 Zhe Yan1,2
Zhe Yan1,2 Ana Pérez-de-Castro3
Ana Pérez-de-Castro3 Maria J. Díez3
Maria J. Díez3 Samuel F. Hutton4
Samuel F. Hutton4 Richard G. F. Visser2Anne-Marie A. Wolters2
Richard G. F. Visser2Anne-Marie A. Wolters2 Yuling Bai2*Junming Li1*
Yuling Bai2*Junming Li1*- 1The Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing, China
- 2Plant Breeding, Graduate School Experimental Plant Sciences, Wageningen University & Research, Wageningen, Netherlands
- 3Instituto Universitario de Conservación y Mejora de la Agrodiversidad Valenciana, Ciudad Politécnica de la Innovación, Universitat Politècnica de València, Valencia, Spain
- 4Gulf Coast Research and Education Center, University of Florida, Gainesville, FL, United States
Tomato yellow leaf curl virus (TYLCV) is a virus species causing epidemics in tomato (Solanum lycopersicum) worldwide. Many efforts have been focused on identification of resistance sources by screening wild tomato species. In many cases, the accession numbers were either not provided in publications or not provided in a consistent manner, which led to redundant screenings. In the current study, we summarized efforts on the screenings of wild tomato species for TYLCV resistance from various publications. In addition, we screened 708 accessions from 13 wild tomato species using different inoculation assays (i.e., whitefly natural infection and Agrobacterium-mediated inoculation) from which 138 accessions exhibited no tomato yellow leaf curl disease (TYLCD) symptoms. These symptomless accessions include 14 accessions from S. arcanum, 43 from S. chilense, 1 from S. chmielewskii, 28 from S. corneliomulleri, 5 from S. habrochaites, 4 from S. huaylasense, 2 from S. neorickii, 1 from S. pennellii, 39 from S. peruvianum, and 1 from S. pimpinellifolium. Most of the screened S. chilense accessions remained symptomless. Many symptomless accessions were also identified in S. arcanum, S. corneliomulleri, and S. peruvianum. A large number of S. pimpinellifolium accessions were screened. However, almost all of the tested accessions showed TYLCD symptoms. Further, we studied allelic variation of the Ty-1/Ty-3 gene in few S. chilense accessions by applying virus-induced gene silencing and allele mining, leading to identification of a number of allele-specific polymorphisms. Taken together, we present a comprehensive overview on TYLCV resistance and susceptibility in wild tomato germplasm, and demonstrate how to study allelic variants of the cloned Ty-genes in TYLCV-resistant accessions.
Introduction
Tomato yellow leaf curl disease (TYLCD) has been a global constraint to tomato (Solanum lycopersicum) production since the 1980s (Moriones and Navas-Castillo, 2000). Up till now, TYLCD is still one of the most devastating diseases of tomato. Infected susceptible tomato plants show symptoms that include yellowing, curling, and cupping of leaves, severe stunting and abortion of flowers and fruits, all of which can lead to yield reduction of up to 100% (Abhary et al., 2007). TYLCD can be caused by a cluster of related virus species including tomato yellow leaf curl virus (TYLCV), which belongs to the genus Begomovirus of the Geminiviridae family. TYLCV has a wide host range that includes tomato (S. lycopersicum), sweet pepper (Capsicum annuum), chili pepper (C. chinense), tobacco (Nicotiana tabacum), common bean (Phaseolus vulgaris), petunia (Petunia × hybrida), and lisianthus (Eustoma grandiflora) (Díaz-Pendón et al., 2010).
In nature, TYLCV is transmitted exclusively by the sweet potato whitefly Bemisia tabaci (Genn.) in a persistent-circulative manner (Gronenborn, 2007). B. tabaci is an invasive pest with global importance since more than 175 countries officially report the presence of B. tabaci (CABI 2017; Bemisia tabaci. In: Invasive Species Compendium1). B. tabaci is a complex consisting of at least 24 distinct species (De Barro et al., 2011). The Bemisia Middle East-Asia Minor 1 (MEAM1/B) and Mediterranean (MED/Q) are regarded as the most invasive and damaging species, and these are also the species that transmit TYLCV to tomato (De Barro et al., 2011; Ning et al., 2015). Serious damage in tomato production attributed to TYLCV was first reported in Israel in 1959 (Cohen and Antignus, 1994). Since then, the list of tomato production regions reporting TYLCV epidemics has grown (Barboza et al., 2014; Chinnaraja et al., 2016; Mabvakure et al., 2016). Among the countries for which B. tabaci incidence was reported, more than 70 also reported the occurrence of TYLCV [CABI 2016; TYLCV (leaf curl). In: Invasive Species Compendium2].
Domesticated tomato is known to be vulnerable to TYLCV infection, but resistance exists in wild tomato species (Ji et al., 2007b). Accessions exhibiting no TYLCD symptoms upon infection have been reported in a number of species, including S. arcanum, S. cheesmaniae, S. chilense, S. chmielewskii, S. corneliomulleri, S. galapagense, S. habrochaites, S. neorickii, S. pennellii, S. peruvianum, and S. pimpinellifolium (Ji et al., 2007b; Vidavski et al., 2008; De la Peña et al., 2010; Pereira-Carvalho et al., 2010; Tomás et al., 2011). So far, three TYLCV resistance genes have been cloned. Ty-1 and Ty-3 originate from S. chilense accession LA1969 and LA2779, respectively (Zamir et al., 1994; Ji et al., 2007a). They are two alleles of the same gene that is located on the long arm of tomato chromosome 6 and encodes an RNA-dependent RNA polymerase (RDR) (Verlaan et al., 2011, 2013). Ty-2, originating from S. habrochaites accession B6013 and located on the long arm of chromosome 11, encodes an NB-LRR gene (Yang et al., 2014; Yamaguchi et al., 2018). ty-5 is a loss-of-function mutant allele of the pelota gene located on chromosome 4. The mutation in ty-5 is caused by a T-to-G transversion in the coding region, which occured in cultivated tomato (Lapidot et al., 2015). However, it has also been suggested that ty-5 is derived from a complex of S. peruvianum accessions (Anbinder et al., 2009). In addition to these cloned genes, two resistance loci Ty-4 and Ty-6 have been mapped. Ty-4 is identified from S. chilense LA1932. This locus is located on the long arm of chromosome 3 and has a minor effect toward TYLCV resistance (Ji et al., 2009). Ty-6 is the most recently identified TYLCV resistance locus on the long arm of chromosome 10, presumably originating from S. chilense accessions LA1938 and LA2779 (Hutton and Scott, 2014).
Up till now, introgressions of Ty-1, Ty-2, and Ty-3 into cultivated tomato have been the major focus in breeding programs. Ty-2 based resistance can be overcome by TYLCV-related Tomato yellow leaf curl Sardinia virus (TYLCSV) (Barbieri et al., 2010) and breakdown of Ty-2 mediated resistance has been demonstrated recently by an isolate of the Mild strain of TYLCV (TYLCV-Mld) (Ohnishi et al., 2016). Ty-1-mediated resistance is not suitable to use under high disease pressure which leads to resistance breakage in some cases (García-Cano et al., 2008). Resistance breakage facilitates TYLCD epidemics, which urges plant breeders to continuously search for effective novel sources of resistance in the wild tomato gene pool. In multiple research programs wild tomato germplasm has been screened in order to identify accessions that can be utilized as sources of TYLCV resistance. Although these efforts have resulted in the identification of a number of sources exhibiting no TYLCD symptoms, the accession numbers of these sources were not consistently provided in publications, which led to redundant screenings in some cases. In this article, we summarize the results of previous resistance screening efforts and the use of different resistance resources in tomato introgression breeding. In addition, we report the identification of 138 tomato accessions that were TYLCD symptomless in a large-scale screening of 708 accessions from 13 wild tomato species. Finally, we discuss the potential use of the newly identified resources for TYLCV resistance in tomato breeding, in the context of donor species, the cloned Ty genes and viral titer levels.
Materials and Methods
Plant Materials
Wild tomato accessions were collected from Tomato Genetics Resource Center (TGRC), World Vegetable Center in Taiwan (previously the Asian Vegetable Research and Development Center, AVRDC), Centre for Genetic Resources, Netherlands (CGN), and Kentucky State University (KSU). The tomato cultivar S. lycopersicum cv. Moneymaker (MM) was included as susceptible control.
Whitefly-Mediated Natural Inoculation
Germplasm accessions were screened from July till September in the years 2012, 2013, and 2014 for TYLCD resistance using field assays and natural infection with whiteflies at the Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing. Seeds were germinated in petri-dishes on moist sterilized filter paper. Germinated seedlings were transferred to pots and placed into a plastic tunnel greenhouse. After transplanting, plants were continuously exposed to natural infection of whiteflies by opening the tunnel ventilation. From July till September in Beijing, China, natural incidence of whitefly with large vector population facilitates TYLCD epidemics. Experimental design was a randomized complete block design with two blocks and one to eight plants per plot depending on the number of available germinated seedlings. S. lycopersicum cv. MM was included as susceptible control and used in each block (five plants per block). An inbred line, TO-937 derived from S. pimpinellifolium material, was included in each block as a tolerant control.
Agrobacterium-Mediated Inoculation
For TYLCV disease assay using Agrobacterium-mediated inoculation in Wageningen, Netherlands, an infectious TYLCV-IL clone (pTYCz40a) was used. The method has been described in detail by Verlaan et al. (2011). The full length clone of TYLCV-IL genome was maintained in Agrobacterium tumefaciens strain LBA4404. Agrobacterium culture was grown, centrifuged and the pellet resuspended to OD600 = 0.5. Tomato plants used to screen for TYLCV resistance were grown under greenhouse conditions. The greenhouse was maintained at 23°C, 60% humidity and a 16/8 h day/night cycle. TYLCV Agro-inoculation was performed on plants at approximately three true leaves stage (around 21 days after sowing) as described by Verlaan et al. (2011). For most of the tested wild species, four plants per accession were inoculated and two plants were mock-inoculated. Only for S. peruvianum, S. chilense, and S. lycopersicum var. cerasiforme accessions, eight plants per accession were inoculated with the virus and four plants were mock-inoculated.
Disease Assessment
In China, plant responses were evaluated three times at 8, 10, and 12 weeks after sowing for TYLCD symptom development. On each date, each plant was rated using a 0 to 4 disease severity index (DSI) described by Friedmann et al. (1998), where 0 indicates no TYLCD symptoms, and 4 means severe TYLCD symptoms, remarkable yellowing, curling leaves and significant stunting in plant size. Intermediate scores, 0.5, 1.5, 2.5, and 3.5 were incorporated for more precise disease severity scoring. The final disease score of each accession was taken into account and the value was presented as mean of all the tested individuals. Only accessions that remained asymptomatic throughout the whole evaluation period have been regarded as resistant genotypes. At Wageningen, plants were scored for symptom development at 25, 35, 45, and 55 days after virus inoculation. Symptom severity was scored using the same scale as described above. Final results were the disease scores of the last evaluation at 55 days post inoculation. At 45 days after TYLCV inoculation, top young leaves were harvested for DNA isolation using cetyltrimethyl ammonium bromide (CTAB) based protocol (Fulton et al., 1995). The presence of TYLCV was detected by PCR using primers TYLCV-Picó-F and TYLCV-Picó-R as described by Picó et al. (1999a).
Virus-Induced Gene Silencing (VIGS) and Allele Mining
We used the virus-induced gene silencing (VIGS) construct targeting Ty-1/Ty-3 and followed the agro-infiltration procedures as described in Verlaan et al. (2013). To quantify viral titer and obtain full length cDNA sequences of the Ty-1/Ty-3 alleles, top leaves of plants infiltrated with VIGS constructs followed by TYLCV infection were harvested and grinded in liquid nitrogen using mortar and pestle. Total DNA was isolated using the CTAB based protocol (Fulton et al., 1995). For quantification of virus accumulation, the forward primer TYLCV-IS 1678F and the reverse primer TYLCV-CONS 1756R were used as described by Powell et al. (2012). Elongation factor 1α (EF) gene was used as a reference with primers: EF-F (5′-ATTGGAAACGGATATGCCCCT-3′) and EF-R (5′-TCCTTACCTGAACGCCTGTCA-3′). The amounts of viral DNA were calculated using the ΔΔCt method as described by Livak and Schmittgen (2001). Quantitative real-time PCR (qPCR) was carried in 10 μl reactions with a Bio-Rad iCycler iQ5 using SYBR Green Supermix according to the manufacturer’s protocol (Bio-Rad). Total RNA was extracted by using the RNeasy Plant Mini Kit (Qiagen) following the manufacturer’s protocol. DNase treatment was performed on 1 μg RNA (DNase I Amp. Grade) as described by the manufacturer (Invitrogen) and cDNA was synthesized using the iScript cDNA Synthesis Kit following the protocol (Bio-Rad).
For allele mining, full length cDNA sequences of the Ty-1/Ty-3 gene were obtained by PCR using a high-fidelity Phusion DNA polymerase (Thermo Fisher Scientific) with primers Ty-F7 and Ty-R5 (Verlaan et al., 2013). cDNA amplification was carried out with 30 cycles of denaturation (10 s at 98°C), annealing (30 s at 68°C), and extension (2 min at 72°C) followed by 7 min of further extension at 72°C. Blunt ended PCR products were cloned using the Zero Blunt PCR Cloning Kit (Invitrogen). After ligation, the constructs were transformed into competent Escherichia coli One Shot® TOP10 cells (Invitrogen). Recombinant clones were selected based on colony PCR using primers M13F (5′-GTAAAACGACGGCCAG-3′) and primer R7 that hybridizes within the insert (Verlaan et al., 2013). Plasmids of positive colonies were isolated using the QIAprep Spin Miniprep Kit (Qiagen). Plasmids were sequenced with different primer combinations covering the full length of the Ty-1/Ty-3 gene (M13F, F3, F7, F6, F4, and M13R) (Verlaan et al., 2013). Sequences were analyzed using SeqMan Pro 9 (DNA Star). The cDNA sequences of the Ty-1/Ty-3 alleles in MM, S. chilense LA1969 and S. chilense LA2779 were obtained from Verlaan et al. (2013); S. chilense LA1932, LA1938, and LA1971 from Caro et al. (2015). Alignments were made with MegAlign (DNA Star).
Results
Evaluation of Wild Tomato Species for TYLCD Resistance
We tested 701 accessions from 13 wild tomato species for resistance to TYLCD. The species that were represented in this screen included S. arcanum (24 accessions), S. cheesmaniae (7 accessions), S. chilense (51 accessions), S. chmielewskii (3 accessions), S. corneliomulleri (44 accessions), S. galapagense (2 accessions), S. habrochaites (36 accessions), S. huaylasense (4 accessions), S. lycopersicoides (2 accessions), S. neorickii (3 accessions), S. pennellii (38 accessions), S. peruvianum (81 accessions), and S. pimpinellifolium (406 accessions) (Supplementary Table S1).
These accessions were subjected to a field screening with natural whitefly infection. To eliminate phenotypic variations and to assess uniformity of the infection across the trial, the susceptible control S. lycopersicum cv. MM plants were placed randomly at different positions within the experimental block. Meanwhile, an inbred line, TO-937 derived from S. pimpinellifolium material was included as a tolerant control. TO-937 has the introgression of type IV leaf glandular trichomes which limits whitefly (B. tabaci) access and feeding (Rodríguez-López et al., 2011). All MM plants exhibited a homogenous and highly susceptible response (DSI of 4). All plants of line TO-937 showed clear TYLCD symptoms (average DSI of 3). The wild accessions screened in the present study showed a range of phenotypic reactions to TYLCD infection. For simplicity, phenotypic responses to TYLCD were initially categorized into two groups, symptomless and symptomatic. Among the accessions tested, we identified 133 symptomless accessions (Figure 1 and Supplementary Table S1). Proportionally, S. huaylasense and S. chilense displayed the most resistant accessions, with all four S. huaylasense accessions tested and more than 80% of the S. chilense accessions tested exhibiting no TYLCD symptoms (Figure 1). Among S. chilense accessions, four of the 54 screened accessions were symptomatic showing mild symptoms (Supplementary Table S2). From species of S. arcanum, S. corneliomulleri, and S. peruvianum a large number of symptomless accessions were identified. Of these three species, symptomatic accessions demonstrated mild to moderate levels of symptoms and severe symptoms were observed only in accessions LA1350 (S. arcanum), PI 199380 (S. corneliomulleri), and LA3218 (S. peruvianum) (Supplementary Table S2).
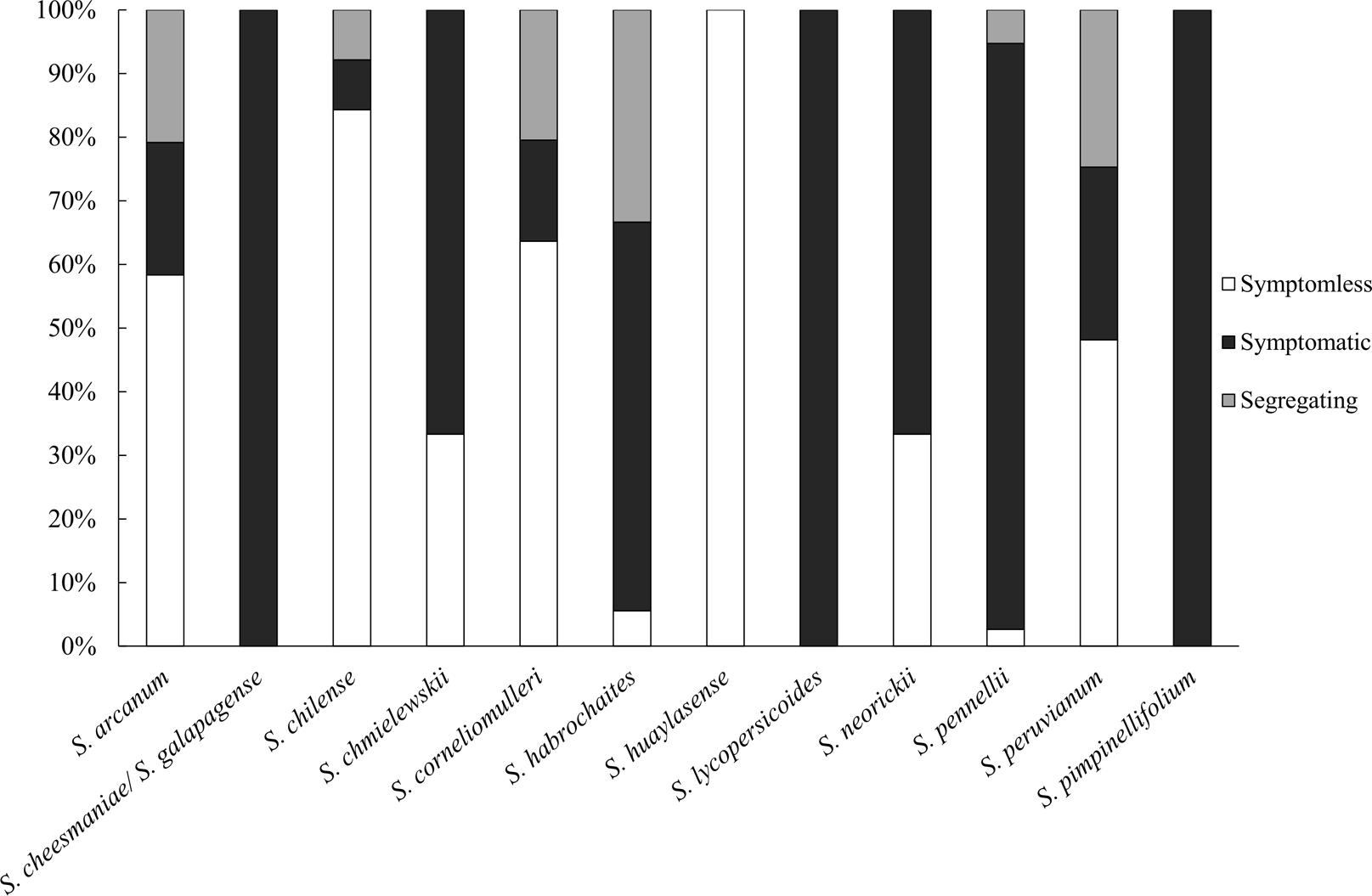
FIGURE 1. Stack bar graph depicting percentage of accessions for each Solanum spp. that exhibited symptomatic, symptomless, or segregating phenotypes upon TYLCD infection. Taxonomy of wild tomato relatives follow the classification system as presented in Peralta et al. (2008).
In contrast, the majority of S. pennellii and S. habrochaites accessions (92 and 61%, respectively) showed clear viral symptoms with severe yellowing and curling (Figure 1, Supplementary Figure S1 and Supplementary Table S2). From S. cheesmaniae, S. galapagense, S. lycopersicoides, and S. pimpinellifolium all the tested accessions displayed TYLCD symptoms (Figure 1, Supplementary Figure S1 and Supplementary Table S2). Particularly, all the 406 S. pimpinellifolium accessions showed TYLCD symptoms (Figure 1 and Supplementary Table S2) although symptom severity varied significantly among accessions. Mild TYLCD symptoms, slight yellowing and curling of the young leaves, were observed only in some accessions including LA1607, LA1344, LA2578, LA1863, LA0398, LA1630, and LA1589 (Supplementary Table S2).
Significant plant-to-plant variation in phenotypic responses was observed within several accessions of tomato wild species, including S. arcanum (5 accessions), S. chilense (4 accessions), S. corneliomulleri (9 accessions), S. habrochaites (12 accessions), S. pennellii (2 accessions), and S. peruvianum (20 accessions) (Figure 1 and Supplementary Table S3). This variation may be due to actual segregation of resistance alleles resulting from the heterogeneous nature of these out-crossing species (Peralta and Spooner, 2005; Moyle, 2008). Alternatively, it may also be due to the presence of escapes, since this test was conducted using natural infection in the field.
At Wageningen, 11 different wild tomato species (including one S. lycopersicum line as control) were screened and evaluated for their responses to TYLCV using Agrobacterium-mediated inoculation (Table 1). This screening panel included 32 accessions of the following species, S. arcanum (1 accession), S. cheesmaniae (2 accessions), S. chilense (1 accession), S. corneliomulleri (1 accession), S. galapagense (1 accession), S. habrochaites (10 accessions), S. lycopersicoides (1 accession), S. neorickii (1 accession), S. peruvianum (11 accessions), and S. pimpinellifolium (3 accessions). Twenty out of the 32 tested accessions showed no TYLCD symptoms (Table 1). Plants of S. arcanum LA2172, S. chilense LA0458, S. corneliomulleri CGN14358, S. lycopersicoides CGN23973, S. neorickii LA0735, and all the tested accessions of S. peruvianum were completely free of symptoms. Plants of S. habrochaites accessions LA4137, LA1777, and LA1718 also exhibited no TYLCV symptoms, while plants of the other tested S. habrochaites accessions showed severe symptoms. S. pimpinellifolium accession CGN15528 did not show viral symptoms. The other tested accessions (LA1584 and PI 365967) of S. pimpinellifolium exhibited clear TYLCV symptoms with yellowing and curling of the young leaves. Plants of the S. galapagense accession and S. cheesmaniae accession LA1409 were also susceptible to TYLCV. S. cheesmaniae accession PI 266375 segregated for its response to TYLCV infection. Of the 10 individuals tested, two plants exhibited no TYLCV symptoms, while the rest displayed a mild to moderate level of susceptibility (Table 1). Among 20 TYLCV symptomless accessions, 14 accessions were tested for virus status. Viral DNA was detected in the top young leaves of all the tested plants (Supplementary Table S4).
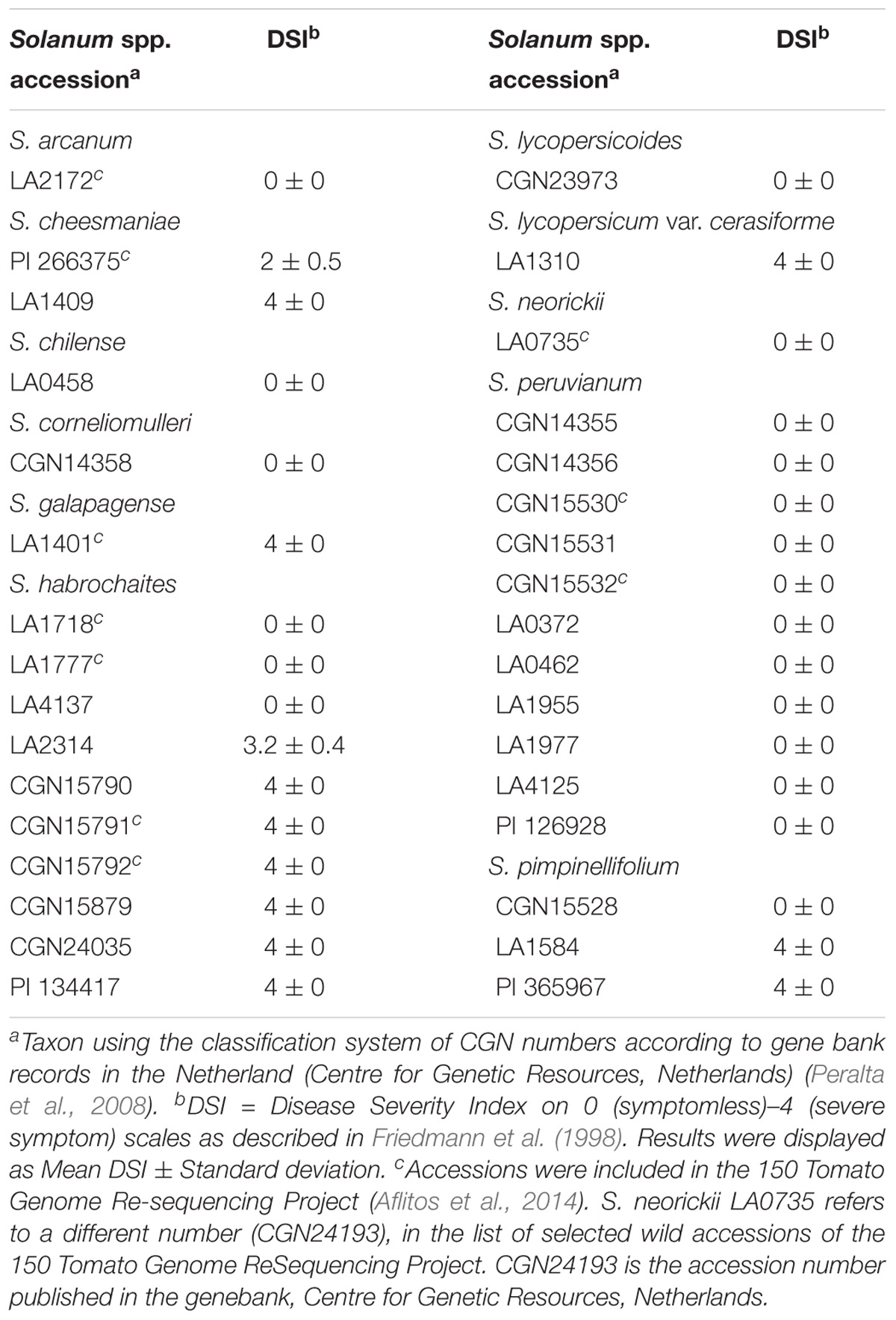
TABLE 1. Average disease severity index of wild tomato species screened for resistance to Tomato Yellow Leaf Curl Virus (TYLCV) using Agrobacterium-mediated inoculation with an infectious TYLCV-IL clone at Plant Breeding, Wageningen University & Research, Netherlands.
Responses of the Same Tomato Species to TYLCV Infection Using Different Inoculation Methods
Wild tomato accessions have been tested in China and Netherlands using natural whitefly infection and agroinoculation, respectively. Some of the accessions were screened using both inoculation methods leading to similar results (Table 2). Plants of S. arcanum accession LA2172, S. chilense LA0458, S. corneliomulleri CGN14358, and six out of the 11 screened S. peruvianum accessions (CGN15530, CGN15532, LA0372, LA1977, LA4125, and PI 126928) were free of viral symptoms (Table 2). While, plants of S. cheesmaniae accession LA1409, six S. habrochaites accessions (CGN15790, CGN15791, CGN15792, CGN15879, CGN24035, and PI 134417) and S. pimpinellifolium accession LA1584 showed severe symptoms (Table 2).
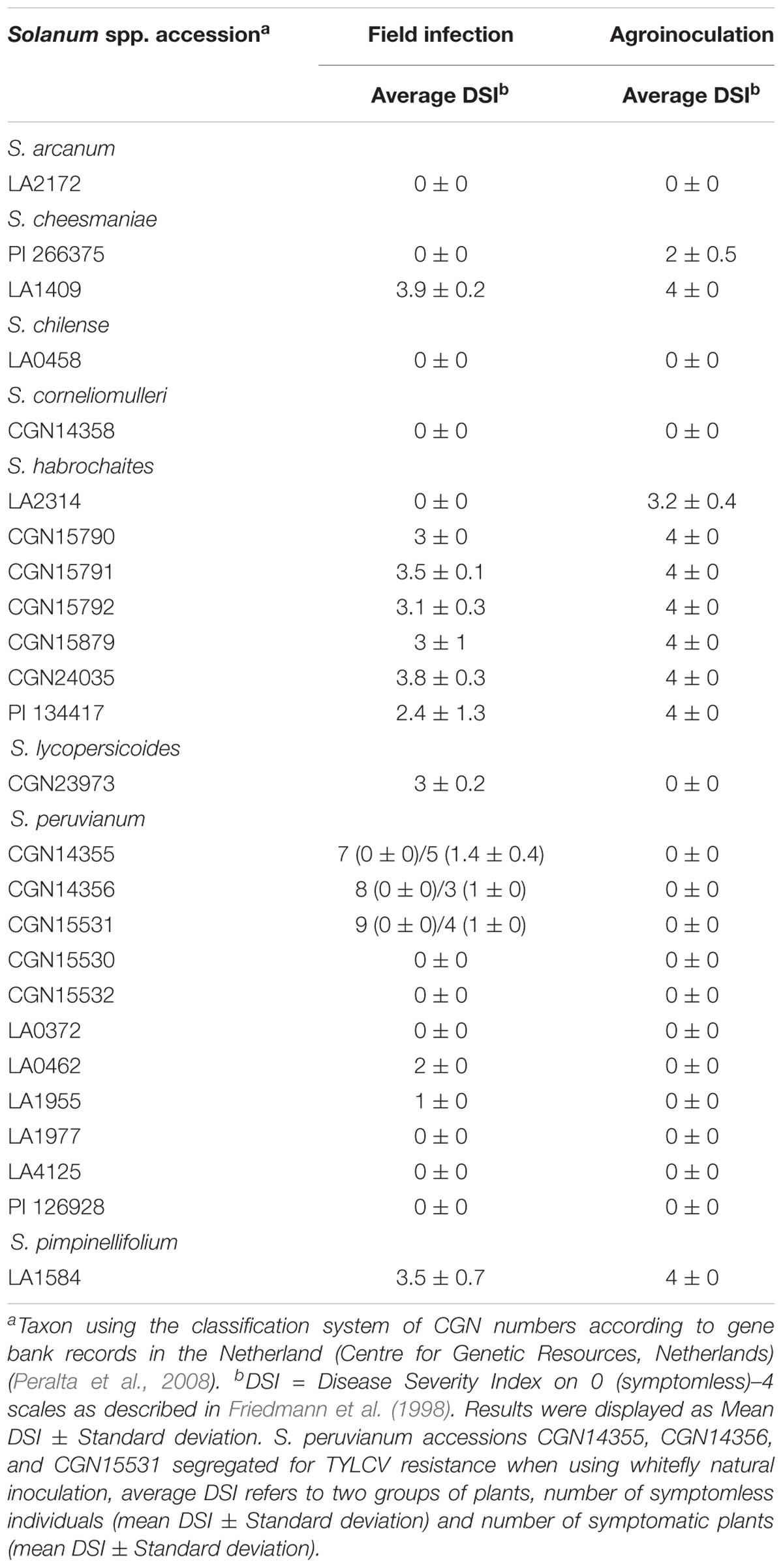
TABLE 2. Disease severity index of TYLCD infection in wild tomato species after artificial Agrobacterium-mediated inoculation and field infection with whitefly.
Also, contrasting results of the same accession were observed. S. habrochaites LA2314, which was symptomless in the field with whitefly infection, exhibited clear TYLCD symptoms with the agroinfection method (Table 2). In contrast, symptomatic accessions in the field test, such as S. lycopersicoides CGN23973 as well as S. peruvianum LA0462 and LA1955, showed no symptoms using Agrobacterium-mediated inoculation (Table 2). Among the accessions that were heterogenic in response to whitefly natural infection, we tested three accessions from S. peruvianum (CGN14355, CGN14356, and CGN15531) with Agrobacterium mediated TYLCV inoculation and all the tested plants were free of viral symptoms (Table 2).
Presence of Functional Ty-1 and Ty-3 Alleles in Different S. chilense Accessions
Previous studies indicated that the resistance to TYLCV in S. chilense accessions LA1932, LA1938, LA1960, and LA1971 is due to the presence of functional Ty-1/Ty-3 alleles (Caro et al., 2015). To verify whether the S. chilense accessions showing no TYLCD symptom in this study carry a functional Ty-1/Ty-3 allele, VIGS was applied to silence the Ty-1/Ty-3 gene in six S. chilense accessions (Table 3). These accessions were selected because relatively more seeds were available.
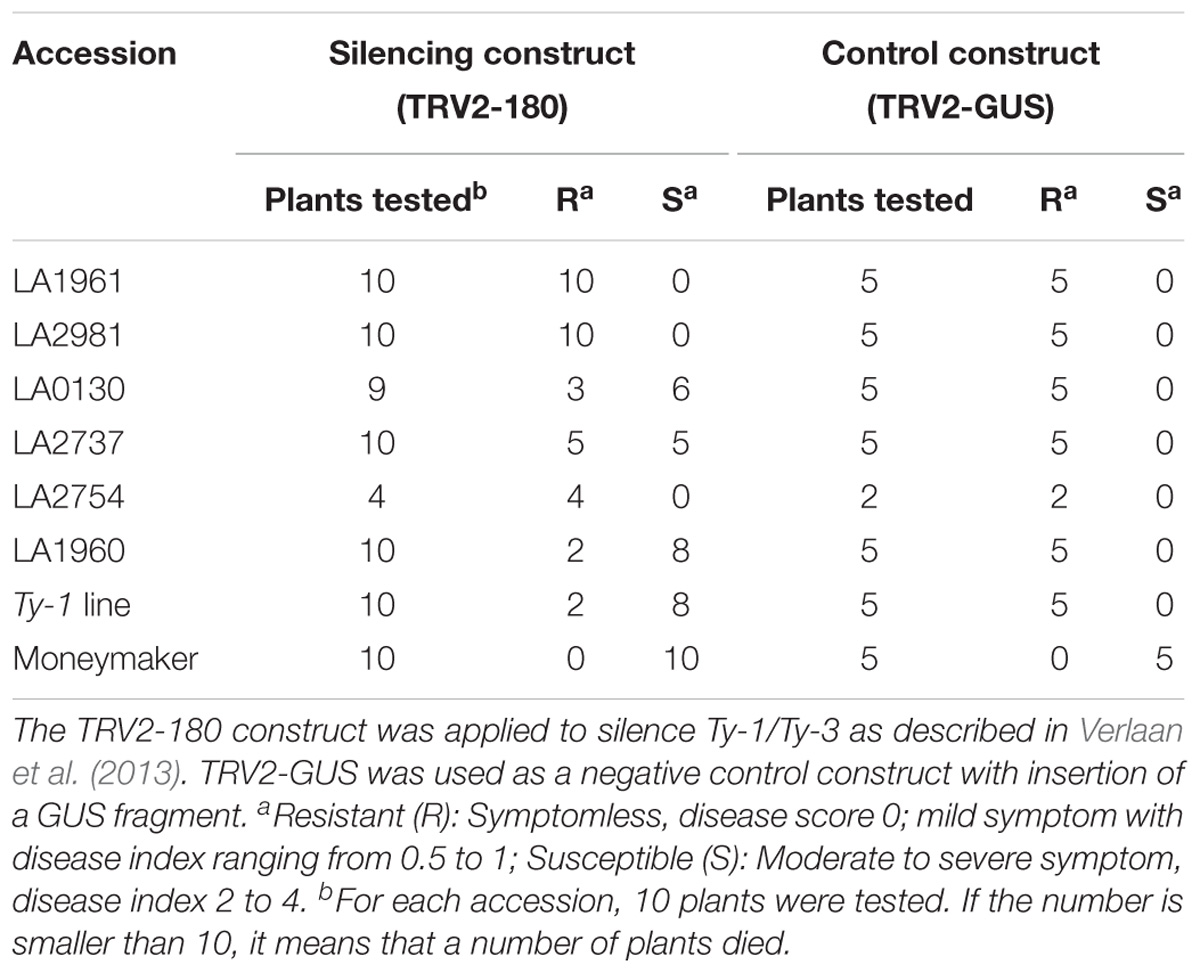
TABLE 3. Silencing of Ty-1/Ty-3 compromises TYLCV resistance in multiple S. chilense accessions.
After TYLCV inoculation, susceptible MM plants showed severe TYLCV symptoms 20 days post inoculation with TYLCV (Table 3). An advanced breeding line harboring the Ty-1 gene (named hereafter as the Ty-1 line) was included as a positive control. Plants of the Ty-1 line, which were infiltrated with the TRV2-GUS construct remained symptomless. While, eight out of 10 plants of the Ty-1 line, which were infiltrated with the TRV2-180 construct to silence the Ty-1 gene, displayed TYLCV symptoms (Table 3). However, not all the TRV2-180 infiltrated plants showed symptoms, showing that the silencing effect of VIGS was not uniform.
All five plants of S. chilense accessions LA1961, LA2981, LA0130, LA2737, LA2754, and LA1960 infiltrated with the TRV2-GUS construct remained resistant, demonstrating that TYLCV resistance in these accessions was not influenced by the VIGS vector, tobacco rattle virus (TRV). Of accessions LA0130, LA2737, and LA1960, a number of plants infiltrated with the TRV2-180 construct showed TYLCV symptoms (Table 3), indicating that the resistance of these accessions was compromised upon silencing the Ty-1 gene (Table 3). We also quantified virus concentration in susceptible TRV2-180 plants of the accessions LA0130, LA2737, and LA1960 and compared it with that of the resistant TRV2-180 plants (no compromised resistance after silencing) and the TRV2-GUS plants. Significantly more virus accumulation was detected in silenced Ty-1/Ty-3 plants compared with that of the other two groups (Figure 2). All the plants of the accessions LA1961 and LA2981 tested with the TRV2-180 construct remained resistant (Table 3). Ten plants of the accession LA2754 were infiltrated with TRV2-180 construct and six died. The remaining ones did not show symptoms after TYLCV inoculation (Table 3).
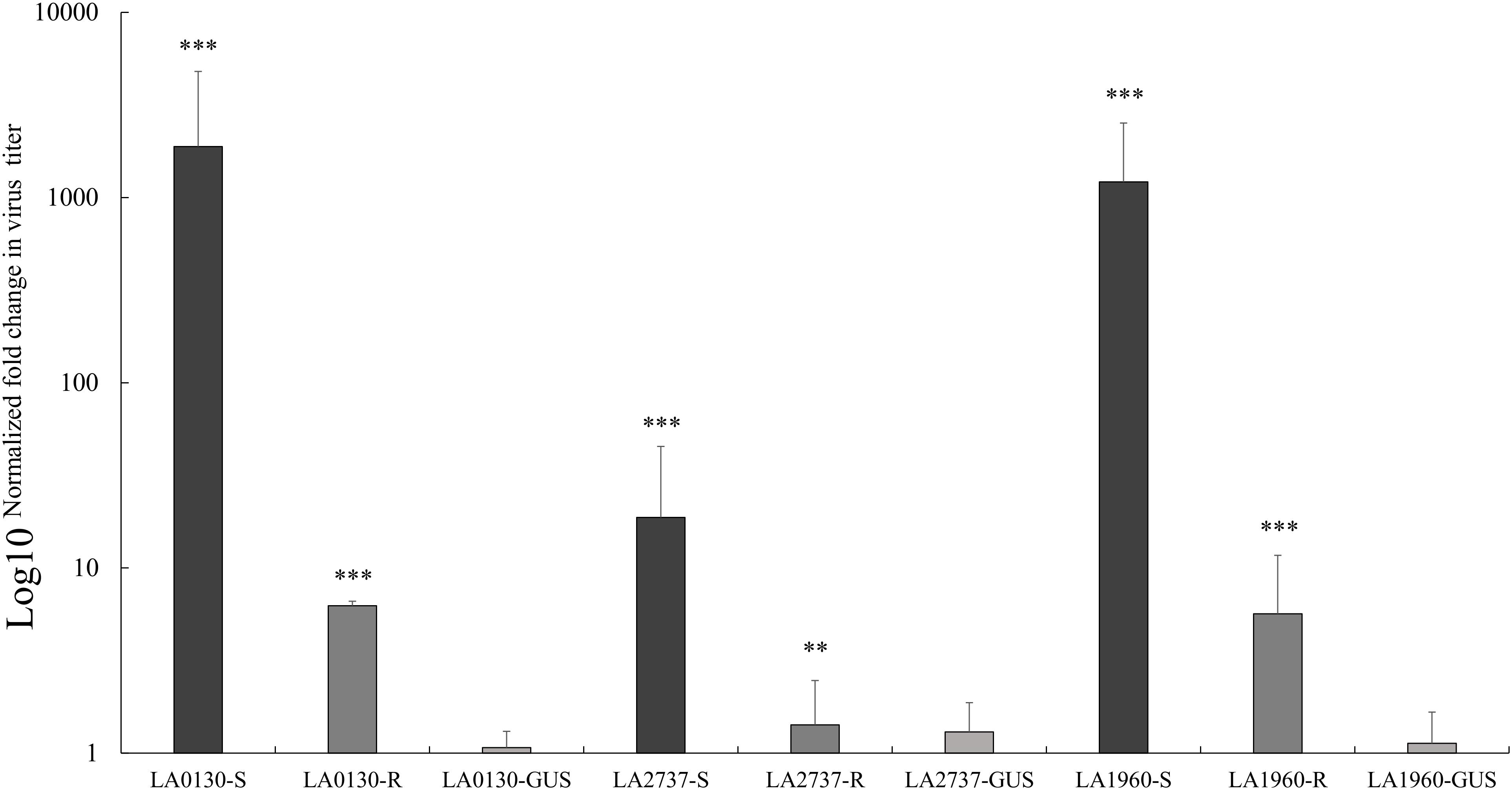
FIGURE 2. Relative quantification of virus accumulation by qPCR in S. chilense accessions LA0130, LA2737, and LA1960 upon silencing Ty-1/Ty-3. For each accession, tested individuals infiltrated with silencing construct were grouped according to their phenotypic responses; R, resistant, S, compromised resistance. The amounts of viral DNA were normalized using the Tomato elongation factor 1α (EF). For each accession, the relative level of viral DNA was calibrated to the DNA level in the respective control plants infiltrated with the TRV2-GUS vector. Error bars represent standard deviation of biological replicates. Virus titer was converted into logarithmic scale (log10) displayed on the vertical axis. Asterisks indicate significant difference between TRV2-180 plants and average of TRV2-GUS plants according to one way analysis of variance. Number of asterisks indicate the degree of significance (∗∗p < 0.01 and ∗∗∗p < 0.001).
Intraspecific Sequence Variations in Ty-1/Ty-3 Resistant Alleles
To explore Ty-1/Ty-3 allele-specific polymorphisms, the sequences of the coding regions from S. chilense accessions LA0130, LA2737, and LA1960 were analyzed (Supplementary Figure S2). Nine polymorphisms representing three unique alleles were identified, of which seven SNPs specific for accession LA0130, one for LA1960 and one for LA2737. All these SNPs differ from the Ty-1/Ty-3 alleles uncovered from previous studies (Verlaan et al., 2013; Caro et al., 2015). Further, the protein sequences obtained by in silico translating the amplified coding sequences were aligned (Supplementary Figure S3). Four allele-specific amino acids (aa) were observed to be unique and not present in other RDR alleles identified before. For S. chilense accession LA1960, the allele-specific aa is F22 (Supplementary Figure S3, yellow marked). LA0130-specific aa are V139, Q805, and K912 (Supplementary Figure S3, yellow marked). In addition, a previous study showed that TYLCV-resistant S. chilense species carry three Ty-1/Ty-3 specific aa (Caro et al., 2015), which are present in the tested accessions (Supplementary Figure S3, red marked).
Previously Published Data on Screening of Accessions With Resistance and Susceptibility to TYLCV
In Spain, numerous wild tomato species have been screened for resistance to TYLCD by researchers at the Institute for the Conservation and Improvement of Agrodiversity (COMAV). These screens utilized whitefly-mediated and Agroinoculation methods with either TYLCV or TYLCSV, and some accessions were tested with both viruses. TYLCSV is the first reported geminivirus in Spain. The screening work started in mid-1990s, when TYLCSV was the predominant TYLCV-related Begomovirus present in Spain (Moriones et al., 1993). In total, 143 accessions were screened for resistance, the results of which we summarize from nine publications (Jordá et al., 1996; Picó and Díez, 1998; Picó et al., 1998, 1999b, 2000; Soler et al., 2000; Pérez de Castro et al., 2004, 2005, 2010).
Among 143 accessions tested, highly and moderately resistant accessions were found in S. chilense (17 accessions), S. lycopersicum var. cerasiforme (1 accession), S. habrochaites (4 accessions), S. lycopersicoides (7 accessions), S. neorickii (1 accession), S. peruvianum (7 accessions), and S. pimpinellifolium (5 accessions) (Supplementary Table S5). Accessions that were resistant against TYLCSV were identified in S. chilense (LA1932 and LA1963), S. habrochaites (ECU336 and ECU436), S. neorickii (ECU301), S. peruvianum (ECU446, PI 126935, and PI 143679), S. pimpinellifolium (UPV16953) and S. lycopersicum var. cerasiforme (ECU464) (Supplementary Table S5). Later, TYLCV was introduced in Spain and was reported worldwide causing serious TYLCD epidemics, thus, this viral species was also included in some of the screenings. All the resistant accessions identified in S. lycopersicoides were effective against TYLCV. Considering the importance of both viral species, in many of the screenings, phenotypic responses to both TYLCV and TYLCSV were evaluated. These intensive screenings resulted in identification of many accessions being resistant to both TYLCV species from S. chilense, S. habrochaites, S. peruvianum, and S. pimpinellifolium, indicating a very strong correlation between resistance to TYLCV and resistance to TYLCSV (Supplementary Table S5). The highest levels of resistance were found among S. chilense and S. peruvianum accessions when challenged with two viral species in different inoculation assays. High variability was found among S. pimpinellifolium and S. habrochaites accessions. Intermediate level of resistance was found in S. pimpinellifolium accessions with mild to moderate viral symptoms (Supplementary Table S5). S. pennellii accessions included in the screenings were rated as susceptible (Supplementary Table S5; Picó et al., 2000; Soler et al., 2000).
In addition to the above mentioned large scale tests carried out at COMAV, we summarized resistant and susceptible accessions in previously published resources (Supplementary Table S5). Worldwide efforts on identification of promising resistance sources against TYLCV viral complex have resulted in various resistant accessions corresponding to S. arcanum, S. cheesmaniae, S. chilense, S. chmielewskii, S. corneliomulleri, S. galapagense, S. habrochaites, S. neorickii, S. pennellii, S. peruvianum, and S. pimpinellifolium (Supplementary Table S5 with literature). Resistant accessions identified up till now from previous publications were mainly from wild species S. chilense, S. habrochaites, S. peruvianum, and S. pimpinellifolium (Supplementary Table S5). All the identified S. chilense accessions showed a high level of resistance exhibiting no viral symptoms. Among S. peruvianum and S. habrochaites accessions, both high levels of resistance (no symptoms) as well as mild to moderate levels of resistance were identified. Resistant accessions from S. pimpinellifolium mostly showed a moderate level of resistance which allows slight viral symptoms (Supplementary Table S5).
Introgression Breeding for TYLCV Resistance From Diverse Tomato Genetic Pools
Currently, Ty-1, Ty-2, and Ty-3 are the primary resistance genes widely used in tomato breeding programs reported in literature. The Ty-4 resistance locus confers only a low level of resistance, while the ty-5 gene is recessive in nature; therefore, the utilization of these genes in tomato breeding programs is restricted (Ji et al., 2009; Lapidot et al., 2015). Ty-6 is an incompletely dominant resistance locus that was more recently identified (Hutton and Scott, 2014). Little is known about Ty-6 or the extent to which it is utilized commercially. Ty-1, Ty-3, Ty-4, and (reportedly) Ty-6 all originated from various S. chilense accessions, including LA1969, LA2779, LA1932, and LA1938 (Zamir et al., 1994; Ji et al., 2007a, 2009; Hutton and Scott, 2014). In some cases, a single S. chilense accession can harbor more than one TYLCV resistance locus (e.g., LA1932 contains both Ty-1 and Ty-4; LA2779 contains both Ty-3 and Ty-6) (Ji et al., 2007a, 2009; Hutton and Scott, 2014; Caro et al., 2015). Resistance in commercial breeding materials can likewise be mediated by a single resistance gene or a joint response of different genes.
At COMAV, promising resistance sources in a number of wild species, such as S. pimpinellifolium, S. peruvianum, and S. chilense, were used to develop advanced resistant generations. UPV16991 was the most resistant S. pimpinellifolium accession identified. Resistance in L102, a UPV16991-derived inbred line, was monogenic and incompletely dominant. The resistance was characterized by a restriction of viral particle accumulation (Pérez de Castro et al., 2007). Pyramiding resistance derived from UPV16991 and the Ty-1 gene increased the level of resistance in different crosses between Ty-1 lines (BC7S1 generation from the cross S. lycopersicum NE-1 and LA3473) and UPV16991-derived lines (Pérez de Castro et al., 2008). In S. peruvianum, PI 126944 was selected for resistance to TYLCV (Picó and Díez, 1998). Three interspecific hybrids were obtained between cultivated tomato NE-1 and PI 126944, and a set of introgression lines into the genetic background of tomato is being developed from these (Picó et al., 2002). In some of these generations the resistance level against TYLCV and TYLCSV has been assessed (Julián et al., 2013). Interspecific hybrids between cultivated tomato (S. lycopersicum) and S. chilense accessions (i.e., LA1932, LA1938, LA1959, LA1960, LA1963, LA1969, LA1971, LA2779, LA3473, UPV20306, UPV20328, and UPV20329) were made leading to different types of populations (Picó et al., 1999c, 2002; Pérez de Castro et al., 2013). In many S. chilense accessions including LA1932, LA1938, LA1960, and LA1971, TYLCV resistance loci have been mapped to tomato chromosome 6, in the Ty-1/Ty-3 region (Caro et al., 2015). Interspecific hybrids were obtained between S. lycopersicum and S. chilense accessions UPV20306, UPV20328, and UPV20329. However, the level of resistance derived from these accessions seemed to be lower than resistance derived from the rest of the exploited sources (Pérez de Castro et al., 2005).
At the World Vegetable Center in Taiwan, breeding for resistance against TYLCD has focused intensely on Ty-2-mediated resistance, and Ty-2 has been extensively exploited in tomato breeding worldwide. CLN2116 is an F7 determinate tomato line developed at AVRDC with Ty-2 introgression (AVRDC, 2001). Three breeding lines namely CLN2513, CLN2514, and CLN2515 contain combined resistance derived from Ty-1 and Ty-2. These lines have been generated by double crossing three tomato lines (CLN399, CH154, and CLN2026) with BL982 (Ty-1-based resistance) as well as CLN2116 (AVRDC, 2002). Multiple determinate tomato lines with TYLCD resistance have been developed including CLN2468A, CLN2468B, CLN2468C, CLN2468D, CLN2469C, CLN2460F, CLN2467E, CLN2467F, and CLN2467G. At the same time, indeterminate tomato lines including CLN2460G, CLN2460H, CLN2460I, CLN2460J, CLN2463O, and CLN2463P are available for breeding purposes. All these breeding lines possess Ty-2 derived originally from H24 (AVRDC, 2003). TYLCD-resistant breeding line CLN2777A was derived from the cross between CL5915 and H24, a carrier of the Ty-2 locus. There are also breeding lines available at AVRDC carrying various combinations of TYLCD resistance loci. CLN3150A-5 has excellent disease resistance, being homozygous for Ty-2 and ty-5 genes (AVRDC, 2016). There are various improved lines developed at the World Vegetable Center with TYLCD resistance mediated by Ty-2 or in combination with other known TYLCD resistance genes (i.e., Ty-1/Ty-3 and ty-5). CLN2498D3 and CLN3024A3 are both (semi-) determinate lines carrying TYLCD resistance gene, Ty-2. CLN3241H-273 and CLN3241Q3 are proper candidates for open field cultivation with a high level of TYLCV resistance achieved by pyramiding Ty-1/Ty-3 and Ty-2 resistance genes. FMTT1733D3 and FMTT1733E3 are indeterminate lines harboring Ty-2 and Ty-3. CLN3736D3 is a semi-determinate open field line with superior TYLCD resistance driven by the combined effort of Ty-1/Ty-3, Ty-2, and ty-5. Cultivars or improved breeding lines developed at the World Vegetable Center are available for breeding purposes or for local agronomic performance testing for future release.
Begomovirus resistance breeding efforts at the University of Florida, Gulf Coast Research and Education Center (GCREC) began in the early 1990s. Although initial breeding efforts involved a larger number of S. chilense accessions, resistance derived from S. chilense accessions LA1932, LA1938, LA2779 and from S. lycopersicum cv. Tyking later became the primary focus of the program. Ty-1 and Ty-3 are widely used for TYLCV resistance in many breeding programs worldwide. However, undesirable horticultural traits are generally coupled with both introgressions, known as linkage drag. Recent breeding efforts at GCREC have focused on reducing the size of Ty-1 and Ty-3 introgressions. Fla.8923 is a product of these efforts, and contains Ty-3 within a 70 Kb S. chilense introgression (Hutton et al., 2015). Similarly, Fla.7907C and Fla.7781B each contain Ty-1 within an approximately 1 Mb introgression (Hutton and Scott, 2017; Hutton, Unpublished). Fla.8624 is a breeding line containing Ty-6 showing an intermediate level of resistance to TYLCV. Fla.8638B is a breeding line pyramiding ty-5 and Ty-6, displaying a high resistance level against a wide range of Begomoviruses (Scott et al., 2015).
Besides the three major centers (COMAV, AVRDC, and GCREC) that have generated advanced breeding lines and released them for breeding purposes, there are other programs aiming at deploying resistant wild tomato species to breed for TYLCV resistance. Various advanced populations have been developed starting from S. habrochaites accessions. The majority of them were derived from accession B6013, which is the donor of the Ty-2 resistance gene (Kalloo and Banerjee, 1990; Picó et al., 2002; Chomdej et al., 2007). Besides accession B6013, promising resistant lines were developed using S. habrochaites accessions LA1777 and LA0386 as well as EELM-889 (Vidavsky and Czosnek, 1998; Tomás et al., 2011). Combined resistance derived from both LA1777 and LA0386 has been introgressed into cultivated tomato. This resulted in segregating families displaying responses to TYLCV ranging from resistance and tolerance to susceptibility. A BC1S4 inbred line (named 902) that is fixed for TYLCV resistance was obtained (Vidavsky and Czosnek, 1998). S. habrochaites accession EELM-889 provides effective resistance against multiple TYLCD associated viruses (Tomás et al., 2011). Segregating populations (F2, F3, and BC1) were developed to further characterize EELM-889 mediated resistance against the Israel strain of TYLCV-IL. TYLCV-IL is the most widespread and economically important TYLCD causing Begomovirus species (Lefeuvre et al., 2010). The resistance to TYLCV-IL is controlled by two distinct loci, one dominant and another recessive (Tomás et al., 2011). In addition, resistant accessions from S. peruvianum were selected including PI 126935, PI 126926, PI 128648, and EC104395 to initiate the introgression of resistance into the cultivated tomato (Pilowsky and Cohen, 1990; Vidavsky et al., 1998; Maruthi et al., 2003). Resistance sources have also been uncovered from S. pimpinellifolium LA1921 and efforts have been taken to further obtain different populations (backcross, F1, and F2) (Banerjee and Kalloo, 1987). Results obtained by testing these populations for their response to TYLCV infection indicated that the resistance mediated by LA1921 was monogenic and incompletely dominant (Banerjee and Kalloo, 1987).
Discussion
In this study, we presented our own and worldwide efforts so far on the identification of potential tomato wild accessions for resistance to TYLCD. Out of more than 700 accessions of 13 wild tomato species, about 140 accessions showed a symptomless response to TYLCD infection, either by agroinfiltration or by natural whitefly infection. Virus replication was detected in most symptomless accessions identified by agroinfiltration, showing that these examined accessions are not immune to TYLCV. Further, unique functional Ty-1/Ty-3 alleles are present in S. chilense accessions LA0130, LA2737, and LA1960. Meanwhile, by summarizing to date the breeding efforts in breeding tomato with TYLCV resistance, we demonstrated that a very small number of resources has been used as resistance genitors in commercial cultivars. TYLCV has a great potential to change due to factors including virus recombination, mutations, additions of satellite and invasion of exogenous whitefly species (Nawaz-ul-Rehman and Fauquet, 2009; Czosnek and Ghanim, 2011; Hosseinzadeh et al., 2014). For example the TYLCV-IS76 strain, a recombinant between TYLCV-IL and the Spanish strain of TYLC-Sardinia virus (TYLCSV-ES), which can accumulate better than its parental strains in cultivars carrying the Ty-1 gene (Belabess et al., 2015, 2016). It has been shown that Ty-2 based resistance can be overcome by TYLCSV and the TYLCV-Mld strain which has a TYLCSV-like C4 protein (Barbieri et al., 2010; Tomás et al., 2011; Ohnishi et al., 2016). Therefore, the TYLCV symptomless accessions identified in this study represent a treasure of resources to tomato breeders.
Our focus of the current study was on the screening of germplasm collection for resistance to TYLCV and we defined resistance only as symptomless response to TYLCV. It should be noted that majority of the accessions was screened in the field using whitefly-mediated natural inoculation. For some accessions symptomless plants may have occurred due to escape or avoidance of whitefly infection. Therefore, the true TYLCV resistance should be determined by retesting these symptomless accessions with controlled inoculation approaches as well as by quantifying virus titers. Here, we discuss several issues in the context with follow-up studies and with the further use of these accessions for breeding purpose.
The Avoidance/Resistance to Whitefly
Previous screening studies on S. peruvianum under whitefly inoculation reported high levels of resistance to TYLCV. The resistance was overcome under graft-inoculated conditions for the same accessions (Azizi et al., 2008), suggesting a possible resistance to whitefly rather than to TYLCV. Similar results were obtained in the present study for S. habrochaites accession LA2314, symptomless in whitefly-mediated screening and symptomatic in Agrobacterium-mediated inoculation (Table 2). These results indicate that symptomless response of S. habrochaites accession LA2314 is possibly associated with the resistance to whitefly, probably owning to the presence of glandular trichomes. In various wild tomato species, the presence of glandular trichomes on the leaf surface contributes to whitefly resistance by entrapping the whiteflies, and thereby possibly changing their feeding behavior (Momotaz et al., 2010; Rodríguez-López et al., 2011; Firdaus et al., 2012; Andrade et al., 2017; Rakha et al., 2017). In Solanum spp., type IV and VI trichomes contribute to a high level of resistance to whitefly, which is attributed to the exudates of glandular trichomes (Kennedy, 2003). Secondary metabolites released by tomato glandular trichomes influence the whiteflies’ preference for or avoidance of specific plants, and their feeding behavior (Bleeker et al., 2009, 2011; Rodríguez-López et al., 2011). Thus, accessions displaying natural resistance against the transmission vector may not be resistant to its transmittable viral species. Before using these symptomless accessions identified upon whitefly natural infection, it is worthwhile to test the selected accessions with the viral species under investigation using Agrobacterium-mediated inoculation.
Resistance to TYLCV or to TYLCV-Like Viruses
In screenings worldwide, accessions were tested with the virus strain that was endemic to the area. This approach likely contributes to the identification of strain-specific resistant accessions that may benefit regional breeding programs. In the field, mixed infections frequently occur. As transmission vector, it is possible for whiteflies to transmit different types of virus as well as various TYLCV-like viral species (Díaz-Pendón et al., 2010). While, in controlled artificial inoculation the viral strain is known. For example, S. lycopersiciodes accession CGN23973 showed a high level of resistance against TYLCV-IL strain when using Agrobacterium-mediated infection. However, it was symptomatic in the field with natural whitefly infection. We did not determine the virus strains present in the field in China. Possibly, CGN23973 may not be resistant to other TYLCV(-like) strains that whitefly transmit in the open field. In addition, plants are continuously exposed to large whitefly populations in the field. TYLCV incidence and severity may be influenced by the level of disease pressure. For example, the Ty-1 conferred resistance was broken under high inoculum pressure (García-Cano et al., 2008). Therefore, in the case of S. lycopersiciodes CGN23973, an alternative possible explanation for the contrasting phenotypic responses can be the high disease pressure with whitefly natural infection.
Alleles of Known TYLCV Resistance Loci
Almost all of the screened accessions of S. chilense did not support any viral symptom development which makes S. chilense the most promising resistance source. In multiple S. chilense accessions including LA1932, LA1938, and LA1971 allelic variants of the Ty-1 gene contribute to the resistance (Caro et al., 2015). Accessions LA1932 and LA1938 also carry other genetic factors (i.e., Ty-4 and Ty-6, respectively) (Ji et al., 2009; Hutton and Scott, 2014). We showed the resistance in three additional S. chilense accessions, LA0130, LA1960, and LA2737, is based on three distinct functional Ty-1/Ty-3 alleles. In the VIGS experiment, the resistance in accessions LA1961, LA2981, and LA2754 was not compromised after silencing the Ty-1 gene, which may be explained by two reasons. One is that other Ty-genes, such as Ty-4 and Ty-6 may be present in these accessions. The other is that Ty-1 was not silenced due to the insufficient effect of VIGS. In our previous studies, we have identified three aa which are specific to S. chilense Ty-1/Ty-3 alleles; as well as two aa that distinguish the Ty-1 allele in LA1969 from Ty-3 in LA2779 (Caro et al., 2015). In this study, three unique functional Ty-1/Ty-3 alleles were discovered for S. chilense LA1932, LA1938, and LA1971. Based on these results, Ty-1 and Ty-3 allele-specific markers can be further developed as in-gene markers for more precise screenings in different breeding programs. Further, as the Ty-2 and ty-5 genes are also cloned (Lapidot et al., 2015; Yamaguchi et al., 2018), gene specific SNPs can be used to check the presence of these two genes in the resistant accessions of S. habrochaites (e.g., LA1777 and LA1718) and S. peruvianum (e.g., CGN15530, CGN15532, and PI 266375). These accessions, together with the other four symptomless accessions (S. arcanum LA2172, S. huaylasense LA1364 and LA1365, S. neorickii LA0735/CGN24193, Table 1 and Supplementary Table S2), belong to the 150 Tomato Genome Re-sequenced genomes (Aflitos et al., 2014), which facilates the allele mining of the Ty-2 and ty-5 genes in these accessions.
Introgression Breeding
Introgression of resistance from wild tomato species into cultivated tomatoes is the common practice for TYLCV resistance breeding. S. chilense and S. peruvianum are the most common sources of resistance commercially used among the wild tomato species (Pérez de Castro et al., 2007). S. chilense accessions screened in this study showed very high resistance levels. Many, if not most, tested accessions remained vigorous and completely symptomless during the whole screening period (Figure 1). This makes S. chilense the first priority to search for potential novel resistance sources in the diverse tomato gene pool. Besides S. chilense, a large number of symptomless accessions have also been identified among the screened accessions of S. arcanum, S. corneliomulleri, and S. peruvianum. These species may serve as the secondary gene pool for TYLCV resistance. However, these species belong to the “peruvianum” complex, a group of species having crossing barriers with S. lycopersicum (Díez and Nuez, 2008). A new wild tomato species segregated from S. peruvianum is described and classified as S. huaylasense (Peralta et al., 2005). Only four accessions of S. huaylasense were screened for their responses upon TYLCV infection, and all these accessions were resistant with no TYLCD symptoms. Thus S. huaylasense may serve as another potential gene pool for TYLCV resistance donors. In addition to crossing barriers, introgression can be hindered by chromosomal inversions between the wild resistant donor and the cultivated tomato such as the inversion present in S. chilense (the donor species of Ty-1) (Verlaan et al., 2011) and S. habrochaites (the donor species of Ty-2) (Wolters et al., 2015). Chromosomal rearrangements lead to suppression of recombination, resulting in linkage drag. In the case of Ty-1, traits with negative effects on several agronomic traits have been introduced along with Ty-1 due to linkage drag (Rubio et al., 2010).
S. pimpinellifolium belongs to the “esculentum” complex that is completely crossable with S. cheesmaniae/S. galapagense and S. lycopersicum, the cultivated tomato (Díez and Nuez, 2008). This makes this species very suitable for breeding programs. Therefore, in this study, great efforts have been made on the screening of 408 S. pimpinellifolium accessions for sources of resistance. However, the majority of the tested accessions displayed severe levels of susceptibility. Only one S. pimpinellifolium accession G1.1554 (CGN15528) was free of TYLCD symptoms (Table 1). Several accessions showed a reduced level of susceptibility exhibiting mild to moderate TYLCD symptoms (Table 1 and Supplementary Table S2). In other studies, few accessions (Hirsute-INRA and LA1478) from S. pimpinellifolium were identified that show resistance to TYLCV with no viral symptoms (Kasrani, 1989). G1.1554 (CGN15528) is currently being exploited for TYLCV resistance (Víquez-Zamora et al., 2014). Two quantitative trait loci (QTLs) contributing to TYLCV resistance were identified on chromosomes 3 and 11 (Víquez-Zamora et al., 2014).
In summary, this is the first time that such a large scale screening has been performed to uncover TYLCV resistance (symptomless) and susceptibility (symptomatic) in wild tomato germplasm. The majority of the symptomless accessions identified in this study have never been reported before in publications. Therefore, these symptomless accessions can be considered as novel sources of TYLCV resistance. Moreover, the other merit of this study is the summary of previous efforts on screening and using wild tomato accessions in breeding for TYLCV resistance. The latter makes it clear that a very small number of TYLCV resistance sources is presently used. Finally, with few S. chilense accessions, we demonstrate how allelic variants of a cloned Ty-gene can be discovered by VIGS in combination with allele mining. Therefore, this work constitutes a treasure of knowledge for the breeder who is urged to extend the very limited germplasm used to date as a donor for TYLCV resistance in commercial cultivars.
Author Contributions
ZY, JL, A-MAW, and YB have conceived and designed the experiments. ZY, AP-d-C, MJD, and SFH have performed the experiments and analyzed the data. ZY, AP-d-C, MJD, SFH, RGFV, A-MAW, and YB have written and edited the paper.
Funding
The screening work performed at the Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences was financed by the National Key Research and Development Program of China (2016YFD0101700), the National Natural Science Foundation of China (31171975), the Agricultural Science and Technology Innovation Program (CAAS-ASTIP-IVFCAAS), the Key Laboratory of Biology and Genetic Improvement of Horticultural Crops, Ministry of Agriculture, China.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The TYLCV-IL infectious clone was kindly provided by Professor Eduardo Rodríguez Bejarano (Universidad de Málaga, Spain).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01198/full#supplementary-material
FIGURE S1 | Phenotypic responses of different wild Solanum species upon inoculation with tomato yellow leaf curl disease. Representatives of different tomato wild species are shown displaying variations in their symptom severity. From left to right are accessions corresponding to different Solanum species: S. habrochaites CGN15391, S. pennellii LA1809, S. lycopersicoides CGN23973, S. chmielewskii CGN15815, S. cheesmaniae LA1409, S. neorickii CGN15816, S. peruvianum CGN15795, S. corneliomulleri LA1944, and S. lycopersicum cv. MM. For each accession, two different individuals are shown.
FIGURE S2 | Alignment of full-length cDNA sequences of the Ty-1/Ty-3 allele in S. chilense accessions LA0130, LA2737, and LA1960. Sequences of S. lycopersicum “Moneymaker” (MM), the Ty-1 allele from S. chilense LA1969 and the Ty-3 allele from S. chilense LA2779 were described by Verlaan et al. (2013). Ty-1/Ty-3 alleles in S. chilense LA1932, LA1938, and LA1971 were obtained from Caro et al. (2015).
FIGURE S3 | Alignment of protein sequences of the Ty-1/Ty-3 alleles. Protein sequences are derived from cDNA sequences in Supplementary Figure S2. Unique amino acids present in either LA1960 or LA0130 are highlighted in yellow. The three amino acids specific for the Ty-1/Ty-3 allele are marked in red.
TABLE S1 | Summary of responses of wild tomato species upon natural infection with tomato yellow leaf curl disease at the Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing.
TABLE S2 | Average disease severity index ratings of Solanum spp. accessions upon natural infection with tomato yellow leaf curl disease at the Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing.
TABLE S3 | Wild tomato accessions displaying variation in phenotypic responses upon natural infection with tomato yellow leaf curl disease at the Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing.
TABLE S4 | Polymerase chain reaction detection of tomato yellow leaf curl virus (TYLCV) in wild accessions screened with Agrobacterium-mediated inoculation. Analyses were performed at 55 days post inoculation in young tissues of three tested individuals of each accession. Plant response was evaluated according to the intensity of viral DNA amplification and symptom expression.
TABLE S5 | Previous published data on resistant/tolerant and susceptible wild tomato accessions against tomato yellow leaf curl virus complex.
Footnotes
- ^ https://www.cabi.org/isc/datasheet/8927
- ^ https://www.cabi.org/isc/datasheet/55402
- ^ http://avrdc.org/seed/improved-lines/fresh-market-tomato/
References
Abhary, M., Patil, B. L., and Fauquet, C. M. (2007). “Molecular biodiversity, taxonomy, and nomenclature of Tomato Yellow Leaf Curl-like Viruses,” in Tomato Yellow Leaf Curl Virus Disease: Management, Molecular Biology, Breeding for Resistance, ed. H. Czosnek (Dordrecht: Springer), 85–118. doi: 10.1007/978-1-4020-4769-5_6
Aflitos, S., Schijlen, E., Jong, H., Ridder, D., Smit, S., Finkers, R., et al. (2014). Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plant J. 80, 136–148. doi: 10.1111/tpj.12616
Anbinder, I., Reuveni, M., Azari, R., Paran, I., Nahon, S., Shlomo, H., et al. (2009). Molecular dissection of tomato leaf curl virus resistance in tomato line TY172 derived from Solanum peruvianum. Theor. Appl. Genet. 119, 519–530. doi: 10.1007/s00122-009-1060-z
Andrade, M. C., da Silva, A. A., Neiva, I. P., Oliveira, I. R. C., De Castro, E. M., Francis, D. M., et al. (2017). Inheritance of type IV glandular trichome density and its association with whitefly resistance from Solanum galapagense accession LA1401. Euphytica 213:52. doi: 10.1007/s10681-016-1792-1
AVRDC (2002). “Pyramiding tomato leaf curl virus resistance genes by marker-assisted selection,” in AVRDC Report 2001, eds T. Kalb and G. Kuo (Tainan: Asian Vegetable Research and Development Center), 12–13.
AVRDC (2003). “Breeding of Solanaceous crops,” in AVRDC Report 2002, ed. T. Kalb (Tainan: Asian Vegetable Research and Development Center), 2–4.
AVRDC (2016). “ SOUTH ASIA,”in Annual Report 2015, ed. M. Mecozzi (Shanhua: AVRDC - The World Vegetable Center), 38.
AVRDC. (2001). “Resistance to yellow leaf curl in Thailand and Southeast Asia,” in AVRDC Report 2000, eds G. Kuo, D. Abbass, and T. Kalb (Tainan: Asian Vegetable Research and Development Center), 110–112.
Azizi, A., Mozafari, J., and Shams-bakhsh, M. (2008). Phenotypic and molecular screening of tomato germplasm for resistance to Tomato yellow leaf curl virus. Iran. J. Biotechnol. 6, 199–206.
Banerjee, M. K., and Kalloo, M. K. (1987). Sources and inheritance of resistance to leaf curl virus in Lycopersicon. Theor. Appl. Genet. 73, 707–710. doi: 10.1007/BF00260780
Barbieri, M., Acciarri, N., Sabatini, E., Sardo, L., Accotto, G. P., and Pecchioni, N. (2010). Introgression of resistance to two Mediterranean virus species causing tomato yellow leaf curl into a valuable traditional tomato variety. J. Plant Pathol. 92, 485–493.
Barboza, N., Blanco-Meneses, M., Hallwass, M., Moriones, E., and Inoue-Nagata, A. K. (2014). First report of Tomato yellow leaf curl virus in tomato in Costa Rica. Plant Dis. 98:699. doi: 10.1094/PDIS-08-13-0881-PDN
Belabess, Z., Dallot, S., El-Montaser, S., Granier, M., Majde, M., Tahiri, A., et al. (2015). Monitoring the dynamics of emergence of a non-canonical recombinant of Tomato yellow leaf curl virus and displacement of its parental viruses in tomato. Virology 486, 291–306. doi: 10.1016/j.virol.2015.09.011
Belabess, Z., Peterschmitt, M., Granier, M., Tahiri, A., Blenzar, A., and Urbino, C. (2016). The non-canonical tomato yellow leaf curl virus recombinant that displaced its parental viruses in southern Morocco exhibits a high selective advantage in experimental conditions. J. Gen. Virol. 97, 3433–3445. doi: 10.1099/jgv.0.000633
Bleeker, P. M., Diergaarde, P. J., Ament, K., Guerra, J., Weidner, M., Schütz, S., et al. (2009). The role of specific tomato volatiles in tomato-whitefly interaction. Plant Physiol. 151, 925–935. doi: 10.1104/pp.109.142661
Bleeker, P. M., Diergaarde, P. J., Ament, K., Schütz, S., Johne, B., Dijkink, J., et al. (2011). Tomato-produced 7-epizingiberene and R-curcumene act as repellents to whiteflies. Phytochemistry 72, 68–73. doi: 10.1016/j.phytochem.2010.10.014
Caro, M., Verlaan, M. G., Julián, O., Finkers, R., Wolters, A. M., Hutton, S. F., et al. (2015). Assessing the genetic variation of Ty-1 and Ty-3 alleles conferring resistance to tomato yellow leaf curl virus in a broad tomato germplasm. Mol. Breed. 35:132. doi: 10.1007/s11032-015-0329-y
Chinnaraja, C., Ramkissoon, A., Ramsubhag, A., and Jayaraj, J. (2016). First report of tomato yellow leaf curl virus infecting tomatoes in trinidad. Plant Dis. 100:1958. doi: 10.1094/PDIS-04-16-0446-PDN
Chomdej, O., Chatchawankanpanich, O., Kositratana, W., and Chunwongse, J. (2007). Response of resistant breeding lines of tomato germplasm and their progenies with Seedathip3 to Tomato Yellow Leaf Curl Virus, Thailand isolate (TYLCTHV-[2]). Songklanakarin J. Sci. Technol. 29, 1469–1477.
Cohen, S., and Antignus, Y. (1994). “Tomato yellow leaf curl virus, a whitefly-borne geminivirus of tomatoes,” in Advances in Disease Vector Research, ed. Kerry F. Harris (New York, NY: Springer), 259–288. doi: 10.1007/978-1-4612-2590-4_10
Czosnek, H., and Ghanim, M., (2011). “Bemisia tabaci–Tomato yellow leaf curl virus interaction causing worldwide epidemics,” in The Whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) Interaction with Geminivirus-Infected Host Plants. ed. W. M. O. Thompson (Dordrecht: Springer), 51–67. doi: 10.1007/978-94-007-1524-0_3
De Barro, P. J., Liu, S. S., Boykin, L. M., and Dinsdale, A. B. (2011). Bemisia tabaci: a statement of species status. Annu. Rev. Entomol. 56, 1–19. doi: 10.1146/annurev-ento-112408-085504
De la Peña, R., Kadirvel, P., Venkatesan, S., Kenyon, L., and Hughes, J. (2010). “Integrated approaches to manage Tomato Yellow Leaf Curl Viruses,” in Biocatalysis and Biomolecular Engineering, eds C. T. Hou and J.-F. Shaw (Hoboken NJ: John Wiley & Sons), 105–132. doi: 10.1002/9780470608524-ch8
Díaz-Pendón, J. A., Cañizares, M. C., Moriones, E., Bejarano, E. R., Czosnek, H., and Navas-Castillo, J. (2010). Tomato yellow leaf curl viruses: ménage à trois between the virus complex, the plant and the whitefly vector. Mol. Plant Pathol. 11, 441–450. doi: 10.1111/j.1364-3703.2010.00618.x
Díez, M. J., and Nuez, F. (2008). “Tomato,” in Vegetables II. Handbook of Plant Breeding. eds J. Prohens and F. Nuez (New York, NY: Springer), 249–323. doi: 10.1007/978-0-387-74110-9_7
Firdaus, S., van Heusden, A. W., Hidayati, N., Supena, E. D. J., Visser, R. G. F., and Vosman, B. (2012). Resistance to Bemisia tabaci in tomato wild relatives. Euphytica 1871, 31–45. doi: 10.1007/s10681-012-0704-2
Friedmann, M., Lapidot, M., Cohen, S., and Pilowsky, M. (1998). A novel source of resistance to Tomato yellow leaf curl virus exhibiting a symptomless reaction to viral infection. J. Am. Soc. Hortic. Sci. 123, 1004–1007.
Fulton, T. M., Chunwongse, J., and Tanksley S. D. (1995). Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep. 13, 207–209. doi: 10.1007/BF02670897
García-Cano, E., Resende, R. O., Boiteux, L. S., Giordano, L. B., Fernández-Muñoz, R., and Moriones, E. (2008). Phenotypic expression, stability, and inheritance of a recessive resistance to monopartite begomoviruses associated with Tomato yellow leaf curl disease in tomato. Phytopathology 98, 618–627. doi: 10.1094/PHYTO-98-5-0618
Gronenborn, B. (2007). “The tomato yellow leaf curl virus genome and function of its proteins,” in Tomato Yellow Leaf Curl Virus Disease: Management, Molecular Biology, Breeding for Resistance, ed. H. Czosnek (Dordrecht: Springer), 67–84. doi: 10.1007/978-1-4020-4769-5_5
Hosseinzadeh, M. R., Shams-Bakhsh, M., Osaloo, S. K., and Brown, J. K. (2014). Phylogenetic relationships, recombination analysis, and genetic variability among diverse variants of tomato yellow leaf curl virus in Iran and the Arabian Peninsula: further support for a TYLCV center of diversity. Arch. Virol. 159, 485–497. doi: 10.1007/s00705-013-1851-z
Hutton, S. F., and Scott, J. W. (2014). Ty-6, a major begomovirus resistance gene located on chromosome 10. Rept. Tomato Genet. Coop. 64, 14–18.
Hutton, S. F., and Scott, J. W. (2017). Fla. 7907C: a Fla. 7907 near-isogenic tomato inbred line containing the begomovirus resistance gene, Ty-1. HortScience 52, 658–660. doi: 10.21273/HORTSCI11827-17
Hutton, S. F., Ji, Y., and Scott, J. W. (2015). Fla. 8923: a tomato breeding line with begomovirus resistance gene Ty-3 in a 70-kb Solanum chilense introgression. HortScience 50, 1257–1259.
Ji, Y., Schuster, D. J., and Scott, J. W. (2007a). Ty-3, a begomovirus resistance locus near the Tomato yellow leaf curl virus resistance locus Ty-1 on chromosome 6 of tomato. Mol. Breed. 20, 271–284. doi: 10.1007/s11032-007-9089-7
Ji, Y., Scott, J. W., Hanson, P., Graham, E., and Maxwell, D. P. (2007b). “Sources of resistance, inheritance, and location of genetic loci conferring resistance to members of the tomato-infecting begomoviruses,” in Tomato Yellow Leaf Curl Virus Disease: Management, Molecular Biology, Breeding for Resistance, ed. H. Czosnek (Dordrecht: Springer), 343–362. doi: 10.1007/978-1-4020-4769-5_20
Ji, Y., Scott, J. W., Schuster, D. J., and Maxwell, D. P. (2009). Molecular mapping of Ty-4, a new Tomato yellow leaf curl virus resistance locus on chromosome 3 of tomato. J. Am. Soc. Hortic. Sci. 134, 281–288.
Jordá, C., Picó, B., Díez, M. J., and Nuez, F. (1996). “Cribado de germoplasma resistente a TYLCV. Desarrollo de un método de diagnóstico adecuado,” in Proceedings of the VIII Congreso Nacional de la Sociedad Española de Fitopatología. Córdoba, 218.
Julián, O., Herráiz, J., Corella, S., di-Lolli, I., Soler, S., Díez, M. J., et al. (2013). Initial development of a set of introgression lines from Solanum peruvianum PI 126944 into tomato: exploitation of resistance to viruses. Euphytica 193, 183–196. doi: 10.1007/s10681-013-0896-0
Kalloo, M. K., and Banerjee, M. K. (1990). Transfer of Tomato leaf curl virus resistance from Lycopersicon hirsutum f. glabratum to L. esculentum. Plant Breed. 105, 156–159. doi: 10.1111/j.1439-0523.1990.tb00469.x
Kasrani, M. A. (1989). Inheritance of resistance to Tomato yellow leaf curl virus (TYLCV) in Lycopersicon pimpinellifolium. Plant Dis. 73, 435–437. doi: 10.1094/PD-73-0435
Kennedy, G. G. (2003). Tomato, pests, parasitoids, and predators: tritrophic interactions involving the genus Lycopersicon. Annu. Rev. Entomol. 48: 51–72. doi: 10.1146/annurev.ento.48.091801.112733
Lapidot, M., Karniel, U., Gelbart, D., Fogel, D., Evenor, D., Kutsher, Y., et al. (2015). A novel route controlling begomovirus resistance by the messenger RNA surveillance factor pelota. PLoS Genet. 11:e1005538. doi: 10.1371/journal.pgen.1005538
Lefeuvre, P., Martin, D. P., Harkins, G., Lemey, P., Gray, A. J., Meredith, S., et al. (2010). The spread of tomato yellow leaf curl virus from the Middle East to the world. PLoS Pathog. 6:e1001164. doi: 10.1371/journal.ppat.1001164
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Mabvakure, B., Martin, D. P., Kraberger, S., Cloete, L., van Brunschot, S., Geering, A. D. W., et al. (2016). Ongoing geographical spread of Tomato yellow leaf curl virus. Virology 498, 257–264. doi: 10.1016/j.virol.2016.08.033
Maruthi, M. N., Czosnek, H., Vidavski, F., Tarba, S.-Y., Milo, J., Leviatov, S., et al. (2003). Comparison of resistance to Tomato leaf curl virus (India) and Tomato yellow leaf curl virus (Israel) among Lycopersicon wild species, breeding lines and hybrids. Eur. J. Plant Pathol. 109, 1–11. doi: 10.1023/A:1022091905911
Momotaz, A., Scott, J. W., and Schuster, D. J. (2010). Identification of quantitative trait loci conferring resistance to Bemisia tabaci in an F2 population of Solanum lycopersicum × Solanum habrochaites accession LA1777. J. Am. Soc. Hortic. Sci. 135, 134–142.
Moriones, E., and Navas-Castillo, J. (2000). Tomato yellow leaf curl virus, an emerging virus complex causing epidemics worldwide. Virus Res. 71, 123–134. doi: 10.1016/S0168-1702(00)00193-3
Moriones, E., Arno, J., Accotto, G. P., Noris, E., and Cavallarin, L. (1993). First report of tomato yellow leaf curl virus in Spain. Plant Dis.77:953. doi: 10.1094/PD-77-0953B
Moyle, L. C. (2008). Ecological and evolutionary genomics in the wild tomatoes (Solanum sect. Lycopersicon). Evolution 62, 2995–3013. doi: 10.1111/j.1558-5646.2008.00487.x
Nawaz-ul-Rehman, M. S., and Fauquet, C. M. (2009). Evolution of geminiviruses and their satellites. FEBS Lett. 583, 1825–1832. doi: 10.1016/j.febslet.2009.05.045
Ning, W., Shi, X., Liu, B., Pan, H., Wei, W., Zeng, Y., et al. (2015). Transmission of Tomato yellow leaf curl virus by Bemisia tabaci as affected by whitefly sex and biotype. Sci. Rep. 5:10744. doi: 10.1038/srep10744
Ohnishi, J., Yamaguchi, H., and Saito, A. (2016). Analysis of the Mild strain of tomato yellow leaf curl virus, which overcomes Ty-2. Arch.Virol. 161, 2207–2217. doi: 10.1007/s00705-016-2898-4
Peralta, I. E., Knapp, S., and Spooner, D. M. (2005). New species of wild tomatoes (Solanum section Lycopersicon: Solanaceae) from Northern Peru. Syst. Bot. 30, 424-434. doi: 10.1600/0363644054223657
Peralta, I. E., and Spooner, D. M. (2005). “Morphological characterization and relationships of wild tomatoes (Solanum L. Sect. Lycopersicon),” in Monographs In Systematic Botany, eds R. C. Keating, V. Hollowell, and T. B. Croat (St. Louis, MO: Missouri Botanical Garden), 227–257.
Peralta, I. E., Spooner, D. M., and Knapp, S. (2008). Taxonomy of wild tomatoes and their relatives (Solanum sect. Lycopersicoidesxs, sect. Juglandifolia, sect. Lycopersicon; Solanaceae). Syst. Bot. Monogr. 84, 1–186.
Pereira-Carvalho, R. C., Boiteux, L. S., Fonseca, M. E. N., Díaz-Pendón, J. A., Moriones, E., Fernández-Muñoz, R., et al. (2010). Multiple resistance to Meloidogyne spp. and bipartite and monopartite Begomovirus spp. in wild Solanum (Lycopersicon) accessions. Plant Dis. 94, 179–185. doi: 10.1094/PDIS-94-2-0179
Pérez de Castro, A., Díez, M. J., and Nuez, F. (2004). Identificación de nuevas fuentes de resistencia al virus del rizado amarillo del tomate (TYLCV). Actas de Horticult. 41, 119–122.
Pérez de Castro, A., Díez, M. J., and Nuez, F. (2005). Caracterización de entradas de Lycopersicon peruvianum y L. chilense por su resistencia al Tomato yellow leaf curl virus (TYLCV). Actas Portuguesas Horticult. 8, 48–54.
Pérez de Castro, A., Díez, M. J., and Nuez, F. (2007). Inheritance of Tomato yellow leaf curl virus resistance derived from Solanum pimpinellifolium UPV16991. Plant Dis. 91, 879–885. doi: 10.1094/PDIS-91-7-0879
Pérez de Castro, A., Díez, M. J., and Nuez, F. (2008). Exploiting partial resistance to Tomato yellow leaf curl virus derived from Solanum pimpinellifolium UPV16991. Plant Dis. 92, 1083–1090. doi: 10.1094/PDIS-92-7-1083
Pérez de Castro, A., Díez, M. J., and Nuez, F. (2010). Resistencia a la enfermedad del rizado amarillo del tomate en la especie silvestre Solanum lycopersicoides. Actas de Horticul. 55, 169–170.
Pérez de Castro, A., Julián, O., and Díez, M. J. (2013). Genetic control and mapping of Solanum chilense LA1932, LA1960 and LA1971-derived resistance to Tomato yellow leaf curl disease. Euphytica 190, 203–214. doi: 10.1007/s10681-012-0792-z
Picó, B., and Díez, M. J. (1998). “Screening Lycopersicon spp. for resistance to TYLCV,” in Second International Workshop on Bemisia and Geminiviral Disease, USDA-ARS, San Juan, 43.
Picó, B., Díez, M. J., and Nuez, F. (1998). Evaluation of whitefly-mediated inoculation techniques to screen Lycopersicon esculentum and wild relatives for resistance to Tomato yellow leaf curl virus. Euphytica 101, 259–271. doi: 10.1023/A:1018353806051
Picó, B., Díez, M. J., and Nuez, F. (1999a). Improved diagnostic techniques for tomato yellow leaf curl virus in tomato breeding programs. Plant Dis. 83, 1006–1012. doi: 10.1094/PDIS.1999.83.11.1006
Picó, B., Ferriol, M., Diez, M. J., and Nuez, F. (1999c). Developing tomato breeding lines resistant to tomato yellow leaf curl virus. Plant Breed. 118, 537–542. doi: 10.1046/j.1439-0523.1999.00427.x
Picó, B., Ferriol, M., Diez, M., and Nuez, F. (1999b). Cribado de fuentes de resistencia de Lycopersicon spp. al Tomato Yellow Leaf Curl Virus mediante agroinoculación en disco de hoja. Actas Horticult. 24, 105–112.
Picó, B., Herraiz, J., Ruiz, J., and Nuez, F. (2002). Widening the genetic basis of virus resistance in tomato. Sci. Hortic. 94, 73–89. doi: 10.1016/S0304-4238(01)00376-4
Picó, B., Sifres, A., Elía, M., Díez, M. J., and Nuez, F. (2000). Searching for new resistance sources to tomato yellow leaf curl virus within a highly variable wild Lycopersicon genetic pool. Acta Physiol. Plant. 22, 344–350. doi: 10.1007/s11738-000-0051-0
Pilowsky, M., and Cohen, S. (1990). Tolerance to tomato yellow leaf curl virus derived from Lycopersicon peruvianum. Plant Dis. 74, 248–250. doi: 10.1094/PD-74-0248
Powell, M. E., Cuthbertson, A. G., Bell, H. A., Boonham, N., Morris, J., and Northing, P. (2012). First record of the Q biotype of the sweetpotato whitefly, Bemisia tabaci, intercepted in the UK. Eur. J. Plant Pathol. 133, 797–801. doi: 10.1007/s10658-012-9976-6
Rakha, M., Hanson, P., and Ramasamy, S. (2017). Identification of resistance to Bemisia tabaci Genn. in closely related wild relatives of cultivated tomato based on trichome type analysis and choice and no-choice assays. Genet. Resour. Crop Evol. 64, 247–260. doi: 10.1007/s10722-015-0347-y
Rodríguez-López, M., Garzo, E., Bonani, J., Fereres, A., Fernández-Muñoz, R., and Moriones, E. (2011). Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the preference and feeding behavior of Bemisia tabaci and reduce the spread of Tomato yellow leaf curl virus. Phytopathology 101, 1191–1201. doi: 10.1094/PHYTO-01-11-0028
Rubio, F., García-Martínez, S., Alonso, A., Grau, A., Valero, M., and Ruiz, J. J. (2010). “Introgressing resistance genes into traditional tomato cultivars: effects on yield and quality,” in Proceedings XXVIIIth International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on New Developments in Plant Genetics and Breeding. ed. J.M. Leitao (Lisbon: ISHS Acta Horticulturae), 29–33. doi: 10.17660/ActaHortic.2012.935.3
Scott, J. W., Hutton, S. F., and Freeman, J. H. (2015). Fla. 8638B and Fla. 8624 tomato breeding lines with begomovirus resistance genes ty-5 plus Ty-6 and Ty-6, respectively. HortScience 50, 1405–1407.
Soler, S., Pico, B., Sifres, A., Diez, M., De Frutos, R., and Nuez, F. (2000). “Multiple virus resistance in a collection of Lycopersicon spp.,” in Proceedings of the Fifth Congress of the European Foundation for Plant Pathology (Taormina, Italy: European Foundation for Plant Pathology), 17.
Tomás, D. M., Cañizares, M. C., Abad, J., Fernández-Muñoz, R., and Moriones, E. (2011). Resistance to Tomato yellow leaf curl virus accumulation in the tomato wild relative Solanum habrochaites associated with the C4 viral protein. Mol. Plant Microbe Interact. 24, 849–861. doi: 10.1094/MPMI-12-10-0291
Verlaan, M. G., Hutton, S. F., Ibrahem, R. M., Kormelink, R., Visser, R. G. F., Scott, J. W., et al. (2013). The tomato yellow leaf curl virus resistance genes Ty-1 and Ty-3 are allelic and code for DFDGD-class RNA-dependent RNA polymerases. PLoS Genet. 9:e1003399. doi: 10.1371/journal.pgen.1003399
Verlaan, M. G., Szinay, D., Hutton, S. F., de Jong, H., Kormelink, R., Visser, R. G. F., et al. (2011). Chromosomal rearrangements between tomato and Solanum chilense hamper mapping and breeding of the TYLCV resistance gene Ty-1. Plant J. 68, 1093–1103. doi: 10.1111/j.1365-313X.2011.04762.x
Vidavski, F., Czosnek, H., Gazit, S., Levy, D., and Lapidot, M. (2008). Pyramiding of genes conferring resistance to Tomato yellow leaf curl virus from different wild tomato species. Plant Breed. 127, 625–631. doi: 10.1111/j.1439-0523.2008.01556.x
Vidavsky, F., and Czosnek, H. (1998). Tomato breeding lines resistant and tolerant to tomato yellow leaf curl virus issued from Lycopersicon hirsutum. Phytopathology 88, 910–914. doi: 10.1094/PHYTO.1998.88.9.910
Vidavsky, F., Leviatov, S., Milo, J., Rabinowitch, H., Kedar, N., and Czosnek, H. (1998). Response of tolerant breeding lines of tomato, Lycopersicon esculentum, originating from three different sources (L. peruvianum, L. pimpinellifolium and L. chilense) to early controlled inoculation by tomato yellow leaf curl virus (TYLCV). Plant Breed. 117, 165–169. doi: 10.1111/j.1439-0523.1998.tb01472.x
Víquez-Zamora, M., Caro, M., Finkers, R., Tikunov, Y., Bovy, A., Visser, R. G. F., et al. (2014). Mapping in the era of sequencing: high density genotyping and its application for mapping TYLCV resistance in Solanum pimpinellifolium. BMC Genomics 15:1152. doi: 10.1186/1471-2164-15-1152
Wolters, A. M. A., Caro, M., Dong, S., Finkers, R., Gao, J., Visser, R. G. F., et al. (2015). Detection of an inversion in the Ty-2 region between S. lycopersicum and S. habrochaites by a combination of de novo genome assembly and BAC cloning. Theor. Appl. Genet. 128, 1987–1997. doi: 10.1007/s00122-015-2561-6
Yamaguchi, H., Ohnishi, J., Saito, A., Ohyama, A., Nunome, T., Miyatake, K., et al. (2018). An NB-LRR gene, TYNBS1, is responsible for resistance mediated by the Ty-2 Begomovirus resistance locus of tomato. Theor. Appl. Genet. 131, 1345–1362. doi: 10.1007/s00122-018-3082-x
Yang, X., Caro, M., Hutton, S. F., Scott, J. W., Guo, Y., Wang, X., et al. (2014). Fine mapping of the tomato yellow leaf curl virus resistance gene Ty-2 on chromosome 11 of tomato. Mol. Breed. 34, 749–760. doi: 10.1007/s11032-014-0072-9
Keywords: Begomovirus, resistance, Solanum lycopersicum, S. chilense, S. peruvianum, tomato, TYLCV
Citation: Yan Z, Pérez-de-Castro A, Díez MJ, Hutton SF, Visser RGF, Wolters A-MA, Bai Y and Li J (2018) Resistance to Tomato Yellow Leaf Curl Virus in Tomato Germplasm. Front. Plant Sci. 9:1198. doi: 10.3389/fpls.2018.01198
Received: 11 May 2018; Accepted: 26 July 2018;
Published: 20 August 2018.
Edited by:
Sergio Lanteri, Università degli Studi di Torino, ItalyReviewed by:
Rafael Fernández-Muñoz, Consejo Superior de Investigaciones Científicas (CSIC), SpainGian Paolo Accotto, Istituto per la Protezione Sostenibile delle Piante (IPSP), Italy
Copyright © 2018 Yan, Pérez-de-Castro, Díez, Hutton, Visser, Wolters, Bai and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuling Bai, YmFpLnl1bGluZ0B3dXIubmw= Junming Li, bGlqdW5taW5nQGNhYXMuY24=