 Selena Gimenez-Ibanez1,2*†
Selena Gimenez-Ibanez1,2*† Dagmar R. Hann1,3,4†
Dagmar R. Hann1,3,4† Jeff H. Chang5,6
Jeff H. Chang5,6 Cécile Segonzac1,7
Cécile Segonzac1,7 Thomas Boller3
Thomas Boller3 John P. Rathjen1,8
John P. Rathjen1,8- 1The Sainsbury Laboratory, Norwich, United Kingdom
- 2Plant Molecular Genetics Department, Centro Nacional de Biotecnología-Consejo Superior de Investigaciones Científicas, Madrid, Spain
- 3Department of Environmental Sciences, Botanical Institute, University of Basel, Basel, Switzerland
- 4Institute of Genetics, Ludwig-Maximilians-Universität München, Munich, Germany
- 5Department of Botany and Plant Pathology, Oregon State University, Corvallis, OR, United States
- 6Center for Genome Research and Biocomputing, Oregon State University, Corvallis, OR, United States
- 7Department of Plant Science, Plant Genomics and Breeding Institute and Research Institute of Agriculture and Life Sciences, College of Agriculture and Life Sciences, Seoul National University, Seoul, South Korea
- 8Research School of Biology, Australian National University, Acton, ACT, Australia
The plant pathogen Pseudomonas syringae injects about 30 different virulence proteins, so-called effectors, via a type III secretion system into plant cells to promote disease. Although some of these effectors are known to suppress either pattern-triggered immunity (PTI) or effector-triggered immunity (ETI), the mode of action of most of them remains unknown. Here, we used transient expression in Nicotiana benthamiana, to test the abilities of type III effectors of Pseudomonas syringae pv. tomato (Pto) DC3000 and Pseudomonas syringae pv. tabaci (Pta) 11528 to interfere with plant immunity. We monitored the sequential and rapid bursts of cytoplasmic Ca2+ and reactive oxygen species (ROS), the subsequent induction of defense gene expression, and promotion of cell death. We found that several effector proteins caused cell death, but independently of the known plant immune regulator NbSGT1, a gene essential for ETI. Furthermore, many effectors delayed or blocked the cell death-promoting activity of other effectors, thereby potentially contributing to pathogenesis. Secondly, a large number of effectors were able to suppress PAMP-induced defense responses. In the majority of cases, this resulted in suppression of all studied PAMP responses, suggesting that these effectors target common elements of PTI. However, effectors also targeted different steps within defense pathways and could be divided into three major groups based on their suppressive activities. Finally, the abilities of effectors of both Pto DC3000 and Pta 11528 to suppress plant immunity was conserved in most but not all cases. Overall, our data present a comprehensive picture of the mode of action of these effectors and indicate that most of them suppress plant defenses in various ways.
Introduction
Plants respond to infections via an innate immune system that employs external and internal receptors (Jones and Dangl, 2006). Primary perception is based on the ability to discriminate between self and non-self. Plants rely on surface-localized pattern recognition receptors (PRRs) to detect conserved molecules called pathogen-associated molecular patterns (PAMPs), or sense wound- and injury-related molecules called danger-associated molecular patterns (DAMPs) to infer the presence of microbes (Boller and Felix, 2009). This recognition leads to PAMP-triggered immunity (PTI), which is considered to be the first level of plant defense, and restricts pathogen infection in most plant species. To overcome such immunity, successful bacterial pathogens use type III secretion systems (TTSSs) to deliver virulence molecules called effector proteins into eukaryotic cells (Collmer et al., 2002). The second branch of the plant innate immune system recognizes type III effectors inside the plant cell via nucleotide-binding leucine-rich repeat (NB-LRR) resistance (R) proteins and is called effector-triggered immunity (ETI) (Chisholm et al., 2006). This specific recognition leads to strong activation of a similar range of plant responses as PTI and is characteristically associated with programmed cell death known as the hypersensitive response (HR) at the infection site.
Pattern recognition receptors are generally membrane-bound receptor kinases (RKs) or receptor proteins (RPs). Ligand perception at the cell surface causes the immediate formation of stable heterodimers comprised of the specific PRR and certain co-receptors. This constitutes an active receptor complex that initiates intracellular signaling (Chinchilla et al., 2007; Heese et al., 2007; Boutrot et al., 2010; Liu et al., 2012; Cao et al., 2014). In Arabidopsis, the paradigm for PAMP sensing is the perception of the bacterial flagellin protein by the leucine-rich repeat (LRR) RK FLS2 (FLAGELLIN-SENSING 2), which triggers formation of an active receptor complex with the LRR-RK co-receptor BAK1 (BRI1-ASSOCIATED KINASE 1) (Chinchilla et al., 2007; Heese et al., 2007). Similarly, chitin oligosaccharides released from fungal cell walls are perceived by the Arabidopsis lysin motif (LysM) RKs LYK5 (LYSIN MOTIF-CONTAINING RECEPTOR-LIKE KINASE 3) and CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1) that form a receptor complex in which both bind chitin (Liu et al., 2012; Cao et al., 2014). Upon PAMP sensing, the activation of different PRR complexes leads to the selective phosphorylation and activation of specific cytoplasmic receptor-like cytoplasmic kinases (RLCKs). These include BIK1 (BOTRYTIS-INDUCED KINASE 1) downstream of flagellin recognition, and PBL27 downstream of chitin perception (Lu et al., 2010; Kadota et al., 2014; Shinya et al., 2014; Yamada et al., 2016).
Activation of PRRs induces a variety of host responses, including evolution of a burst of Ca2+ and reactive oxygen species (ROS), post-translational activation of mitogen-activated protein kinases (MAPK), induction of defense genes, callose deposition into plant cell walls and functional immunity (Kunze et al., 2004; Zipfel et al., 2004; Chinchilla et al., 2006). Upon PAMP stimulation, one of the earliest responses is changes in ion fluxes across the plasma membrane. This includes an influx of Ca2+ from the apoplast within seconds (Chandra et al., 1997; Blume et al., 2000; Lecourieux et al., 2002; Davies et al., 2006; Segonzac et al., 2011). Downstream of the Ca2+ influx, two distinct branches of signaling occur, one leading to ROS production by NADPH oxidases and the other to the activation of MAPKs and transcriptional changes (Segonzac et al., 2011). The plasma membrane Arabidopsis NADPH oxidase RBOHD (RESPIRATORY BURST OXIDASE HOMOLOG D) is part of the PRR complex and is directly regulated through phosphorylation by the RLCK BIK1 in a calcium-independent manner. This leads to ROS production and antibacterial immunity (Kadota et al., 2014; Li et al., 2014). In contrast, the RLCK PBL27 directly connects the PRR CERK1 with the MAPK kinase kinase MAPKKK5, establishing a link between PRRs and activation of intracellular MAPK cascades (Yamada et al., 2016). However, the fact that the influx of Ca2+ across the plasma membrane is nevertheless required for the PAMP-induced ROS burst and MAPK activation (Segonzac et al., 2011; Kadota et al., 2014), suggests that calcium-based regulation is also required for the ultimate activation of RBOHD and MAPKKK5. Successful initiation of signaling leads to characteristic transcriptional reprogramming. Microbial perception alters the expression of approximately 10% of the whole plant transcriptome (Navarro et al., 2004; Moore et al., 2011). This results in the production of antimicrobial compounds and cell wall-reinforcing materials around the infection site (Hahlbrock et al., 2003), which is supposed to be critical for plant immunity.
Type III effectors contribute collectively to bacterial pathogenesis by targeting host defense pathways (Katagiri et al., 2002). Effectors hijack multiple cellular processes including phytohormone signaling, proteasome-dependent protein degradation, cytoskeleton formation, manipulation of stomatal openings, establishment of intercellular apoplastic living spaces, and vesicle transport (Buttner, 2016; Toruno et al., 2016). Among all these functions, suppression of PTI has emerged as a primary role of bacterial effectors to ensure pathogenesis. Multiple effectors directly target PRRs, their co-receptors, RLCKs and MAPKs in a redundant manner and by multiple strategies to ensure suppression (Macho and Zipfel, 2015; Buttner, 2016). Some examples include the effectors AvrPto, AvrPtoB, HopAO1, and HopF2, which act as direct suppressors of PRRs such as FLS2 and/or their co-receptors BAK1 and CERK1; HopAR1 that targets BIK1 and other RLCKs; and HopAI1 and HopF2 that suppress MAPK signaling by directly modifying these proteins (Macho and Zipfel, 2015; Buttner, 2016; Toruno et al., 2016). Despite the success in identifying effectors using functional and genetic analysis, the modes of action of most of them are yet unknown.
The Gram-negative bacterial species Pseudomonas syringae (P. syringae) comprises at least 50 pathovars that can be distinguished by their host ranges (Sawada et al., 1999). For example, P. syringae pv. tomato (Pto) DC3000 infects tomato and can cause disease on Arabidopsis whereas P. syringae pv. tabaci (Pta) 11528 is infectious on Nicotiana tabacum and can grow to high levels on the model species Nicotiana benthamiana (N. benthamiana). The Arabidopsis-Pto DC3000 system represents the primary model for plant–bacteria interactions (Katagiri et al., 2002). In contrast, the Pta 11528-N. benthamiana system represents an interaction that offers complementary benefits to Arabidopsis, including high amenability to Agrobacterium tumefaciens-mediated transient expression of foreign genes and virus-induced gene silencing (VIGS) for gene knockdowns (Goodin et al., 2008). Pto DC3000 encodes nearly 30 effectors whereas Pta 11528 carries approximately 20 genes with homology to previously described effectors (Chang et al., 2005; Vinatzer et al., 2005; Ferreira et al., 2006; Lindeberg et al., 2006; Schechter et al., 2006; Studholme et al., 2009). Several of these Pto DC3000 effectors are conserved in Pta 11528 but it is currently unknown whether they function similarly to their Pto DC3000 homologs. We currently lack an integrated view of which defense pathways are suppressed by the effector repertoire, how many effectors act redundantly, and the conservation of effector function among homologs in different Pseudomonas strains. Also, although most of the current work has focused on defense responses elicited by flagellin, Pseudomonas contains many more PAMPs which may stimulate alternate pathways. It is not clear whether different PRRs feed into a limited number of signal transduction pathways, or if effectors target certain pathways specifically or act more broadly. Thus, we require a more integrative view of global molecular activities and the range of suppression by bacterial effectors to understand how host processes normally prevent successful infection.
In this study, we transiently expressed type III effector genes in leaves of N. benthamiana to screen and identify those with the ability to suppress defense responses. We analyzed nearly all Pto DC3000 effectors and most Pta 11528 secreted proteins for the ability to interfere with effector induced cell death, a typical hallmark of effector recognition by plant R genes. Secondly, we analyzed their capacities to abolish a broad range of PAMP responses including changes in cytoplasmic Ca2+ levels, production of ROS and induction of defense genes. Strikingly, among the 32 effectors tested, 29 showed suppressive activity in at least one of our assays. This observation is consistent with the hypothesis that one of the most important roles of effector proteins is to suppress host immunity. This work helps to identify P. syringae effectors that are capable of suppressing plant defenses, and to define the spectrum of their activities.
Materials and Methods
Elicitors
Crab shell chitin and flg22 peptide (CKANSFREDRNEDREV) were purchased from Sigma (United Kingdom) and Peptron (South Korea), respectively. Pto DC3000, Pta 11528, and A. tumefaciens (strain C58C1) were grown in L-medium at 28°C on a rotary shaker. Crude A. tumefaciens, Pto DC3000 and Pta 11528 bacterial suspensions were prepared by centrifuging overnight cultures and resuspending bacterial pellets in water.
Statistical Methods
Statistical significance based on t-test analysis was developed by the GraphPad Prism program. Eight independent samples were used to analyze the significance of ROS and Ca2+ generation assays. For quantitative gene expression analysis, statistical significance was calculated from three independent samples.
Bacterial Strains
Agrobacterium tumefaciens strain C58C1 was used for transient assays transformed with pGWB14 (Pto DC3000 and Pta 11528 effector libraries) (Nakagawa et al., 2007). Both the Pto DC3000 and Pta 11528 effector libraries were described previously (Chang et al., 2005; Gimenez-Ibanez et al., 2014). The Pta 11528 effector library contains individual effector genes under control of the 35S promoter fused in-frame to a sequence encoding three C-terminal hemagglutinin (HA) epitope tags (Gimenez-Ibanez et al., 2014). In contrast, the full-length Pto DC3000 effectors used in this study were recombined into pDONR207 (Invitrogen) (Chang et al., 2005). These plasmids, a Gateway BP II kit (Invitrogen), and the pGWB14 (Nakagawa et al., 2007) destination vector were used to generate expression constructs in which each Pto DC3000 effector gene is under the control of the 35S promoter and fused to a sequence encoding three HA epitope tags. These constructs were verified by DNA sequencing and then transferred to A. tumefaciens strain C58C1. Additional bacterial strains used in this study were Pseudomonas syringae pv. tomato (Pto) DC3000, Pto DC3000 hrcC, Pto DC3000ΔavrPto, Pto DC3000ΔavrPtoB, and Pto DC3000ΔavrPtoΔavrPtoB (Lin and Martin, 2005).
A. tumefaciens-Mediated Transient Expression Assays
For transient gene expression, A. tumefaciens C58C1 was syringe infiltrated in N. benthamiana leaves at OD600 = 0.3–0.5 in 10 mM MgCl2 and 10 mM MES. Samples were collected 2 days post-inoculation for ROS, Ca2+, MAPK and gene expression assays. For co-infiltration assays, each of the two A. tumefaciens strains containing the respective effector gene were prepared at OD600 = 0.5 in 10 mM MgCl2 and 10 mM MES and then mixed in equal volumes. This solution was infiltrated in N. benthamiana leaves and phenotypes were scored between 1 to 7 days post-inoculation as indicated in the text.
Virus-Induced Gene Silencing (VIGS)
Virus-induced gene silencing was performed using a tobacco rattle virus vector as previously described (Peart et al., 2002).
Measurement of Reactive Oxygen Species (ROS) Generation
Leaf disks (0.38 cm2) were floated on water overnight and ROS released by the leaf tissue were measured using a chemiluminescent assay (Keppler et al., 1989). The water was replaced with 200 μl of a solution containing 20 μM luminol (Sigma, St. Louis, MO, United States) and 1 μg of horseradish peroxidase (Fluka, Buchs, Switzerland). ROS was elicited with 100 nM flg22 or 100 μg/ml of chitin in all experiments. Elicitation in the absence of any PAMP (water treatment) was included as a negative control. Luminescence was measured over a time period of 30 min using the Photek camera system (East Sussex, United Kingdom), and data were recorded as total counts.
Measurement of Ca2+ Burst Generation
Transgenic N. benthamiana plants expressing the Ca2+ sensor 35S:Aequorin (N. benthamiana SLJR15) were used to measure intracellular Ca2+ concentrations (Segonzac et al., 2011). Leaf disks (0.38 cm2) were floated overnight on an aqueous 2.5 μM coelenterazine solution in the dark at room temperature. The Ca2+ influx was elicited with water, flg22 (100 nM) or chitin (100 μg/ml), and luminescence was measured using the Photek camera system as total counts over a time period of 30 min.
Quantitative RT-PCR
Nicotiana benthamiana leaf disks expressing each individual effector were collected 2 days post-infiltration (dpi), and then floated overnight in water. Leaf disks were subsequently elicited with water (Mock EV control), 100 nM flg22 or 100 μg/ml chitin for 60 min and frozen in liquid nitrogen. Total RNA was extracted by using TRIzol-Reagent (Sigma), and the absence of genomic DNA was checked by PCR amplification of the housekeeping NbEF1α gene by using 2.5 μg of RNA. For analysis of gene expression, first-strand cDNA was synthesized from 2.5 μg of RNA using SuperScript RNA H-Reverse Transcriptase (Invitrogen, United Kingdom) and an oligo (dT) primer, according to the manufacturer’s instructions. For quantitative PCR, 2 μl of cDNA was combined with SYBR master mix. PCRs were performed in triplicate with a PTC-200 Peltier Thermal Cycler (MJ Research, Waltham, MA, United States), and the data were collected and analyzed with Chromo 4 Continuous Fluorescence detection system. The NbEF1α RNA was analyzed as an internal control and used to normalize the values for transcript abundance. All samples were related to the Mock EV negative control. Primers for genes used here are as follows: NbCyp71D20, 5′-AAGGTCCACCGCACCATGTCCTTAGAG-3′ and 5′-AAGAATTCCTTGCCCCTTGAGTACTTGC-3′; NbACRE132, 5′-AAGGTCCAGCGAAGTCTCTGAGGGTGA-3′ and 5′-AAGAATTC-CAATCCTAGCTCTGGCTCCTG-3′; and NbEF1α gene 5′-AAGGTCCAGTATGCCTGGGTGCTTGAC-3′ and 5′-AAGAATTCACAGGGACAGTTCCAATACCA-3′.
Cell Death Assays on N. benthamiana
For elicitation of the HR by different P. syringae pathovars, overnight bacterial cultures were pelleted, resuspended in sterile 10 mM MgCl2 and infiltrated into the leaves of transgenic N. benthamiana plants at high densities (5 × 107 cfu/ml). The development of cell death was scored between 1 to 4 dpi.
Results
Construction of Effector Libraries for Pto DC3000 and Pta 11528
Pto DC3000 and Pta 11528 encode nearly 30 and 20 effectors, respectively (Chang et al., 2005; Vinatzer et al., 2005; Ferreira et al., 2006; Lindeberg et al., 2006; Schechter et al., 2006; Studholme et al., 2009). To investigate the functions of most of these, we analyzed two libraries containing 22 Pto DC3000 and 10 Pta 11528 effector genes, respectively. Both libraries were described previously (Chang et al., 2005; Gimenez-Ibanez et al., 2014) and express each effector from a T-DNA under control of the strong 35S promoter, fused in frame to a sequence encoding three HA epitope tags. There were seven pairs of homologous effectors shared between the Pto DC3000 and Pta 11528 libraries sampled here, with varying levels of amino acid conservation. Based on protein sequence identity to their Pto DC3000 homologs, some Pta 11528 effectors were almost identical, namely HopO1-1Pta11528 (99%, Supplementary Figure S1) and HopT1-1Pta11528 (99%, Supplementary Figure S2); others showed high levels of identity such as AvrPtoBPta11528 (70%, Supplementary Figure S3) and HopX1Pta11528 (72%, Supplementary Figure S4); whereas another group including AvrPtoPta11528 (42%, Supplementary Figure S5), HopI1Pta11528 (55%, Supplementary Figure S6) and HopF1Pta11528 (49%, Supplementary Figure S7) showed significantly less homology with their respective Pto DC3000 homologs. A complete list of the effector genes used in this study is in Table 1.

TABLE 1. List of Pto DC3000 and Pta 11528 effectors included in this study.
Several Pto DC3000 and Pta 11528 Effectors Elicit Cell Death in N. benthamiana
Mutations affecting individual effector genes typically have no or only subtle effects on bacterial pathogenicity, due to redundancy among effectors (Kvitko et al., 2009). To overcome this, we used Agrobacterium tumefaciens to transiently express individual effector genes in N. benthamiana (Hann and Rathjen, 2007). Western blot analysis showed that most effector proteins accumulated to detectable levels in N. benthamiana (Table 1 and Supplementary Figure S8). We detected 25 out of 32 proteins at 2 dpi. An additional seven effectors that could not be detected by western blots suppressed defense responses in at least one assay performed in this work, indicating that these proteins were expressed at undetectable levels in the plant cell. Overall, there is evidence that all Pto DC3000 and Pta 11528 effector proteins accumulated within the plant cell after transient expression.
Four Pto DC3000 and three Pta 11528 effectors elicited various forms of necrosis when transiently expressed in N. benthamiana leaves, whereas no phenotype was observed in control tissue expressing an EV construct (Figure 1A). Leaves when transiently expressing hopAD1PtoDC3000, avrE1Pta11528, hopW1-1Pta11528, or hopT1-1Pta11528 displayed strong cell death within two or three dpi. However, expression of avrE1Pta11528 induced a stronger and faster necrosis compared to expression of hopAD1PtoDC3000, hopW1-1Pta11528, and hopT1-1Pta11528. In contrast, expression of hopM1PtoDC3000 induced only a patchy necrosis within 3 to 4 dpi, whereas hopAA1-1PtoDC3000 led to appearance of shiny areas, obvious only on the abaxial leaf surface that never developed into full necrosis. Leaves expressing hopQ1-1PtoDC3000 displayed only mild chlorosis within 4 days. In our experiments, such chlorotic areas did not normally progress further into necrosis. Finally, we observed mild chlorotic areas associated with avrPtoBPta11528 and hopAR1Pta11528 expression that appeared consistently at 7 dpi (data not shown). Effector protein accumulation did not correlate with the intensity of necrosis. For example, neither AvrE1Pta11528 nor HopAD1PtoDC3000 proteins could be detected by western blots, but they both induced fast and strong necrosis. Overall, 7 of 32 effector genes caused cell death when expressed transiently in N. benthamiana leaves.
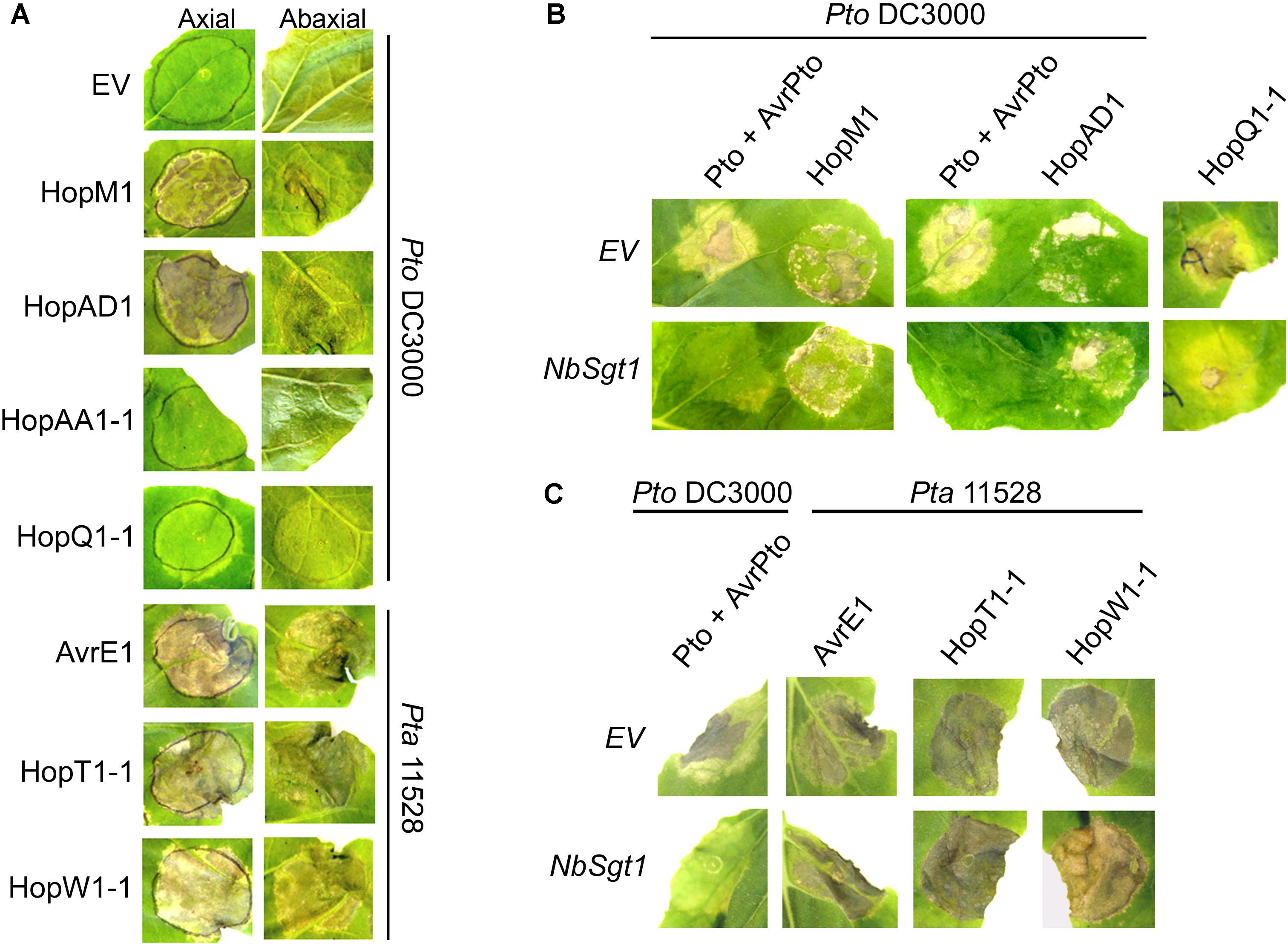
FIGURE 1. Several Pto DC3000 and Pta 11528 type-III effectors induce cell death in N. benthamiana leaves in an NbSgt1-independent manner. (A) Ability of Pto DC3000 and Pta 11528 effectors to elicit cell death in N. benthamiana when delivered by A. tumefaciens. Four-week-old N. benthamiana leaves were syringe-infiltrated with an A. tumefaciens C58C1 strain expressing an EV or each individual effector gene driven by the 35S promoter. Pictures were taken 4 days after inoculation from both sides of the plant leaf (axial and abaxial). All effector genes were tested but those that didn’t show a response are not represented. Similar results were obtained in two independent experiments. (B) Silencing of NbSgt1 does not compromise cell death induced by hopM1PtoDC3000 and hopAD1PtoDC3000. Four-week-old N. benthamiana leaves silenced for EV or NbSgt1 with a tobacco rattle virus (TRV) vector were syringe-infiltrated with A. tumefaciens C58C1 strains individually expressing hopM1PtoDC3000 or hopAD1PtoDC3000 effectors. hopQ1-1PtoDC3000, or avrPtoPtoDC3000 co-expressed with Pto, were included on the same leaf as positive controls for NbSgt1 silencing. Pictures were taken 4 days after infiltration for hopM1PtoDC3000, hopAD1PtoDC3000, and Pto/avrPtoPtoDC3000, and 8 days after infiltration for hopQ1-1PtoDC3000. Similar results were obtained in three independent experiments. (C) Silencing of NbSgt1 does not compromise cell death induced by avrE1Pta11528, hopT1-1Pta11528, and hopW1-1Pta11528 when transiently expressed in N. benthamiana. Four-week-old N. benthamiana leaves silenced for EV and NbSgt1 with a TRV vector were syringe-infiltrated with A. tumefaciens C58C1 strains expressing one of avrE1Pta11528, hopT1-1Pta11528, or hopW1-1Pta11528 effector genes. Co-infiltrated avrPtoPtoDC3000 and Pto were included on the same leaf as a positive control for NbSgt1 silencing. Pictures were taken 4 days after inoculation. Similar results were obtained in three independent experiments.
Plant cell death is associated with effector recognition and subsequent HR in resistant plants as well as with the formation of lesions in susceptible plants (Alfano and Collmer, 2004). To investigate the nature of the cell death phenotype caused by individual expression of several Pto DC3000 and Pta 11528 effectors, we used VIGS to knock down the expression of NbSgt1 in N. benthamiana. NbSgt1 is required for NB-LRR proteins to trigger the defense-associated HR in response to effector recognition (Peart et al., 2002). Three weeks post-silencing, we transiently expressed hopM1PtoDC3000, hopAD1PtoDC3000, hopQ1-1PtoDC3000, avrE1Pta11528, hopT1-1Pta11528, and hopW1-1Pta11528 in EV- or NbSgt1-silenced plants. Because of the difficulty in scoring the mild phenotype of hopAA1-1PtoDC3000 expression, we excluded this gene from the assay. In addition, we co-expressed avrPtoPtoDC3000 and the protein kinase gene Pto as a control. Pto in complex with the NB-LRR protein Prf recognizes AvrPtoPtoDC3000 in vivo, leading to induction of a NbSgt1-dependent HR (Scofield et al., 1996; Tang et al., 1996; Peart et al., 2002; Mucyn et al., 2006). As expected, silencing of NbSgt1 compromised the cell death phenotype developed by the AvrPtoPtoDC3000/Pto control in N. benthamiana, and also by hopQ1-1PtoDC3000, which was reported previously to be NbSgt1-dependent (Wei et al., 2007). In contrast, cell death induced by any of the Pto DC3000 or Pta 11528 effectors was unaffected in both EV- and NbSgt1-silenced plants (Figures 1B,C). Thus, the necrosis induced by hopM1PtoDC3000, hopAD1PtoDC3000, avrE1Pta11528, hopT1-1Pta11528, and hopW1-1Pta11528 was not due to recognition by plant R genes. Taken together, these results suggest that NbSgt1 does not play a role in the cell death induced by these effectors.
Pto DC3000 and Pta 11528 Effectors Redundantly Suppress NbSgt1-Independent Effector-Induced Cell Death
Some effector proteins can mask the ability of other effectors to trigger cell death (Jackson et al., 1999; Jamir et al., 2004; Guo et al., 2009). We investigated whether the cell death phenotypes induced by certain Pto DC3000 or Pta 11528 effectors could be suppressed by others encoded by these pathovars. We transiently co-delivered the necrosis-inducing effector hopAD1PtoDC3000 with each of the remaining effectors of the Pto DC3000 library. Among the 21 genes screened, 8 effectors completely blocked hopAD1PtoDC3000 induced cell death, whereas 7 additional genes significantly delayed the timing of necrosis appearance (Figure 2A). Effector genes that completely blocked cell death included avrPtoBPtoDC3000, hopT1-1PtoDC3000, hopF2PtoDC3000, hopK1PtoDC3000, hopQ1-1PtoDC3000, hopV1PtoDC3000, hopY1PtoDC3000, and hopD1PtoDC3000. In addition, effector genes that delayed the appearance of cell death were avrPtoPtoDC3000, hopO1-1PtoDC3000, hopA1PtoDC3000, hopI1PtoDC3000, hopG1PtoDC3000, hopN1PtoDC3000, and hopX1PtoDC3000. These effectors interfered not only with hopAD1PtoDC3000 but also with hopM1PtoDC3000 induced necrosis, with the exception of hopX1PtoDC3000 (Supplementary Figure S9). To test whether a similar activity is present in the Pta 11528 effector inventory, we screened these genes for the capacity to suppress hopT1-1Pta11528 induced cell death. Similar to previous results, most Pta 11528 effectors interfered with necrosis induced by this protein (Figure 2B). Expression of avrPtoBPta11528, hopF1Pta11528, hopX1Pta11528 completely suppressed hopT1-1Pta11528 induced cell death, whereas avrPtoPta11528, hopO1-1Pta11528, hopI1Pta11528, and hopAR1Pta11528 delayed it. We next investigated whether the cell death phenotypes induced by the Pto DC3000 effectors hopAD1 and hopM1 could be suppressed by an effector encoded within the Pta 11528 repertoire, such as avrPtoB. The effector avrPtoBPta11528 also interfered with the cell death phenotypes induced by both hopAD1PtoDC3000 and hopM1PtoDC3000 (Supplementary Figure S10). Therefore, our data indicate that the vast majority of Pto DC3000 and Pta 11528 effectors can interfere with cell death induced by certain effectors within the same bacterial pathovar and in the case of avrPtoBPta11528, also across bacterial pathovars.
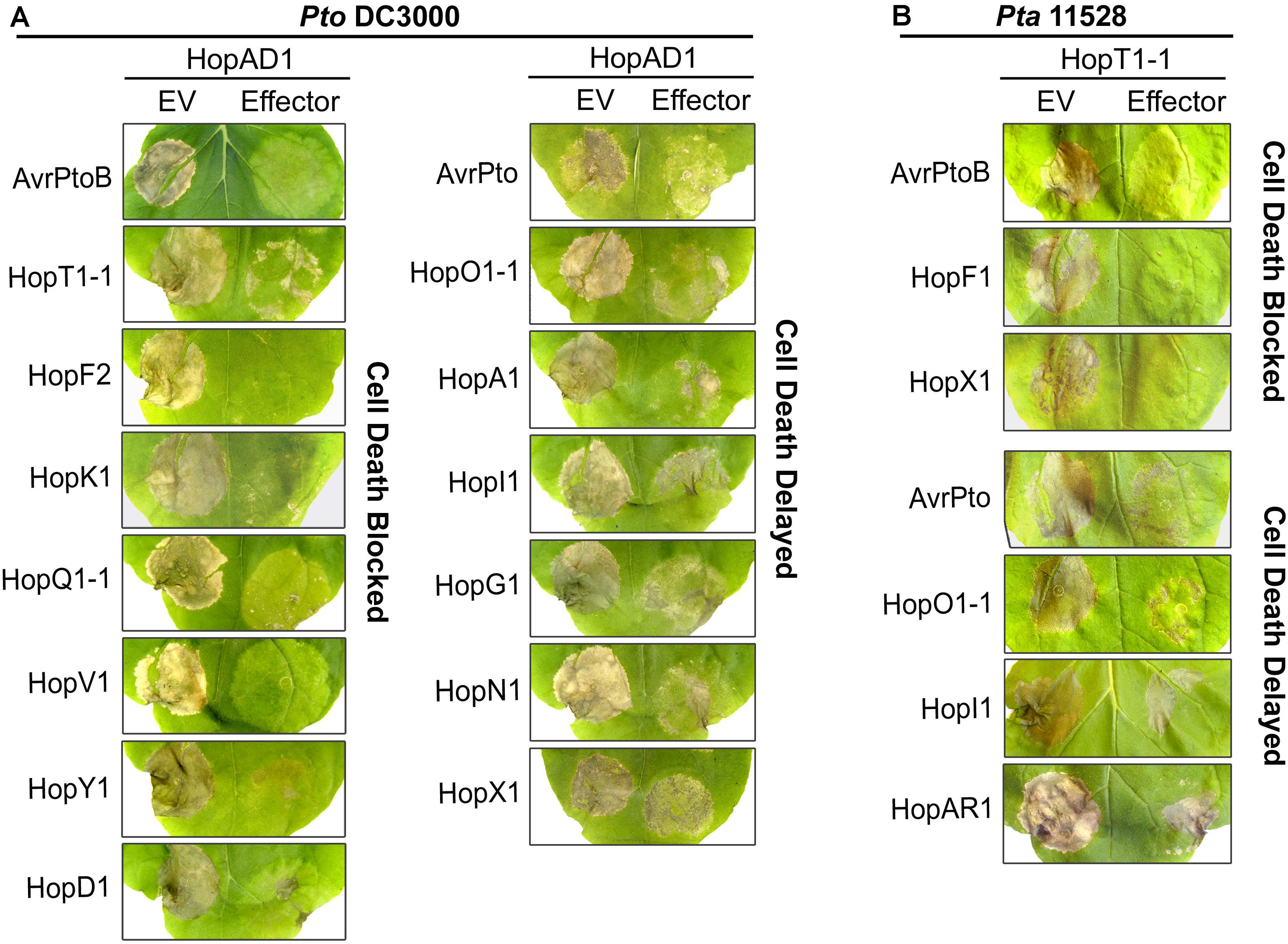
FIGURE 2. Most Pto DC3000 and Pta 11528 effectors can interfere with effector-induced cell death. (A) Suppression of hopAD1PtoDC3000 induced cell death by Pto DC3000 effectors. The effector hopAD1PtoDC3000 was co-expressed in N. benthamiana leaves with each effector in the Pto DC3000 repertoire, or an EV control. Each of the two A. tumefaciens strains containing the respective effector gene or an EV control were prepared at OD600 = 0.5 then mixed in equal volumes. Cell death induced by hopAD1PtoDC3000 was scored 4 days after infiltration. Similar results were obtained in three independent experiments. (B) Suppression of hopT1-1Pta11528 induced cell death by the Pta 11528 effector repertoire. hopT1-1Pta11528 was co-expressed with each individual effector in the Pta 11528 repertoire or an EV control in N. benthamiana. Pictures were taken 4 days after inoculation and similar results were obtained in three independent experiments.
We next analyzed the ability of Pto DC3000 effectors to antagonize specific R gene-dependent HR events in N. benthamiana. In tomato, the unrelated effectors AvrPtoPtoDC3000 and AvrPtoBPtoDC3000 trigger disease resistance in plants carrying Pto and Prf (Scofield et al., 1996; Tang et al., 1996; Kim et al., 2002; Mucyn et al., 2006). Thus, we next assessed the ability of transgenic N. benthamiana plants expressing the tomato Pto and Prf genes (N. benthamiana R411A) (Balmuth and Rathjen, 2007) for the ability to trigger HR upon infiltration of high bacterial densities of Pto DC3000, or isogenic strains lacking avrPtoPtoDC3000 (Pto DC3000ΔavrPto), avrPtoBPtoDC3000 (Pto DC3000ΔavrPtoB) or both (Pto DC3000ΔavrPtoΔavrPtoB). We additionally included a Pto DC3000 strain lacking a functional TTSS required for effector secretion as a negative control (Pto DC3000 hrcC). Three days post-inoculation, Pto DC3000, Pto DC3000ΔavrPto, and Pto DC3000ΔavrPtoB elicited strong HRs (Supplementary Figure S11), indicating that the effectors secreted by Pto DC3000 do not interfere with the AvrPtoPtoDC3000 and/or AvrPtoBPtoDC3000-dependent recognition by Pto/Prf.
Multiple Pto DC3000 and Pta 11528 Effectors Target PAMP(s) Signaling Pathways at Different Steps
To identify suppressive effectors, we screened our collection for the ability to suppress the PAMP-dependent Ca2+ and ROS bursts, and activation of defensive marker genes in N. benthamiana. For these experiments, we used two PAMPs; the flg22 peptide derived from bacterial flagellin (Chinchilla et al., 2007), and the oligosaccharide chitin which is a component of fungal cell walls. These two PAMPs are structurally unrelated and perceived by different receptor complexes (Boutrot and Zipfel, 2017). Each effector was expressed individually in N. benthamiana for 2 days and the harvested tissue subjected to treatments with either flg22 or chitin followed by one of the three assays. The first was a ROS assay done in wild-type (WT) N. benthamiana leaves. The second assayed for an increase in cytosolic Ca2+ concentrations using transgenic N. benthamiana plants expressing the 35S:Aequorin reporter (N. benthamiana SLJR15) (Segonzac et al., 2011). Finally, we assayed induction of the well-characterized defense genes NbCyp71D20 and NbACRE132 by quantitative RT-PCR (Navarro et al., 2004; Segonzac et al., 2011). The majority of Pto DC3000 and Pta 11528 effectors interfered with PAMP-induced responses and could be divided into three major groups based on their range of suppressive activities. Firstly, broad-range suppressors of all tested PAMP-induced early responses (Group A). Secondly, suppressors of the PAMP-induced ROS burst, without affecting Ca2+ influx (Group B). And finally, effectors that suppressed PAMP-induced transcriptional activation of defense genes without affecting other outputs (Group C).
Group A: Broad-Range Suppressors of PAMP-Induced Early Responses
As expected, leaf tissue expressing an EV control and treated with either flg22 or chitin led to an increase in cytosolic Ca2+ levels (Figure 3A), the production of ROS (Figure 3B) and the strong induction of both NbCyp71D20 and NbACRE132 defensive marker genes (Figures 3C,D). We found that six Pto DC3000 and four Pta 11528 effectors compromised the activation of all tested responses to both PAMPs simultaneously (Figure 3). These effectors included the Pto DC3000 effectors AvrPtoPtoDC3000, AvrPtoBPtoDC3000, HopM1PtoDC3000, HopAD1PtoDC3000, HopAA1-1PtoDC3000 and HopQ1-1PtoDC3000, and the Pta 11528 effectors AvrPtoPta11528, AvrPtoBPta11528, HopT1-1Pta11528, and HopW1-1Pta11528. AvrEPta11528 was omitted from these experiments because of the strong cell death phenotype caused by its expression. Interestingly, in contrast to the other effectors of this group, AvrPtoPtoDC3000 and AvrPtoPta11528 only mildly suppressed or had no effect on chitin-induced defense responses, while they strongly compromised all flg22-induced outputs. Overall, 10 effectors from Pto DC3000 and Pta 11528 acted as early broad-range suppressors of PAMP-induced responses. These results suggest that multiple specificities and strategies may converge for simultaneous suppression of early PAMP-triggered immunity elicited by multiple microbial elicitors.
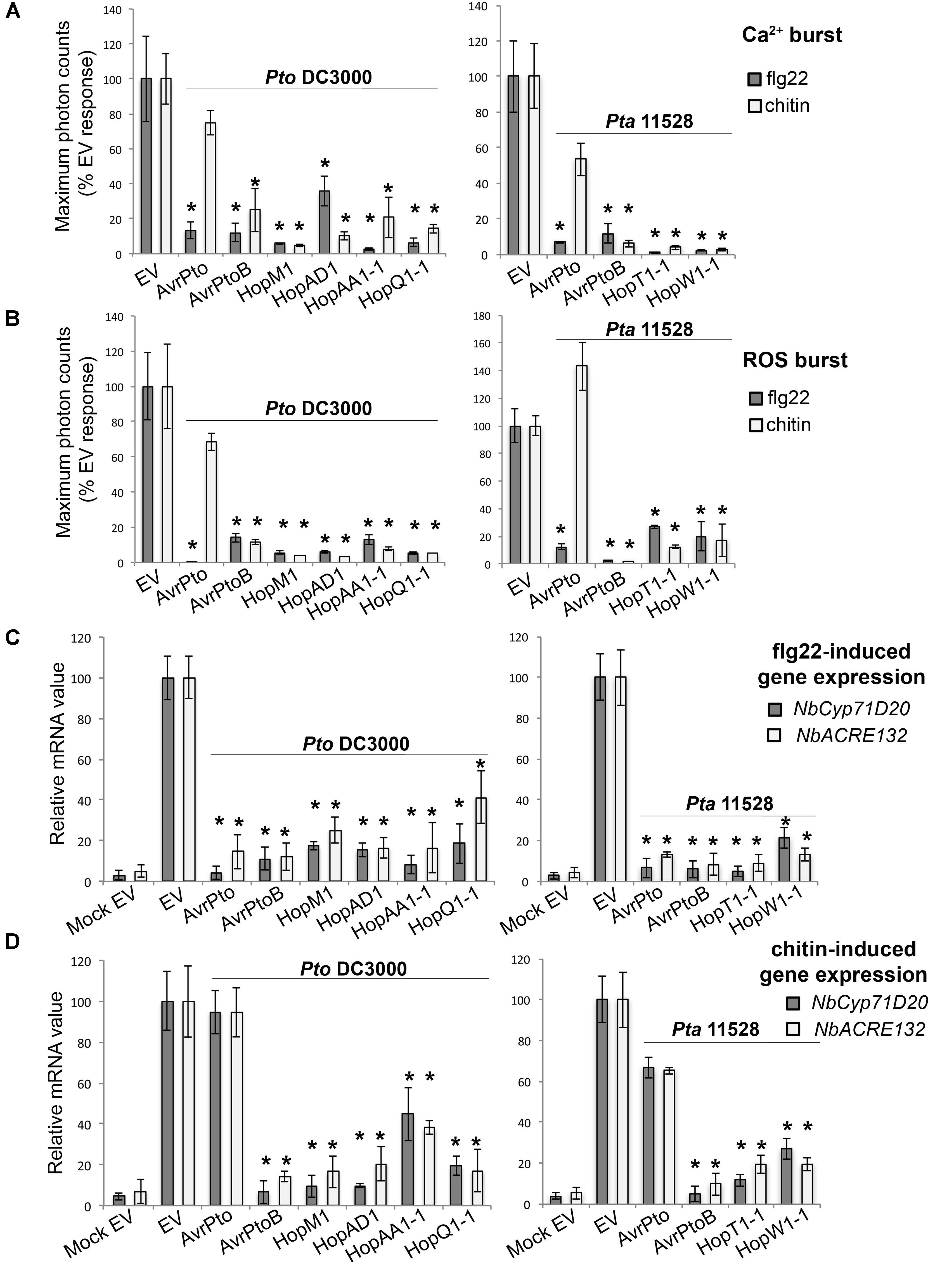
FIGURE 3. Group A effectors: broad-range suppressors of early PAMP-induced responses. (A) Ca2+ burst in N. benthamiana SLJR15 transgenic plants transiently expressing an EV or each Group A effector as indicated after treatment with 100 nM flg22 or 100 μg/ml chitin. Data are presented relative (%) to control EV (PAMP-treated). Error bars represent the standard error of the mean (SEM; n = 8). Statistical significance compared to N. benthamiana tissue transiently expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). (B) ROS burst in N. benthamiana plants transiently expressing an EV or each Group A effector from Pto DC3000 or Pta 11528 after treatment with 100 nM flg22 or 100 μg/ml chitin. Data are presented relative (%) to control EV (PAMP-treated). Error bars represent the standard error of the mean (SEM; n = 8). Statistical significance compared to N. benthamiana tissue transiently expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). (C) Quantitative RT-PCR analysis of NbCyp71D20 and NbACRE132 gene expression 60 min after treatment with 100 nM flg22 in N. benthamiana leaf tissue transiently expressing an EV control or each Group A effector protein as indicated. A mock induction treatment in N. benthamiana leaf tissue transiently expressing EV (Mock EV) is included as a negative control. All samples were normalized against the housekeeping gene NbEF1α and the measurements represent the ratio of expression levels (%) compared to the flg22-induced EV sample. Error bars represent (SEM; n = 3). Statistical significance compared to PAMP-induced N. benthamiana tissue expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). (D) Quantitative RT-PCR analysis of NbCyp71D20 and NbACRE132 defense gene expression 60 min after treatment with 100 μg/ml chitin in N. benthamiana leave tissue transiently expressing an EV control or each Group A effector protein as indicated. A mock induction treatment in N. benthamiana leaf tissue transiently expressing EV (Mock EV) is included as a negative control. All samples were normalized against the housekeeping gene NbEF1α and the measurements represent the ratio of expression levels (%) compared to the chitin-induced EV sample. Error bars represent (SEM; n = 3). Statistical significance compared to PAMP-induced N. benthamiana tissue expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). The results shown in (A–D) are representative of three independent experiments. All effector genes were tested but those that didn’t show any response of suppression are not represented.
Group B: Suppressors of PAMP-Induced ROS Burst
Apoplastic generation of ROS upon microbial perception plays a key role in the activation of disease resistance mechanisms in plants (Yoshioka et al., 2008). We found three effectors, HopT1-1PtoDC3000, HopX1Pta11528, and HopAR1Pta11528 that suppressed PAMP-induced ROS production but not the Ca2+ influx (Figures 4A,B). HopT1-1PtoDC3000 did not distinguish between flg22 and chitin, whereas HopX1Pta11528 and HopAR1Pta11528 suppressed chitin-induced ROS more efficiently than flg22 ROS. Only HopT1-1PtoDC3000 suppressed induction of the defense markers NbCyp71D20 and NbACRE132 by both elicitors, whereas HopX1Pta11528 and HopAR1Pta11528 did not suppress defense gene induction (Figures 4C,D). We designated these Group B effectors, based on their ability to suppress the ROS burst upon PAMP perception without interfering with the Ca2+ influx. The specificity of HopX1Pta11528 and HopAR1Pta11528 for suppression of chitin-induced ROS suggests that the chitin and flg22 perception systems may differ in some key components required for ROS production. It is remarkable that HopT1-1PtoDC3000 and HopX1Pta11528 behaved differently from their respective Pta 11528 and Pto DC3000 homologs. Compared to HopT1-1PtoDC3000, HopT1-1Pta11528 acted as a broad-range suppressor of early PAMP-induced responses (Group A), whereas HopX1PtoDC3000 did not compromise the chitin-triggered ROS burst that was typically suppressed in the presence of HopX1Pta11528. The capacity of several effectors to suppress ROS production upon PAMP sensing supports the importance of apoplastic ROS production in immunity to bacterial pathogens.
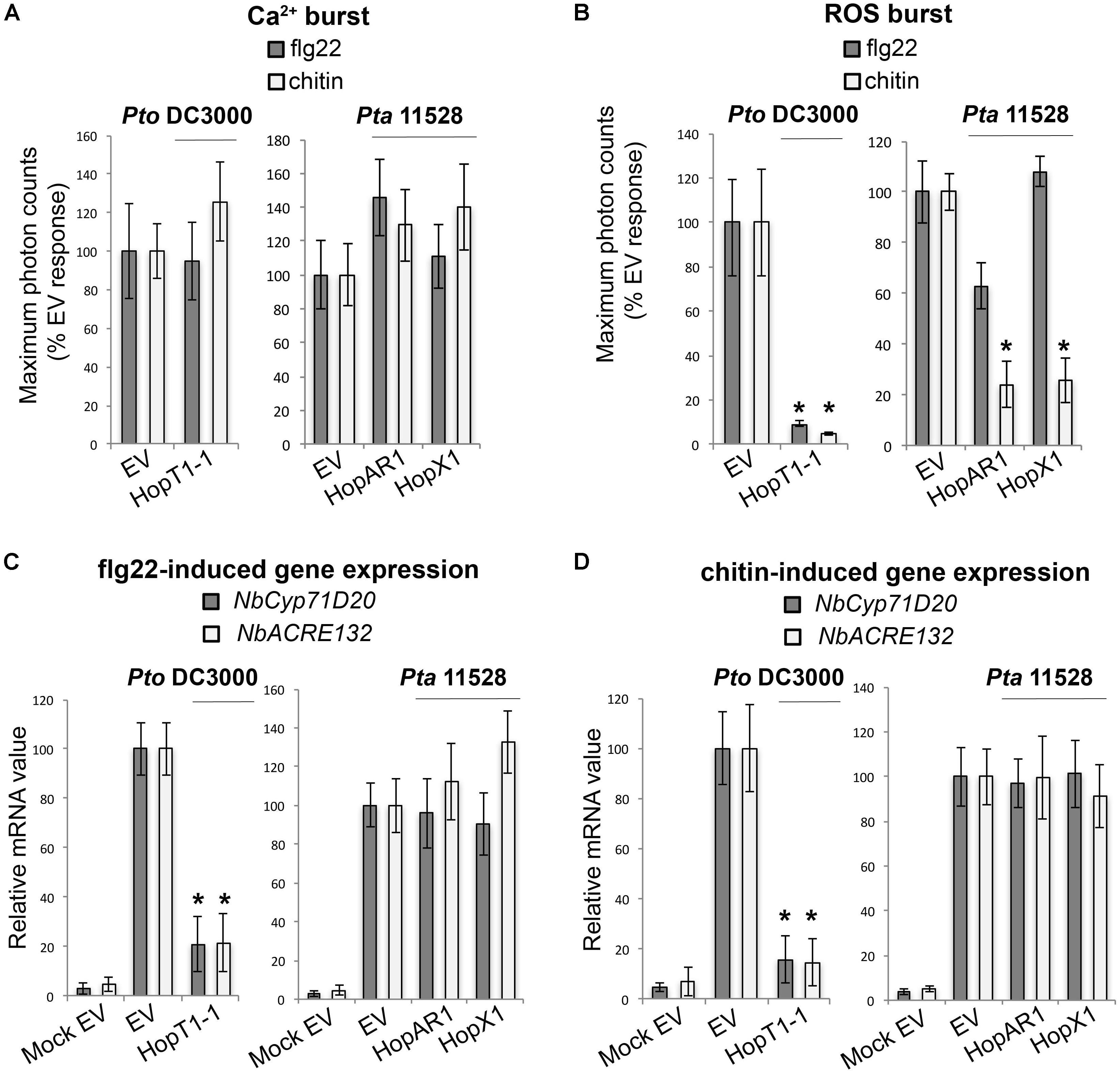
FIGURE 4. Group B effectors: suppressors of PAMP-induced ROS burst. (A) Ca2+ burst in N. benthamiana SLJR15 transgenic plants transiently expressing an empty EV or each Group B effector as indicated after treatment with 100 nM flg22 or 100 μg/ml chitin. Data are presented relative (%) to control EV (PAMP-treated). Error bars represent the standard error of the mean (SEM; n = 8). Statistical significance compared to N. benthamiana tissue transiently expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). (B) ROS burst in N. benthamiana plants transiently expressing an EV or each Group B effector as indicated after treatment with 100 nM flg22 or 100 μg/ml chitin. Data are presented relative (%) to control EV (PAMP-treated). Error bars represent the standard error of the mean (SEM; n = 8). Statistical significance compared to N. benthamiana tissue transiently expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). (C) Quantitative RT-PCR analysis of NbCyp71D20 and NbACRE132 defense gene expression 60 min after treatment with 100 nM flg22 in N. benthamiana leaf tissue transiently expressing an EV control or each Group B effector as indicated. A mock induction treatment in N. benthamiana leaf tissue transiently expressing EV (Mock EV) is included as a negative control. All samples were normalized against the housekeeping gene NbEF1α and the measurements represent the ratio of expression levels (%) compared to the flg22-induced EV sample. Error bars represent (SEM; n = 3). Statistical significance compared to PAMP-induced N. benthamiana tissue expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). (D) Quantitative RT-PCR analysis of NbCyp71D20 and NbACRE132 defense gene expression 60 min after treatment with 100 μg/ml chitin in N. benthamiana leaf tissue transiently expressing an EV control or each Group A effector as indicated. A mock induction treatment in N. benthamiana leaf tissue transiently expressing EV (Mock EV) is included as a negative control. All samples were normalized against the housekeeping gene NbEF1α and the measurements represent the ratio of expression levels (%) compared to the chitin-induced EV sample. Error bars represent (SEM; n = 3). Statistical significance compared to PAMP-induced N. benthamiana tissue expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). The results shown in (A–D) are representative of three independent experiments. All effector genes were tested but those that didn’t show any response of suppression are not represented.
Group C: Suppressors of PAMP-Induced Transcriptional Activation of Defense Genes
Recognition of microbial elicitors orchestrates an extensive defense-oriented transcriptional reprogramming of the affected cell (Navarro et al., 2004; Wan et al., 2008). In this study, we identified six effectors which downregulated flg22- and chitin-induced transcriptional activation of NbCyp71D20 and NbACRE132 genes. These included the Pto DC3000 effectors HopF2PtoDC3000, HopAF1PtoDC3000, HopI1PtoDC3000, and HopH1PtoDC3000, and the Pta 11528 effectors HopF1Pta11528 and HopI1Pta11528. These effectors reduced rather than abolished gene induction, to a range of about 40–70% of the EV controls. Each effector caused very similar reductions in the expression of each marker gene. None of them compromised the Ca2+ and ROS bursts produced after flg22 or chitin perception (Figures 5A,B). Among them, HopH1PtoDC3000 showed the strongest ability to compromised flg22- and chitin-induced expression of both NbCyp71D20 and NbACRE132 genes (Figures 5C,D), whereas the remaining effectors showed reproducibly intermediate levels of suppression. HopF1Pta11528 and HopI1Pta11528 consistently reduced flg22- and chitin-induced gene expression to a similar extent as their Pto DC3000 homologs. The fact that effector proteins from both Pto DC3000 and Pta 11528 compromised PAMP-induced gene expression shows that this function is conserved between strains. In all cases, these effectors compromised transcriptional responses to both flg22 and chitin, suggesting that these pathways converge at some point downstream of each PRR complex.
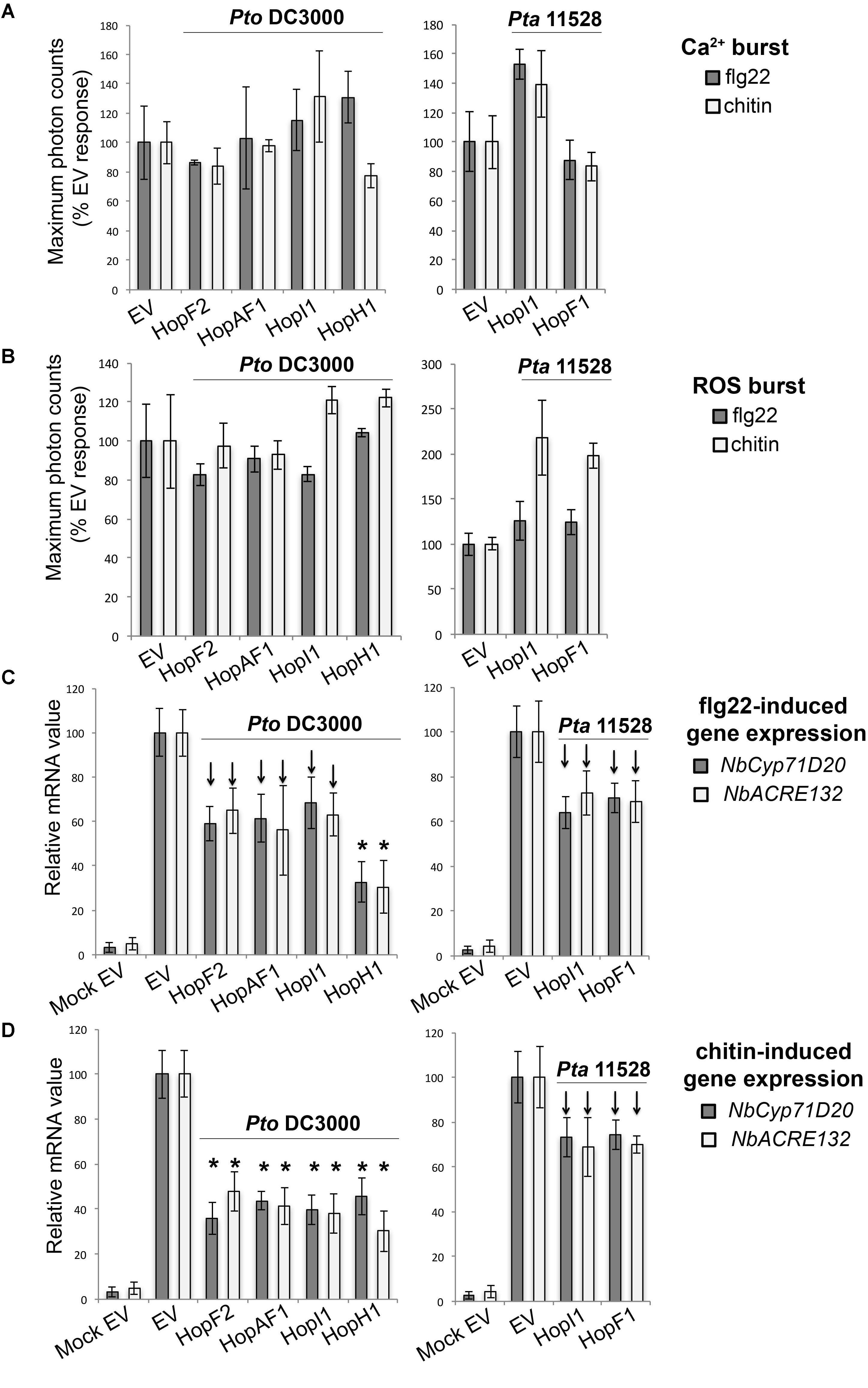
FIGURE 5. Group C effectors: suppressors of PAMP-induced transcriptional activation of defense genes. (A) Ca2+ burst in N. benthamiana SLJR15 transgenic plants transiently expressing an EV or each Group C effector as indicated after treatment with 100 nM flg22 or 100 μg/ml chitin. Data are presented relative (%) to control EV (PAMP-treated). Error bars represent the standard error of the mean (SEM; n = 8). Statistical significance compared to N. benthamiana tissue transiently expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). (B) ROS burst in N. benthamiana plants transiently expressing an EV or each Group C effector as indicated after treatment with 100 nM flg22 or 100 μg/ml chitin. Data are presented relative (%) to control EV (PAMP-treated). Error bars represent the standard error of the mean (SEM; n = 8). Statistical significance compared to N. benthamiana tissue transiently expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). (C) Quantitative RT-PCR analysis of NbCyp71D20 and NbACRE132 defense gene expression 60 min after treatment with 100 nM flg22 in N. benthamiana leaf tissue transiently expressing an EV control or each Group C effector as indicated. A mock induction treatment in N. benthamiana leaf tissue transiently expressing EV (Mock EV) is included as a negative control. All samples were normalized against the housekeeping gene NbEF1α and the measurements represent the ratio of expression levels (%) compared to the flg22-induced EV sample. Error bars represent (SEM; n = 3). Statistical significance compared to PAMP-induced N. benthamiana tissue expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). Arrows indicate consistent reduction observed in independent experiments. (D) Quantitative RT-PCR analysis of NbCyp71D20 and NbACRE132 defense gene expression 60 min after treatment with 100 μg/ml chitin in N. benthamiana leaf tissue transiently expressing an EV control or each Group C effector proteins as indicated. A mock induction treatment in N. benthamiana leaf tissue transiently expressing EV (Mock EV) is included as a negative control. All samples were normalized against the housekeeping gene NbEF1α and the measurements represent the ratio of expression levels (%) compared to the chitin-induced EV sample. Error bars represent (SEM; n = 3). Statistical significance compared to PAMP-induced N. benthamiana tissue expressing EV is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). Arrows indicate consistent reduction observed in independent experiments. The results shown in (A–D) are representative of three independent experiments. All effector genes were tested but those that didn’t show any response of suppression are not represented.
HopAD1 and HopM1 Compromise PAMP-Dependent ROS Production in the Presence of the Cell Death Blocker HopY1
Some Pto D3000 effectors both suppress PAMP responses and elicit cell death when expressed transiently in N. benthamiana leaves. In order to determine whether cell death caused the absence of PAMP-induced defense responses, we suppressed the hopAD1PtoDC3000 and hopM1PtoDC3000 induced necroses by co-expressing them with the previously described effector hopY1PtoDC3000 in N. benthamiana. This effector blocked the necroses triggered by hopAD1PtoDC3000 and hopM1PtoDC3000, but did not compromise PAMP responses in any of our assays. Both hopAD1PtoDC3000 and hopM1PtoDC3000 suppressed the ROS burst upon flg22 and chitin treatment to the same extent independent of the presence of hopY1PtoDC3000 (Figure 6), suggesting that ROS suppression upon PAMP treatment by these effectors is likely the outcome of their activities within the plant cell rather than due to the death of the infiltrated tissue.
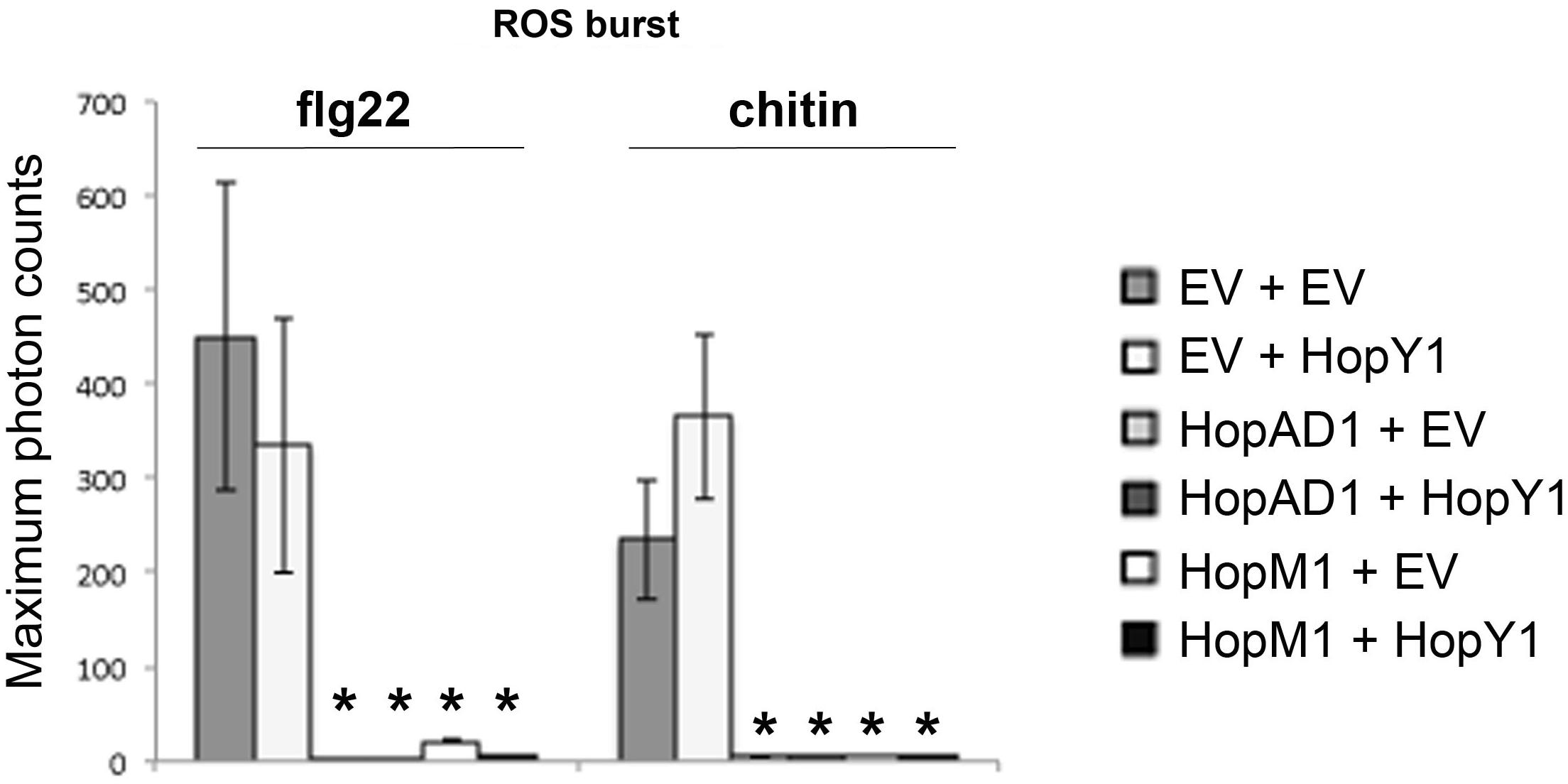
FIGURE 6. HopAD1 and HopM1 compromise PAMP-dependent ROS production in the presence of the cell death blocker HopY1. ROS burst in N. benthamiana plants transiently co-expressing an empty vector (EV), hopAD1PtoDC3000 or hopM1PtoDC3000 with the effector hopY1PtoDC3000 or the EV control as indicated after treatment with 100 nM flg22 or 100 μg/ml chitin. Error bars represent the standard error of the mean (SEM; n = 8). Statistical significance compared to N. benthamiana tissue transiently co-expressing EV (EV + EV. PAMP-treated) is indicated by asterisks (Student’s t-test, ∗p ≤ 0.01). The result shown is representative of two independent experiments.
Discussion
Bacterial pathogens use a TTSS to deliver effector proteins into eukaryotic cells. This mechanism enables the bacterium to grow to high levels and produce disease symptoms (Collmer et al., 2002). We found that 20 of the 22 Pto DC3000 effectors and 9 of 10 Pta 11528 effectors tested in this study showed suppressive activity in at least one of defense assay when overexpressed transiently using Agrobacterium-mediated transformation. This is consistent with the notion that a major role of effector proteins is to suppress plant innate immunity, and that they do so at different times and points in the signaling pathways. A comprehensive picture of the activity of effector molecules inside plant cells is summarized in Figures 7, 8.
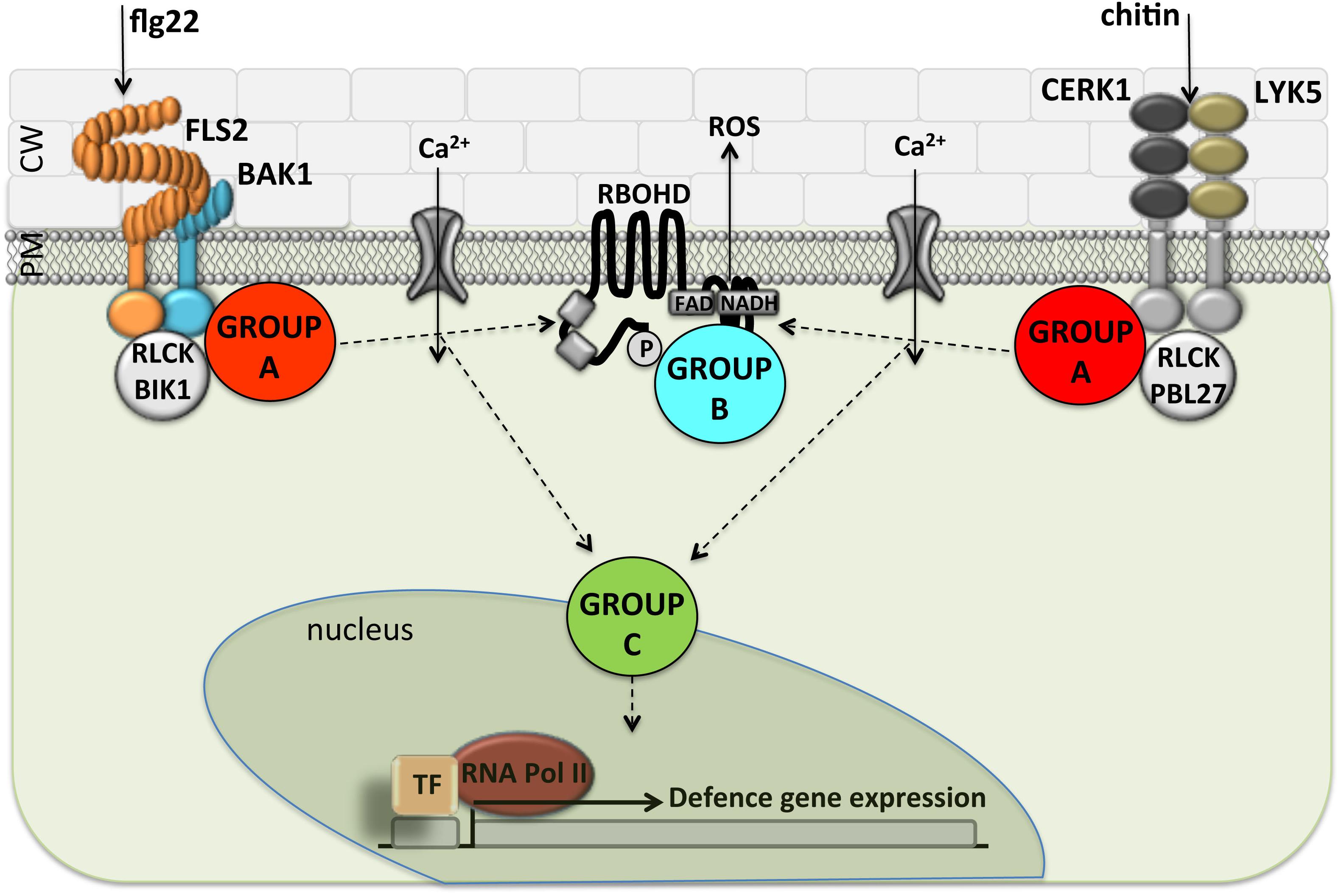
FIGURE 7. Separation of P. syringae type-III effectors into functional groups based on the ability to suppress different PAMP responses. The scheme represents the FLS2 and LYK5 PRR complexes at the plasma membrane and shows the major complex components. The cellular events measured here downstream of PAMP perception; Ca2+ influx, ROS generation and changes in defense gene activation, are depicted according to the model in Segonzac et al. (2011). As an exception, the Group A effectors AvrPtoPtoDC3000 and AvrPtoPta11528 suppressed flg22-induced outputs but had no effect on chitin-induced defense responses. Arrows are indicative and drawn for conceptual simplicity to show the order of events, but junctions may exist differently from depicted. PM, plasma membrane; CW, cell wall; ROS, reactive oxygen compound synthesis; TF, transcription factor.

FIGURE 8. Summary of results. Summary of the ability of all Pto DC3000 and Pta 11528 effectors tested in this manuscript to interfere with effector-induced cell death in N. benthamiana and the activation of PAMP-dependent defense responses including the Ca2+ influx, generation of ROS and defense gene expression. Dark gray indicates complete suppression. Light gray indicates significant reduction compared to positive control. White indicates no suppression. ND, not done. The cell death suppression box indicates those effectors for which cell death was compromised, namely hopAD1PtoDC3000 or hopM1PtoDC3000 for Pto DC3000, and hopT1-1Pta11528 for Pta 11528.
Evolutionary Aspects of Type III Effectors Conserved in Both Pto DC3000 and Pta 11528
Comparing the activities of the effector repertoires of Pto DC3000 and Pta 11528 is a useful first step toward an understanding how these bacteria cause disease on different host species. We generated a library of 10 Pta 11528 effector genes, which constitute about half of the effectors present in the genome (Studholme et al., 2009). Seven of these effectors have homologs in the Pto DC3000 effector library analyzed in this work. However, the level of conservation varies among them. Some effectors show high levels of identity, including HopO1-1Pta11528 and HopT1-1Pta11528, which are 99 and 98% identical at the amino acid level with their respective Pto DC3000 homologs (Supplementary Figures S1, S2). Putative catalytic functions identified by prediction programs are present in both the Pta 11528 and Pto DC3000 effector homologs. Therefore, it seems likely that they are conserved functionally. Despite this, the transgenic expression phenotypes of the Pta 11528 and Pto DC3000 HopT1-1 homologs differed significantly. While HopT1-1Pta11528 elicited strong necrosis in N. benthamiana, HopT1-1PtoDC3000 did not. Although the effectors share high identity at the amino acid level, they differ in five residues that likely confer the basis for this behavior. Likewise, the cysteine protease HopX1Pta11528 is highly similar to its Pto DC3000 homolog and catalytic residues required for putative enzymatic activity are conserved (Supplementary Figure S4). However, only the Pta 11528 version of HopX1 interfered with the chitin-induced ROS burst. It was previously reported that HopX1Pta11528 but not HopX1PtoDC3000 targets conserved JAZ transcriptional repressors to activate jasmonate (JA) hormone signaling (Gimenez-Ibanez et al., 2014). In contrast to Pto DC3000, the Pta 11528 strain does not produce coronatine (COR), a phytotoxin that mimics the active form of JA, and therefore, exploits an alternative evolutionary strategy to activate the pathway through the HopX1Pta11528 effector. HopX1 and COR biosynthetic genes generally do not co-exist in a single strain, but in the few cases where both occur, the HopX1 alleles contain mutations in functionally essential residues (Yang et al., 2017), suggesting that redundancy between COR and HopX1 might have inactivated and/or allowed the effector to evolve toward other functions. In contrast, the Pto DC3000 E3-ligase effector AvrPtoB also shares high level of amino acid identity (70%) with AvrPtoBPta11528 (Supplementary Figure S3) and the amino acids critical for E3 ligase activity (F479A, T450, F525, and P533) are present in AvrPtoBPta11528. This suggests that AvrPtoBPta11528 is also an active E3-ligase in plant cells and congruently, both Pto DC3000 and Pta 11528 homologs showed similar abilities to interfere with PAMP-triggered immunity (Janjusevic et al., 2006; Ntoukakis et al., 2009). Interestingly, although AvrPtoPta11528, HopI1Pta11528, and HopF1Pta11528 show significantly less homology with their respective Pto DC3000 homologs, they showed functional conservation in PAMP-induced defense suppression assays (Supplementary Figures S5–S7). The putative homolog AvrPtoPta11528 shares just 42% sequence identity at the amino acid level. The N- and C-terminal regions are highly conserved but there is significant diversity in the central region required for both the virulence and avirulence functions of this effector in tomato (Chang et al., 2001; Wulf et al., 2004; Xing et al., 2007). Similarly, HopI1Pta11528 shares 55% identity with HopI1PtoDC3000 (Supplementary Figure S6), and both the N- and C-terminal regions are highly conserved maintaining a chloroplast targeting signal and the J domain required for virulence (Jelenska et al., 2007). However, 152 central amino acids containing the proline- and glutamine- (P/Q)-rich repeat region of unknown function are completely absent in HopI1Pta11528. Despite these differences, both the AvrPto and HopI1 homologs in Pto DC3000 and Pta 11528 strains showed similar abilities to suppress PAMP-induced defense responses, and thus the non-conserved regions may not be required for virulence functions in N. benthamiana. The differences between the Pta 11528 effectors and the Pto DC3000 homologs might indicate functional diversification as a consequence of divergent evolution, as well as diversification to escape recognition.
Cell Death Promotion and Suppression by Pto DC3000 and Pta 11528 Effectors
Several Pto DC3000 and Pta 11528 effectors elicited cell death in N. benthamiana. Five effectors including hopM1PtoDC3000, hopAD1PtoDC3000, hopT1-1Pta11528, avrE1Pta11528, and hopW1-1Pta11528 strongly elicited necrosis whereas two effectors, hopAA1-1PtoDC3000 and hopQ1-1PtoDC3000 displayed mild chlorosis when expressed transiently in N. benthamiana leaves. This supports previous observations indicating that multiple P. syringae effectors trigger cell death in N. benthamiana (Vinatzer et al., 2006; Wei et al., 2007; Wroblewski et al., 2009; Choi et al., 2017). For example, hopM1PtoDC3000, hopAA1-1PtoDC3000, hopQ1-1PtoDC3000, avrE1PtoDC3000, hopT1-1PtoDC3000, and hopAD1PtoDC3000 effectors elicit cell death in N. benthamiana when delivered by Pseudomonas fluorescens heterologously expressing a P. syringae TTSS (Wei et al., 2007) or though Pto DC3000 secretion effector polymutants (Wei et al., 2007, 2015). Similar to our experiments, transient expression of a number of effectors from Pto DC3000, P. syringae pv. actinidiae (Psa), and P. syringae pv. syringae (Psy) B728a, showed the ability of hopQ1-1, avrE1, hopT1-1, hopM1, and hopAA1-1 to induce cell death in N. benthamiana (Vinatzer et al., 2006; Wroblewski et al., 2009; Choi et al., 2017). It is noteworthy that in our assays, hopT1-1PtoDC3000 did not trigger cell death in N. benthamiana, although this has been previously reported (Wei et al., 2007; Wroblewski et al., 2009).
Cell death is also associated with effector recognition by R proteins. This has been reported for HopQ1-1PtoDC3000, and to a much weaker extent for HopAD1PtoDC3000 and the Pma ES4326 effector HopW1-1 in N. benthamiana (Wei et al., 2007, 2015; Lee et al., 2008). HopQ1-1PtoDC3000 and HopAD1PtoDC3000 are recognized in N. benthamiana and consequently, Pto DC3000 lacking the hopQ1-1 or hopAD1 genes is capable of causing disease symptoms comparable to the virulent Pta 11528 (Wei et al., 2007, 2015). The R protein Roq1 mediates recognition of Xanthomonas and Pseudomonas effector proteins XopQ and HopQ1 in the Nicotiana genus (Schultink et al., 2017), whereas in Arabidopsis, a genetic interaction between Qpm3.1 and hopW1-1 determines resistance to bacterial infection (Luo et al., 2017). However, several lines of evidence suggest that host R proteins do not recognize all effectors that elicited cell death here. Firstly, necrosis induced by hopM1PtoDC3000, hopAD1PtoDC3000, hopT1-1Pta11528, avrE1Pta11528, and hopW1-1Pta11528 effector genes was not dependent on NbSgt1, which is typically required for R gene function (McDowell and Dangl, 2000). The NbSgt1-independence of avrE1 induced cell death has been reported previously (Choi et al., 2017). However, cell death induced by the Psy B728a effector hopM1 and the Psa effectors hopT1-1 and hopW1-1 were reported to be at least partially dependent on NbSgt1 (Vinatzer et al., 2006; Choi et al., 2017). Therefore, further analyses are required to determine to what extent the cell death caused by these effectors is dependent on NbSgt1 in N. benthamiana. On the other hand, NbSgt1 may not be required for all R gene-mediated plant defenses and thus, correlation of NbSgt1-dependency with effector recognition by R proteins should be undertaken cautiously. Secondly, Pta 11528 is virulent on N. benthamiana despite possessing a HopW1-1 homolog. It is possible that some of these effectors would not elicit cell death when expressed under native conditions, i.e., in bacteria and delivered to the plant cell via the TTSS. And thirdly, there is precedence for host cell death to be associated with effector virulence, as demonstrated for members of the AvrE1 and HopM1 effector families (Badel et al., 2006; Boureau et al., 2006; Ham et al., 2006). Both avrE1 and hopM1 are functionally redundant and reside in the conserved effector locus (CEL) of the P. syringae Hrp pathogenicity island (DebRoy et al., 2004; Badel et al., 2006; Kvitko et al., 2009). HopM1 and AvrE1 are required to establish a water-soaked aqueous living space in plants that is crucial for virulence (Xin et al., 2016). Whether HopAA1-1, another effector encoded in the CEL locus, or the remaining cell death-inducing effectors of Pto DC3000 and Pta 11528 function similarly to HopM1/AvrE in altering apoplastic environmental conditions is unknown. The conservation of necrosis-inducing effectors in various bacteria suggests this function may be an important virulence strategy for bacterial infections on plants.
A number of Pto DC3000 and Pta 11528 effectors masked the activity of cell death-inducing effectors when co-expressed transiently in N. benthamiana. Similar data exist for suppression of effector-triggered cell death phenotypes by Pto DC3000 and Psa effectors (Jamir et al., 2004; Guo et al., 2009; Wei et al., 2015; Choi et al., 2017). Indeed, most Pto DC3000 effectors that acted as cell death suppressors in this work where previously identified as suppressors of the HR response induced by HopA1 in tobacco when delivered by Pseudomonas fluorescens heterologously expressing a P. syringae TTSS (Guo et al., 2009). Notably, it has been reported recently that HopQ1-mediated cell death suppression in N. benthamiana is due to attenuation of Agrobacterium-mediated protein expression rather than a genuine virulence activity. Thus, it is possible that additional effectors may act in a similar fashion (Adlung and Bonas, 2017). We speculate that necrosis induction by effectors must be tightly regulated to establish and maintain an optimal intracellular niche. Although it is possible that some effectors would not interfere with cell death when expressed at native levels, the potent activity of some of them suggests that cell death suppression may occur during infection to at least some extent. On the other hand, the ability of some effectors to interfere with the outcomes of others identifies which effectors perturbate common defensive pathways. HopAD1PtoDC3000-, HopM1PtoDC3000-, and HopT1-1Pta11528-dependent cell death could be efficiently suppressed by multiple effectors from both Pto DC3000 and Pta 11528 repertoires, which include both AvrPtoB homologs. This supports recent work by Wei et al. (2015) indicating interplay between HopAD1 and AvrPtoB in regulating HopAD1-dependent cell death. Despite this, Pto DC3000 effectors could not collectively suppress the HR events activated upon recognition of AvrPtoB and/or AvrPto in transgenic N. benthamiana plants expressing the tomato Pto and Prf resistance genes (Balmuth and Rathjen, 2007). In addition, neither the effector complements of Pto DC3000 or Pta 11528 were able to collectively suppress HopQ1-1 or HopAD1 induced defenses in N. benthamiana (Wei et al., 2007, 2015). Interplay between effectors within the bacterial repertoires is emerging as an important but poorly understood phenomenon controlling defensive phenotypic outcomes (Wei et al., 2015; Huett, 2017). More studies are needed in this area to understand the complex network of interactions between the effectors among the repertoires and how this translates into an effective immune response or a virulence strategy to promote disease.
Pseudomonas syringae Effectors Differentially Suppress Innate Immune Responses
In recent years, many effectors have been shown to suppress PAMP-induced defenses (Macho and Zipfel, 2015; Buttner, 2016; Toruno et al., 2016). Here, we analyzed the ability of effectors to interfere with PAMP-triggered defenses activated between seconds to minutes to hours after elicitation, which has allowed us to group effectors into three major categories based on their range of suppressive activities.
Group A effectors acted as broad-range suppressors of all early PAMP-induced responses tested. AvrPtoPtoDC3000 and AvrPtoBPtoDC3000 which fall into this group, are well known suppressors of flg22-induced defenses by targeting PRR complexes directly at the plasma membrane (Chinchilla et al., 2007; Heese et al., 2007; Gohre et al., 2008; Xiang et al., 2008). The additional Pto DC3000 and Pta 11528 effectors within Group A may also target PRR complexes due to their comprehensive suppression activities. Indeed, direct inactivation or destabilization of PAMP receptor complexes at the plasma membrane is a common strategy of multiple effectors (Macho and Zipfel, 2015; Buttner, 2016; Toruno et al., 2016). HopM1PtoDC3000 suppresses vesicle trafficking to overcome host innate immunity (Nomura et al., 2006). Our data indicates additional roles for this effector in early PAMP signaling suppression. Both HopM1PtoDC3000 and HopQ1-1PtoDC3000 associate with multiple 14-3-3 proteins (Nomura et al., 2006; Giska et al., 2013; Li et al., 2013; Lozano-Duran et al., 2014). 14-3-3 proteins are highly conserved eukaryotic regulatory adapters whose interaction with client proteins can regulate protein activity and remarkably, chemical inhibition of 14-3-3s results in suppression of the PAMP-triggered ROS burst (Lozano-Duran et al., 2014). Some 14-3-3 proteins associate with several defense-related proteins in planta, including the FLS2 co-receptor BAK1 (Chang et al., 2009). Whether HopM1PtoDC3000 and HopQ1-1PtoDC3000 act through interaction with 14-3-3s remains to be explored.
Group B effectors suppressed the PAMP-induced ROS burst not the Ca2+ influx. These included HopT1-1PtoDC3000, HopX1Pta11528, and HopAR1Pta11528. These effectors abolished the PAMP-induced ROS burst with differential impact on activation of defense gene markers. While HopT1-1PtoDC3000 compromised flg22- and chitin-dependent induction of defense genes, HopX1Pta11528 and HopAR1Pta11528 did not. Furthermore, their ROS-suppressive action was restricted to chitin-induced responses. The P. syringae pv. phaseolicola effector HopAR1 suppresses PTI by directly targeting BIK1 and other RLCKs (Zhang et al., 2010), which contrast with our results. However, the hopAR1Pta11528 and hopAR1Pphrace3 versions of this effector differ significantly in their protein sequences. HopAR1Pta11528 contains an extra 44 N-terminal amino acids (in a relatively small effector of 311 amino acids), whereas the common C-terminal domain shares a moderate 78% identity, which may account for the observed differences. In an alternative model, HopX1Pta11528 targets conserved JAZ transcriptional repressors to activate JA signaling and promote infection in Arabidopsis (Gimenez-Ibanez et al., 2014), and therefore, additional targets would be expected for this effector. This is unsurprising as increasing evidence suggests that effector proteins target multiple host proteins simultaneously in a “death by a thousand cuts” strategy to abolish defense responses (Navarro et al., 2008; Shan et al., 2008; Gimenez-Ibanez et al., 2009).
Group C effectors are suppressors of PAMP-induced gene induction. This group only compromised PAMP-induced gene expression but not the other outputs. They include the Pto DC3000 effectors HopF2PtoDC3000, HopAF1PtoDC3000, HopI1PtoDC3000, and HopH1PtoDC3000, and the Pta 11528 effectors HopF1Pta11528 and HopI1Pta11528. Because this group suppressed gene induction but not plasma membrane-related events, we suggest that they act later in signal transduction perhaps proximal to or within the nucleus. In this group falls HopF2PtoDC3000, which was previously shown to suppress Arabidopsis immunity by targeting the co-receptor BAK1 (Zhou et al., 2014). This finding is difficult to reconcile with our data as this effector did not suppress early flg22-induced Ca2+ and ROS bursts in our experiments to any extent. This is similar to HopB1PtoDC3000, one of the few effectors that did not suppress any of the defensive outputs tested in this work, despite the fact that it acts as a protease that targets BAK1 (Li et al., 2016). On the other hand, HopAF1PtoDC3000 suppresses ethylene production (Washington et al., 2016) whereas HopI1 targets plant heat shock chaperone protein Hsp70 (Jelenska et al., 2010). None of these functions explains the ability of these effectors to suppress PAMP-induced gene expression. Thus, we expect alternative targets for these effectors to disrupt transcriptional activation. Effectors that interfere with the transcriptional machinery are not yet known from Pto DC3000, but are well described in other pathogens such as Xanthomonas which produces transcription activator-like effectors (TALEs) (Boch et al., 2009; Moscou and Bogdanove, 2009). Whether Pto DC3000 also injects direct modulators of host gene expression remains an open question for the future.
Most Effectors Interfere With Responses Elicited by Different PAMPs
Most effectors interfered with both flg22- and chitin-induced defense responses simultaneously, indicating that a considerable overlap exists between these pathways. One exception was the AvrPto homologs of both Pto DC3000 and Pta 11528 which suppressed flagellin signaling but not chitin responses. This was surprising because AvrPtoPtoDC3000 was previously reported to block chitin-induced defense gene expression in transgenic Arabidopsis protoplasts (Shan et al., 2008). It is possible that the chitin-suppressing effect of AvrPtoPtoDC3000 was simply due to overexpression in Arabidopsis. Alternatively, the kinase domain of the Arabidopsis chitin receptor/receptor complex might be simply a better target for AvrPtoPtoDC3000 than its N. benthamiana homolog (Shan et al., 2008; Xiang et al., 2008). Therefore, it remains to be determined how chitin-induced signaling in N. benthamiana evades AvrPto-mediated suppression. Conversely, HopX1Pta11528 and HopAR1Pta11528 suppressed chitin-induced ROS generation, but not that generated by flg22 treatment. This might be simply a consequence of the different strength of ROS levels induced by both elicitors. While flg22 induces a very strong response, the chitin response is relatively mild. Therefore, smaller effects might be more easily revealed using chitin as an elicitor. Overall, much of our data suggest that chitin- and flg22-induced signaling converge soon after PAMP perception, since most effectors suppressed defense responses induced by both elicitors.
Final Considerations
Nicotiana benthamiana has emerged as an important and widely used experimental system to study plant biology. Its advantages include its large leaves, its high amenability for A. tumefaciens-mediated transient expression, the possibility to rapidly test the involvement of host factors using VIGS, and the reproducibility of the data (77, 78). Agrobacterium-mediated transient expression has already been used to study some effectors that suppress plant defense responses at the molecular level (24, 79). Thus, the efficiency and versatility of the agroinfiltration technique in N. benthamiana prompted us to investigate the ability of a large number of P. syringae type III effectors to suppress a collection of defense responses activated upon PAMP treatment. However, it should be taken into account that overexpression of effector proteins can led to misleading results by causing protein imbalances, promiscuous interactions, and regulation of pathways that are associated with the degree of overexpression rather than the function of the protein. On the other hand, overexpression of effector proteins in plant cells, combined with loss-of-function analyses, is a common and powerful approach to understanding effector function. Identification of effector targets is the next step to unraveling how pathogens can overcome plant immunity and promote pathogenesis. This will lead ultimately to new strategies for crop protection in the field.
Author Contributions
DH, JR, and SG-I designed the experiments. DH, CS, and SG-I conducted the experiments. DH, JR, and SG-I analyzed the data. JR, JC, TB, and SG-I wrote the manuscript. All authors edited and approved the final manuscript.
Funding
This work was funded by the Gatsby Charitable Foundation and the Spanish Ministry of Science and Innovation Grant BIO2014-55884-JIN to SG-I.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer BK declared a past co-authorship with one of the authors JC to the handling Editor.
Acknowledgments
We thank Professor Dr. Jeffery L. Dangl for providing full-length Pto DC3000 effectors recombined into pDONR207 (Chang et al., 2005) and Dr. Gregory Martin for the Pto DC3000ΔavrPtoBΔavrPto strain. In addition we thank Dr. Birgit Schulze for careful revision of the manuscript and Anna Egger for technical support. We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.00688/full#supplementary-material
References
Adlung, N., and Bonas, U. (2017). Dissecting virulence function from recognition: cell death suppression in Nicotiana benthamiana by XopQ/HopQ1-family effectors relies on EDS1-dependent immunity. Plant J. 91, 430–442. doi: 10.1111/tpj.13578
Alfano, J. R., and Collmer, A. (2004). Type III secretion system effector proteins: double agents in bacterial disease and plant defense. Annu. Rev. Phytopathol. 42, 385–414. doi: 10.1146/annurev.phyto.42.040103.110731
Badel, J. L., Shimizu, R., Oh, H. S., and Collmer, A. (2006). A Pseudomonas syringae pv. tomato avrE1/hopM1 mutant is severely reduced in growth and lesion formation in tomato. Mol. Plant Microbe Interact. 19, 99–111. doi: 10.1094/MPMI-19-0099
Balmuth, A., and Rathjen, J. P. (2007). Genetic and molecular requirements for function of the Pto/Prf effector recognition complex in tomato and Nicotiana benthamiana. Plant J. 51, 978–990. doi: 10.1111/j.1365-313X.2007.03199.x
Blume, B., Nurnberger, T., Nass, N., and Scheel, D. (2000). Receptor-mediated increase in cytoplasmic free calcium required for activation of pathogen defense in parsley. Plant Cell 12, 1425–1440. doi: 10.1105/tpc.12.8.1425
Boch, J., Scholze, H., Schornack, S., Landgraf, A., Hahn, S., Kay, S., et al. (2009). Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509–1512. doi: 10.1126/science.1178811
Boller, T., and Felix, G. (2009). A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 60, 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
Boureau, T., Elmaarouf-Bouteau, H., Garnier, A., Brisset, M. N., Perino, C., Pucheu, I., et al. (2006). DspA/E, a type III effector essential for Erwinia amylovora pathogenicity and growth in planta, induces cell death in host apple and nonhost tobacco plants. Mol. Plant Microbe Interact. 19, 16–24. doi: 10.1094/MPMI-19-0016
Boutrot, F., Segonzac, C., Chang, K. N., Qiao, H., Ecker, J. R., Zipfel, C., et al. (2010). Direct transcriptional control of the Arabidopsis immune receptor FLS2 by the ethylene-dependent transcription factors EIN3 and EIL1. Proc. Natl. Acad. Sci. U.S.A. 107, 14502–14507. doi: 10.1073/pnas.1003347107
Boutrot, F., and Zipfel, C. (2017). Function, discovery, and exploitation of plant pattern recognition receptors for broad-spectrum disease resistance. Annu. Rev. Phytopathol. 55, 257–286. doi: 10.1146/annurev-phyto-080614-120106
Buttner, D. (2016). Behind the lines-actions of bacterial type III effector proteins in plant cells. FEMS Microbiol. Rev. 40, 894–937. doi: 10.1093/femsre/fuw026
Cao, Y., Liang, Y., Tanaka, K., Nguyen, C. T., Jedrzejczak, R. P., Joachimiak, A., et al. (2014). The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. eLife 3:e03766. doi: 10.7554/eLife.03766
Chandra, S., Stennis, M., and Low, P. S. (1997). Measurement of Ca2+ fluxes during elicitation of the oxidative burst in aequorin-transformed tobacco cells. J. Biol. Chem. 272, 28274–28280. doi: 10.1074/jbc.272.45.28274
Chang, I. F., Curran, A., Woolsey, R., Quilici, D., Cushman, J. C., Mittler, R., et al. (2009). Proteomic profiling of tandem affinity purified 14-3-3 protein complexes in Arabidopsis thaliana. Proteomics 9, 2967–2985. doi: 10.1002/pmic.200800445
Chang, J. H., Tobias, C. M., Staskawicz, B. J., and Michelmore, R. W. (2001). Functional studies of the bacterial avirulence protein AvrPto by mutational analysis. Mol. Plant Microbe Interact. 14, 451–459. doi: 10.1094/MPMI.2001.14.4.451
Chang, J. H., Urbach, J. M., Law, T. F., Arnold, L. W., Hu, A., Gombar, S., et al. (2005). A high-throughput, near-saturating screen for type III effector genes from Pseudomonas syringae. Proc. Natl. Acad. Sci. U.S.A. 102, 2549–2554. doi: 10.1073/pnas.0409660102
Chinchilla, D., Bauer, Z., Regenass, M., Boller, T., and Felix, G. (2006). The Arabidopsis receptor kinase FLS2 Binds flg22 and determines the specificity of flagellin perception. Plant Cell 18, 465–476. doi: 10.1105/tpc.105.036574
Chinchilla, D., Zipfel, C., Robatzek, S., Kemmerling, B., Nurnberger, T., Jones, J. D., et al. (2007). A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 448, 497–500. doi: 10.1038/nature05999
Chisholm, S. T., Coaker, G., Day, B., and Staskawicz, B. J. (2006). Host-microbe interactions: shaping the evolution of the plant immune response. Cell 124, 803–814. doi: 10.1016/j.cell.2006.02.008
Choi, S., Jayaraman, J., Segonzac, C., Park, H. J., Park, H., Han, S. W., et al. (2017). Pseudomonas syringae pv. actinidiae type III effectors localized at multiple cellular compartments activate or suppress innate immune responses in Nicotiana benthamiana. Front. Plant Sci. 8:2157. doi: 10.3389/fpls.2017.02157
Collmer, A., Lindeberg, M., Petnicki-Ocwieja, T., Schneider, D. J., and Alfano, J. R. (2002). Genomic mining type III secretion system effectors in Pseudomonas syringae yields new picks for all TTSS prospectors. Trends Microbiol. 10, 462–469. doi: 10.1016/S0966-842X(02)02451-4
Davies, D. R., Bindschedler, L. V., Strickland, T. S., and Bolwell, G. P. (2006). Production of reactive oxygen species in Arabidopsis thaliana cell suspension cultures in response to an elicitor from Fusarium oxysporum: implications for basal resistance. J. Exp. Bot. 57, 1817–1827. doi: 10.1093/jxb/erj216
DebRoy, S., Thilmony, R., Kwack, Y. B., Nomura, K., and He, S. Y. (2004). A family of conserved bacterial effectors inhibits salicylic acid-mediated basal immunity and promotes disease necrosis in plants. Proc. Natl. Acad. Sci. U.S.A. 101, 9927–9932. doi: 10.1073/pnas.0401601101
Ferreira, A. O., Myers, C. R., Gordon, J. S., Martin, G. B., Vencato, M., Collmer, A., et al. (2006). Whole-genome expression profiling defines the HrpL regulon of Pseudomonas syringae pv. tomato DC3000, allows de novo reconstruction of the Hrp cis clement, and identifies novel coregulated genes. Mol. Plant Microbe Interact. 19, 1167–1179. doi: 10.1094/MPMI-19-1167
Gimenez-Ibanez, S., Boter, M., Fernandez-Barbero, G., Chini, A., Rathjen, J. P., and Solano, R. (2014). The bacterial effector HopX1 targets JAZ transcriptional repressors to activate jasmonate signaling and promote infection in Arabidopsis. PLoS Biol. 12:e1001792. doi: 10.1371/journal.pbio.1001792
Gimenez-Ibanez, S., Hann, D. R., Ntoukakis, V., Petutschnig, E., Lipka, V., and Rathjen, J. P. (2009). AvrPtoB targets the LysM receptor kinase CERK1 to promote bacterial virulence on plants. Curr. Biol. 19, 423–429. doi: 10.1016/j.cub.2009.01.054
Giska, F., Lichocka, M., Piechocki, M., Dadlez, M., Schmelzer, E., Hennig, J., et al. (2013). Phosphorylation of HopQ1, a type III effector from Pseudomonas syringae, creates a binding site for host 14-3-3 proteins. Plant Physiol. 161, 2049–2061. doi: 10.1104/pp.112.209023
Gohre, V., Spallek, T., Haweker, H., Mersmann, S., Mentzel, T., Boller, T., et al. (2008). Plant pattern-recognition receptor FLS2 is directed for degradation by the bacterial ubiquitin ligase AvrPtoB. Curr. Biol. 18, 1824–1832. doi: 10.1016/j.cub.2008.10.063
Goodin, M. M., Zaitlin, D., Naidu, R. A., and Lommel, S. A. (2008). Nicotiana benthamiana: its history and future as a model for plant-pathogen interactions. Mol. Plant Microbe Interact. 21, 1015–1026. doi: 10.1094/MPMI-21-8-1015
Guo, M., Tian, F., Wamboldt, Y., and Alfano, J. R. (2009). The majority of the type III effector inventory of Pseudomonas syringae pv. tomato DC3000 can suppress plant immunity. Mol. Plant Microbe Interact. 22, 1069–1080. doi: 10.1094/MPMI-22-9-1069
Hahlbrock, K., Bednarek, P., Ciolkowski, I., Hamberger, B., Heise, A., Liedgens, H., et al. (2003). Non-self recognition, transcriptional reprogramming, and secondary metabolite accumulation during plant/pathogen interactions. Proc. Natl. Acad. Sci. U.S.A. 100(Suppl. 2), 14569–14576. doi: 10.1073/pnas.0831246100
Ham, J. H., Majerczak, D. R., Arroyo-Rodriguez, A. S., Mackey, D. M., and Coplin, D. L. (2006). WtsE, an AvrE-family effector protein from Pantoea stewartii subsp. stewartii, causes disease-associated cell death in corn and requires a chaperone protein for stability. Mol. Plant Microbe Interact. 19, 1092–1102. doi: 10.1094/MPMI-19-1092
Hann, D. R., and Rathjen, J. P. (2007). Early events in the pathogenicity of Pseudomonas syringae on Nicotiana benthamiana. Plant J. 49, 607–618. doi: 10.1111/j.1365-313X.2006.02981.x
Heese, A., Hann, D. R., Gimenez-Ibanez, S., Jones, A. M., He, K., Li, J., et al. (2007). The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Proc. Natl. Acad. Sci. U.S.A. 104, 12217–12222. doi: 10.1073/pnas.0705306104
Huett, A. (2017). Combinatorial actions of bacterial effectors revealed by exploiting genetic tools in yeast. Mol. Syst. Biol. 13:911. doi: 10.15252/msb.20167447
Jackson, R. W., Athanassopoulos, E., Tsiamis, G., Mansfield, J. W., Sesma, A., Arnold, D. L., et al. (1999). Identification of a pathogenicity island, which contains genes for virulence and avirulence, on a large native plasmid in the bean pathogen Pseudomonas syringae pathovar phaseolicola. Proc. Natl. Acad. Sci. U.S.A. 96, 10875–10880. doi: 10.1073/pnas.96.19.10875
Jamir, Y., Guo, M., Oh, H. S., Petnicki-Ocwieja, T., Chen, S., Tang, X., et al. (2004). Identification of Pseudomonas syringae type III effectors that can suppress programmed cell death in plants and yeast. Plant J. 37, 554–565. doi: 10.1046/j.1365-313X.2003.01982.x
Janjusevic, R., Abramovitch, R. B., Martin, G. B., and Stebbins, C. E. (2006). A bacterial inhibitor of host programmed cell death defenses is an E3 ubiquitin ligase. Science 311, 222–226. doi: 10.1126/science.1120131
Jelenska, J., Van Hal, J. A., and Greenberg, J. T. (2010). Pseudomonas syringae hijacks plant stress chaperone machinery for virulence. Proc. Natl. Acad. Sci. U.S.A. 107, 13177–13182. doi: 10.1073/pnas.0910943107
Jelenska, J., Yao, N., Vinatzer, B. A., Wright, C. M., Brodsky, J. L., and Greenberg, J. T. (2007). A J domain virulence effector of Pseudomonas syringae remodels host chloroplasts and suppresses defenses. Curr. Biol. 17, 499–508. doi: 10.1016/j.cub.2007.02.028
Jones, J. D., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Kadota, Y., Sklenar, J., Derbyshire, P., Stransfeld, L., Asai, S., Ntoukakis, V., et al. (2014). Direct regulation of the NADPH oxidase RBOHD by the PRR-associated kinase BIK1 during plant immunity. Mol. Cell 54, 43–55. doi: 10.1016/j.molcel.2014.02.021
Katagiri, F., Thilmony, R., and He, S.Y. (2002). “The Arabidopsis thaliana–Pseudomonas syringae interaction,” in The Arabidopsis Book, eds C. R. Somerville, and E. M. Meyerowitz (Rockville, MD: American Society of Plant Biologists).
Keppler, L. D., Atkinson, M. M., and Baker, C. J. (1989). Active oxygen production during a bacteria-induced hypersensitive reaction in tobacco suspension cells. Phytopathology 79, 974–978. doi: 10.1094/Phyto-79-974
Kim, Y. J., Lin, N. C., and Martin, G. B. (2002). Two distinct Pseudomonas effector proteins interact with the Pto kinase and activate plant immunity. Cell 109, 589–598. doi: 10.1016/S0092-8674(02)00743-2
Kunze, G., Zipfel, C., Robatzek, S., Niehaus, K., Boller, T., and Felix, G. (2004). The N terminus of bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants. Plant Cell 16, 3496–3507. doi: 10.1105/tpc.104.026765
Kvitko, B. H., Park, D. H., Velasquez, A. C., Wei, C. F., Russell, A. B., Martin, G. B., et al. (2009). Deletions in the repertoire of Pseudomonas syringae pv. tomato DC3000 type III secretion effector genes reveal functional overlap among effectors. PLoS Pathog. 5:e1000388. doi: 10.1371/journal.ppat.1000388
Lecourieux, D., Mazars, C., Pauly, N., Ranjeva, R., and Pugin, A. (2002). Analysis and effects of cytosolic free calcium increases in response to elicitors in Nicotiana plumbaginifolia cells. Plant Cell 14, 2627–2641. doi: 10.1105/tpc.005579
Lee, M. W., Jelenska, J., and Greenberg, J. T. (2008). Arabidopsis proteins important for modulating defense responses to Pseudomonas syringae that secrete HopW1-1. Plant J. 54, 452–465. doi: 10.1111/j.1365-313X.2008.03439.x
Li, L., Kim, P., Yu, L., Cai, G., Chen, S., Alfano, J. R., et al. (2016). Activation-dependent destruction of a co-receptor by a Pseudomonas syringae effector dampens plant immunity. Cell Host Microbe 20, 504–514. doi: 10.1016/j.chom.2016.09.007
Li, L., Li, M., Yu, L., Zhou, Z., Liang, X., Liu, Z., et al. (2014). The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase RbohD to control plant immunity. Cell Host Microbe 15, 329–338. doi: 10.1016/j.chom.2014.02.009
Li, W., Yadeta, K. A., Elmore, J. M., and Coaker, G. (2013). The Pseudomonas syringae effector HopQ1 promotes bacterial virulence and interacts with tomato 14-3-3 proteins in a phosphorylation-dependent manner. Plant Physiol. 161, 2062–2074. doi: 10.1104/pp.112.211748
Lin, N. C., and Martin, G. B. (2005). An avrPto/avrPtoB mutant of Pseudomonas syringae pv. tomato DC3000 does not elicit Pto-mediated resistance and is less virulent on tomato. Mol. Plant Microbe Interact. 18, 43–51. doi: 10.1094/MPMI-18-0043
Lindeberg, M., Cartinhour, S., Myers, C. R., Schechter, L. M., Schneider, D. J., and Collmer, A. (2006). Closing the circle on the discovery of genes encoding Hrp regulon members and type III secretion system effectors in the genomes of three model Pseudomonas syringae strains. Mol. Plant Microbe Interact. 19, 1151–1158. doi: 10.1094/MPMI-19-1151
Liu, T., Liu, Z., Song, C., Hu, Y., Han, Z., She, J., et al. (2012). Chitin-induced dimerization activates a plant immune receptor. Science 336, 1160–1164. doi: 10.1126/science.1218867
Lozano-Duran, R., Bourdais, G., He, S. Y., and Robatzek, S. (2014). The bacterial effector HopM1 suppresses PAMP-triggered oxidative burst and stomatal immunity. New Phytol. 202, 259–269. doi: 10.1111/nph.12651
Lu, D., Wu, S., Gao, X., Zhang, Y., Shan, L., and He, P. (2010). A receptor-like cytoplasmic kinase, BIK1, associates with a flagellin receptor complex to initiate plant innate immunity. Proc. Natl. Acad. Sci. U.S.A. 107, 496–501. doi: 10.1073/pnas.0909705107
Luo, Q., Liu, W. W., Pan, K. D., Peng, Y. L., and Fan, J. (2017). Genetic interaction between Arabidopsis Qpm3.1 locus and bacterial effector gene hopW1-1 underlies natural variation in quantitative disease resistance to Pseudomonas infection. Front. Plant Sci. 8:695. doi: 10.3389/fpls.2017.00695
Macho, A. P., and Zipfel, C. (2015). Targeting of plant pattern recognition receptor-triggered immunity by bacterial type-III secretion system effectors. Curr. Opin. Microbiol. 23, 14–22. doi: 10.1016/j.mib.2014.10.009
McDowell, J. M., and Dangl, J. L. (2000). Signal transduction in the plant immune response. Trends Biochem. Sci. 25, 79–82. doi: 10.1016/S0968-0004(99)01532-7
Moore, J. W., Loake, G. J., and Spoel, S. H. (2011). Transcription dynamics in plant immunity. Plant Cell 23, 2809–2820. doi: 10.1105/tpc.111.087346
Moscou, M. J., and Bogdanove, A. J. (2009). A simple cipher governs DNA recognition by TAL effectors. Science 326:1501. doi: 10.1126/science.1178817
Mucyn, T. S., Clemente, A., Andriotis, V. M., Balmuth, A. L., Oldroyd, G. E., Staskawicz, B. J., et al. (2006). The tomato NBARC-LRR protein Prf interacts with Pto kinase in vivo to regulate specific plant immunity. Plant Cell 18, 2792–2806. doi: 10.1105/tpc.106.044016
Nakagawa, T., Kurose, T., Hino, T., Tanaka, K., Kawamukai, M., Niwa, Y., et al. (2007). Development of series of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. J. Biosci. Bioeng. 104, 34–41. doi: 10.1263/jbb.104.34
Navarro, L., Jay, F., Nomura, K., He, S. Y., and Voinnet, O. (2008). Suppression of the microRNA pathway by bacterial effector proteins. Science 321, 964–967. doi: 10.1126/science.1159505
Navarro, L., Zipfel, C., Rowland, O., Keller, I., Robatzek, S., Boller, T., et al. (2004). The transcriptional innate immune response to flg22. Interplay and overlap with Avr gene-dependent defense responses and bacterial pathogenesis. Plant Physiol 135, 1113–1128. doi: 10.1104/pp.103.036749
Nomura, K., Debroy, S., Lee, Y. H., Pumplin, N., Jones, J., and He, S. Y. (2006). A bacterial virulence protein suppresses host innate immunity to cause plant disease. Science 313, 220–223. doi: 10.1126/science.1129523
Ntoukakis, V., Mucyn, T. S., Gimenez-Ibanez, S., Chapman, H. C., Gutierrez, J. R., Balmuth, A. L., et al. (2009). Host inhibition of a bacterial virulence effector triggers immunity to infection. Science 324, 784–787. doi: 10.1126/science.1169430
Peart, J. R., Lu, R., Sadanandom, A., Malcuit, I., Moffett, P., Brice, D. C., et al. (2002). Ubiquitin ligase-associated protein SGT1 is required for host and nonhost disease resistance in plants. Proc. Natl. Acad. Sci. U.S.A. 99, 10865–10869. doi: 10.1073/pnas.152330599
Sawada, H., Suzuki, F., Matsuda, I., and Saitou, N. (1999). Phylogenetic analysis of Pseudomonas syringae pathovars suggests the horizontal gene transfer of argK and the evolutionary stability of hrp gene cluster. J. Mol. Evol. 49, 627–644. doi: 10.1007/PL00006584
Schechter, L. M., Vencato, M., Jordan, K. L., Schneider, S. E., Schneider, D. J., and Collmer, A. (2006). Multiple approaches to a complete inventory of Pseudomonas syringae pv. tomato DC3000 type III secretion system effector proteins. Mol. Plant Microbe Interact. 19, 1180–1192. doi: 10.1094/MPMI-19-1180
Schultink, A., Qi, T., Lee, A., Steinbrenner, A. D., and Staskawicz, B. (2017). Roq1 mediates recognition of the Xanthomonas and Pseudomonas effector proteins XopQ and HopQ1. Plant J. 92, 787–795. doi: 10.1111/tpj.13715
Scofield, S. R., Tobias, C. M., Rathjen, J. P., Chang, J. H., Lavelle, D. T., Michelmore, R. W., et al. (1996). Molecular basis of gene-for-gene specificity in bacterial speck disease of tomato. Science 274, 2063–2065. doi: 10.1126/science.274.5295.2063
Segonzac, C., Feike, D., Gimenez-Ibanez, S., Hann, D. R., Zipfel, C., and Rathjen, J. P. (2011). Hierarchy and roles of pathogen-associated molecular pattern-induced responses in Nicotiana benthamiana. Plant Physiol. 156, 687–699. doi: 10.1104/pp.110.171249
Shan, L., He, P., Li, J., Heese, A., Peck, S. C., Nurnberger, T., et al. (2008). Bacterial effectors target the common signaling partner BAK1 to disrupt multiple MAMP receptor-signaling complexes and impede plant immunity. Cell Host Microbe 4, 17–27. doi: 10.1016/j.chom.2008.05.017
Shinya, T., Yamaguchi, K., Desaki, Y., Yamada, K., Narisawa, T., Kobayashi, Y., et al. (2014). Selective regulation of the chitin-induced defense response by the Arabidopsis receptor-like cytoplasmic kinase PBL27. Plant J. 79, 56–66. doi: 10.1111/tpj.12535
Studholme, D. J., Ibanez, S. G., Maclean, D., Dangl, J. L., Chang, J. H., and Rathjen, J. P. (2009). A draft genome sequence and functional screen reveals the repertoire of type III secreted proteins of Pseudomonas syringae pathovar tabaci 11528. BMC Genomics 10:395. doi: 10.1186/1471-2164-10-395
Tang, X., Frederick, R. D., Zhou, J., Halterman, D. A., Jia, Y., and Martin, G. B. (1996). Initiation of plant disease resistance by physical interaction of AvrPto and Pto kinase. Science 274, 2060–2063. doi: 10.1126/science.274.5295.2060
Toruno, T. Y., Stergiopoulos, I., and Coaker, G. (2016). Plant-pathogen effectors: cellular probes interfering with plant defenses in spatial and temporal manners. Annu. Rev. Phytopathol. 54, 419–441. doi: 10.1146/annurev-phyto-080615-100204
Vinatzer, B. A., Jelenska, J., and Greenberg, J. T. (2005). Bioinformatics correctly identifies many type III secretion substrates in the plant pathogen Pseudomonas syringae and the biocontrol isolate P. fluorescens SBW25. Mol. Plant Microbe Interact. 18, 877–888. doi: 10.1094/MPMI-18-0877
Vinatzer, B. A., Teitzel, G. M., Lee, M. W., Jelenska, J., Hotton, S., Fairfax, K., et al. (2006). The type III effector repertoire of Pseudomonas syringae pv. syringae B728a and its role in survival and disease on host and non-host plants. Mol. Microbiol. 62, 26–44. doi: 10.1111/j.1365-2958.2006.05350.x
Wan, J., Zhang, X. C., Neece, D., Ramonell, K. M., Clough, S., Kim, S. Y., et al. (2008). A LysM receptor-like kinase plays a critical role in chitin signaling and fungal resistance in Arabidopsis. Plant Cell 20, 471–481. doi: 10.1105/tpc.107.056754
Washington, E. J., Mukhtar, M. S., Finkel, O. M., Wan, L., Banfield, M. J., Kieber, J. J., et al. (2016). Pseudomonas syringae type III effector HopAF1 suppresses plant immunity by targeting methionine recycling to block ethylene induction. Proc. Natl. Acad. Sci. U.S.A. 113, E3577–E3586. doi: 10.1073/pnas.1606322113
Wei, C. F., Kvitko, B. H., Shimizu, R., Crabill, E., Alfano, J. R., Lin, N. C., et al. (2007). A Pseudomonas syringae pv. tomato DC3000 mutant lacking the type III effector HopQ1-1 is able to cause disease in the model plant Nicotiana benthamiana. Plant J. 51, 32–46. doi: 10.1111/j.1365-313X.2007.03126.x
Wei, H. L., Chakravarthy, S., Mathieu, J., Helmann, T. C., Stodghill, P., Swingle, B., et al. (2015). Pseudomonas syringae pv. tomato DC3000 Type III secretion effector polymutants reveal an interplay between HopAD1 and AvrPtoB. Cell Host Microbe 17, 752–762. doi: 10.1016/j.chom.2015.05.007
Wroblewski, T., Caldwell, K. S., Piskurewicz, U., Cavanaugh, K. A., Xu, H., Kozik, A., et al. (2009). Comparative large-scale analysis of interactions between several crop species and the effector repertoires from multiple pathovars of Pseudomonas and Ralstonia. Plant Physiol. 150, 1733–1749. doi: 10.1104/pp.109.140251
Wulf, J., Pascuzzi, P. E., Fahmy, A., Martin, G. B., and Nicholson, L. K. (2004). The solution structure of type III effector protein AvrPto reveals conformational and dynamic features important for plant pathogenesis. Structure 12, 1257–1268. doi: 10.1016/j.str.2004.04.017
Xiang, T., Zong, N., Zou, Y., Wu, Y., Zhang, J., Xing, W., et al. (2008). Pseudomonas syringae effector AvrPto blocks innate immunity by targeting receptor kinases. Curr. Biol. 18, 74–80. doi: 10.1016/j.cub.2007.12.020
Xin, X. F., Nomura, K., Aung, K., Velasquez, A. C., Yao, J., Boutrot, F., et al. (2016). Bacteria establish an aqueous living space in plants crucial for virulence. Nature 539, 524–529. doi: 10.1038/nature20166
Xing, W., Zou, Y., Liu, Q., Liu, J., Luo, X., Huang, Q., et al. (2007). The structural basis for activation of plant immunity by bacterial effector protein AvrPto. Nature 449, 243–247. doi: 10.1038/nature06109
Yamada, K., Yamaguchi, K., Shirakawa, T., Nakagami, H., Mine, A., Ishikawa, K., et al. (2016). The Arabidopsis CERK1-associated kinase PBL27 connects chitin perception to MAPK activation. EMBO J. 35, 2468–2483. doi: 10.15252/embj.201694248
Yang, L., Teixeira, P. J., Biswas, S., Finkel, O. M., He, Y., Salas-Gonzalez, I., et al. (2017). Pseudomonas syringae type III effector HopBB1 promotes host transcriptional repressor degradation to regulate phytohormone responses and virulence. Cell Host Microbe 21, 156–168. doi: 10.1016/j.chom.2017.01.003
Yoshioka, H., Bouteau, F., and Kawano, T. (2008). Discovery of oxidative burst in the field of plant immunity: looking back at the early pioneering works and towards the future development. Plant Signal. Behav. 3, 153–155. doi: 10.4161/psb.3.3.5537
Zhang, J., Li, W., Xiang, T., Liu, Z., Laluk, K., Ding, X., et al. (2010). Receptor-like cytoplasmic kinases integrate signaling from multiple plant immune receptors and are targeted by a Pseudomonas syringae effector. Cell Host Microbe 7, 290–301. doi: 10.1016/j.chom.2010.03.007
Zhou, J., Wu, S., Chen, X., Liu, C., Sheen, J., Shan, L., et al. (2014). The Pseudomonas syringae effector HopF2 suppresses Arabidopsis immunity by targeting BAK1. Plant J. 77, 235–245. doi: 10.1111/tpj.12381
Keywords: Pseudomonas, effector, PTI, cell death, suppression, Nicotiana benthamiana
Citation: Gimenez-Ibanez S, Hann DR, Chang JH, Segonzac C, Boller T and Rathjen JP (2018) Differential Suppression of Nicotiana benthamiana Innate Immune Responses by Transiently Expressed Pseudomonas syringae Type III Effectors. Front. Plant Sci. 9:688. doi: 10.3389/fpls.2018.00688
Received: 25 January 2018; Accepted: 04 May 2018;
Published: 23 May 2018.
Edited by:
Zhengqing Fu, University of South Carolina, United StatesReviewed by:
Brian H. Kvitko, University of Georgia, United StatesFrederik Börnke, Leibniz-Institut für Gemüse-und Zierpflanzenbau (IGZ), Germany
Copyright © 2018 Gimenez-Ibanez, Hann, Chang, Segonzac, Boller and Rathjen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Selena Gimenez-Ibanez, c2VsZW5hLmdpbWVuZXpAY25iLmNzaWMuZXM=
†These authors have contributed equally to this work.