 Sona Fekecsová1,2
Sona Fekecsová1,2 Maksym Danchenko
Maksym Danchenko Lubica Uvackova
Lubica Uvackova Ludovit Skultety
Ludovit Skultety Martin Hajduch
Martin Hajduch- 1Department of Developmental and Reproduction Biology, Institute of Plant Genetics and Biotechnology, Slovak Academy of Sciences, Nitra, Slovakia
- 2Faculty of Natural Sciences, Comenius University, Bratislava, Slovakia
- 3Institute of Virology, Slovak Academy of Sciences, Bratislava, Slovakia
The aim of the work was to test a relatively simple proteomics approach based on phenol extraction and two-dimensional gel electrophoresis (2-DE) with 7 cm immobilized pH gradient strips for the determination of clinically relevant proteins in wheat grain. Using this approach, 157 2-DE spots were quantified in biological triplicate, out of which 55 were identified by matrix-assisted laser desorption/ionization – time of flight tandem mass spectrometry. Clinically relevant proteins associated with celiac disease, wheat dependent exercise induced anaphylaxis, baker’s asthma, and food allergy, were detected in 24 2-DE spots. However, alcohol-soluble gliadins were not detected with this approach. The comparison with a recent quantitative study suggested that gel-based and gel-free proteomics approaches are complementary for the detection and quantification of clinically relevant proteins in wheat grain.
Introduction
The main component of the wheat grain are storage proteins with gluten as the major part representing as much as 80% of total protein content (Ferranti et al., 2007). Gluten is a mixture of gliadins and glutenins that differ in their electrophoretic mobility (Payne et al., 1985; Jacobsen et al., 2007). Gluten is also the main allergen in the wheat grain and is responsible for nutritive intolerances such as celiac disease or gluten-sensitive enteropathy (Rubio-Tapia et al., 2009), and various allergies (Battais et al., 2008; Sapone et al., 2012; Mauro Martin et al., 2014). In addition to storage proteins, wheat grain allergens include enzymatic and structural proteins such as prolamins, cupins, and Bet v1 protein family (Breiteneder and Mills, 2005). Out of these, prolamins are dominant and include α-amylase and protease inhibitors, 2S albumins, and non-specific lipid transfer proteins (nsLTPs; Breiteneder and Radauer, 2004; Mills et al., 2004).
Protein two-dimensional gel electrophoresis (2-DE) has been extensively used to characterize wheat grain proteins. For instance, 2-DE followed with immunoblotting and tandem mass spectrometry (MS/MS) resulted into the identification of nine subunits of low molecular weight (LMW) glutenins, serpin, α-amylase inhibitor, and α-gliadin in wheat flour (Akagawa et al., 2007). The combination of 2-DE and MS/MS identified several allergenic proteins, such as serpins, in dough liquor of four wheat cultivars under abiotic stress (Sancho et al., 2008). Similar approaches detected heat responsible allergenic proteins, such as α-amylase inhibitors or serpins, in the endosperm of developing wheat grains under heat stress (Hurkman et al., 2009). Additionally, several allergenic proteins were detected in Korean sprouting wheat cultivars using 2-DE and matrix assisted laser desorption/ionization-tandem Time of Flight (MALDI-TOF) MS/MS (Kamal et al., 2009). Importantly, 20 allergenic proteins in wheat grains were detected using proteomics approach based on 2-DE in combination with 17 cm immobilized pH gradient (IPG) strips and MS/MS (Yang et al., 2011). Similarly, 2-DE in combination with isoelectric focusing (IEF) capillary tube gels, three different proteases, and MS/MS resulted in the detection of 476 2-DE spots out of which 233 were identified, including well-known allergens (Dupont et al., 2011). The 2-DE was also used to analyze wheat with genetically altered omega-5 gliadin content (Altenbach et al., 2014). Interestingly, this study showed that unique genetic transformation events with the same RNA interference construct may have differential effects on the wheat grain proteome (Altenbach et al., 2014). This study highlights the importance of proteomic analyses in the study of genetic transformations (Altenbach et al., 2014).
The above studies showed that a 2-DE approach is effective in the characterization of wheat grain proteins. However, 2-DE can be labor, resources, and time consuming, especially when long IPG strips are used. The aim of this study was to test a relatively simple 2-DE approach based on 7 cm IPG strips for the detection of clinically relevant proteins of wheat grain.
Materials and Methods
Plant Material and Protein Extraction
Seeds of winter wheat cultivar Viginta were obtained from SELEKT LtD, Bučany, Slovak Republic. Proteins were extracted in biological triplicate from 500 mg of dry seeds. Seeds were ground in liquid nitrogen and proteins were extracted with phenol-based extraction media [50% (v/v) phenol, 0.45 M sucrose, 5 mM EDTA, 0.2% (v/v) 2-mercaptoethanol, 50 mM Tris–HCl, pH 8.8]. Sample was stirred and homogenized for 30 min at 4°C. The phenol phase was removed after centrifugation at 5000 × g for 10 min at 4°C. Proteins were precipitated from the phenol phase by the addition of five volumes of ice-cold 0.1 M ammonium acetate in 100% methanol, and incubated at -20°C overnight. The protein pellet was extensively washed twice using 0.1 M ice cold ammonium acetate in 100% methanol, followed by 80% ice cold acetone, and finally with 70% ice cold ethanol and precipitates were collected by centrifugation for 15 min., 5000 ×g at 4°C. Total protein concentration was determined using the Bradford (1976) assay with Bovine Serum Albumin as the standard.
Two-Dimensional Gel Electrophoresis
Samples (50 μg protein) were diluted in 100 μl of IEF buffer [8 M urea, 2 M thiourea, 2% (w/v) CHAPS, 2% (v/v) Triton X-100, 50 mM dithiothreitol], 3 μl of ampholytes were added, and loaded onto 7 cm IPG strips of pH 3–10 (ReadyStripTM IPG Strips BioRad) for IEF. Isoelectric focusing was carried out using Protean IEF Cell (Bio-Rad) with the following conditions: 150 V for 150 VH, 500 V for 500 VH, and 4000 V for 15,000 VH including initial active rehydration for 12 h at 50 V. For the second dimension (SDS-PAGE), IPG strips were incubated in SDS equilibration buffer [1.5 M Tris-HCl pH 6.8, 6 M urea, 30% (v/v) glycerol, 5% (w/v) SDS) for 15 min with 2% (w/v) dithiothreitol] followed by a second equilibration step of 15 min with the equilibration buffer containing 2.5% (w/v) iodoacetamide. The equilibrated strips were loaded on the top of 10% polyacrylamide gel and the electrophoresis was run at 80 V until the dye reached the bottom of the gel. Gels were stained for 16 h with Coomassie Brilliant Blue G-250 at room temperature. The 2-DE gels were digitalized using a GS-800 Calibrated Densitometer (Bio-Rad) at 300 dpi and 16 bit grayscale. Digitalized gels were analyzed with PDQuest 8.0 software (Bio-Rad).
Protein Digestion and Mass Spectrometry
Excised 2-DE plugs were washed with 300 μl destaining solution (50% acetonitrile in 50 mM ammonium bicarbonate) and dehydrated in 100% acetonitrile. After removal of acetonitrile, 2-DE spots were rehydrated with trypsin (Promega) and digested at 37°C overnight. The digestion was stopped with formic acid and extracted tryptic peptides were stored at -80°C until MS/MS analysis with a TOF/TOF mass spectrometer in combination with MALDI using an ultrafleXtreme instrument equipped with a 355 nm smartbeam-2 laser, capable of pulsing frequency 1 kHz (Bruker). Peptides were concentrated to 20 μl using Concentrator plus (Eppendorf). After that, concentrated peptides were desalted by μ-C18 ZipTips (Merck Millipore). Next, 1 μl of purified digests were spotted onto 800 μm AnchorChip MALDI target (Bruker) and α-cyano-4-hydroxycinnamic acid (CHCA) matrix (0.7 mg⋅ml-1 in 85% acetonitrile, 0.1% trifluoroacetic acid, 1 mM ammonium phosphate) was added.
The mass spectrometer was operated by flexControl 3.3 software (Bruker). For every position 4000 shots were summed in positive reflector mode in the range of 700–3500 mass to charge (m/z). Following that, up to 25 of the most intense precursor peaks per sample were selected for the MS/MS analysis with the minimal signal to noise (S/N) ratio set to 15. Abundant trypsin and keratin peaks were specified in the exclusion list. Fragmentation spectra were acquired by accumulation of 3000 laser shots in positive reflector LIFT mode. Fragmentation was achieved by laser induced dissociation (LID) mechanism by 50% increase in laser power, without the introduction of a collision gas. Simultaneously detector voltage was boosted by 80%.
Processing of MS/MS Data
Acquired spectra were processed by flexAnalysis 3.3 software (Bruker). A sophisticated numerical annotation procedure (SNAP) algorithm was used for peak picking to calculate exact monoisotopic masses. For the precursor spectra the S/N threshold was set to 10 and the resulting spectra were externally recalibrated against data from an adjacent spot containing nine peptides of the Peptide calibration standard 2 (Bruker). For the fragment spectra S/N threshold was set to 5, also baseline subtraction (TopHat algorithm), and smoothing (Savitzky-Golay algorithm 3 cycles with 0.15 m/z width) were applied.
The MS/MS peak lists were imported into the ProteinScape 2.1 proteomic data management software (Bruker). Peptide identification was performed by an in-house Mascot 2.3 server (Matrix Science), querying against the non-redundant Triticeae plant protein UniProt database downloaded on April, 2014 (100 981 entries). Additionally, protein assignments were verified by searches against the SwissProt database from June 2014 (545 388 sequences) that included major contaminants such as trypsin or keratin. Search parameters were the following: fixed cysteine carbamidomethylation, variable methionine oxidation, one missed trypsin cleavage site, 40 ppm precursor mass tolerance, 0.5 Da fragment mass tolerance. Protein identifications were accepted if at least two different matched peptides had ion score higher than 30, meaning p < 0.05.
For allergenicity assessments, identified proteins were queried against the Allergome database1 containing 2994 allergen entries, using Allergome Aligner module with an embedded NCBI blastp v.2.2.18 algorithm. Only hits with 100% sequence identity were accepted as clinically relevant allergens. The MS/MS proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD002067.
Results
The 2-DE-based proteomics approach in combination with 7 cm IPG strips (Figure 1) quantified 157 2-DE spots in biological triplicate (Supplementary Figure S1; Supplementary Table S1) out of which 55 were identified (Table 1). Identified proteins were classified according to previous apporaches (Bevan et al., 1998) into five functional classes (Figure 2). The most abundant class was 37 proteins associated with destination and storage, followed by nine proteins associated with metabolism and five energy proteins (Figure 2). This study also detected two signaling proteins and two proteins associated with disease/defense (Table 1). All identified proteins were assigned on the 2-DE gel (Supplementary Figure S2). The most abundant protein on this reference map is the high molecular weight (HMW) glutenin subunit (GS) with a relative volume (%V) of 8.8 (spot number 2909) followed by 11-S seed storage domain containing protein (3302) with %V of 5.6 (Table 1).
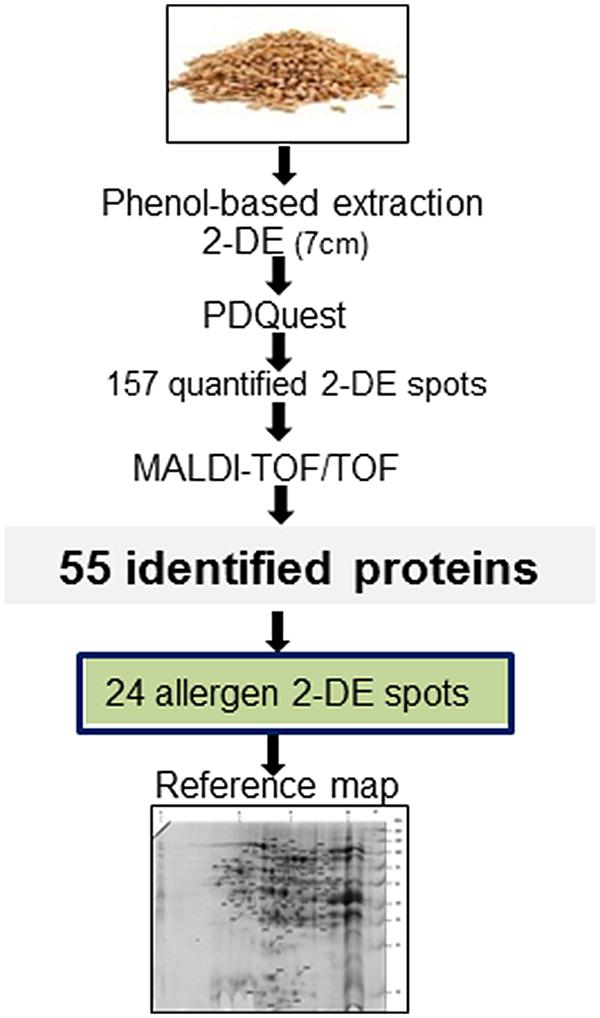
FIGURE 1. Experimental workflow. Proteins were isolated from the winter wheat variety Viginta using a phenol-based method and resolved by 7 cm immobilized pH gradient (IPG) strip. After analysis with PDQuest, two-dimensional gel electrophoresis (2-DE) spots were excised from the gels, and analyzed by MALDI-TOF/TOF. From 55 identified 2-DE spots, 24 contained clinically relevant proteins. These proteins were assigned onto a 2-DE reference map.
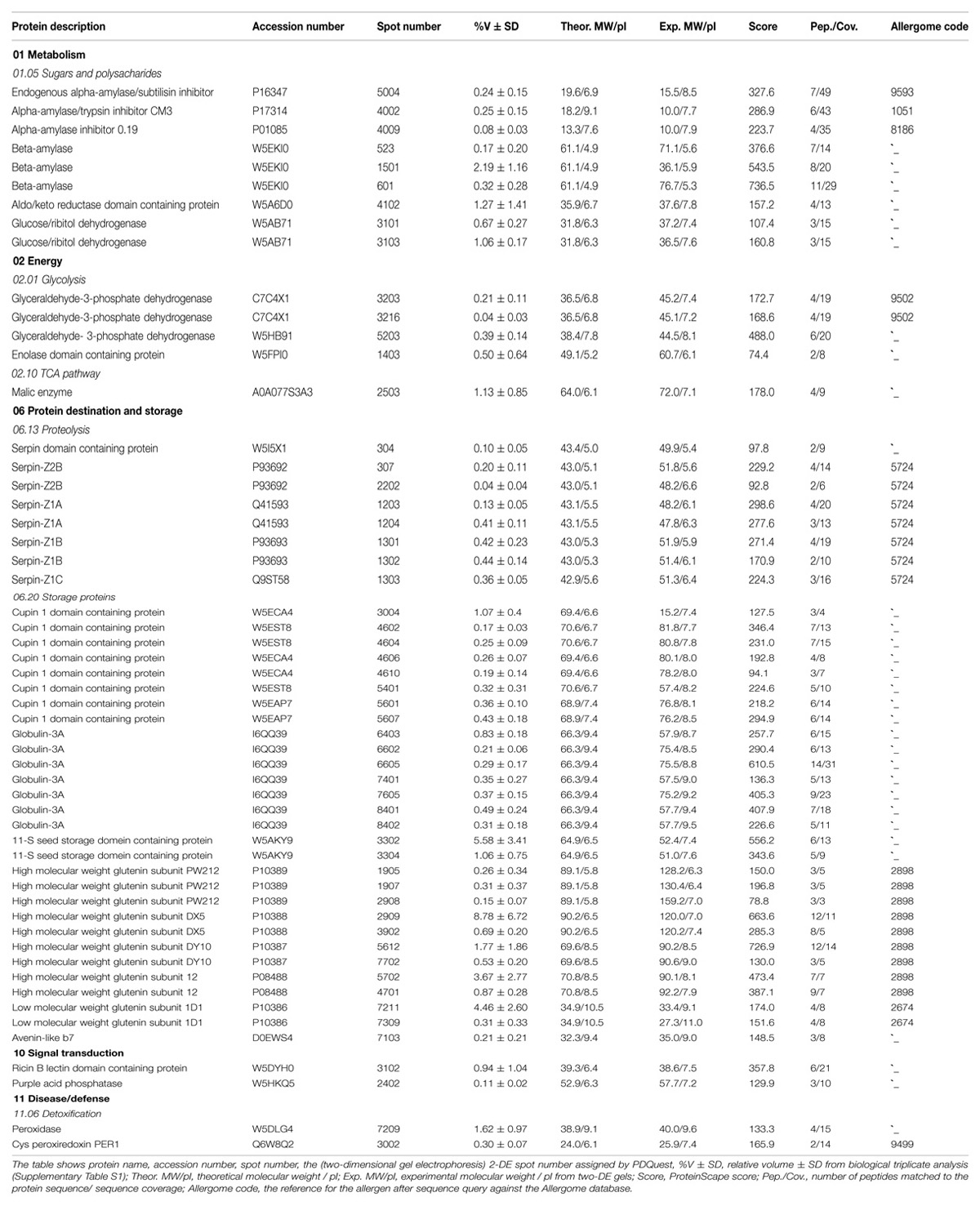
TABLE 1. The list of 55 identified and quantified proteins in wheat grain extracts.
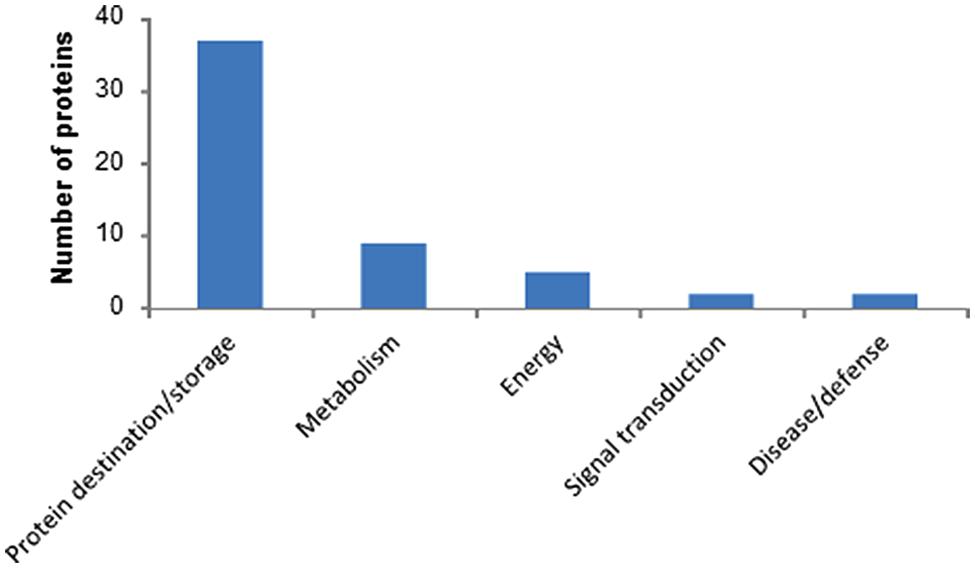
FIGURE 2. The 55 identified wheat grain proteins were classified into five functional categories.
As Much as 45% of Identified Proteins were Associated with Various Allergies or Food Intolerances
To determine clinical relevance of the identified proteins, sequences were queried against the Allergome database1 which contains 2994 allergen entries (Supplementary Table S2). This approach detected clinically relevant proteins in 24 2-DE spots which represented 13 non-redundant accession numbers (Table 1). Out of these, nine 2-DE spots were identified as HMW GS, seven as serpins, three as α-amylase inhibitors, two as LMW GS, two as glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and one as Cys peroxiredoxin (PER1; Table 1). All these proteins were assigned on the 2-DE gel in order to establish the reference map of clinically relevant proteins of wheat grain (Figure 3). All 24 2-DE spots presented on this reference map were color-coded based on protein identification to visualize regions of the 2-DE gel with a prevalence of clinically relevant proteins (Figure 3). The most abundant protein is HMW GS (2-DE spot number 2909) followed LMW GS (7211; Table 1). The 2-DE spots identified as serpin (2202), GAPDH (3216), GAPDH, and α-amylase inhibitor (4009) (3203) showed the lowest abundance on this reference map (Figure 3; Table 1). To reveal overall abundances of the detected clinically relevant proteins, relative volumes of each 2-DE spot were summed based on protein identifications. Using this approach, the summed (total) relative volumes for each of eight detected clinically relevant proteins was established (Figure 4). It was revealed that HMW GS, LMW GS, and serpins are highly abundant in wheat grain (Figure 4).

FIGURE 3. The 2-DE reference map of 24 clinically relevant proteins representing 13 unique accession numbers (six proteins).
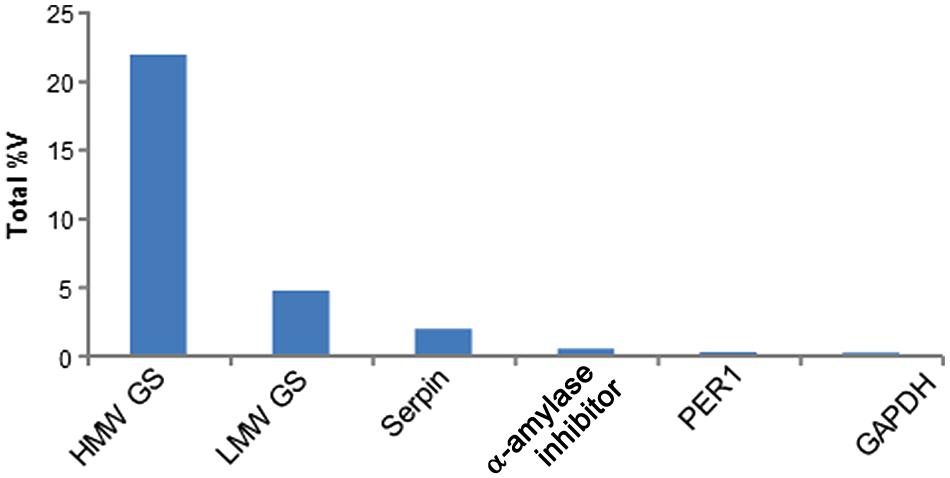
FIGURE 4. Summed abundance of the six non-redundant allergenic proteins detected in wheat grain extracts. The total abundance is shown as relative volume (%V).
Discussion
The aim of this work was to test a 2-DE proteomics approach with 7 cm IPG strips and phenol-based protein extraction for the detection of clinically relevant proteins in wheat grain. Classical methods for protein isolation from wheat grain are based on iso-propanol extraction (van den Broeck et al., 2009). We successfully implemented this method and have previously determined quantities of wheat grain proteins using gel-free proteomics approach (Uvackova et al., 2013a,b). In the present study we tested the phenol-based extraction protocol (Hurkman and Tanaka, 1986), which also solubilizes membrane proteins often excluded from alcohol-based protein extractions. Previously, our group efficiently used this protocol for the characterization of seed proteins in soybean (Hajduch et al., 2005; Danchenko et al., 2009; Klubicova et al., 2012), canola (Hajduch et al., 2006), castor (Houston et al., 2009), Arabidopsis (Hajduch et al., 2010), and flax (Klubicova et al., 2010, 2013).
In the present study we detected nine 2-DE spots as HMW GS (Table 1; Figure 3), which influence the viscoelastic properties of wheat flour (Masci et al., 1998), and may cause wheat dependent exercise-induced anaphylaxis (WDEIA) when digested (Hofmann et al., 2012). The present study was particularly successful in the detection of wheat grain allergens associated with Baker’s asthma (Salcedo et al., 2011; Olivieri et al., 2013). Five 2-DE spots were identified as serpin (Table 1; Figure 3), which are involved in food allergy and Baker’s asthma (Salcedo et al., 2011; Mameri et al., 2012). Three 2-DE spots were identified as an α-amylase inhibitor, important contributors to Baker’s asthma (Tatham and Shewry, 2008; Salcedo et al., 2011), food allergies (James et al., 1997), and WDEIA (Hofmann et al., 2012). Additionally, one 2-DE spot was detected as PER1 which is a confirmed wheat allergen likely associated with Baker’s asthma (Pahr et al., 2012). However, this study did not detect the 27 kDa albumin, which was shown to be associated with Baker’s asthma (Weiss et al., 1993) or the alcohol-soluble gliadin proteins involved in celiac disease (Wieser, 1996; Allred and Ritter, 2010).
The majority of wheat grain allergenic proteins detected in the present study were not quantified in our recent MS-based study (Uvackova et al., 2013b). This finding is in agreement with a recent investigation of soybean under flooding stress, where only 9 out of 115 proteins were detected by both gel-based and gel-free proteomics approaches in root tips (Yin et al., 2014). Similar results have been shown in the analysis of the honey bees hemolymph proteome, where only 27% of proteins were detected with both approaches (Bogaerts et al., 2009).
Based on this, it is tempting to speculate that gel-based and gel-free approaches are complementary for the detection and quantification of wheat grain allergenic proteins. However, the complementarity of gel-based and gel-free proteomics approaches was suggested previously (Luque-Garcia et al., 2011; Abdallah et al., 2012). The combination of gel-based and gel-free proteomics was shown to be effective for the analyses of soybean under flooding (Yin et al., 2014), phytopathogenic fungus Botrytis cinerea (Gonzalez-Fernandez et al., 2013), Nicotiana tabacum trichomes (Van Cutsem et al., 2011), the honeybee hemolymph proteome (Bogaerts et al., 2009), or during soybean seed filling (Agrawal et al., 2008).
Conclusion
This study has demonstrated that phenol-based protein extraction in combination with 2-DE and 7 cm IPG strips is capable of determining clinically relevant proteins in wheat grain extracts. However, important clinically relevant proteins, such as alcohol-soluble gliadins were not detected with this approach. The comparison of these data with previous work suggests that gel-based and gel-free proteomics are complementary approaches for the determination of clinically relevant proteins in wheat grain extracts.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was supported by the Scientific Grant Agency of the Ministry of Education of the Slovak Republic and Slovak Academy of Sciences (VEGA-2/0016/14) and European Community under project no 26220220180: Building Research Centre “AgroBioTech.”
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2015.00433/abstract
Footnotes
References
Abdallah, C., Dumas-Gaudot, E., Renaut, J., and Sergeant, K. (2012). Gel-based and gel-free quantitative proteomics approaches at a glance. Int. J. Plant Genomics 2012, 494572. doi: 10.1155/2012/494572
Agrawal, G. K., Hajduch, M., Graham, K., and Thelen, J. J. (2008). In-depth investigation of the soybean seed-filling proteome and comparison with a parallel study of rapeseed. Plant Physiol. 148, 504–518. doi: 10.1104/pp.108.119222
Akagawa, M., Handoyo, T., Ishii, T., Kumazawa, S., Morita, N., and Suyama, K. (2007). Proteomic analysis of wheat flour allergens. J. Agric. Food Chem. 55, 6863–6870. doi: 10.1021/jf070843a
Allred, L. K., and Ritter, B. W. (2010). Recognition of gliadin and glutenin fractions in four commercial gluten assays. J. AOAC Int. 93, 190–196.
Altenbach, S. B., Tanaka, C. K., and Allen, P. V. (2014). Quantitative proteomic analysis of wheat grain proteins reveals differential effects of silencing of omega-5 gliadin genes in transgenic lines. J. Cereal Sci. 59, 118–125. doi: 10.1016/j.jcs.2013.11.008
Battais, F., Richard, C., Jacquenet, S., Denery-Papini, S., and Moneret-Vautrin, D. A. (2008). Wheat grain allergies: an update on wheat allergens. Eur. Ann. Allergy Clin. Immunol. 40, 67–76.
Bevan, M., Bancroft, I., Bent, E., Love, K., Goodman, H., Dean, C., et al. (1998). Analysis of 1.9 Mb of contiguous sequence from chromosome 4 of Arabidopsis thaliana. Nature 391, 485–488. doi: 10.1038/35140
Bogaerts, A., Baggerman, G., Vierstraete, E., Schoofs, L., and Verleyen, P. (2009). The hemolymph proteome of the honeybee: gel-based or gel-free? Proteomics 9, 3201–3208. doi: 10.1002/pmic.200800604
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Breiteneder, H., and Mills, E. N. C. (2005). Plant food allergens – structural and functional aspects of allergenicity. Biotechnol. Adv. 23, 395–399. doi: 10.1016/j.biotechadv.2005.05.004
Breiteneder, H., and Radauer, C. (2004). A classification of plant food allergens. J. Allergy Clin. Immunol. 113, 821–830. doi: 10.1016/j.jaci.2004.01.779
Danchenko, M., Skultety, L., Rashydov, N. M., Berezhna, V. V., Mátel, L., Salaj, T., et al. (2009). Proteomic analysis of mature soybean seeds from the chernobyl area suggests plant adaptation to the contaminated environment. J. Proteome Res. 8, 2915–2922. doi: 10.1021/pr900034u
Dupont, F. M., Vensel, W. H., Tanaka, C. K., Hurkman, W. J., and Altenbach, S. B. (2011). Deciphering the complexities of the wheat flour proteome using quantitative two-dimensional electrophoresis, three proteases and tandem mass spectrometry. Proteome Sci. 9, 10. doi: 10.1186/1477-5956-9-10
Ferranti, P., Marnone, G. R., Picariello, G., and Addeo, F. (2007). Mass spectrometry analysis of gliadins in celiac disease. J. Mass Spectrom. 42, 1531–1548. doi: 10.1002/jms.1361
Gonzalez-Fernandez, R., Aloria, K., Arizmendi, J. M., and Jorrin-Novo, J. V. (2013). Application of label-free shotgun nUPLC-MSE and 2-DE approaches in the study of Botrytis cinerea mycelium. J. Proteome Res. 12, 3042–3056. doi: 10.1021/pr3010937
Hajduch, M., Casteel, J. E., Hurrelmeyer, K. E., Song, Z., Agrawal, G. K., and Thelen, J. J. (2006). Proteomic analysis of seed filling in Brassica napus. Developmental characterization of metabolic isozymes using high-resolution two-dimensional gel electrophoresis. Plant Physiol. 141, 32–46. doi: 10.1104/pp.105.075390
Hajduch, M., Ganapathy, A., Stein, J. W., and Thelen, J. J. (2005). A systematic proteomic study of seed filling in soybean. Establishment of high-resolution two-dimensional reference maps, expression profiles, and an interactive proteome database. Plant Physiol. 137, 1397–1419. doi: 10.1104/pp.104.056614
Hajduch, M., Hearne, L. B., Miernyk, J. A., Casteel, J. E., Joshi, T., Agrawal, G. K., et al. (2010). Systems analysis of seed filling in Arabidopsis: using general linear modeling to assess concordance of transcript and protein expression. Plant Physiol. 152, 2078–2087. doi: 10.1104/pp.109.152413
Hofmann, S. C., Fischer, J., Eriksson, C., Gref, O. B., Biedermann, T., and Jakob, T. (2012). IgE detection to alpha/beta/gamma-gliadin and its clinical relevance in wheat-dependent exercise-induced anaphylaxis. Allergy 67, 1457–1460. doi: 10.1111/all.12020
Houston, N. L., Hajduch, M., and Thelen, J. J. (2009). Quantitative proteomics of seed filling in castor: comparison with Soybean and rapeseed reveals differences between photosynthetic and nonphotosynthetic seed metabolism. Plant Physiol. 151, 857–868. doi: 10.1104/pp.109.141622
Hurkman, W. J., and Tanaka, C. K. (1986). Solubilization of plant membrane-proteins for analysis by two-dimensional gel-electrophoresis. Plant Physiol. 81, 802–806. doi: 10.1104/pp.81.3.802
Hurkman, W. J., Vensel, W. H., Tanaka, C. K., Whitehand, L., and Altenbach, S. B. (2009). Effect of high temperature on albumin and globulin accumulation in the endosperm proteome of the developing wheat grain. J. Cereal Sci. 49, 12–23. doi: 10.1016/j.jcs.2008.06.014
Jacobsen, S., Grove, H., Jensen, K. N., Sørensen, H. A., Jessen, F., Hollung, K., et al. (2007). Multivariate analysis of 2-DE protein patterns - Practical approaches. Electrophoresis 28, 1289–1299. doi: 10.1002/elps.200600414
James, J. M., Sixbey, J. P., Helm, R. M., Bannon, G. A., and Burks, A. W. (1997). Wheat alpha-amylase inhibitor: a second route of allergic sensitization. J. Allergy Clin. Immunol. 99, 239–244. doi: 10.1016/S0091-6749(97)70103-9
Kamal, A. H. M., Kim, K. H., Shin, D. H., Seo, H. S., Shin, K. H., Park, G. S., et al. (2009). Proteomics profile of pre-harvest sprouting wheat by using MALDI-TOF mass spectrometry. Plant Omics J. 2, 110–119.
Klubicova, K., Danchenko, M., Skultety, L., Berezhna, V. V., Rashydov, N. M., and Hajduch, M. (2013). Radioactive Chernobyl environment has produced high-oil flax seeds that show proteome alterations related to carbon metabolism during seed development. J. Proteome Res. 12, 4799–4806. doi: 10.1021/pr400528m
Klubicova, K., Danchenko, M., Skultety, L., Berezhna, V. V., Uvackova, L., Rashydov, N. M., et al. (2012). Soybeans grown in the Chernobyl area produce fertile seeds that have increased heavy metal resistance and modified carbon metabolism. PLoS ONE 7:e48169. doi: 10.1371/journal.pone.0048169
Klubicova, K., Danchenko, M., Skultety, L., Miernyk, J. A., Rashydov, N. M., Berezhna, V. V., et al. (2010). Proteomics analysis of flax grown in Chernobyl area suggests limited effect of contaminated environment on seed proteome. Environ. Sci. Technol. 44, 6940–6946. doi: 10.1021/es100895s
Luque-Garcia, J. L., Cabezas-Sanchez, P., and Camara, C. (2011). Proteomics as a tool for examining the toxicity of heavy metals. Trac Trends Anal. Chem. 30, 703–716. doi: 10.1016/j.trac.2011.01.014
Mameri, H., Denery-Papini, S., Pietri, M., Tranquet, O., Larré, C., Drouet, M., et al. (2012). Molecular and immunological characterization of wheat Serpin (Tri a 33). Mol. Nutr. Food Res. 56, 1874–1883. doi: 10.1002/mnfr.201200244
Masci, S., D’ovidio, R., Lafiandra, D., and Kasarda, D. D. (1998). Characterization of a low-molecular-weight glutenin subunit gene from bread wheat and the corresponding protein that represents a major subunit of the glutenin polymer. Plant Physiol. 118, 1147–1158. doi: 10.1104/pp.118.4.1147
Mauro Martin, I. S., Garicano Vilar, E., Collado Yurrutia, L., and Ciudad Cabanas, M. J. (2014). Is gluten the great etiopathogenic agent of disease in the xxi century? Nutr. Hosp. 30, 1203–1210.
Mills, E. N. C., Jenkins, J. A., Alcocer, M. J. C., and Shewry, P. R. (2004). Structural, biological, and evolutionary relationships of plant food allergens sensitizing via the gastrointestinal tract. Crit. Rev. Food Sci. Nutr. 44, 379–407. doi: 10.1080/10408690490489224
Olivieri, M., Biscardo, C. A., Palazzo, P., Pahr, S., Malerba, G., Ferrara, R., et al. (2013). Wheat IgE profiling and wheat IgE levels in bakers with allergic occupational phenotypes. Occup. Environ. Med. 70, 617–622. doi: 10.1136/oemed-2012-101112
Pahr, S., Constantin, C., Mari, A., Scheiblhofer, S., Thalhamer, J., Ebner, C., et al. (2012). Molecular characterization of wheat allergens specifically recognized by patients suffering from wheat-induced respiratory allergy. Clin. Exp. Allergy 42, 597–609. doi: 10.1111/j.1365-2222.2012.03961.x
Payne, P. I., Holt, L. M., Jarvis, M. G., and Jackson, E. A. (1985). Two-dimensional fractionation of the endosperm proteins of bread wheat (Triticum aestivum) – biochemical and genetic-studies. Cereal Chem. 62, 319–326.
Rubio-Tapia, A., Kyle, R. A., Kaplan, E. L., Johnson, D. R., Page, W., Erdtmann, F., et al. (2009). Increased prevalence and mortality in undiagnosed celiac disease. Gastroenterology 137, 88–93. doi: 10.1053/j.gastro.2009.03.059
Salcedo, G., Quirce, S., and Diaz-Perales, A. (2011). Wheat allergens associated with Baker’s asthma. J. Investig. Allergol. Clin. Immunol. 21, 81–92.
Sancho, A. I., Gillabert, M., Tapp, H., Shewry, P. R., Skeggs, P. K., and Mills, E. N. C. (2008). Effect of environmental stress during grain filling on the soluble proteome of wheat (Triticum aestivum) dough liquor. J. Agric. Food Chem. 56, 5386–5393. doi: 10.1021/jf800209b
Sapone, A., Bai, J. C., Ciacci, C., Dolinsek, J., Green, P. H., Hadjivassiliou, M., et al. (2012). Spectrum of gluten-related disorders: consensus on new nomenclature and classification. BMC Med. 10:13. doi: 10.1186/1741-7015-10-13
Tatham, A. S., and Shewry, P. R. (2008). Allergens in wheat and related cereals. Clin. Exp. Allergy 38, 1712–1726.
Uvackova, L., Skultety, L., Bekesova, S., Mcclain, S., and Hajduch, M. (2013a). The MS(E)-proteomic analysis of gliadins and glutenins in wheat grain identifies and quantifies proteins associated with celiac disease and baker’s asthma. J. Proteomics 93, 65–73. doi: 10.1016/j.jprot.2012.12.011
Uvackova, L., Skultety, L., Bekesova, S., Mcclain, S., and Hajduch, M. (2013b). MSE based multiplex protein analysis quantified important allergenic proteins and detected relevant peptides carrying known epitopes in wheat grain extracts. J. Proteome Res. 12, 4862–4869. doi: 10.1021/pr400336f
Van Cutsem, E., Simonart, G., Degand, H., Faber, A. M., Morsomme, P., and Boutry, M. (2011). Gel-based and gel-free proteomic analysis of Nicotiana tabacum trichomes identifies proteins involved in secondary metabolism and in the (a)biotic stress response. Proteomics 11, 440–454. doi: 10.1002/pmic.201000356
van den Broeck, H. C., America, A. H. P., Smulders, M. J. M., Bosch, D., Hamer, R. J., Gilissen, L. J., et al. (2009). A modified extraction protocol enables detection and quantification of celiac disease-related gluten proteins from wheat. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 877, 975–982. doi: 10.1016/j.jchromb.2009.02.035
Weiss, W., Vogelmeier, C., and Gorg, A. (1993). Electrophoretic characterization of wheat-grain allergens from different cultivars involved in bakers asthma. Electrophoresis 14, 805–816. doi: 10.1002/elps.11501401126
Wieser, H. (1996). Relation between gliadin structure and coeliac toxicity. Acta Paediatr. Suppl. 412, 3–9. doi: 10.1111/j.1651-2227.1996.tb14239.x
Yang, L. M., Tian, D. G., Luo, Y. M., Zhang, R. Y., Ren, C. M., and Zhou, X. (2011). Proteomics-based identification of storage, metabolic, and allergenic proteins in wheat seed from 2-DE gels. Afr. J. Agric. Res. 6, 808–816.
Keywords: Triticum aestivum, gel-based, quantification, MALDI-TOF/TOF, 2-DE, 7 cm IPG, grain, allergen
Citation: Fekecsová S, Danchenko M, Uvackova L, Skultety L and Hajduch M (2015) Using 7 cm immobilized pH gradient strips to determine levels of clinically relevant proteins in wheat grain extracts. Front. Plant Sci. 6:433. doi: 10.3389/fpls.2015.00433
Received: 26 February 2015; Accepted: 27 May 2015;
Published online: 12 June 2015
Edited by:
Jesus V. Jorrin Novo, University of Cordoba, SpainReviewed by:
Benjamin Schwessinger, University of California, Davis, USALoïc Rajjou, AgroParisTech – Paris Institute of Technology for Life, Food and Environmental Sciences, France
Copyright © 2015 Fekecsová, Danchenko, Uvackova, Skultety and Hajduch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin Hajduch, Department of Developmental and Reproduction Biology, Institute of Plant Genetics and Biotechnology, Slovak Academy of Sciences, Akademicka 2, P.O. Box 39A, Nitra, Slovakia,aGFqZHVjaEBzYXZiYS5zaw==