 Changyu Jin†
Changyu Jin† Dong Li†
Dong Li† Chenhao Gao
Chenhao Gao Kaige Liu
Kaige Liu Shuanghui Qi
Shuanghui Qi Shaowei Duan
Shaowei Duan Zixiong Li
Zixiong Li Jingyun Gong
Jingyun Gong Jianjun Wang
Jianjun Wang Jiangbo Hai
Jiangbo Hai Mingxun Chen*
Mingxun Chen*- State Key Laboratory of Crop Stress Biology for Arid Areas and College of Agronomy, Northwest A&F University, Yangling, China
Previous studies have shown that several ACYL–ACYL CARRIER PROTEIN DESATURASE (AtAAD) members in Arabidopsis thaliana are responsible for oleic acid (C18:1) biosynthesis. Limited research has been conducted on another member, AtAAD5, and its paralog BnAAD5 in the closely related and commercially important plant, Brassica napus. Here, we found that AtAAD5 was predominantly and exclusively expressed in developing embryos at the whole seed developmental stages. The aad5 mutation caused a significant decrease in the amounts of oil and C18:1, and a considerable increase in the content of stearic acid (C18:0) in mature seeds, suggesting that AtAAD5 functioned as an important facilitator of seed oil biosynthesis. We also cloned the full-length coding sequence of BnAAD5-1 from the A3 subgenome of the B. napus inbred line L111. We showed that ectopic expression of BnAAD5-1 in the A. thaliana aad5-2 mutant fully complemented the phenotypes of the mutant, such as lower oil content and altered contents of C18:0 and C18:1. These results help us to better understand the functions of AAD members in A. thaliana and B. napus and provide a promising target for genetic manipulation of B. napus.
Introduction
Seed fatty acids (FAs) and FA-derived complex lipids not only provide nutrients for humans and livestock (Li et al., 2006; Graham, 2008), but also serve as raw materials for industries and biofuel production (Durrett et al., 2008; Lu et al., 2011). Biosynthesis of seed oil is under the control of multiple genes, and occurs in plant cells in three steps (Baud et al., 2008; Itabe, 2010; Chapman and Ohlrogge, 2012). The first step is the production of pyruvate and other substances during glycolysis. Catabolysis of pyruvate and other substances leads to the FA precursor acetyl-CoA, which results in biosynthesis of C16-18 FAs in plastids. In the second step, FA derivatives are formed at acyl chains. FA formation occurs in the cytoplasm after chain elongation and desaturation of most C16-18 FAs from the first step. Finally, triacylglycerols are formed to store the new oil in oil bodies.
There are six main types of FAs in seed oil from A. thaliana: palmitic acid (C16:0), stearic acid (C18:0), oleic acid (C18:1Δ9), α-linoleic acid (C18:2), α-linolenic acid (C18:3), and eicosenoic acid (C20:1Δ11); many minor FAs also exist in A. thaliana (<3 mol% each). The ACYL–ACYL CARRIER PROTEIN (ACP) DESATURASE (AAD) members have specificity for the acyl chain length of the particular substrate and generate double bonds between particular carbon atoms (Kachroo et al., 2007; Bryant et al., 2016). They are the only enzymes catalyzing the conversion of C18:0 into C18:1 in plant cells, and thus their activity primarily controls the saturated to monounsaturated FAs ratio (Kachroo et al., 2007).
The A. thaliana genome contains seven AtAAD genes, including SUPPRESSOR OF SALICYLIC ACID INSENSITIVE2 (AtSSI2/AtFAB2, AT2G43710), AtAAD1 (AT5G16240), AtAAD2 (AT3G02610), AtAAD3 (AT5G16230), AtAAD4 (AT3G02620), AtAAD5 (AT3G02630), and AtAAD6 (AT1G43800) (Kachroo et al., 2007; Bryant et al., 2016). AtSSI2, AtAAD1, AtAAD3, AtAAD4, and AtAAD5 have all been found to desaturate C18:0-ACP at the Δ9 position, and AtSSI2 and AtAAD3 were also shown to have specific activity on C16:0-ACP (Kachroo et al., 2007). Expression of AtSSI2 occurs in the endosperm and embryo (Le et al., 2010). Stearic acid (C18:0) is accumulated in the ssi2 mutant in seed and vegetative tissues. This accumulation indicates reduced Δ9 desaturation, because it occurs by depleting C18:1Δ9 (Lightner et al., 1994). AtAAD1 is strictly localized in the embryo (Le et al., 2010), and the aad1 mutation causes a significant increase in C18:0 and a decrease in C18:1Δ9 contents in the embryo (Bryant et al., 2016). AtAAD2 is most strongly expressed in the peripheral endosperm, and AtAAD3 is most strongly expressed in the chalazal endosperm (Le et al., 2010). The aad2 mutants exhibit an approximately 30% reduction in the levels of C18:1Δ9 plus C20:1Δ13, while the levels of C18:1Δ9 plus C20:1Δ13 in aad4 mutants are not significantly changed in the endosperm and seed coat (Bryant et al., 2016). AtAAD3, which functions independently of AtSSI2, is responsible for the biosynthesis of C16:1Δ9, C18:1Δ11, and C20:1Δ13 in the endosperm and seed coat (Bryant et al., 2016). Taken together, the specific expression patterns and effects of AtSSI2 and AtAAD1-4 on seed FA biosynthesis have been gained a lot in A. thaliana. In addition, AAD genes occur in several plant species, and have been previously isolated and/or characterized (Shanklin and Somerville, 1991; Thompson et al., 1991; Cahoon et al., 1996, 1997; Lindqvist et al., 1996; Cahoon and Browse, 1998; Whittle et al., 2005). However, little is currently known about AtAAD5, and its paralog BnAAD5 in Brassica napus, the commercially important close relative of A. thaliana.
In the current study, we found that AtAAD5 is specifically expressed in the embryo during seed development in A. thaliana. We demonstrated that AtAAD5 promotes oil and C18:1 biosynthesis in A. thaliana seeds. We also cloned and functionally characterized B. napus AAD5-1 (BnAAD5-1), showing that it exhibits a conserved role with AtAAD5 in regulating seed FA accumulation when expressed in A. thaliana.
Materials and Methods
Plant Materials and Growth Conditions
The Col-0 ecotype was used as the wild type A. thaliana control, and the mutants were aad5-1 (SALK_129779C), and aad5-2 (SALK_035968C) in the Col-0 background. All A. thaliana plants were grown at 22°C with a 16 h/8 h light/dark photoperiod, which has been reported in detail previously (Duan et al., 2017). The B. napus inbred line L111 was maintained in the greenhouse of South Campus, Northwest A&F University, China. T-DNA mutant were genotyped using specific primers (Supplementary Table S1).
Gene Cloning of BnAAD5-1 from B. napus
Primers were designed to amplify the BnAAD5-1 gene based on the full-length coding domain sequence of BnAAD5-1 (GenBank Number XP_013735719.1). Developing seeds were used as a source of total RNA to synthesize template complementary DNA (cDNA). Seeds were collected from the B. napus inbred line L111 15 days after pollination. We used the pMD18-T vector for cloning (TaKaRa Bio, Dalian, China), and eight single colonies were picked randomly and sequenced by Sangon Biotechnology (Shanghai, China). Cloning primers are listed in Supplementary Table S1.
Plasmid Construction
To obtain the construct of 35S:BnAAD5-1, the amplified full-length coding regions of BnAAD5-1 were digested with Xma I and Spe I and then were cloned into pGreen-35S; this was driven by the 35S promoter. To construct pAtAAD5:GUS, the 824-bp AtAAD5 genomic region including a 283-bp promoter region, ATG, and a 538-bp region downstream of the ATG start codon in sequence was amplified and then cloned into pHY107 (Liu et al., 2007). Plasmid construction primers are listed in Supplementary Table S1.
Generation of A. thaliana Transgenic Plants
The pAtAAD5:GUS and 35S:BnAAD5-1 constructs were transformed into Agrobacterium tumefaciens GV3101 and were used to transform A. thaliana wild type and aad5-2 plants, respectively, and the floral dip method was utilized (Clough and Bent, 1998). We used Basta® selection and genotyping to confirm that plants were transgenic until T3 homozygous lines were obtained.
Morphological Observation of Mature Seeds
Mature A. thaliana seeds were randomly selected from major inflorescences, specifically from siliques in the basal region, and photographed using an OLYMPUS SZ 61 stereomicroscope.
Seed FA Measurement
Mature A. thaliana seeds for FA determination were collected from siliques in the basal region of the major inflorescences of 16 individual plants sown in different pots arranged in a randomized block design. Seed FA determination was conducted as previously described (Poirier et al., 1999; Chen et al., 2012). In brief, seeds were infused into the methanol solution containing 1 M HCl at 80°C for 2 h, which would convert FAs into the corresponding methyl esters. Then, FA methyl esters were extracted with the hexane, and were subsequently quantified by a gas chromatograph (GC-2014; Shimadzu).
Gene Expression Analysis
Total RNA samples were isolated from A. thaliana young siliques or B. napus developing seeds with the MiniBEST Plant RNA Extraction Kit (TaKaRa) and their corresponding cDNA samples were biosynthesized with PrimerScript RT (TaKaRa). Reverse transcription-PCR (RT-PCR) and quantitative RT-PCR (qRT-PCR) were conducted for three biological replicates. SYBR Green Master Mix (TaKaRa) was utilized for qRT-PCR analysis. The A. thaliana house-keeping gene AtEF1aA4 was regarded as an internal control. Primers used for the RT-PCR and qRT-PCR analyses are listed in Supplementary Table S1.
Results
Analysis of AtAAD5 Expression Pattern
Previous RT-PCR results showed that AtAAD5 was widely expressed in A. thaliana tissues, including leaves, stems, roots, flowers, and siliques (Kachroo et al., 2007). To better investigate the spatiotemporal expression pattern of AtAAD5, we obtained 19 independent lines of pAtAAD5:GUS from a wild type background. GUS staining patterns were similar among most of the lines; therefore, one representative line was used for GUS staining analysis. The result showed that AtAAD5 was expressed in several tissues, including hypocotyl vascular bundles (Figure 1A), root tips (Figures 1A,B), cotyledons (Figure 1B), and young expanding true leaves (Figure 1B). Notably, AtAAD5 was highly present in developing embryos at different stages (Figures 1G–L). However, no GUS staining was observed in other tissues, such as expanded true leaves (Figure 1C), cauline leaves (Figures 1D), flowers (Figure 1D), young siliques (Figure 1E), seed coats (Figures 1F–L), or endosperms (Figures 1F–L). These results suggested that AtAAD5 controls seed traits mainly occurring in the A. thaliana embryo at the whole seed developmental stages (Baud et al., 2002; Fait et al., 2006; Graham, 2008; Baud and Lepiniec, 2009).
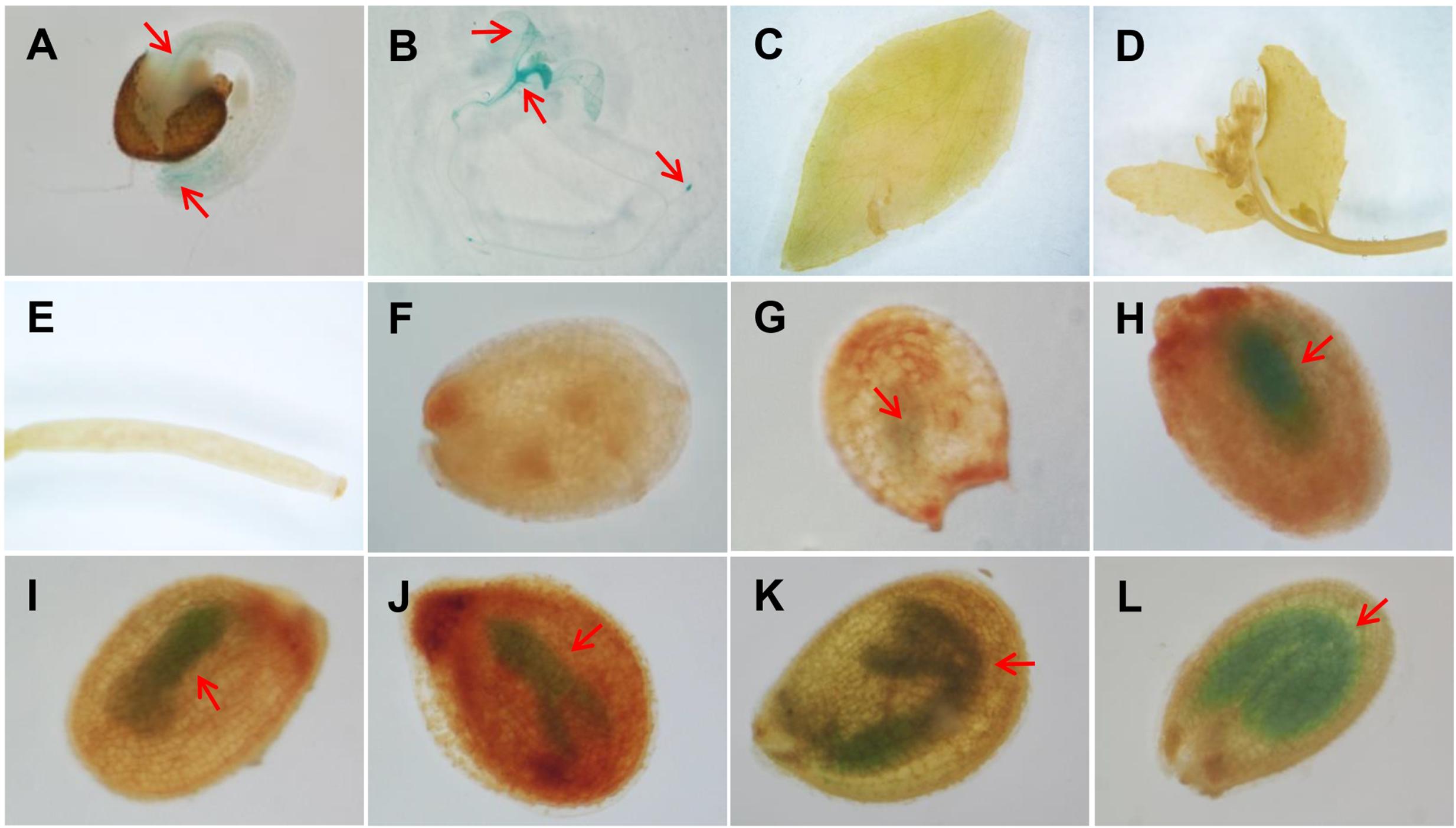
FIGURE 1. The expression pattern of AtAAD5 as determined by GUS staining of the representative A. thaliana pAtAAD5:GUS line. (A,B) seedlings 1 (A) and 5 (B) days after germination; (C) rosette leaves; (D) cauline leaves and flowers; (E) siliques 3 days after pollination; (F–L) developing seeds at different stages (F: zygote; G: globular; H–J: liner cotyledon; K: bending cotyledon; L: mature green cotyledon). Red arrow indicates the position of GUS signal.
AtAAD5 Promotes Oil and Oleic Acid Biosynthesis in Seeds
AtAAD5 was previously screened by Kachroo et al. (2007) for T-DNA insertion mutants, but they did not obtain homozygous lines. In this study, we successfully obtained two T-DNA insertion mutants SALK_129779C and SALK_035968C from the Col-0 ecotype in the 5′ untranslated region and the exon of AtAAD5, respectively, from the Arabidopsis Biological Resources Center (ABRC), which were designated aad5-1 and aad5-2, respectively (Figure 2A). The genotyping PCR result indicated the presence of the two homozygous mutants (Figure 2B). The RT-PCR result showed that the N- and C-terminal AtAAD5 transcripts were not detected in aad5-1 and aad5-2 mutants, respectively (Figure 2C). More PCR product was amplified by C-terminal primers compared to N-terminal primers for the Col-0 RNA samples (Figure 2C), suggesting that the PCR amplification efficiency of the C-terminal primers is higher than that of the N-terminal primers. Notably, the C-terminal AtAAD5 transcript in aad5-1 was almost as strong as the wild type, which needs further investigation (Figure 2C).
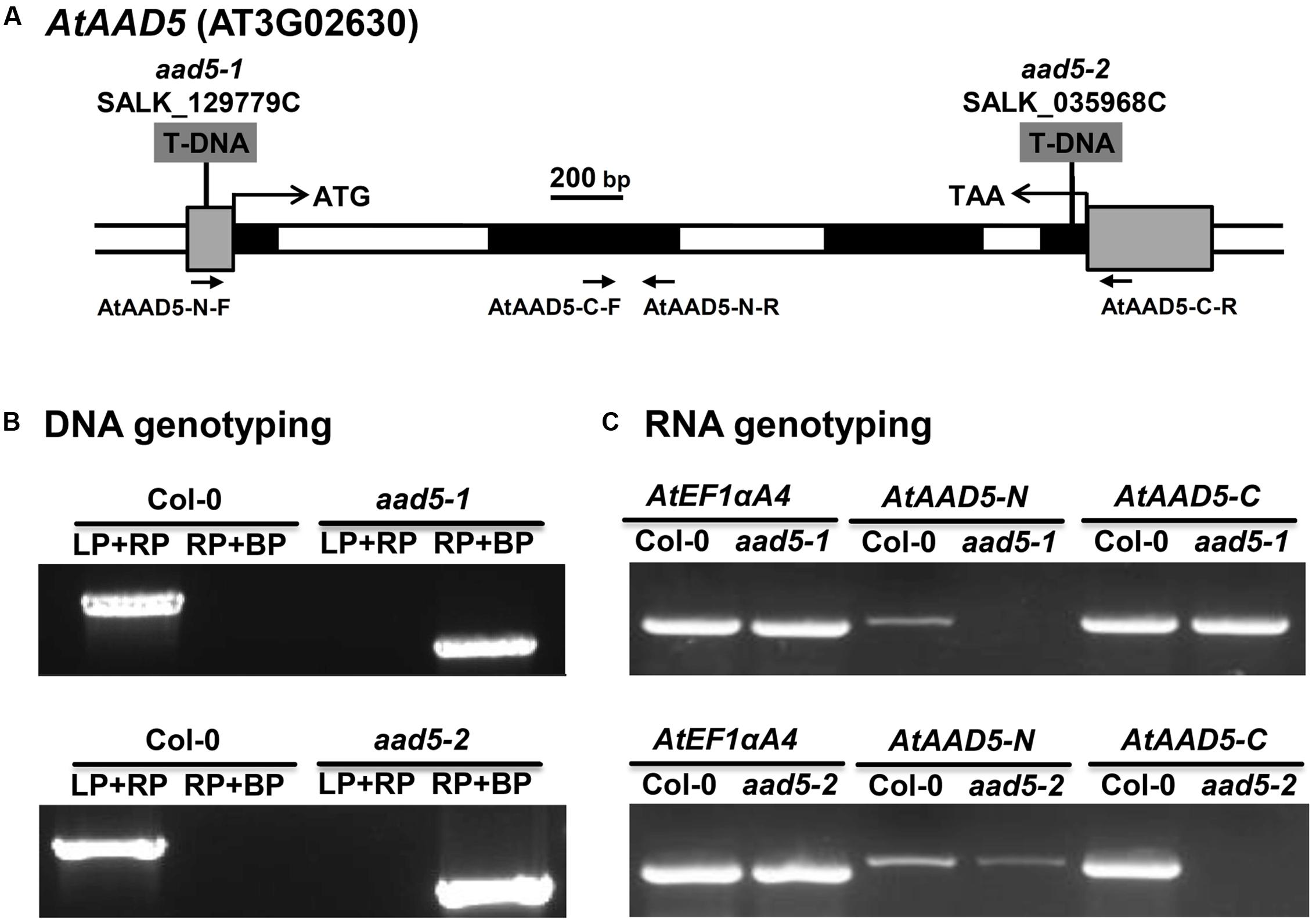
FIGURE 2. Molecular identification of the A. thaliana aad5 mutation. (A) Structure of the AtAAD5 gene indicating the position of the T-DNA insertions in SALK_129779C (aad5-1) and SALK_035968C (aad5-2) mutants. The coding and untranslated regions of AtAAD5 are represented by black and gray boxes, respectively, and introns and other genomic regions are represented by open boxes. Translation start site (ATG) and stop codon (TAA) are indicated. (B) PCR-based DNA genotyping of the two mutants. LP and RP refer to the AtAAD5 gene specific primers and BP refers to T-DNA right-border primer given in Supplementary Table S1. (C) Detection of N- and C- terminal transcripts of AtAAD5 in the two mutants by RT-PCR using primer pairs indicated in (A). AtEF1αA4 was amplified as an internal control.
To explore the biological function of seed FA accumulation, we used mature seeds from wild type and aad5 plants to determine the contents of major FAs. The result showed that the seed oil content was much lower in aad5 mutants than in the wild type seeds (Figures 3A,B). In aad5 seeds, there was a significant increase in the amount of C18:0 and a significant decrease in the C18:1 content (Figure 3C), suggesting that AAD5 plays a role in the desaturation of C18:0-ACP. However, we did not observe clear differences among morphological traits of seeds, including color of the seed coat, the size of the seed, or the dry weight of the seed (Supplementary Figure S1) between mature seeds of wild type and aad5 plants. These results suggested that AtAAD5 promotes seed oil and oleic acid biosynthesis in the A. thaliana embryo.
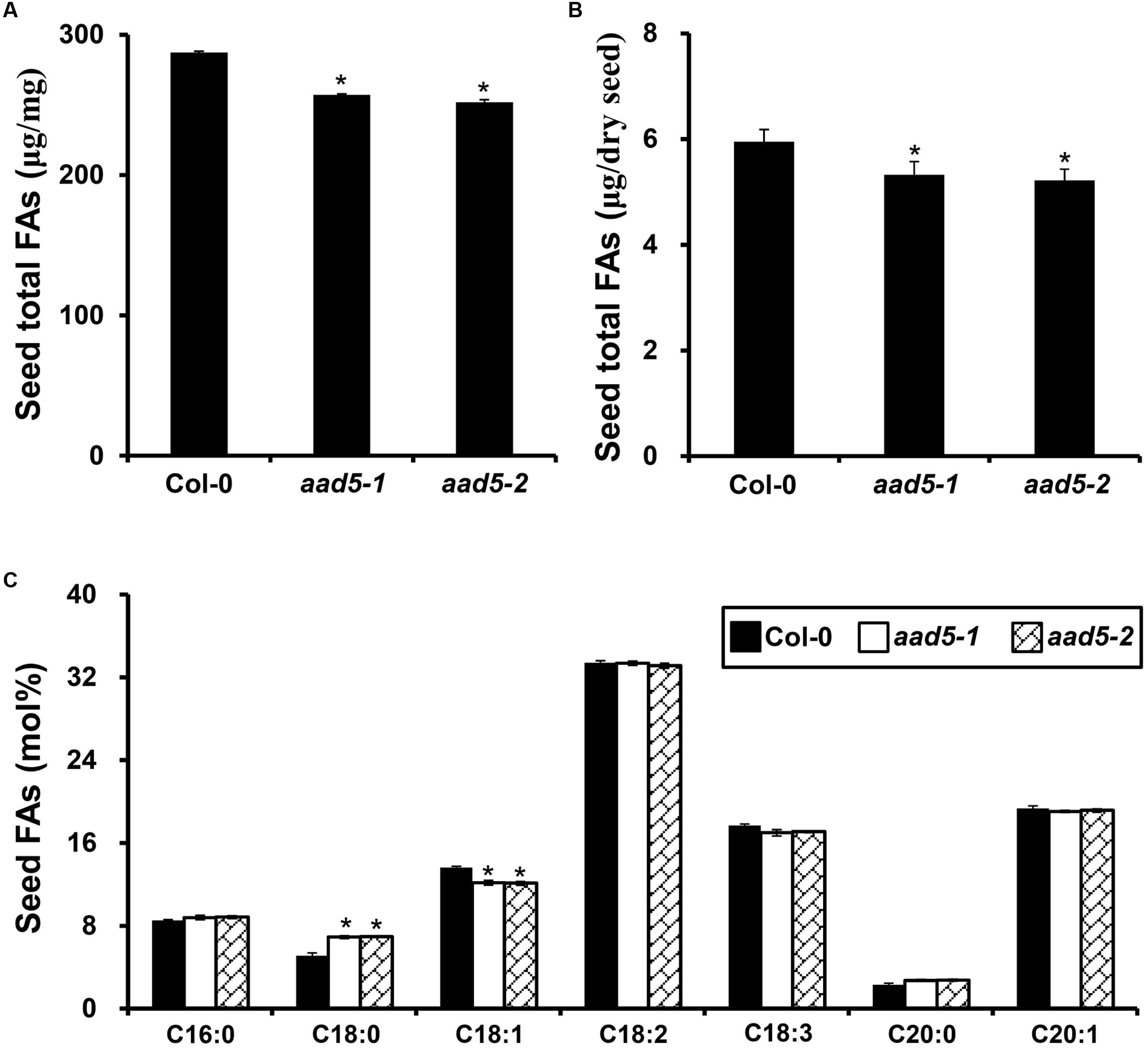
FIGURE 3. Effect of AtAAD5 on A. thaliana seed FA biosynthesis. (A) Comparison of seed total FA content (μg/mg) between wild type (Col-0) and aad5 plants. (B) Comparison of seed total FA content (μg/seed) between the wild type (Col-0) and aad5 plants. (C) Comparison of contents of major seed FA compositions between the wild type (Col-0) and aad5 plants. Asterisks indicate significant differences in the seed total FA content (A,B) and the contents of major seed FA compositions (C) compared to that in the wild type (two-tailed paired Student’s t-test, P ≤ 0.05). DW, dry weight. Values are means ± SD (n = 5). Error bars indicate standard deviation.
Analysis of BnAAD5-1 Sequence
We named the seven BnAAD5 paralogs from the B. napus genome BnAAD5 1-7 (Figure 4). As shown in Figure 4A, the cloned BnAAD5-1 (L111) sequence from the B. napus inbred line L111 was the same as the BnAAD5-1 protein sequence (XP_013735719.1) of the B. napus cultivar Zhongshuang11 (ZS11). BnAAD5-1 is located on the A3 subgenome of the B. napus cultivar ZS11, and our sequence results suggested that we had cloned BnAAD5-1 from the L111 A3 subgenome. BnAAD5-1 (L111) was also predicted to share 96% amino acid sequence identity with AtAAD5 (Figure 4A).
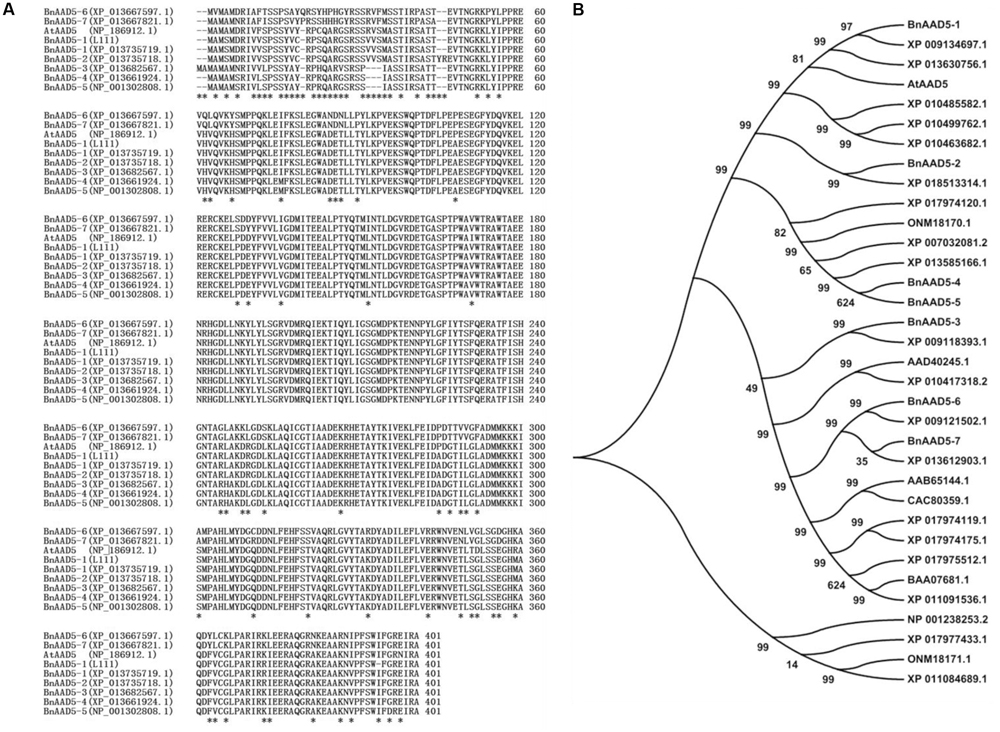
FIGURE 4. Sequence analysis of AAD5 proteins from A. thaliana, B. napus, and other oil-producing plant species. (A) Alignment of AAD5 protein sequences from A. thaliana and B. napus. The alignment was conducted using MUSCLE (http://www.ebi.ac.uk/Tools/msa/muscle/) and discrepant amino acids are indicated by asterisks. (B) Phylogenetic analysis of 34 AAD5 family proteins from 11 oil-producing plant species. A neighbor-joining tree (Jones–Taylor–Thornton model) was generated by MEGA6. A bootstrap analysis with 1,000 replicates was performed to assess the statistical reliability of the tree topology. The accession numbers corresponding to the species names are as follows: AtAAD5 (A. thaliana); BnAAD5 1-7 (B. napus); XP_009134697.1, XP_018513314.1, XP_009118393.1, and XP_009121502.1 (B. rapa); XP_013630756.1, XP_013585166.1, and XP_013612903.1 (B. oleracea); AAD40245.1 (B. juncea); ONM18170.1 and ONM18171.1 (Zea mays); XP_010499762.1, XP_010485582.1, XP_010463682.1, and XP_010417318.2 (Camelina sativa); BAA07681.1, XP_011091536.1, and XP_011084689.1 (Sesamum indicum); AAB65144.1 and CAC80359.1 (Helianthus annuus); NP_001238253.2 (Glycine max); XP_017977433.1, XP_017975512.1, XP_007032081.2, XP_017974119.1, XP_017974175.1, XP_017974120.1 (Theobroma cacao).
We performed a phylogenetic analysis to investigate the evolutionary relationship between BnAAD5-1 and 33 AAD5 proteins from 11 oil-producing plant species. The analysis indicated that BnAAD5-1 is most related to the three AAD5 sequences, including BrAAD5 (XP_009134697.1) from B. rapa, BoAAD5 (XP_013630756.1) from B. oleracea, and AtAAD5 from A. thaliana (Figure 4B).
BnAAD5-1 Fully Rescues the FA Phenotype of A. thaliana aad5-2 Seeds
To further elucidate the function of BnAAD5-1 in seed FA biosynthesis, we over-expressed it in the A. thaliana aad5-2 mutant, using the construct 35S:BnAAD5-1 (Figure 5A). A total of 23 independent T1 transgenic plants were obtained following Basta® selection, and five independent transgenic lines (aad5-2 35S:BnAAD5-1 T3) were confirmed by PCR amplification of the BnAAD5-1 gene with the specific primers 35S_Pro/BnAAD5-1_R1 (Figure 5A; Supplementary Table S1). Expression of the BnAAD5-1 gene in these transgenic plants was measured by qRT-PCR, and was determined to be highest in the transgenic line aad5-2 35S:BnAAD5-1#10, whereas its expression was not detected in the wild type or aad5-2 plants (Figure 5B). We observed that ectopic expression of BnAAD5-1 fully rescued aad5-2 seed phenotypes, such as lower oil content (Figure 5C) and altered contents of C18:0 and C18:1 (Figure 5D). Although the aad5-2 35S:BnAAD5-1#8 transgenic line showed the lowest expression of BnAAD5-1, the contents of total FAs, C18:0, and C18:1 were close to those of other transgenic plants (Figures 5B–D). This indicated that BnAAD5-1 regulates seed FA accumulation in a dose-independent manner when overexpressed in A. thaliana. These results together suggested that BnAAD5-1 has a similar function to AtAAD5.
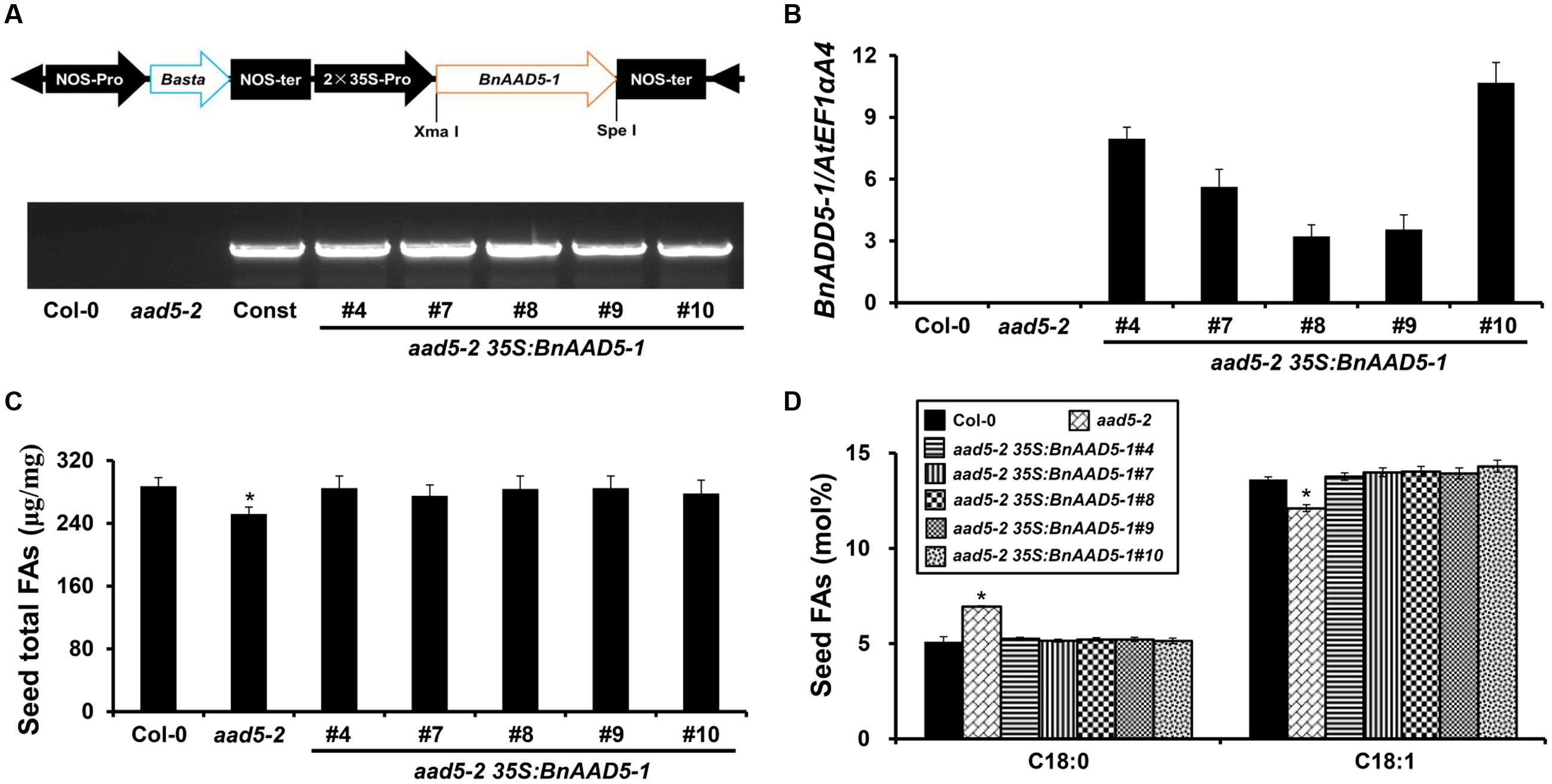
FIGURE 5. Effect of BnAAD5-1 on seed FA accumulation when expressed in the A. thaliana aad5-2 mutant. (A) Schematic diagram of the constitutive expression cassette of the BnAAD5-1 gene in the binary vector pGreen 2 × 35S (Top figure) and PCR-based DNA genotyping of aad5-2 35S:BnAAD5-1 transgenic plants with the specific primers: 35S_Pro and BnAAD5-1_R1 (Bottom figure). RB, right border; LB, left border; NOS-pro, nopaline synthase promoter; NOS-ter, nopaline synthase terminator; Basta®, glyphosate; 35S-pro, CaMV 35S promoter, Const: 35S:BnAAD5-1 construct. (B) Comparative analysis of BnAAD5-1 expression by qRT-PCR in the wild type (Col-0), aad5-2, and aad5-2 35S:BnAAD5-1 plants. Expression levels were normalized against the expression of the internal control, AtEF1αA4. Error bars indicate standard deviation. (C) Quantitative comparison of total FA content between the wild type (Col-0), aad5-2, and aad5-2 35S:BnAAD5-1 seeds. Asterisks indicate statistically significant differences in total FA content of aad5-2 seeds compared to that of wild type seeds (two-tailed paired Student’s t-test, P ≤ 0.05). Values are means ± SD (n = 5). Error bars indicate standard deviation. (D) Quantitative comparison of FA compositions of C18:0 and C18:1 between the wild type (Col-0), aad5-2, and aad5-2 35S:BnAAD5-1 seeds. Asterisks indicate statistically significant differences in contents of FA compositions of aad5-2 seeds compared to that of wild type seeds (two-tailed paired Student’s t-test, P ≤ 0.05). Values are means ± SD (n = 5). Error bars indicate standard deviation.
Discussion
The increase and optimization of FA composition in oil-producing plant seeds is the most important objective for breeders. Several of the seven AtAAD genes, including AtSSI2, AtAAD1, AtAAD2, AtAAD3, and AtAAD4, have been functionally identified for seed FA biosynthesis in A. thaliana (Kachroo et al., 2007; Bryant et al., 2016). However, little is known about the role of AtAAD5, and its paralog BnAAD5 in seed FA accumulation. Our results provide two major lines of evidence for a conserved and important role for AAD5 in mediating total FAs accumulation in seeds and C18:1 accumulation in the embryo in both A. thaliana and B. napus.
First, the aad5 mutation resulted in a considerable increase in the amounts of oil and C18:0, and a significant decrease in the C18:1 content in mature seeds (Figure 3). The expression of AtAAD5 was stably observed in developing embryos, but not in the endosperm and seed coat, during the whole seed developmental stages (Figure 1). C18:1 FA mainly exists in the forms of C18:1Δ9 and C18:1Δ11 in the A. thaliana embryo and endosperm plus seed coat, respectively, and C18:1Δ11 FA only accounts for less than 1 mol% of total FAs in the embryo (Bryant et al., 2016). The previous study showed that AtAAD5 preferentially desaturates C18:0-ACP substrate at the C9 position (Kachroo et al., 2007). These results together suggested that AtAAD5 plays an important role in controlling the conversion of C:18-ACP to C18:1Δ9 in the A. thaliana embryo. Traits of seeds including color of the coat, size of the seed, and weight of the seed were not altered in aad5 mutants, which is consistent with the fact that AtAAD5 was not expressed in seed coat and endosperm (Figure 1F–L). Intricate regulatory networks control FA accumulation in seeds. These networks also require coordinated development of three distinct seed tissues: embryo, endosperm, and seed coat. Therefore, the disruption of the structural gene AtAAD5 might disturb seed embryo development, causing lower seed oil accumulation (Figures 3A,B). AtAAD5 and AtAAD1 are most closely related based on phylogenetic analyses of the AtAAD family; they are 82% identical at the amino acid level (Kachroo et al., 2007). Consistently, AtAAD1 and AtAAD5 showed similar functions on the conversion of C18:0-ACP to C18:1Δ9 in the A. thaliana embryo (Figure 3C; Kachroo et al., 2007). It is worth mentioning that AtAAD1 negatively affects C18:2 biosynthesis (Kachroo et al., 2007), whereas AtAAD5 has no significant effect on the accumulation of C18:2 and other major seed FAs except for C18:0 and C18:1 (Figure 3C). These results indicated that the two genes have some differences in the regulation of seed FA biosynthesis in the A. thaliana embryo.
Second, ectopic expression of BnAAD5-1 cloned from the A3 subgenome of the B. napus inbred line L111 in the aad5-2 mutant fully rescued altered seed FA contents of the mutant (Figure 5). This strongly suggested that BnAAD5-1 exhibits a conserved role with AtAAD5 in regulating seed FA accumulation when expressed in A. thaliana. However, no obvious differences were observed in the contents of oil, C18:0, and C18:1 in seeds between aad5-2 BnAAD5-1 overexpressors and the wild type control (Figure 5). This indicates that the alterations caused by reduced expression of AtAAD5 and increased expression of BnAAD5-1 in A. thaliana do not simply mirror each other. Arabidopsis thaliana and B. napus are both part of Cruciferae, and there are three A. thaliana loci in the B. rapa, B. oleracea, and B. nigra genomes (Kowalski et al., 1994; Osborn et al., 1997; Lagercrantz, 1998; Haberer et al., 2006). Brassica rapa and B. oleracea hybridize to create B. napus (Parkin et al., 1995; Osborn et al., 1997). During B. napus evolution, there was a high frequency of rearrangement, fusion, and deletion of chromosomes (Lagercrantz, 1998), which led to, on average, 2-8 paralogs in the B. napus genome for each gene locus in A. thaliana (Osborn et al., 1997; Cavell et al., 1998). Here we found a single copy of AtAAD5 in the A. thaliana genome as expected, and seven putative BnAAD5 paralogs in the B. napus genome (Figure 4A). Our previous study showed that BnTOP1α-1 from the inbred L111 line has lost 4 amino acid stretches, compared with BnTOP1α-1 (XP_013685667.1) from ZS11, which collectively correspond to 130 amino acids (Gao et al., 2017). However, the cloned BnAAD5-1 from the inbred L111 line has the same sequence as BnAAD5-1 (XP_013735719.1) from ZS11 at the protein level (Figure 4A), and is most related to BrAAD5 (XP_009134697.1), BoAAD5 (XP_013630756.1), and AtAAD5 (Figure 4B). Saturated FA quantity in B. napus has been increased by seed-specific antisense repression of one BrAAD gene from B. rapa (Knutzon et al., 1992). These results indicate that AAD5 might be conserved during evolution of the cruciferous species (A. thaliana, B. rapa, B. oleracea, and B. napus).
In summary, this study is the first to identify that an AtAAD member, AtAAD5, is responsible for converting C18:0-ACP to C18:1 and promoting oil accumulation in the A. thaliana embryo. In addition, we showed that BnAAD5-1 has a conserved function with AtAAD5 in regulating seed FA accumulation when it is expressed in A. thaliana. Brassica napus is grown as a crop primarily for its seed oil. The identification and manipulation of key B. napus genes controlling seed oil and FA accumulation are of fundamental importance for agricultural production. These results suggest that BnAAD5-1 can be used as a promising target to genetically manipulate B. napus and other oil-producing plants to improve the amounts of seed oil, C18:0, and C18:1.
Author Contributions
CJ and DL carried out the experiments. CJ and CG analyzed the data. KL, SQ, SD, ZL, JG, and JW assisted with doing the experiments. MC conceived and designed the experiments. MC and CJ wrote the manuscript. DL, CG, and JH helped to draft the manuscript and revise the manuscript. All authors read and approved the final manuscript.
Funding
This study was financially supported by Young Elite Scientists Sponsorship Program by CAST (Grant no. 2016QNRC001), Natural Science Foundation of China (Grant no. 31501336), Science Fund for The Cultivation of The Excellent Youth Scholars (Z109021517) and Startup Fund for Talents (Z111021402) of Northwest A&F University, and Breeding Project of Yangling (K3360216010).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
The T-DNA insertion mutants used in this study, SALK_129779C and SALK_035968C, were distributed by ABRC.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.01319/full#supplementary-material
FIGURE S1 | Characterization of seed traits from the wild type (Col-0), aad5, and aad5-2 35S:BnAAD5-1 plants. (A) Microscopy of mature seeds that were selected randomly from wild type, aad5, and aad5-2 35S:BnAAD5-1 plants. Bars: 500 μm. (B) Quantitative comparisons of seed size (length and width) between the wild type, aad5, and aad5-2 35S:BnAAD5-1 plants. (C) Quantitative comparison of dry weight of seeds between the wild type, aad5-2, and aad5-2 35S:BnAAD5-1 plants. Error bars indicate standard deviation. No significant differences in seed size (B) or seed weight (C) were observed between wild type and aad5 or aad5-2 35S:BnAAD5-1 (two-tailed paired Student’s t-test, P ≤ 0.05). Values are means ± SD (n = 5). Error bars indicate standard deviation.
References
Baud, S., Boutin, J. P., Miquel, M., Lepiniec, L., and Rochat, C. (2002). An integrated overview of seed development in Arabidopsis thaliana ecotype WS. Plant Physiol. Biochem. 40, 151–160. doi: 10.1016/S0981-9428(01)01350-X
Baud, S., Dubreucq, B., Miquel, M., Rochat, C., and Lepiniec, L. (2008). Storage reserve accumulation in Arabidopsis: metabolic and developmental control of seed filling. Arabidopsis Book 6:e0113. doi: 10.1199/tab.0113
Baud, S., and Lepiniec, L. (2009). Regulation of de novo fatty acid synthesis in maturing oilseeds of Arabidopsis. Plant Physiol. Biochem. 47, 448–455. doi: 10.1016/j.plaphy.2008.12.006
Bryant, F. M., Munoz-Azcarate, O., Kelly, A. A., Beaudoin, F., Kurup, S., and Eastmond, P. J. (2016). ACYL-ACYL CARRIER PROTEIN DESATURASE2 and 3 are responsible for making omega-7 fatty acids in the Arabidopsis aleurone. Plant Physiol. 172, 154–162. doi: 10.1104/pp.16.00836
Cahoon, E. B., and Browse, J. (1998). A determinant of substrate specificity predicted from the acyl-acyl carrier protein desaturase of developing cat’s claw seed. Plant Physiol. 117, 593–598. doi: 10.1104/pp.117.2.593
Cahoon, E. B., Coughlan, S. J., and Shanklin, J. (1997). Characterization of a structurally and functionally diverged acyl-acyl carrier protein desaturase from milkweed seed. Plant Mol. Biol. 33, 1105–1110. doi: 10.1023/A:1005821007291
Cahoon, E. B., Mills, L. A., and Shanklin, J. (1996). Modification of the fatty acid composition of Escherichia coli by coexpression of a plant acyl-acyl carrier protein desaturase and ferredoxin. J. Bacteriol. 178, 936–939. doi: 10.1128/jb.178.3.936-939.1996
Cavell, A. C., Lydiate, D. J., Parkin, I. A., Dean, C., and Trick, M. (1998). Collinearity between a 30-centimorgan segment of Arabidopsis thaliana chromosome 4 and duplicated regions within the Brassica napus genome. Genome 41, 62–69. doi: 10.1139/g97-097
Chapman, K. D., and Ohlrogge, J. B. (2012). Compartmentation of triacylglycerol accumulation in plants. J. Biol. Chem. 287, 2288–2294. doi: 10.1074/jbc.R111.290072
Chen, M. X., Wang, Z., Zhu, Y. N., Li, Z. L., Hussain, N., Xuan, L. J., et al. (2012). The effect of TRANSPARENT TESTA2 on seed fatty acid biosynthesis and tolerance to environmental stresses during young seedling establishment in Arabidopsis. Plant Physiol. 160, 1023–1036. doi: 10.1104/pp.112.202945
Clough, S. J., and Bent, A. F. (1998). Floral dip: a simplified method for Agrobacterium -mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. doi: 10.1046/j.1365-313x.1998.00343.x
Duan, S., Jin, C., Li, D., Gao, C., Qi, S., Liu, K., et al. (2017). MYB76 inhibits seed fatty acid accumulation in Arabidopsis. Front. Plant Sci. 8:226. doi: 10.3389/fpls.2017.00226
Durrett, T. P., Benning, C., and Ohlrogge, J. (2008). Plant triacylglycerols as feedstocks for the production of biofuels. Plant J. 54, 593–607. doi: 10.1111/j.1365-313X.2008.03442.x
Fait, A., Angelovici, R., Less, H., Ohad, I., Urbanczyk-Wochniak, E., Fernie, A. R., et al. (2006). Arabidopsis seed development and germination is associated with temporally distinct metabolic switches. Plant Physiol. 142, 839–854. doi: 10.1104/pp.106.086694
Gao, C., Qi, S., Liu, K., Li, D., Jin, C., Duan, S., et al. (2017). Functional characterization of Brassica napus DNA topoisomerase Iα-1 and its effect on flowering time when expressed in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 486, 124–129. doi: 10.1016/j.bbrc.2017.03.011
Graham, I. A. (2008). Seed storage oil mobilization. Annu. Rev. Plant Biol. 59, 115–142. doi: 10.1146/annurev.arplant.59.032607.092938
Haberer, G., Mader, M. T., Kosarev, P., Spannagl, M., Yang, L., and Mayer, K. F. X. (2006). Large-scale cis-element detection by analysis of correlated expression and sequence conservation between Arabidopsis and Brassica oleracea. Plant Physiol. 142, 1589–1602. doi: 10.1104/pp.106.085639
Itabe, H. (2010). Intracellular lipid droplet-associated proteins: unique members and their biological functions. Biol. Pharm. Bull. 33:341. doi: 10.1248/bpb.33.341
Kachroo, A., Shanklin, J., Whittle, E., Lapchyk, L., Hildebrand, D., and Kachroo, P. (2007). The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Mol. Biol. 63, 257–271. doi: 10.1007/s11103-006-9086-y
Knutzon, D. S., Thompson, G. A., Radke, S. E., Johnson, W. B., Knauf, V. C., and Kridl, J. C. (1992). Modification of Brassica seed oil by antisense expression of a stearoyl-acyl carrier protein desaturase gene. Proc. Natl. Acad. Sci. U.S.A. 89, 2624–2628. doi: 10.1073/pnas.89.7.2624
Kowalski, S. P., Lan, T. H., Feldmann, K. A., and Paterson, A. H. (1994). Comparative mapping of Arabidopsis thaliana and Brassica oleracea chromosomes reveals islands of conserved organization. Genetics 138, 499–510.
Lagercrantz, U. (1998). Comparative mapping between Arabidopsis thaliana and Brassica nigra indicates that Brassica genomes have evolved through extensive genome replication accompanied by chromosome fusions and frequent rearrangements. Genetics 150, 1217–1228.
Le, B. H., Cheng, C., Bui, A. Q., Wagmaister, J. A., Henry, K. F., Pelletier, J., et al. (2010). Global analysis of gene activity during Arabidopsis seed development and identification of seed-specific transcription factors. Proc. Natl. Acad. Sci. U.S.A. 107, 8063–8070. doi: 10.1073/pnas.1003530107
Li, Y., Beisson, F., Pollard, M., and Ohlrogge, J. (2006). Oil content of Arabidopsis seeds: the influence of seed anatomy, light and plant-to-plant variation. Phytochemistry 67, 904–915. doi: 10.1016/j.phytochem.2006.02.015
Lightner, J., Wu, J., and Browse, J. (1994). A mutant of Arabidopsis with increased levels of stearic acid. Plant Physiol. 106, 1443–1451. doi: 10.1104/pp.106.4.1443
Lindqvist, Y., Huang, W. J., and Schneider, G. J. (1996). Crystal structure of delta9 stearoyl-acyl carrier protein desaturase from castor seed and its relationship to other di-iron proteins. EMBO J. 15, 4081–4092.
Liu, C., Zhou, J., Bracha-Drori, K., Yalovsky, S., Ito, T., and Yu, H. (2007). Specification of Arabidopsis floral meristem identity by repression of flowering time genes. Development 134, 1901–1910. doi: 10.1242/dev.003103
Lu, C. F., Napier, J. A., Clemente, T. E., and Cahoon, E. B. (2011). New frontiers in oilseed biotechnology: meeting the global demand for vegetable oils for food, feed, biofuel, and industrial applications. Curr. Opin. Biotechnol. 22, 252–259. doi: 10.1016/j.copbio.2010.11.006
Osborn, T. C., Kole, C., Parkin, I. A., Sharpe, A. G., Kuiper, M., Lydiate, D. J., et al. (1997). Comparison of flowering time genes in Brassica rapa, B. napus and Arabidopsis thaliana. Genetics 146, 1123–1129.
Parkin, I. A., Sharpe, A. G., Keith, D. J., and Lydiate, D. J. (1995). Identification of the A and C genomes of amphidiploid Brassica napus (oilseed rape). Genome 38, 1122–1131. doi: 10.1139/g95-149
Poirier, Y., Ventre, G., and Caldelari, D. (1999). Increased flow of fatty acids toward beta-oxidation in developing seeds of Arabidopsis deficient in diacylglycerol acyltransferase activity or synthesizing medium-chain-length fatty acids. Plant Physiol. 121, 1359–1366. doi: 10.1104/pp.121.4.1359
Shanklin, J., and Somerville, C. (1991). Stearoyl-acyl-carrier-protein desaturase from higher plants is structurally unrelated to the animal and fungal homologs. Proc. Natl. Acad. Sci. U.S.A. 88, 2510–2514. doi: 10.1073/pnas.88.6.2510
Thompson, G. A., Scherer, D. E., Foxall-Van Aken, S., Kenny, J. W., Young, H. L., Shintani, D. K., et al. (1991). Primary structures of the precursor and mature forms of stearoyl-acyl carrier protein desaturase from safflower embryos and requirement of ferredoxin for enzyme activity. Proc. Natl. Acad. Sci. U.S.A. 88, 2578–2582. doi: 10.1073/pnas.88.6.2578
Keywords: AAD5, seed oil, stearic acid, oleic acid, Arabidopsis thaliana, Brassica napus
Citation: Jin C, Li D, Gao C, Liu K, Qi S, Duan S, Li Z, Gong J, Wang J, Hai J and Chen M (2017) Conserved Function of ACYL–ACYL CARRIER PROTEIN DESATURASE 5 on Seed Oil and Oleic Acid Biosynthesis between Arabidopsis thaliana and Brassica napus. Front. Plant Sci. 8:1319. doi: 10.3389/fpls.2017.01319
Received: 03 May 2017; Accepted: 13 July 2017;
Published: 25 July 2017.
Edited by:
Basil J. Nikolau, Iowa State University, United StatesReviewed by:
Yuanheng Cai, Stony Brook University, United StatesDezhi Wu, Zhejiang University, China
Qian Wang, Zhejiang Wanli University, China
Copyright © 2017 Jin, Li, Gao, Liu, Qi, Duan, Li, Gong, Wang, Hai and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mingxun Chen, Y214Nzg2QG53YWZ1LmVkdS5jbg==
†These authors have contributed equally to this work.