. 78, 371–384. doi: 10.1128/AEM.06119-11
Harms, P. A., Sorden, S. D., Halbur, P. G., Bolin, S. R., Lager, K. M., Morozov, I., et al. (2001). Experimental reproduction of severe disease in CD/CD pigs concurrently infected with type 2 porcine circovirus and porcine reproductive and respiratory syndrome virus. Vet. Pathol. 38, 528–539. doi: 10.1354/vp.38-5-528
Hosni, T., Moretti, C., Devescovi, G., Suarez-Moreno, Z. R., Fatmi, M. B., Guarnaccia, C., et al. (2011). Sharing of quorum-sensing signals and role of interspecies communities in a bacterial plant disease. ISME J. 5, 1–14. doi: 10.1038/ismej.2011.65
Ichikawa, K., and Aoki, T. (2000). New leaf spot disease of Cymbidium species caused by Fusarium subglutinans and Fusarium proliferatum. J. Gen. Plant Pathol. 66, 213–218. doi: 10.1007/PL00012948
Karyeija, R. F., Kreuze, J. F., Gibson, R. W., and Valkonen, J. P. T. (2000). Synergistic interactions of a Potyvirus and a phloem-limited crinivirus in sweet potato plants. Virology 269, 26–36. doi: 10.1006/viro.1999.0169
Kaur, P., Sivasithamparam, K., Li, H., and Barbetti, M. J. (2011). Pre-inoculation with Hyaloperonospora parasitica reduces incubation period and increases severity of disease caused by Albugo candida in a Brassica juncea variety resistant to downy mildew. J. Gen. Plant Pathol. 77, 101–106. doi: 10.1007/s10327-011-0293-2
Klement, Z., and Lovrekovich, L. (1961). Defense reactions induced by phytopathogenic bacteria in bean pods. Phytopatology 41, 217–227. doi: 10.1111/j.1439-0434.1961.tb04050.x
Kreuze, J. F., Perez, A., Untiveros, M., Quispe, D., Fuentes, S., Barker, I., et al. (2009). Complete viral genome sequence and discovery of novel viruses by deep sequencing of small RNAs: a generic method for diagnosis, discovery and sequencing of viruses. Virology 388, 1–7. doi: 10.1016/j.virol.2009.03.024
Kůdela, V., Krejzar, V., and Pánková, I. (2010). Pseudomonas corrugata and Pseudomonas marginalis associated with the collapse of tomato plants in rockwool slab hydroponic culture. Plant Prot. Sci. 46, 1–11.
Kuzdraliński, A., Szczerba, H., Tofil, K., Filipiak, A., Garbarczyk, E., Dziadko P., et al. (2014). Early PCR-based detection of Fusarium culmorum, F. graminearum, F. sporotrichioides and F. poae on stem bases of winter wheat throughout Poland. Eur. J. Plant Pathol. 140, 491–502. doi: 10.1007/s10658-014-0483-9
Lamprecht, S. C., Tewoldemedhin, Y. T., Botha, W. J., and Calitz, F. J. (2011). Species complex associated with maize crowns and roots in the KwaZulu-Natal Province of South Africa. Plant Dis. 95, 1153–1158. doi: 10.1094/PDIS-02-11-0083
Larignon, P., Fulchic, R., Cere, L., and Dubos, B. (2001). Observation on black dead arm in French vineyards. Phytopathol. Mediterr. 40, S336–S342.
Le, D. P., Smith, M., Hudler, G. W., and Aitken, E. (2014). Detection and identification of the causal pathogens, and their control. Crop Prot. 65, 153–167. doi: 10.1016/j.cropro.2014.07.021
Le May, C., Potage, G., Andrivon, D., Tivoli, B., and Outreman, Y. (2009). Plant disease complex: antagonism and synergism between pathogens of the Ascochyta blight complex on Pea. J. Phytopathol. 157, 715–721. doi: 10.1111/j.1439-0434.2009.01546.x
Li, Y. P., You, M. P., Finnegan, P. M., Khan, T. N., Lanoiselet, V., Eyres, N., et al. (2012). First report of black spot caused by Boeremia exigua var. exigua on field pea in Australia. Plant Dis. 96, 148.3.
Li, Y. P., You, M., Khan, T., Finnegan, P. M., and Barbetti, M. J. (2011). First report of Phoma herbarum on field pea (Pisum sativum) in Australia. Plant Dis. 95, 1590–1590. doi: 10.1094/PDIS-07-11-0594
Llama-Palacios, A., López-Solanilla, E., and Rodríguez-Palenzuela, P. (2002). The ybiT gene of Erwinia chrysanthemi codes for a putative ABC transporter and is involved in competitiveness against endophytic bacteria during infection. Appl. Environ. Microbiol. 68, 1624–1630. doi: 10.1128/AEM.68.4.1624-1630.2002
Lunden, S., Meng, B., Avery, J., and Qiu, W. (2010). Association of Grapevine fanleaf virus, Tomato ringspot virus and Grapevine rupestris stem pitting-associated virus with a grapevine vein-clearing complex on var. Chardonnay. Eur. J. Plant Pathol. 126, 135–144. doi: 10.1007/s10658-009-9527-y
Ma, L., Cao, Y. H., Cheng, M. H., Huang, Y., Mo, M. H., Wang, Y., et al. (2013). Phylogenetic diversity of bacterial endophytes of Panax notoginseng with antagonistic characteristics towards pathogens of root-rot disease complex. Antonie Van Leeuwenhoek 113, 299–312. doi: 10.1007/s10482-012-9810-3
Macho, A. P., Zumaquero, A., Ortiz-Martín, I., and Beuzón, C. R. (2007). Competitive index in mixed infections: a sensitive and accurate assay for the genetic analysis of Pseudomonas syringae-plant interactions. Mol. Plant Pathol. 8, 437–450. doi: 10.1111/j.1364-3703.2007.00404.x
Malathrakis, N. E., and Goumas, D. E. (1987). Bacterial soft rot of tomato in plastic greenhouses in Crete. Ann. Appl. Biol. 111, 115–123. doi: 10.1111/j.1744-7348.1987.tb01438.x
Martin, R. R., MacFarlane, S., Sabandzovic, S., Quito, D., Poucdel, B., and Tzanetakis, I. E. (2013). Viruses and virus diseases of Rubus. Plant Dis. 97, 168–182. doi: 10.1094/PDIS-04-12-0362-FE
Mazzaglia, A., Studholme, D. J., Taratufolo, M. C., Cai, R., Almeida, N. F., Goodman, T., et al. (2012). Pseudomonas syringae pv. actinidiae (PSA) isolates from recent bacterial canker of kiwifruit outbreaks belong to the same genetic lineage. PLoS ONE 7:e36518. doi: 10.1371/journal.pone.0036518
Mazzola, M. (1998). Elucidation of the microbial complex having a causal role in the development of apple replant disease in Washington. Phytopathology 88, 930–938. doi: 10.1094/PHYTO.1998.88.9.930
Mbega, E. R., Mabagala, R. B., Adriko, J., Lund, O. S., Wulff, E. G., and Mortensen, C. N. (2012). Five species of Xanthomonads associated with bacterial leaf spot symptoms in tomato from Tanzania. Plant Dis. 96, 760.2. doi: 10.1094/PDIS-01-12-0105-PDN
McMullen, M. P., Bergstrom, G. C., De Wolf, E., Dill-Macky, R., Hershman, D., Shaner, G., et al. (2012). A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 96, 1712–1728. doi: 10.1094/PDIS-03-12-0291-FE
Miao, Z. Q., Li, S. D., Liu, X. Z., Chen, Y., Li, Y., Wang, Y., et al. (2006). The causal microorganisms of Panax notoginseng root rot disease. Sci. Agric. Sinica 39, 1371–1378.
Moura, M. L., Jacques, L. A., Brito, L. M., Mourao, I. M., and Duclos, J. (2005). Tomato pith necrosis caused by P-corrugata and P-mediterranea: severity of damages and crop loss assessment. Acta Hort. 695, 365–372.
Mugnai, L., Graniti, A., and Surico, G. (1999). Esca (black measles) and brown wood-streaking: two old and elusive diseases of grapevines. Plant Dis. 83, 404–418. doi: 10.1094/PDIS.1999.83.5.404
Murphy, J. F., and Bowen, K. L. (2006). Synergistic disease in pepper caused by the mixed infection of Cucumber mosaic virus and Pepper mottle virus. Phytopathology 96, 240–247. doi: 10.1094/PHYTO-96-0240
Naidu, R., Rowhani, A., Fuchs, M., Golino, D., and Martelli, G. P. (2014). Grapevine leafroll: a complex viral disease affecting a high value fruit crops. Plant Dis. 98, 1172–1185. doi: 10.1094/PDIS-08-13-0880-FE
Nguyen, P. T. H., Pettersson, O. V., Olsson, P., and Liljeroth, E. (2010). Identification of Colletotrichum species associated with anthracnose disease of coffee in Vietnam. Eur. J. Plant Pathol. 127, 73–87. doi: 10.1007/s10658-009-9573-5
Niblett, C. L., and Claflin, L. E. (1978). Corn lethal necrosis-A new virus disease of corn in Kansas. Plant Dis. Rep. 62, 15–19.
Nikolaki, S., and Tsiamis, G. (2013). Microbial diversity in the era of omic technologies. BioMed. Res. Int. 2013, 958719. doi: 10.1155/2013/958719
O’Donnell, K., Ward, T. J., Aberra, D., Kistler, H. C., Aoki, T., Orwig, N., et al. (2008). Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fung. Genet. Biol. 45, 1514–1522. doi: 10.1016/j.fgb.2008.09.002
Olatinwo, R. O., Hanson, E. J., and Schilder, A. M. C. (2003). A first assessment of the cranberry fruit rot complex in Michigan. Plant Dis. 87, 550–556. doi: 10.1094/PDIS.2003.87.5.550
Oliver, J. D. (2010). Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 34, 415–425.
Omer, M. E. H., and Wood, R. K. S. (1969). Growth of Pseudomonas phaseolicola in susceptible and resistant bean plants. Ann. Appl. Biol. 63, 103–116. doi: 10.1111/j.1744-7348.1969.tb05471.x
Oudemans, P. V., Caruso, F. L., and Stretch, A. W. (1998). Cranberry fruit rot in the Northeast: a complex disease. Plant Dis. 82, 1176–1184. doi: 10.1094/PDIS.1998.82.11.1176
Papavasileiou, A., Testempasis, S., Michailides, T. J., and Karaoglanidis, G. S. (2014). Frequency of brown rot fungi on blossoms and fruit in stone fruit orchards in Greece. Plant Pathol. 64, 416–424. doi: 10.1111/ppa.12264
Pemberton, I. J., Smith, G. R., Philley, G. L., Rouquette, F. M., and Yuen, G. Y. (1998). First Report of Pythium ultimum, P. irregulare, Rhizoctonia solani AG4, and Fusarium proliferatum from Arrowleaf Clover (Trifolium vesiculosum): a disease complex. Plant Dis. 82, 128.2.
Peters, B. M., Jabra-Rizk, M. A., O’May, G. A., Costerton, J. W., and Shirtliff, M. E. (2012). Polymicrobial interactions: impact on pathogenesis and human disease. Clin. Microbiol. Rev. 25, 193–213. doi: 10.1128/CMR.00013-11
Petkowski, J. E., de Boer, R. F., Norng, S., Thomson, F., and Minchinton, E. J. (2013). Pythium species associated with root rot complex in winter-grown parsnip and parsley crops in south eastern Australia. Australas. Plant Pathol. 42, 403–411. doi: 10.1007/s13313-013-0211-5
Pio-Ribeiro, G., Wyatt, S. D., and Kuhn, C. W. (1978). Cowpea stunt: a disease caused by a synergistic interaction of two viruses. Phytopathology 68, 1260–1265. doi: 10.1094/Phyto-68-1260
Purcell, A. (2013). Paradigms: examples from the bacterium Xylella fastidiosa. Annu. Rev. Phytopathol. 51, 339–356. doi: 10.1146/annurev-phyto-082712-102325
Ramsey, M. D. (1990). Etiology of root and stalk rots of maize in north Queensland. Disease development and associated fungi. Australas. Plant Pathol. 19, 2–12. doi: 10.1071/APP9900002
Roberts, F. A., and Sivasithamparam, K. (1986). Identity and pathogenicity of Rhizoctonia spp. associated with bare patch disease of cereals at a field site in Western Australia. Neth. J. Plant Pathol. 92, 185–195. doi: 10.1007/BF01977685
Sagar, V., and Sugha, S. K. (1997). Role of individual and combined inocula on the development of pea root rot. Indian Phytopathol. 50, 499–503.
Salvaudon, L., De Moraes, C. M., and Mesche, M. C. (2013). Outcomes of co-infection by two potyviruses: implications for the evolution of manipulative strategies. Proc. Biol. Soc. 280, 20122959. doi: 10.1098/rspb.2012.2959
Sampietro, D. A., Díaz, C. G., Gonzalez, V., Vattuone, M. A., Ploper, L. D., Catalan, C. A., et al. (2011). Species diversity and toxigenic potential of Fusarium graminearum complex isolates from maize fields in northwest Argentina. Int. J. Food Microbiol. 145, 359–364. doi: 10.1016/j.ijfoodmicro.2010.12.021
Sarver, B. A. J., Ward, T. J., Gale, L. R., Broz, K., Kistler, H. C., Nicholson, P., et al. (2011). Novel Fusarium head blight pathogens from Nepal and Louisiana revealed by multilocus genealogical concordance. Fung. Genet. Biol. 48, 1096–1107. doi: 10.1016/j.fgb.2011.09.002
Sawant, I. S., Narkar, S. P., Shetty, D. S., Upadhyay, A., and Sawant, S. D. (2012). Emergence of Colletotrichum gloeosporioides sensu lato as the dominant pathogen of anthracnose disease of grapes in India as evidenced by cultural, morphological and molecular data. Australas. Plant Pathol. 41, 493–504. doi: 10.1007/s13313-012-0143-5
Saygili, H., Aysan, Y., Ustun, N., Mirik, M., and Sahin, F. (2008). “Tomato pith necrosis disease caused by Pseudomonas species in Turkey,” in Pseudomonas Syringae Pathovars and Related Pathogens Identification, Epidemiology, and Genomics, eds M. B. Fatmi, A. Collmer, and N.S. Iacobellis (Berlin: Springer), 357–366.
Scarlett, C. M., Fletcher, J. T., Roberts, P., and Lelliott, R. A. (1978). Tomato pith necrosis caused by Pseudomonas corrugata n. sp. Anna. Appl. Biol. 88, 105–114. doi: 10.1111/j.1744-7348.1978.tb00684.x
Scheets, K. (1998). Maize chlorotic mottle machlomovirus and wheat streak mosaic rymovirus concentrations increase in the synergistic disease corn lethal necrosis. Virology 242, 28–38. doi: 10.1006/viro.1997.8989
Short, F. L., Murdoch, S. L., and Ryan, R. P. (2014). Polybacterial human disease: the ills of social networking. Trends Microbiol. 22, 508–516. doi: 10.1016/j.tim.2014.05.007
Strausbaugh, C. A., and Eujayl, I. A. (2012). Influence of sugarbeet tillage systems on the Rhizoctonia-bacterial root rot complex. J. Sugar Beet Res. 49, 57–78. doi: 10.5274/jsbr.49.3.57
Strausbaugh, C. A., and Gillen, A. M. (2008). Bacteria and yeast associated with sugar beet root rot at harvest in the intermountain west. Plant Dis. 92, 357–363. doi: 10.1094/PDIS-92-3-0357
Studholme, D. J., Glover, R. H., and Boonham, N. (2011). Application of high-throughput DNA sequencing in phytopathology. Annu. Rev. Phytopathol. 49, 87–105. doi: 10.1146/annurev-phyto-072910-095408
Suheri, H., and Price, T. V. (2001). The epidemiology of purple leaf blotch on leeks in Victoria, Australia. Eur. J. Plant Pathol. 107, 503–510. doi: 10.1023/A:1011244104920
Syller, J. (2012). Facilitative and antagonistic interactions between plant viruses in mixed infections. Mol. Plant Pathol. 13, 204–216. doi: 10.1111/j.1364-3703.2011.00734.x
Tairo, F., Jones, R. A. C., and Valkonen, J. P. T. (2006). Potyvirus complexes in sweet potato: occurrence in Australia, serological and molecular resolution, and analysis of the Sweet potato virus 2 (SPV2) component. Plant Dis. 90, 1120–1128. doi: 10.1094/PD-90-1120
Tantaoui, A., Ouinten, M., Geiger, J.-P., and Fernandez, D. (1996). Characterization of a single clonal lineage of Fusarium oxysporum f. sp. albedinis causing Bayoud disease of date palm in Morocco. Phytopathology 86, 787–792. doi: 10.1094/Phyto-86-787
Tatineni, S., Graybosch, R. A., Hein, G. L., Wegulo, S. N., and French, R. (2010). Wheat cultivar-specific disease synergism and alteration of virus accumulation during co-infection with Wheat streak mosaic virus and Triticum mosaic virus. Phytopathology 100, 230–238. doi: 10.1094/PHYTO-100-3-0230
Tomioka, K., Nishikawa, J., Moriwaki, J., and Sato, T. (2013). Anthracnose of Madagascar periwinkle caused by species belonging to the Colletotrichum gloeosporioides species complex. J. Gen. Plant Pathol. 79, 374–377. doi: 10.1007/s10327-013-0469-z
Tran, H. S., You, M., Li, Y., Khan, T. N., and Barbetti, M. J. (2014). First report of Phoma glomerata on field pea (Pisum sativum) in Australia. Plant Dis. 98, 427. doi: 10.1094/PDIS-08-13-0809-PDN
Tringe, S. G., and Hugenholtz, P. (2008). A renaissance for the pioneering 16S rRNA gene. Curr. Opin. Microbiol. 11, 442–446. doi: 10.1016/j.mib.2008.09.011
van Dijk, E. L., Auger, H., Jaszczyszyn, Y., and Thermes, C. (2014). Ten years of next-generation sequencing technology. Trends Gentet. 30, 418–426. doi: 10.1016/j.tig.2014.07.001
van Niekerk, J. M., Fourie, P. H., Halleen, F., and Crous, P. W. (2006). Botryosphaeria spp. as grapevine trunk disease pathogens. Phytopathol. Mediterr. 45, S43–S54.
Venter, J. C., Remington, K., Heidelberg, J. F., Halpern, A. L., Rusch, D., and Eisen, J. A. (2004). Environmental genome shotgun sequencing of the Sargasso Sea. Science 304, 66–74. doi: 10.1126/science.1093857
Vettraino, A. M., Morel, O., Perlerou, C., Robin, C., Diamandis, S., and Vannini, A. (2005). Occurrence and distribution of Phytophthora species in European chestnut stands, and their association with Ink Disease and crown decline. Eur. J. Plant Pathol. 111, 169–180. doi: 10.1007/s10658-004-1882-0
Vitullo, D., De Curtis, F., Palmieri, D., and Lima, G. (2014). Milkwort (Polygala myrtifolia L.) decline is caused by Fusarium oxysporum and F. solani in Southern Italy. Eur. J. Plant Pathol. doi: 10.1007/s10658-014-0514-6
Voigt, K., Schleier, S., and Bruckner, B. (1995). Genetic variability in Gibberella fujikuroi and some related species of the genus Fusarium based on random amplification of poly-morphic DNA (RAPD). Curr. Genet. 27, 528–535. doi: 10.1007/BF00314443
Wang, G.-F., Xie, G.-L., Zhu, B., Huang, J. S., Liu, B., Kawicha, P., et al. (2010). Identification and characterization of the Enterobacter complex causing mulberry (Morus alba) wilt disease in China. Eur. J. Plant Pathol. 126, 465–478. doi: 10.1007/s10658-009-9552-x
Wang, Y., Gaba, V., Yang, J., Palukaitis, P., and Gal-On, A. (2002). Characterization of synergy between Cucumber mosaic virus and Potyviruses in cucurbit hosts. Phytopathology 92, 51–58. doi: 10.1094/PHYTO.2002.92.1.51
Watson, M. T., Tian, T., Estabrook, E., and Falk, B. W. (1998). A small RNA resembling the beet western yellows luteovirus ST9-associated RNA is a component of the California Carrot motley dwarf complex. Phytopathology 88, 164–170. doi: 10.1094/PHYTO.1998.88.2.164
Wen, A., Mallik, I., Alvarado, V. Y., Pasche, J. S., Wang, X., Li, W., et al. (2009). Detection, distribution, and genetic variability of “Candidatus liberibacter” species associated with Zebra complex disease of potato in North America. Plant Dis. 93, 1102–1115. doi: 10.1094/PDIS-93-11-1102
Whitelaw-Weckert, M. A., Rahman, L., Appleby, L. M., Hall, A., Clark, A. C., Waite, H., et al. (2013). Co-infection by Botryosphaeriaceae and Ilyonectria spp. fungi during propagation causes decline of young grafted grapevines. Plant Pathol. 62, 1226–1237. doi: 10.1111/ppa.12059
Wilkie, J. P., and Dye, D. W. (1974). Pseudomonas cichorii causing tomato and celery diseases in New Zealand. N. Z. J. Agric. Res. 17, 123–130. doi: 10.1080/00288233.1974.10420990
Willocquet, L., Aubertot, J. N., Lebard, S., Robert, C., Lannou, C., and Savary, S. (2008). Simulating multiple pest damage in varying winter wheat production situations. Field Crops Res. 107, 12–28. doi: 10.1016/j.fcr.2007.12.013
Willocquet, L., Savary, S., Fernandez, L., Elazegui, F. A., Castilla, N., Zhu, D., et al. (2002). Structure and validation of RICEPEST, a production situation-driven, crop growth model simulating rice yield response to multiple pest injuries for tropical Asia. Ecol. Model. 153, 247–268. doi: 10.1016/S0304-3800(02)00014-5
Wilson, J. P. (2002). Fungi associated with the stalk rot complex of pearl millet. Plant Dis. 86, 833–839. doi: 10.1094/PDIS.2002.86.8.833
Wing, K. B., Pritts, M. P., and Wilcox, W. F. (1994). Strawberry black root rot: a review. Adv. Strawberry Res. 13, 13–19.
Yli-Mattila, T., Gagkaeva, T., Ward, T. J., Aoki, T., Kistler, H. C., and O’Donnell, K. (2009). A novel Asian clade within the Fusarium graminearum species complex includes a newly discovered cereal head blight pathogen from the Far East of Russia. Mycologia 101, 841–852. doi: 10.3852/08-217
Keywords: microbial cooperation, co-infection, synergism, interspecies interactions, disease complex
Citation: Lamichhane JR and Venturi V (2015) Synergisms between microbial pathogens in plant disease complexes: a growing trend. Front. Plant Sci. 6:385. doi: 10.3389/fpls.2015.00385
Received: 04 March 2015; Accepted: 13 May 2015;
Published online: 27 May 2015
Edited by:
Simone Ferrari, Sapienza Università di Roma, ItalyReviewed by:
Johann Weber, University of Lausanne, SwitzerlandGabor Jakab, University of Pécs, Hungary
Copyright © 2015 Lamichhane and Venturi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jay Ram Lamichhane, INRA, UAR 1240 Eco-Innov, BP 01, 78850 Thiverval-Grignon, France,amF5cmFtLmxhbWljaGhhbmVAZ21haWwuY29t