 Katarzyna Roguz1*
Katarzyna Roguz1* Andrzej Bajguz2
Andrzej Bajguz2 Agnieszka Gołębiewska2
Agnieszka Gołębiewska2 Magdalena Chmur2
Magdalena Chmur2 Laurence Hill3
Laurence Hill3 Paweł Kalinowski4Jürg Schönenberger5
Paweł Kalinowski4Jürg Schönenberger5 Małgorzata Stpiczyńska1
Małgorzata Stpiczyńska1 Marcin Zych1*
Marcin Zych1*- 1Botanic Garden, Faculty of Biology, University of Warsaw, Warsaw, Poland
- 2Department of Plant Biochemistry and Toxicology, Faculty of Biology and Chemistry, Institute of Biology, University of Bialystok, Bialystok, Poland
- 3Petersham Lodge, Richmond, United Kingdom
- 4Department of Nature Protection and Rural Landscape, Institute of Technology and Life Sciences, Falenty, Poland
- 5Department of Botany and Biodiversity Research, University of Vienna, Vienna, Austria
Fritillaria is a genus consisting of 130 to 140 species of bulbous plants, native to temperate regions of the northern hemisphere. Generally viewed as an insect pollinated genus with the exception of two North American species, Fritillaria gentneri and F. recurva, which are described as hummingbird-pollinated and the Asian species, F. imperialis, described as passerine-pollinated. These pollinator shifts are possibly the result of adaptive changes to the structure and morphology of the nectary, as well as a change in the nectar concentration and composition. A study was conducted in a target group of 56 Fritillaria species, based on the morphology of their nectaries and nectar composition to assess the significance of pollination mode as well as its predisposition for the evolution of bird pollination. All species studied had nectaries located at their tepal base and produced nectar, but their size, shape, color, and composition all varied. Most fritillaries had hexose-rich nectar, in easily accessible and unprotected nectaries. Scanning electron microscope (SEM) analysis revealed that the surface of the nectaries of most Fritillaria species was flat and clearly distinct from that of the surrounding tissues, which might be regarded as an adaptation for insect-pollination. Nectaries of F. imperialis were considerably larger and had dilute nectar without sucrose, which was produced profusely, thereby fulfilling the criteria characteristic of ornithophilous flowers. The copious nectar of presumed hummingbird-pollinated species was rather balanced and of medium sugar concentration. Their large lanceolate nectaries contrasted sharply with the tessellated background of their tepals. These characters might indicate a mixed pollination system that engages both birds and insects. Floral anatomy and microstructure and nectar composition for Fritillaria species in subgenera Korolkowia and Liliorhiza are studied for the first time.
Introduction
The genus Fritillaria L. (type species F. meleagris L.) is a widely distributed member of Liliaceae (lily family). It comprises of 130 to 140 species occurring through most of the northern hemisphere (Tamura, 1998; Rønsted et al., 2005; Tomović et al., 2007; Mabberley, 2008; Day et al., 2014), with centers of speciation in the Mediterranean region, especially in Turkey and Greece (Rix, 1984; Zaharof, 1986; Tekşen and Aytaç, 2011), Iran (Kiani et al., 2017), Western North America (Beetle, 1944; Rønsted et al., 2005), and East Asia (Hill, 2016). Recent phylogenetic analyses indicate that the genus may be paraphyletic with members of the subgenus Liliorhiza, principally North American species, forming a sister clade to the remaining Fritillaria species and the genus Lilium (Day et al., 2014).
Fritillaria are found in a variety of climatic regions and in different habitats, with about half of the genus occurring in locations categorized as open with dry summers (Rønsted et al., 2005). Fritillaries are located across a wide latitudinal range from coasts, riparian zones, meadows, woodland, steppe, deserts, mountain screes, and alpine zones (Xinqi and Mordak, 2000; Ness, 2003; Tekşen and Aytaç, 2011; Zych and Stpiczyńska, 2012; Rix and Strange, 2014).
Fritillaria species are bulbiferous, spring-flowering perennials with an erect flowering stem producing either a single flower or multi-flowered racemes. The flowers are usually actinomorphic and have a typical tulip-like, trimerous, campanulate perianth but with a nodding character (Tamura, 1998). They come in various colors such as white, pink, greenish, yellow, or purplish/reddish. The perianth parts of many species, including the type species, have a characteristic checkerboard pattern; hence, the name of the genus – fritillus, Latin for dicebox (Zych et al., 2014).
This astonishing floral diversity may have developed in response to their pollinator interactions, although, flower visitors have only been recorded for six species of Fritillaria (White, 1789; Hedström, 1983; Búrquez, 1989; Peters et al., 1995; Bernhardt, 1999; Minagi et al., 2005; Kawano et al., 2008; Zox and Gold, 2008; Zych and Stpiczyńska, 2012). These limited records are unlikely to reflect the complete spectrum of pollination vectors. Pollinator effectiveness has only been evaluated for bumblebee-pollinated F. meleagris (Zych and Stpiczyńska, 2012; Zych et al., 2013, 2014) and to a lesser extent the pollination of F. imperialis by birds and insects (Peters et al., 1995).
The first observation of a Fritillaria flower visitor was made by White (1789). He recorded small birds drinking nectar from F. imperialis. After 200 years, similar observations were also made for F. imperialis in other European gardens by Burques (1989) and Peters et al. (1995). Blue and great tits visited and efficiently pollinated flowers. Bumblebees were also seen visiting and pollinating the large and pendant flowers of F. imperialis, but pollinator efficiency was lower than bird visitations (Peters et al., 1995). Although there are no pollinator records for F. imperialis in the wild, it has been proposed that bird pollination is the most likely vector. This hypothesis is supported by the presence of a landing platform suitable for passerine birds and large pendant flowers with large volumes of diluted nectar (Peters et al., 1995). For some North American species (F. gentneri and F. recurva), hummingbird pollination has been recorded in natural habitats (Pendergrass and Robinson, 2005; Cronk and Ojeda, 2008), possibly attracted by their bright red flowers. The only Fritillaria species occurring in both Asia and North America, the typically dark-flowered F. camschatcensis, is habitually visited by flies (Zox and Gold, 2008). Flowers of this species emit a disagreeable smell like rotting flesh, and it might be described as a typical example of carrion fly-pollination syndrome (Willmer, 2011). There are only six fritillaries that have been noted to have a sweet scent: F. liliacea (King, 2001), F. obliqua (Beck, 1953), F. striata (Santana, 1984), F. stribrnyi (personal observation LH), F. tortifolia (personal observation LH), and F. yuminensis (Leon et al., 2009), all presumably visited by bee species. Bees have only been recorded to visit F. ayakoana (Minagi et al., 2005) and F. meleagris (Hedström, 1983; Stpiczyńska et al., 2012; Zych et al., 2014).
Generally, flower pollinators are attracted by a combination of visual and olfactory cues. Although the data is limited, one might assume this is true for Fritillaria. Floral scent, but to the best of our knowledge, it has only been explored in studies for F. meleagris (Hedström, 1983). In the study conducted by Zych and Stpiczyńska (2012), many bees collected Fritillaria pollen, but the most common floral food-reward appeared to be nectar. This was secreted by perigonal nectaries positioned adaxially on each of the six perianth segments (Stpiczyńska et al., 2012). Until now, the most comprehensive description of Fritillaria nectar diversity was given by Rix and Rast (1975), who studied nectar sugars in 37 European and Asiatic species of the genus. These authors found that nectar generally contained all three common nectar sugars, namely sucrose, glucose, and fructose. The only exception in their dataset was F. imperialis which produced no sucrose, confirming an earlier report by Wykes (1952). Rix and Rast (1975) concluded that nectar composition and, in particular, the fructose/glucose ratio may be an important diagnostic character in the infrageneric taxonomy of Fritillaria.
The position, morphology, and structure of nectar-secreting glands have been key taxonomic characters and have been investigated by many researchers, notably in Asiatic species by Bakhshi Khaniki and Persson (1997) and for F. meleagris by Stpiczyńska et al. (2012). For example, nectaries are lanceolate to linear in members of the subgenus Fritillaria, circular in the subgenus Petilium (L.) Baker, and in the subgenus Rhinopetalum (Fisch. ex Alexand) the nectaries are deeply depressed and situated in sac-like projections (Bakhshi Khaniki and Persson, 1997). This latter study, however, provides only general morphological descriptions and information concerning the ultrastructure of nectaries, is only available for F. meleagris (Stpiczyńska et al., 2012; Zych and Stpiczyńska, 2012). Studies of nectaries and nectar characteristics are important, not only from a taxonomic point of view but also with respect to ecological and evolutionary studies of the genus. For example, based on nectar characteristics, Rix and Rast (1975) posited that bees and wasps were the main pollinators of Fritillaria. In fact, floral visitors of Fritillaria flowers include insects of the orders Hymenoptera (mostly various species of bees and wasps), Diptera, Lepidoptera, and Coleoptera (Hedström, 1983; Bernhardt, 1999; Naruhashi et al., 2006; Kawano et al., 2008; Zych and Stpiczyńska, 2012; Zych et al., 2013). Although usually concealed by the perianth, in some Fritillaria species optically copious nectaries also play an important ecological role in guiding the pollinators to the nectar once they have been attracted to the flowers by other traits (tepal color, scent, etc.). This appears to be the case in ornithophilous F. imperialis (Cronk and Ojeda, 2008) and, perhaps, also in other bird-pollinated species. However, despite the diversity and wide distribution of the genus, these aspects of Fritillaria diversity are still neglected to a great extent. In particular, detailed studies of the flower, and, especially the study of nectar-secreting structures and nectar composition have only been conducted for a very limited number of species. To date, the most complete analysis using light microscopy (LM) and scanning electron microscopy (SEM) was completed by Bakhshi Khaniki and Persson (1997) for members of four subgenera represented in Central Asia: Fritillaria, Petilium (L.) Baker, Rhinopetalum (Fisch. ex Alexand) Baker, and Theresia (K. Koch) Baker. However, no information is available for fritillaries from North America or the Far East. This paper represents the first study of floral anatomy and microstructure, as well as nectar composition, for a broad range of Fritillaria species, including, for the first time, taxa from two subgenera: Korolkowia Rix and Liliorhiza (Kellogg) Benth. & Hook.f. Our objectives were (1) to verify the presence of secretory tissues in selected members of the genus, (2) to investigate the microstructure of their nectaries, and (3) to compare nectar production and composition in the taxa studied in order to shed light on the evolution of their pollination systems.
Materials and Methods
Plant material used for this study was obtained from Fritillaria species cultivated at the University of Warsaw Botanic Garden (BG), from the private collections of the coauthors (LH and PK) and from the private collection of Colin Everett (Somerton, Somerset, United Kingdom; CE). Many Fritillaria species are very rare in cultivation, and the number of specimens used for each type of analysis varied because of the availability of fresh plant material (accession numbers for species in this investigation are listed in Table 1).
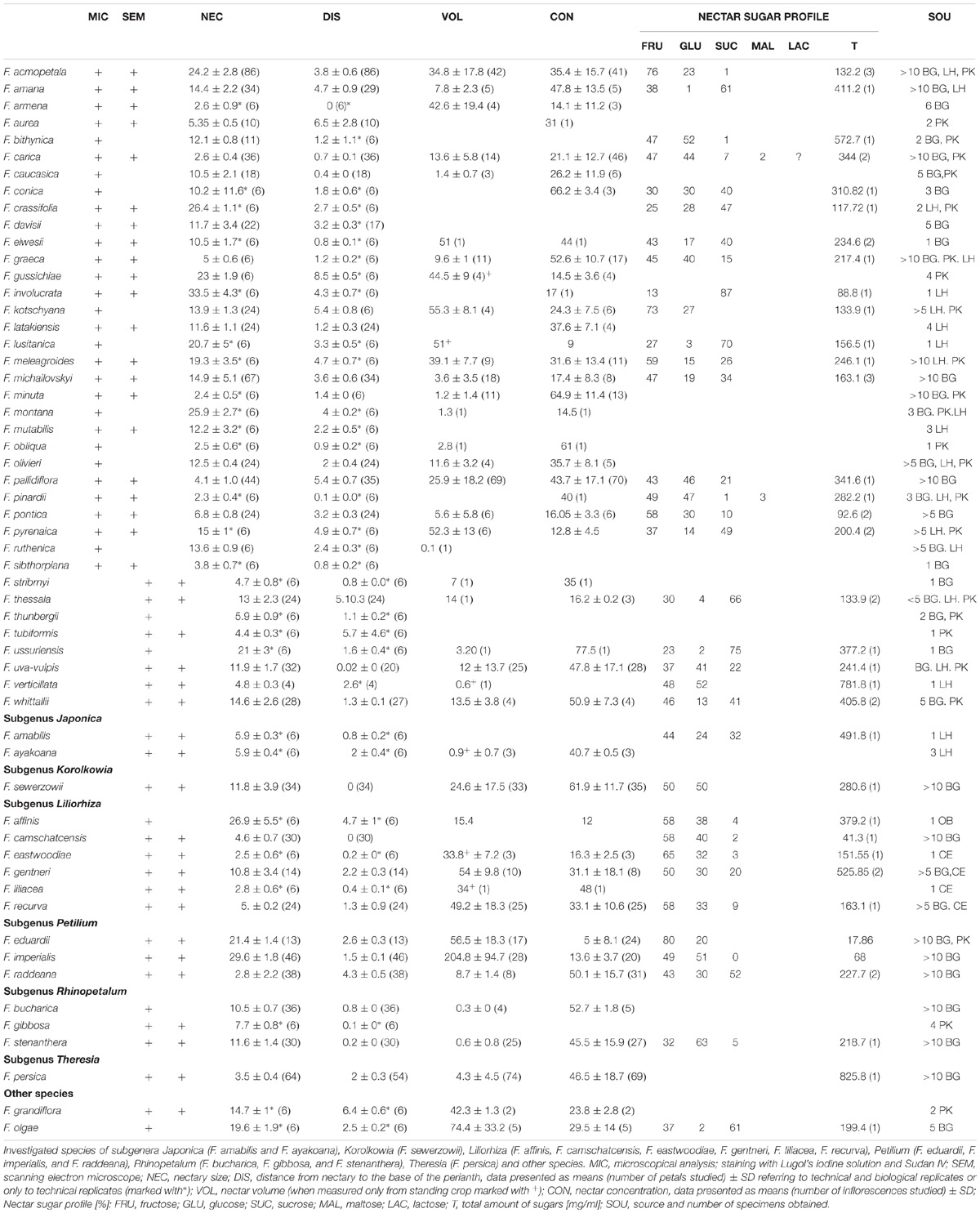
TABLE 1. Investigated species of the subgenus Fritillaria.
Microscopical Observations
All microscopical examinations were conducted for flowers in full anthesis. Flowers from 1 to 10 were selected for morphometric measurements of the nectary size and position. If less than three specimens were available, all flowers from one individual were measured, and if more plants were available, the flowers studied were selected randomly. The study was conducted with the use of a digital caliper Borletti DIN 862 (Borletti, Italy), tethered to a computer to record the values. Shape, size, structure, and color of the nectaries were observed in the fresh material using a Nikon SMZ 1000 stereomicroscope (Nikon Corp., Japan).
SEM Observations
Three areas were selected for SEM observations on the outer tepals: the nectaries, the area distal to the nectaries, and the tip of the tepals. Sections of nectaries collected in the greenhouse and from PK collection were prepared by fixing nectary tissue in 2.5% glutaraldehyde in phosphate buffer (pH 7.4; 0.1 M). After three washes in phosphate buffer, they were postfixed in 2% (w/v) osmium tetroxide solution for 2 h and were dehydrated in a graded ethanol series. After dehydration, samples were subjected to critical point drying using liquid CO2 and were sputter-coated with gold. Nectaries gathered from the collections of LH and CE were transported from the United Kingdom to Warsaw in 70% ethanol. Subsequently, the material was prepared for SEM as described above, and the sample was examined using a SEM LEO 1430VP (Zeiss, Germany) and Zeiss Libra 120 (Zeiss, Germany).
Seven representative species (F. eduardii, F. gentneri, F. michailovskyi, F. persica, F. recurva, F. raddeana, and F. sewerzowii), either characterized by visually different nectary structures or representing closely related species, were prepared as semi-thin nectary sections.
Plant material was prepared by fixing nectary tissue in 2.5% glutaraldehyde in phosphate buffer (pH 7.4; 0.1 M). The samples were then washed three times before postfixation in 2% (w/v) osmium tetroxide solution for 2 h and were dehydrated in a graded ethanol series. After dehydration, they were infiltrated with LR White resin. Succeeding polymerization at 60°C, the nectaries sections were cut with a glass knife. The semi-thin sections (0.9–1.0 μm thick), stained with an aqueous solution of 1% methylene blue/1% Azure II (1:1) for 5–7 min on a hot plate (60°C), were prepared for LM and analyzed for general histology.
Hand-cut sections of the nectaries of all studied species were also prepared for histochemical investigations by means of LM. The size of epidermal and parenchymal cells and the depth of nectariferous tissue were measured. Subsequently, hand-cut sections were stained with an alcoholic solution of Sudan IV for lipids and with Lugol’s iodine solution for starch. Sections of nectaries were also stained with aniline blue and were examined by means of fluorescence microscopy (FM) in order to test for the presence of callose in cell walls.
Nectar Sampling
Flowers for nectar sampling in the collections of BG and PK were first selected during the bud stage and were bagged with nylon mesh (net 0.5 mm) to prevent visits by insects. During anthesis but before anther dehiscence, nectar was sampled. In the BG collection flowers, progress was checked daily in the morning and in the afternoon for the presence of nectar. Nectar sampling in the LH and CE collections was from unbagged flowers open to animal visitors. All nectar was sampled with microcapillary pipettes from nectaries of all six tepals and was combined as one sample per flower. In the case of F. camschatcensis, nectar volumes were very small and sampling with microcapillaries pipettes was performed under a Nikon SMZ 1000 stereomicroscope (Nikon Corp., Japan). The collected nectar was subsequently expelled from microcapillaries onto a refractometer prism RL-4 (PZO, Poland) in order to measure nectar sugar concentration.
Nectar was also sampled to assess nectar sugar composition. Nectar from a standing crop of unbagged flowers was collected for this purpose. Particular care was taken during nectar collection, to avoid any contamination by pollen, phloem soap, or any other plant tissue. However, as most of the Fritillaria have downward facing flowers the risk of pollen contamination was low.
No attempts were made to emasculate flowers prior to sampling, so that sugar composition represents nectar as encountered by visitors. Nectar from one to three flowers of each species was placed into 1.5 ml Eppendorf tubes prior to analysis using high performance liquid chromatography (HPLC). The samples were frozen (−20°C) until required. Nectar was diluted with water to a volume of 50 μl (10 μl of nectar + 40 μl of water). The sample was filtered through spin columns using a 0.4 μm pore size membrane filter before injection. The supernatant was then loaded into the insert. An Agilent 1260 Infinity Series HPLC system with autoinjector, refrigerated autosampler compartment, thermostatted column compartment, quaternary pump with in-line vacuum degasser, and refractive index detector was used. A ZORBAX Carbohydrate Analysis Column (4.6 mm × 250 mm, 5 μm) was used for sugar separation and analysis. A 10 μl aliquot sample or standard solution was injected. The separation was conducted at 30°C with the mobile phase comprising acetonitrile:water (70:30, v/v) at a flow rate of 1.4 ml/min. The analytical data were integrated using the Agilent OpenLab CDS ChemStation software for liquid chromatography (LC) systems. Identification of sugars was performed by comparing retention times of individual sugars in the reference vs. test solution. The content of glucose, fructose, sucrose, maltose, and lactose was assayed based on comparisons of peak areas obtained for the samples investigated with those of the reference solutions.
Results
Nectary Location and Structure, Nectar Secretion, Concentration, and Composition
In all species, six nectaries were located at the base of the tepals (Figures 1, 2). The mean distance from the base of the perianth for all species studied was 2.3 ± 2.1 mm (means calculated only for technical replicates, if only one specimen was available, or means resulting from technical replicates were used to represent each biological replicate; missing SD values represent a single accession), in the range of 0.0 to 8.5 mm (Table 1). In all but one studied species, the nectaries of both outer and inner tepals were equally accessible to potential pollinators. Only in F. persica were the nectaries of the outer tepals not visible.
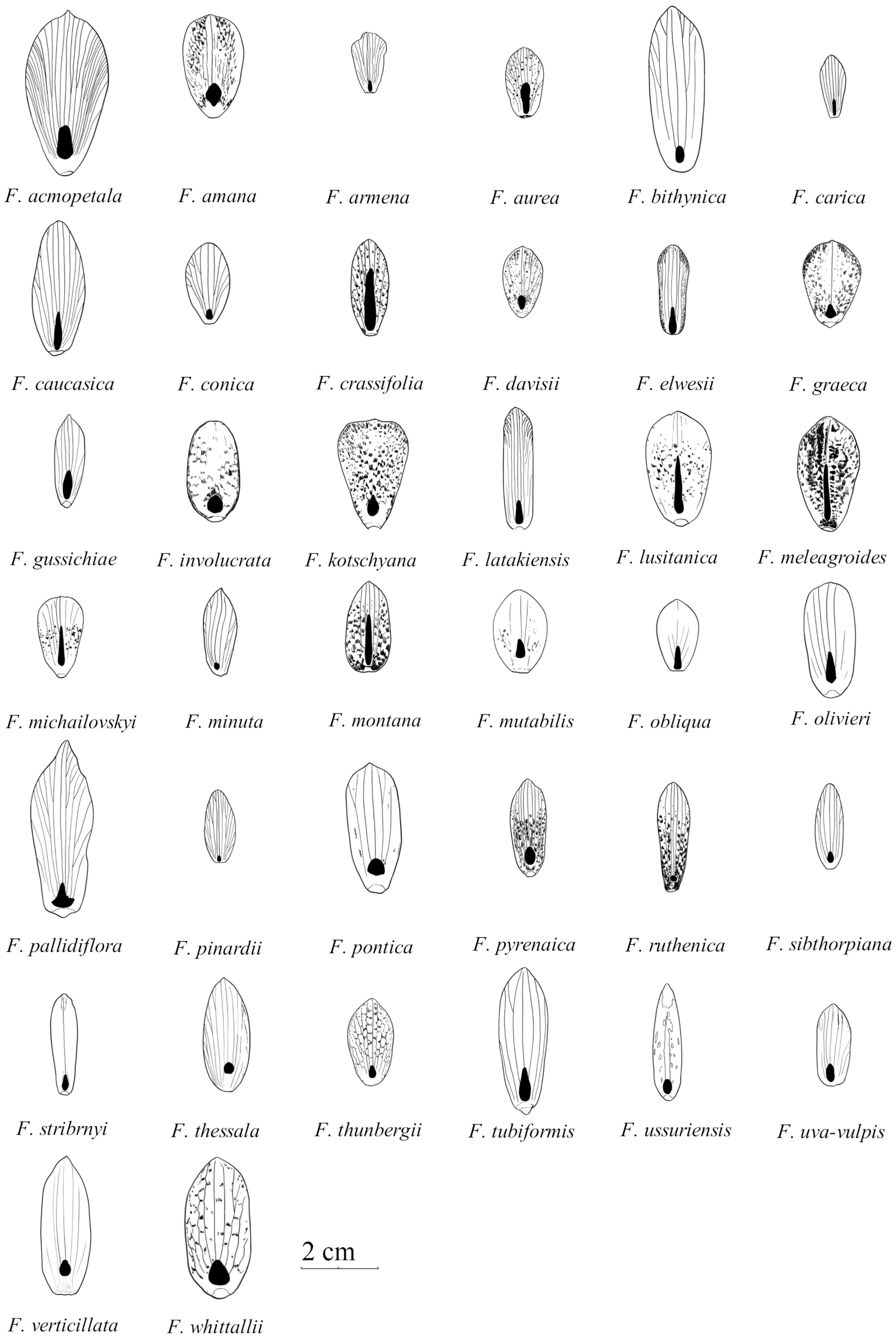
FIGURE 1. Schematic drawings of outer tepals and nectaries (filled with black) in species studied of the subgenus Fritillaria. Size graded according to natural size of studied tepals (Drawn by Jan Kryciński).
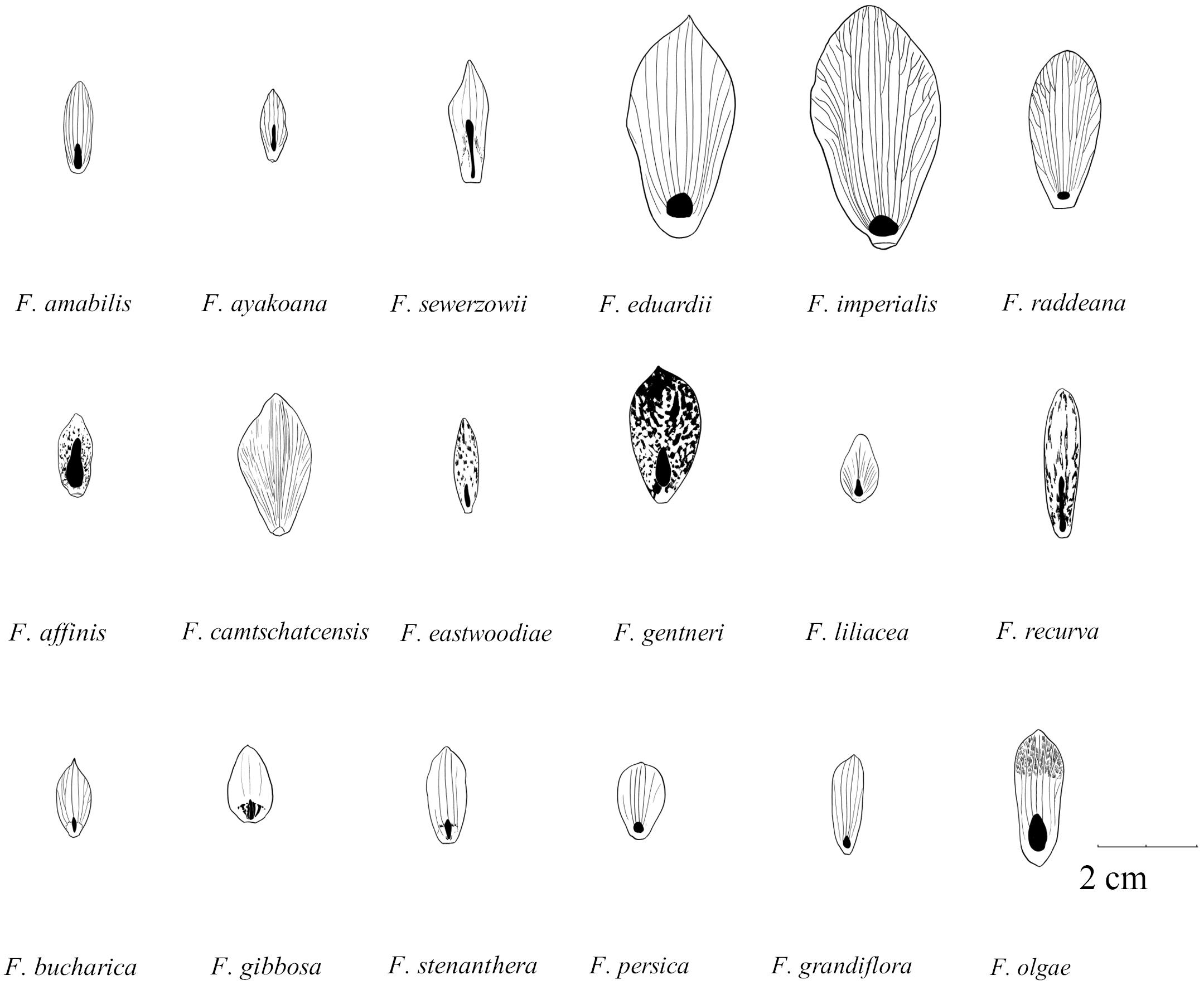
FIGURE 2. Schematic drawings of outer tepals and nectaries (filled with black) in selected species of subgenera: Japonica (F. amabilis and F. ayakoana); Petilium (F. eduardii, F. imperialis, and F. raddeana); Liliorhiza (F. affinis, F. camschatcensis, F. eastwoodiae, F. gentneri, F. liliacea, and F. recurva); Rhinopetalum (F. bucharica, F. gibbosa, and F. stenanthera); Korolkowia (F. sewerzowii); Theresia (F. persica); and other species (F. olgae and F. tubiformis) (Drown by Jan Kryciński).
Nectary cells were smaller, flatter, and more regular in shape than other epidermal cells (not shown). In each case, the nectaries consisted of a single-layered epidermis (without stomata) and several layers of subepidermal parenchyma (Figures 3D,E)1. The cytoplasm of epidermal cells contained a large nucleus, small vacuoles, and plastids. Plastids were also present in deeper layers of the nectaries’ parenchyma. Vascular bundles contained both xylem and phloem elements. Subepidermal nectary parenchyma consisted of 2–5 layers (Figure 3F). Staining with Lugol’s iodine solution revealed no starch grains, with the exception of members of the subgenus Petilium, where staining revealed the presence of numerous starch grains in the plastids of epidermal and subepidermal cells (Figure 4B).
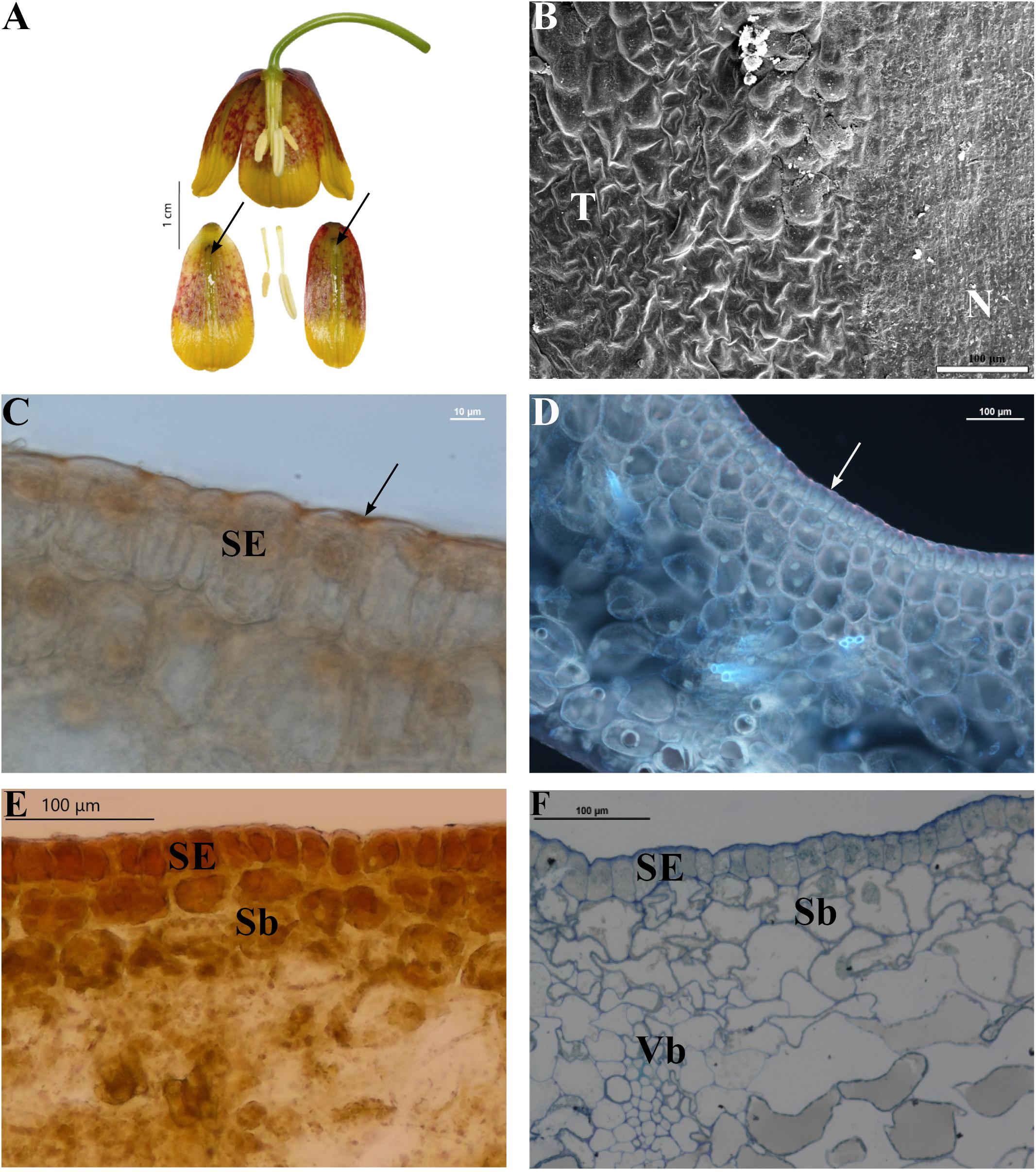
FIGURE 3. Flowers and nectaries of F. michailovskyi at full anthesis. Macro, SEM, and LM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Part of outer tepal showing flat nectary cells (N) and slightly convex cells of surrounding tepals area (T). (C) Cuticule on the surface of secretory epidermis (SE) of outer tepals stained with Sudan IV, arrow indicates lipids layer on the surface of cuticule of nectary. (D) Staining with aniline blue does not reveal the presence of callose in the walls of nectary cells. Arrow indicates lipids on the cuticle of the nectary of outer tepals. (E) Treatment with Lugol’s iodine solution reveals the absence of starch in secretory (SE) and subsecretory parenchyma (Sb). Secretory epidermis with dense cytoplasm. (F) Staining with Azure II showing secretory epidermis (SE), subsecretory parenchyma (Sb), and ground parenchyma with vascular bundle (Vb).
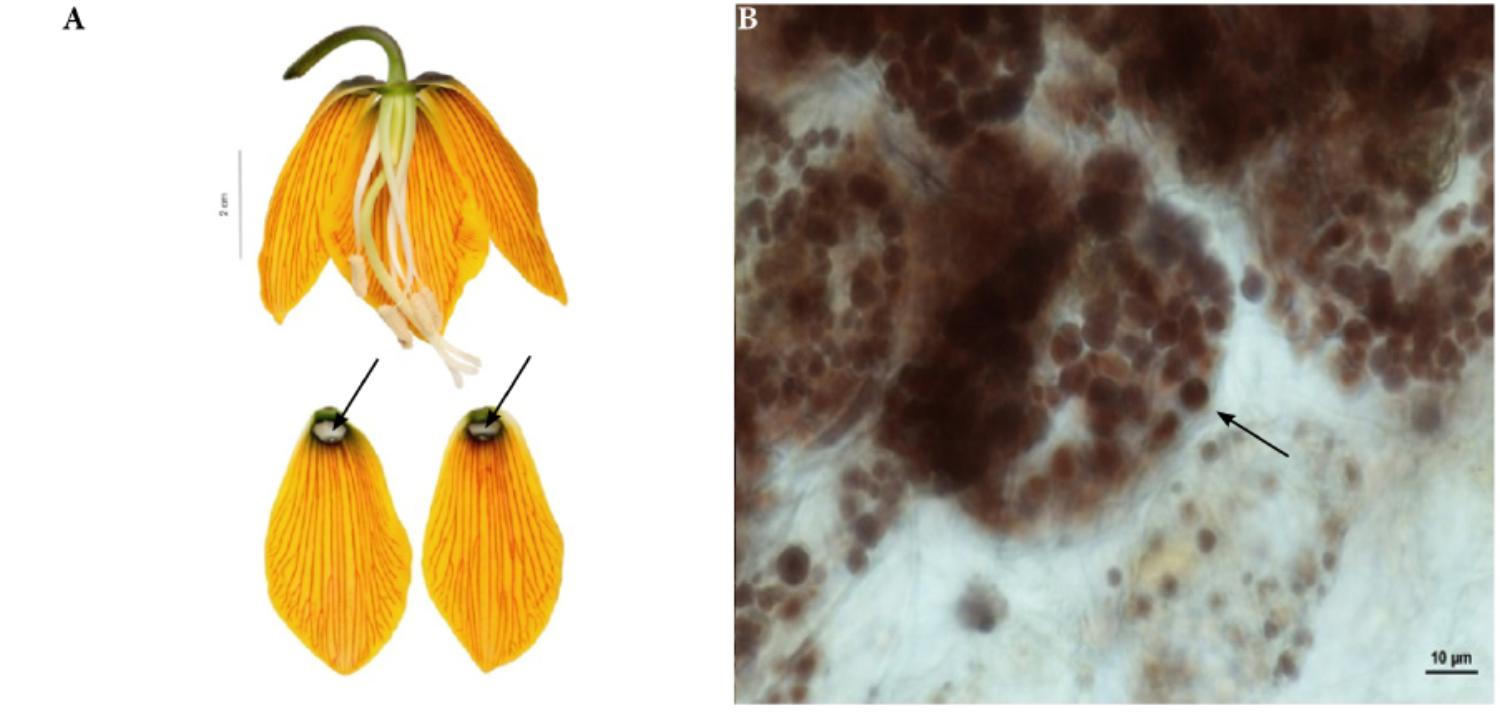
FIGURE 4. Flowers and nectaries of F. imperialis at full anthesis. Macro and LM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Treatment with Lugol’s iodine solution reveals the presence of starch in secretory and subsecretory parenchyma. Grains visible in the plastids of subepidermal cells in F. imperialis outer tepal nectaries.
Staining with Sudan IV revealed the presence of numerous droplets of lipid on the epidermis (Figure 3C) and within nectary cells of all studied species, the cuticule on the surface of secretory epidermis stained red. Staining with aniline blue did not reveal the presence of callose in cell walls (Figure 3D). The mean area occupied by the nectaries of all studied species was 11.8 ± 8.6 mm2, in the range of 1 to 38.2 mm2 (Table 1).
In all of the investigated species, each of the six nectaries located adaxially on perianth segments produced nectar. Nectar passed across the cell wall and was exuded through pores in the cuticle.
The amount of nectar produced depended largely based on the species. On average, Fritillaria flowers produced 30.6 ± 52.2 μl of nectar, in the range of 0.4 to 204.8 μl (means and SDs; N = 498, 41 species; data pooled for all seasons and species investigated). The concentration of nectar, on average, was 39.1 ± 20.63%, in the range of 5 to 77.5% (N = 599, 44 species). Nectar in most species was hexose-rich (with mean total concentration of 278.6 ± 187.8 mg/ml). The sugar profile of nectar was dominated by sucrose and glucose, which were also detected in the nectar of all species (115.4 ± 77.2 mg/ml and 88.5 ± 122.2 mg/ml, respectively; N = 54, 34 species). Fructose was also a significant component of Fritillaria nectar (87.6 ± 84.4 mg/ml), but it was not present in the nectar of all species studied. Traces of maltose and lactose were also detected in the nectar of several species (4 ± 1.8 and 5.1 mg/ml, respectively) (Table 1 and Supplementary Material).
Subgenus Fritillaria
This subgenus was represented by 38 species (Table 1). The nectaries of this subgenus were highly variable, and the nectaries differed greatly in size and area occupied (mean value 12.4 ± 8.8 mm2). The average distance of the nectaries from the base of the perianth was 2.7 ± 2.2 mm. Several species had ovate nectaries of uniform background color (i.e., of poor contrast) and were difficult to differentiate. For example, F. amana had round nectaries, sometimes encircled by a brownish band, especially those of the inner tepals, but the nectaries were generally of a uniform green color. Fritillaria aurea, F. bithynica, F. conica, and F. sibthorpiana had round, slightly depressed, yellow or slightly greenish nectaries, similar in color to the rest of the tepal. Fritillaria davisii and F. pyrenaica (Figure 5A) had linear-lanceolate darker nectaries that did not contrast well against the tessellated brownish background. Fritillaria elwesii had ovate to triangular, greenish nectaries, of a slightly darker hue when compared to that of surrounding tissues. Fritillaria pallidiflora had triangular nectaries, greenish to yellowish and of the same color as the tepals.
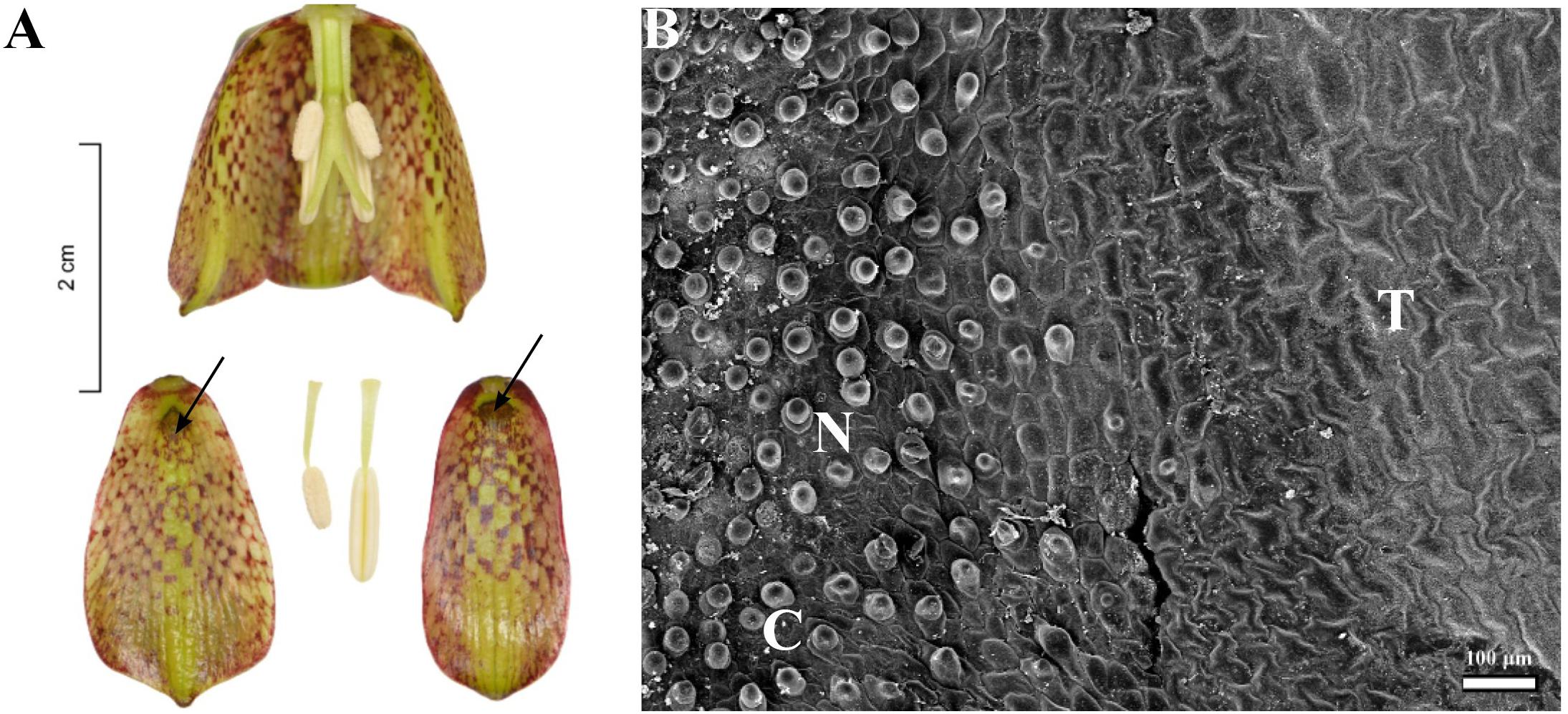
FIGURE 5. Flowers and nectaries of F. pyrenaica. Macro and SEM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Part of outer tepal showing the nectary area (N), comprising conical papillae and slightly convex cells (C) of surrounding tepals area (T).
Fritillaria uva-vulpis had ovate yellowish nectaries and a similarly colored background.
Considerably, more members of the subgenus Fritillaria had contrasting nectaries. Fritillaria acmopetala, F. graeca, F. involucrata, F. latakiensis, F. mutabilis, F. olivieri, F. pontica, F. thessala, and F. verticillata had ovate to obovate, dark nectaries that contrasted strongly against the brighter tepals. Fritillaria kotschyana and F. whittallii also had ovate to obovate, dark nectaries that contrasted strongly against the tessellated tepals. Fritillaria minuta had ovate, slightly depressed nectaries of dark green color and contrasting yellowish tepals. Similarly, F. pinardii and F. carica, had linear-lanceolate, deeply depressed, greenish nectaries, that were only slightly darker than the green-yellow tepals. Sometimes, there was slightly more contrast when the tepals were yellow. Fritillaria lusitanica, F. ussuriensis, F. ruthenica, and F. michailovskyi (Figure 3A) had greenish, oblanceolate nectaries surrounded by reddish, tessellated tepals. Fritillaria thunbergii had greenish, oblanceolate nectaries surrounded by yellowish, tessellated tepals. Fritillaria caucasica, F. obliqua, and F stribrnyi had linear-lanceolate, bright green nectaries contrasting with dark red tepals, like those of F. montana, where the green, slightly depressed nectaries were surrounded by red, tessellated tepals. Fritillaria gussichiae had ovate, bright green nectaries that contrasted strongly against a dark red background. Fritillaria crassifolia had linear nectaries that were usually green and heavily marked with purple. Nectaries were visible but did not contrast strongly against the green, red-tessellated tepals. Similarly, in F. meleagroides, the dark, linear nectaries were surrounded by green tepals with dark red tessellation. Fritillaria armena had nectaries at the base of the perianth. Several species had nectaries close to the base of the perianth (not more than 1 mm distant) or more than 5 mm from it, but for most of the species nectaries arose more than 1 mm but less than 5 mm from the base of the perianth (Table 1).
Scanning electron microscope analysis revealed that in most of the species investigated, the internal surface of the nectary was flat, while the surrounding area and the rest of the tepal was slightly undulate, owing to the slightly convex cells (Figure 3B). Nectaries of F. verticillata also had slightly convex cells. In F. armena, the area of the nectary was also surrounded by a row of elevated, rounded protrusions, also present on the tepals, where they were arranged in rows. In F. davisii, the remainder of the tepal was covered with elevated protrusions. Fritillaria uva-vulpis and F. michailovskyi had rows of elevated cells directly above the nectaries. In F. pyrenaica (Figure 5B), the area of the nectary was comprised of conical papillae. In F. tubiformis, nectary cells had papillae and the epidermal cells of the surrounding area were also slightly convex.
Plants of this subgenus produced variable amounts of nectar (32.3 ± 54.4 μl) of highly variable concentration (38.5 ± 20.6%.) The lowest mean concentration was recorded for F. pyrenaica (12.8 ± 4.5%, 52.3 ± 13 μl). The highest mean value was observed for F. ussuriensis (77.5%, 3.2 μl). The highest production was recorded for F. kotschyana (55.3 ± 8.1 μl, 24.3 ± 7.5%). The smallest volume was recorded for F. verticillata (0.6 μl, but this was too small to measure sugar concentration) (Table 1).
Subgenus Japonica
This subgenus was represented by two species (Table 1). Nectaries of these species were yellowish and ovate-lanceolate. In the case of F. ayakoana (Figure 6A), the base of the nectary was green and it contrasted strongly with the bright tepals. In the middle of the nectary, there were small upwardly curved ridges or protuberances. The area occupied by nectaries of the members of this subgenus varied little and, on average, measured 5.4 ± 1 mm2. The nectaries were placed close to the base of the perianth (Table 1).
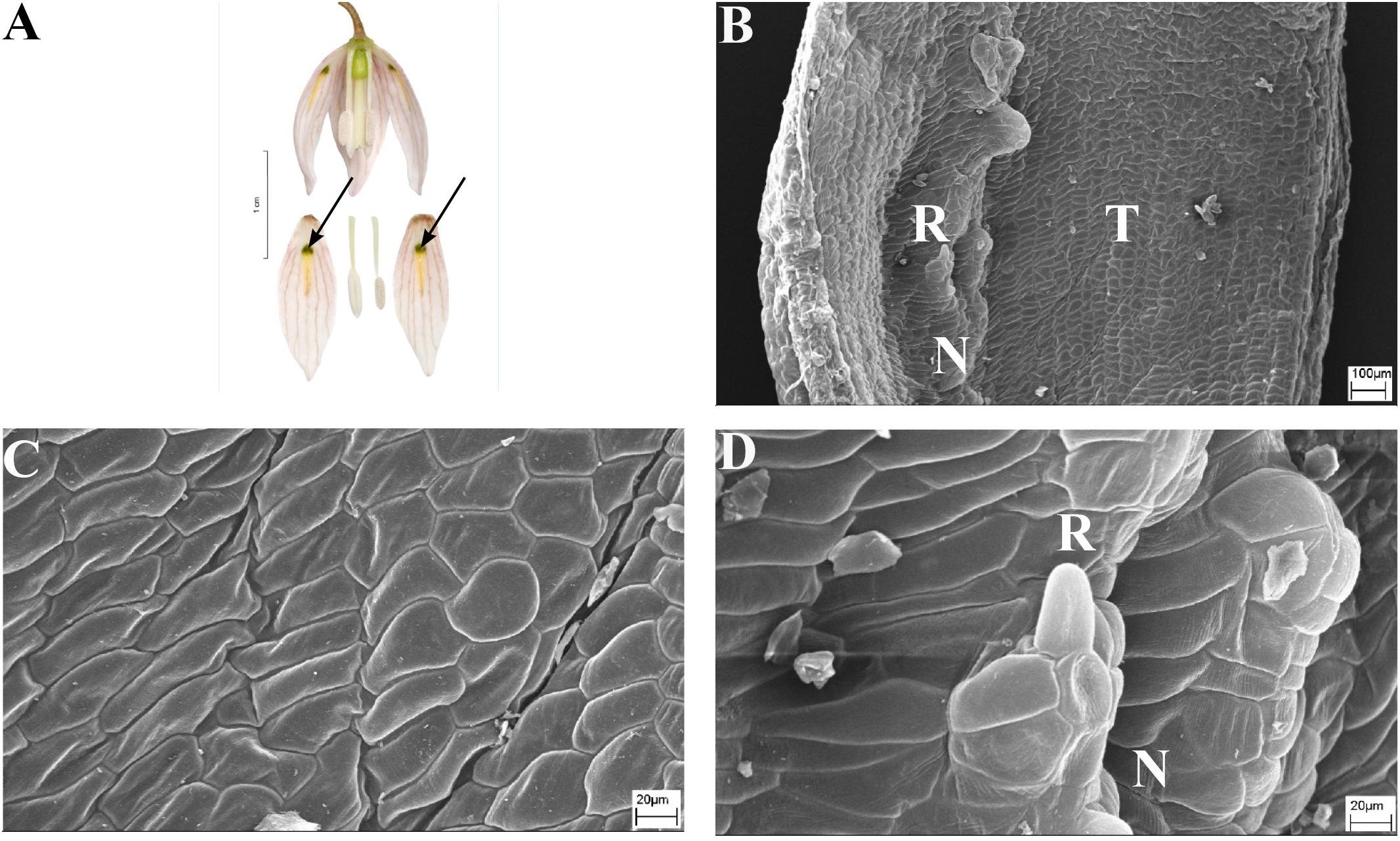
FIGURE 6. Flowers and nectaries of F. ayakoana. Macro and SEM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Part of outer tepal showing the nectar-bearing area (N) with upwardly curved ridge or protuberance on its surface and slightly convex surrounding area (T). (C) Slightly convex cells of outer tepal surface directly above the nectary. (D) Cuticule of the nectary (N) of outer tepal with protuberance (R).
Scanning electron microscope analysis revealed that the surface of the nectaries and the surrounding areas of F. ayakoana (Figures 6B,C) flowers were identical with conical projections (Figures 6B,D).
The layer of subepidermal nectary parenchyma was deeper in F. ayakoana and was four or more cells deep, whereas in F. amabilis, it was 2–3 cells deep.
During anthesis, the entire nectary area was coated with nectar (no data regarding nectar replenishing available). Flowers of the species studied produced, on average, 0.9 ± 0.7 μl nectar of concentration 40.7 ± 0.5% (Table 1).
Subgenus Korolkowia
This is a monotypic subgenus containing F. sewerzowii. Its nectaries were long, elliptical, and depressed in a groove (Figure 7A), which was surrounded by a row of longitudinal papillose ridges (Figure 7B). They did not cover the nectaries, which were clearly visible, green, and were strongly contrasting with the dark background. The area of the nectaries was measured to be 11.8 ± 3.9 mm2, and the nectaries were located at the base of the perianth (Table 1).
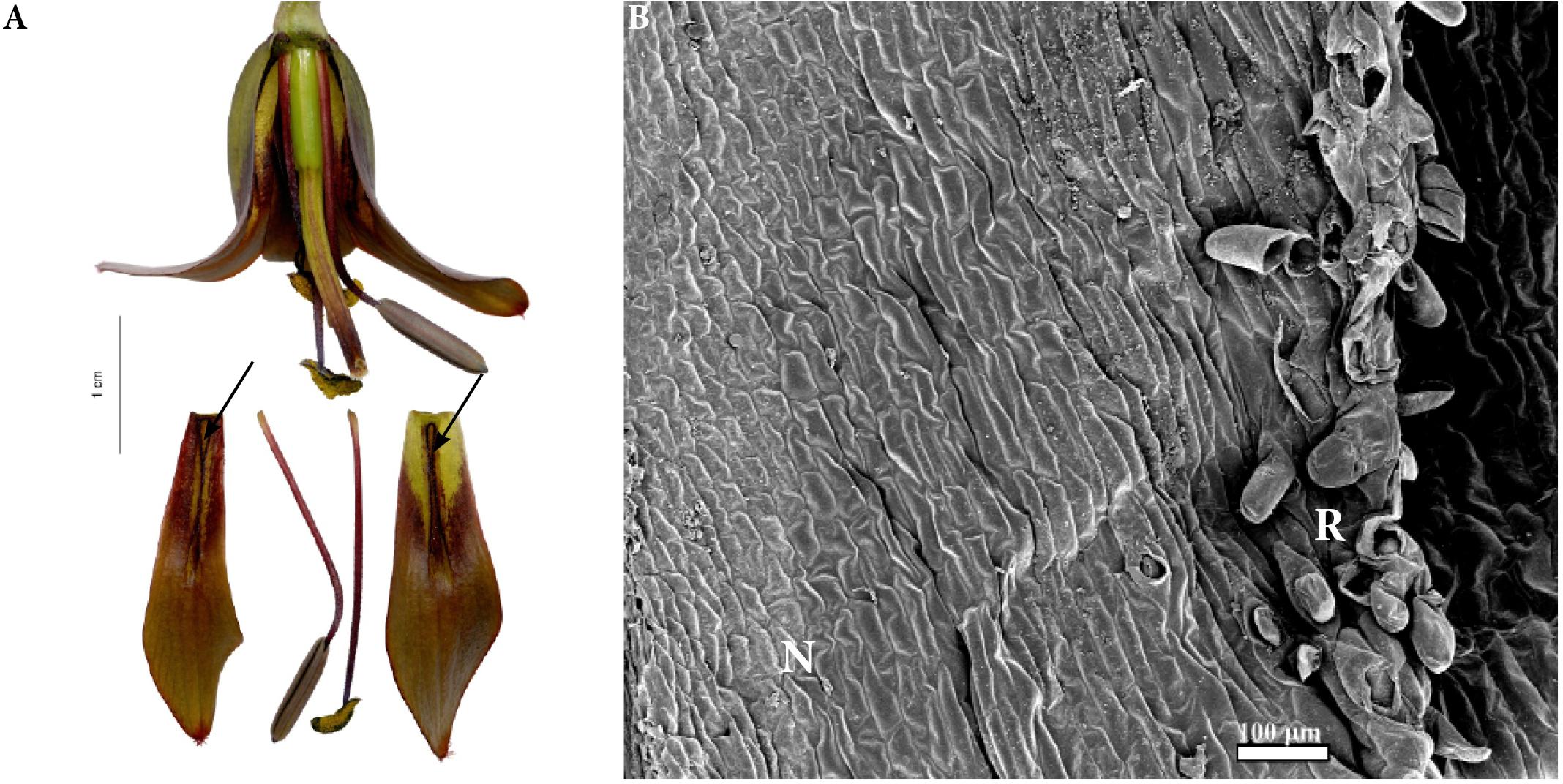
FIGURE 7. Flowers and nectaries of F. sewerzowii. Macro and SEM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Part of outer tepal showing the nectary (N), surrounded by papillose ridges (R).
Scanning electron microscope analysis revealed that the surface of the nectaries was slightly undulate and wrinkled. The cells of the surrounding area were slightly convex (Figure 7B).
During anthesis, the entire nectary was coated with nectar, which was replenished on its removal. Flowers of this species produced, on average, 24.6 ± 17.5 μl nectar of concentration 61.9 ± 11.7% (Table 1).
Subgenus Liliorhiza
This subgenus was represented by six species (Table 1). Nectaries of F. camschatcensis were very narrow, lanceolate and were hidden in the ridges (Figure 8C). Its surface was covered with proturbance and it glistened; therefore, the nectaries always looked as if they contained nectar. The nectaries of other Liliorhiza species were ovate-lanceolate in shape and were not protected by any additional structures. Nectaries of F. affinis, F. recurva, F. gentneri, and F. eastwoodiae were similar in appearance and were brightly colored against a contrasting darker, tessellated background (Figure 9A). The nectaries of F. liliacea (Figure 8A), like the surrounding part of the tepal, were uniformly green and, thus, almost invisible. Fritillaria affinis had the largest nectaries (29.6 ± 5.5 mm2), and the smallest nectaries were recorded for F. liliacea (2.8 ± 0.6 mm2). The nectaries were generally situated close to the base of the perianth (0.9 ± 0.9 mm) (Figure 9A and Table 1).

FIGURE 8. Flowers and nectaries of F. liliacea (A,B) and of F. camschatcensis (C,D), both in full anthesis. (A) Flowers and tepals of F. liliacea, nectaries marked with arrows. (B) Protrusions surrounding the nectary area (P) on F. liliacea outer tepal (T). (C) Flowers and tepals of F. camtschatcensis, nectaries marked with arrows. (D) Protrusions surrounding the nectary area on the outer tepal of F. camschatcensis.
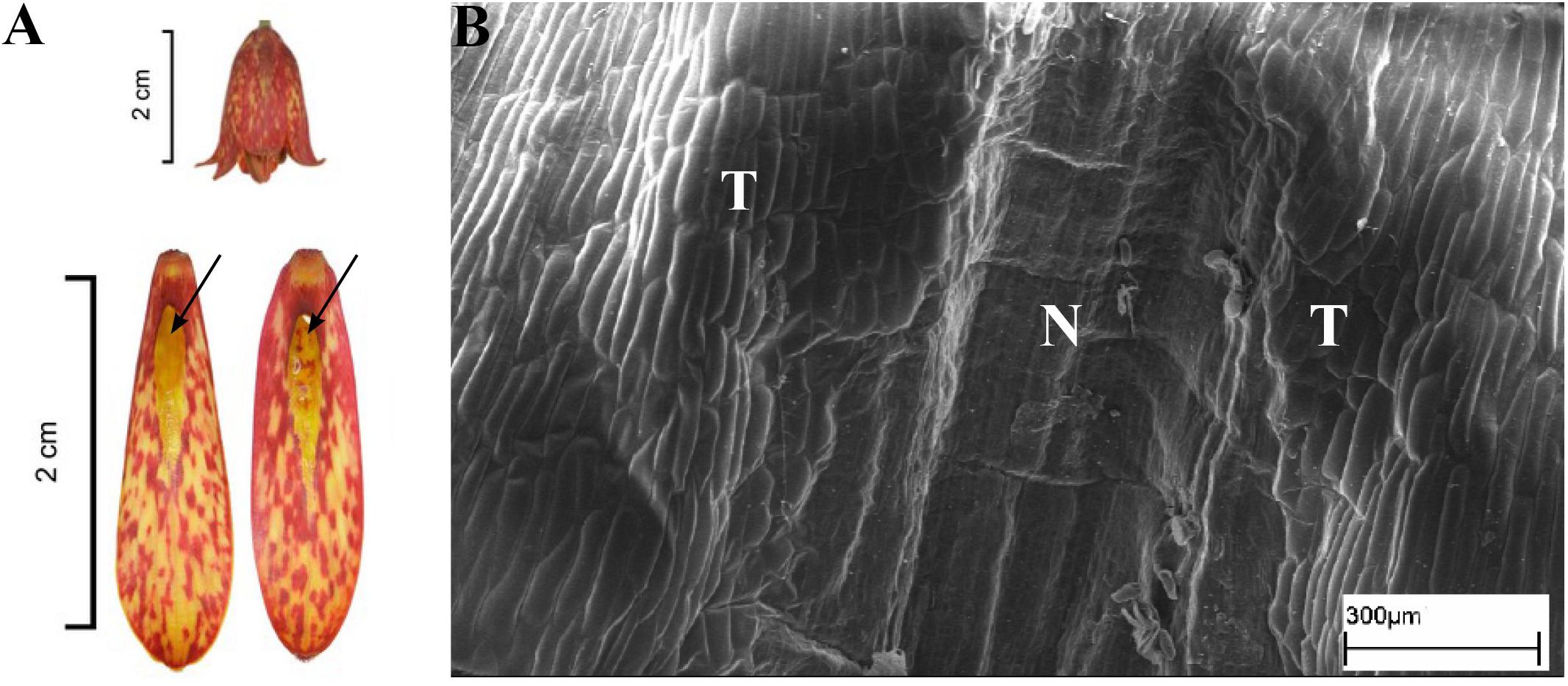
FIGURE 9. Flowers and nectaries of F. gentneri. Macro and SEM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Part of outer tepal showing deeply depressed nectary (N).
Scanning electron microscope analysis revealed that the surface of the depressed nectaries of F. eastwoodiae consisted of slightly convex epidermal cells, as well as in the surrounding area. Fritillaria liliacea and F. camschatcensis also had depressed nectaries surrounded by a row of elevated cells having a grooved surface (Figures 8B,D). Both F. gentneri and F. recurva had depressed nectaries surrounded by elevated cells (Figure 9B).
The subepidermal nectary parenchyma consisted of four or more layers. Only in F. eastwoodiae was the nectary parenchyma shallower and consisted of 2–3 layers.
In F. gentneri and F. recurva, nectar was replenished on its removal. Fritillaria camschatcensis produced very small amount of barely visible, viscous nectar. Owing to the consistency of the nectar and the fact that the nectary surface was glistening, it was not possible to assess nectar replenishment. No data was available for F. eastwoodiae and F. liliacea. Plants of this subgenus produced copious amounts of nectar (48 ± 17.1 μl) of average concentration 30.1 ± 11.9%. The lowest concentration of nectar was recorded for flowers of F. affinis (12%), and the highest was recorded for F. liliacea (48%). The smallest volume of nectar was produced by F. affinis (15.4 μl), and the greatest volume of nectar was produced by F. gentneri (54 ± 9.8 μl) (Table 1).
Subgenus Petilium
This subgenus was represented by three species (Table 1). The nectaries were depressed, elliptic, or round in shape. In F. imperialis and F. eduardii, they were similar in size with an area of 27.7 ± 4.7 mm2 and were similarly located 1.7 ± 0.7 mm above the base of the perianth. Nectaries of F. raddeana (Figure 10A) were smaller (2.8 ± 2.2 mm2) and were located 4.3 ± 0.5 mm above the base of the tepal (Table 1). The white nectaries of F. imperialis (Figure 4A) and F. eduardii contrasted sharply with the surrounding dark green background. Nectaries of F. raddeana were not as strongly contrasting as were those of the two previous species described; they were darker and surrounded by a similarly dark background.
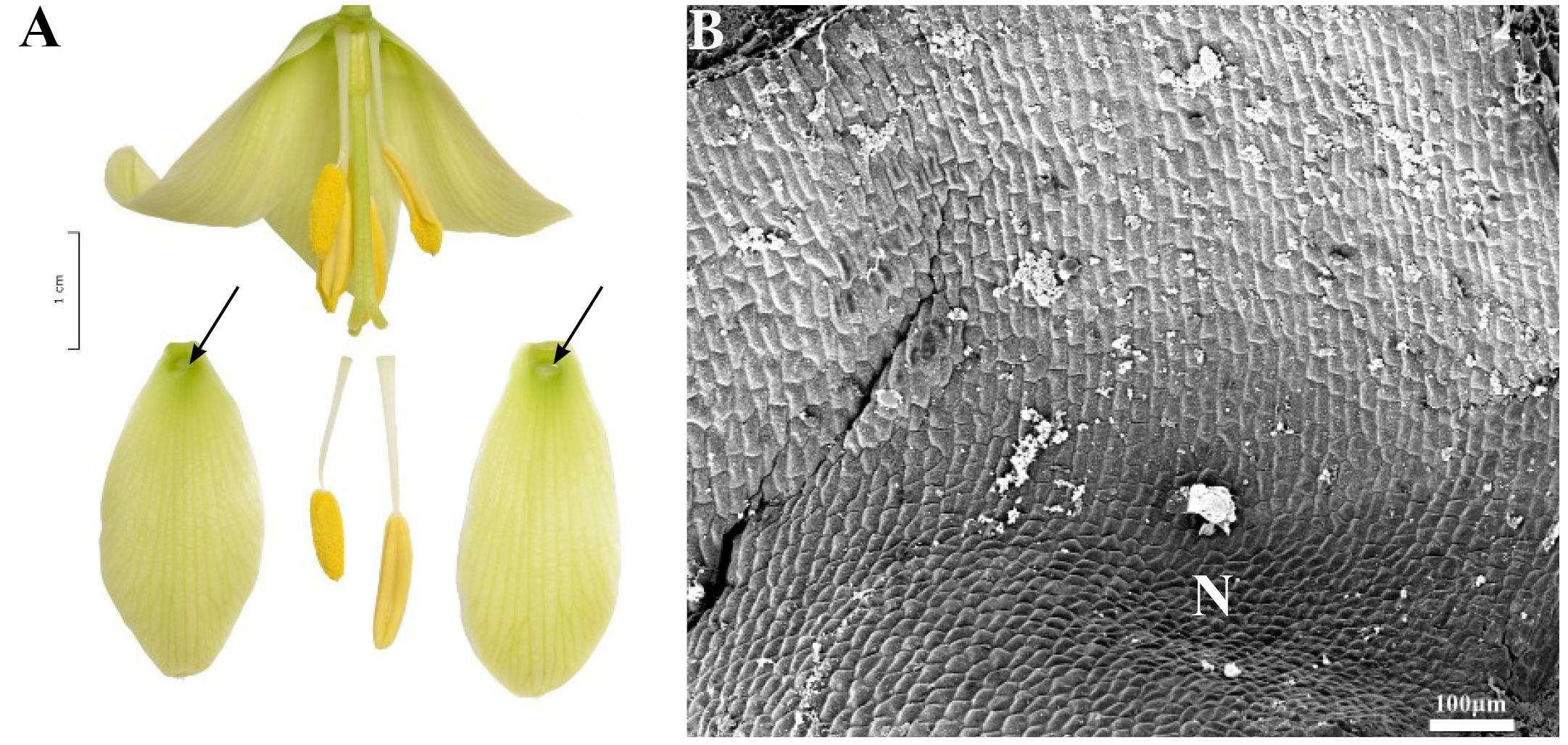
FIGURE 10. Flowers and nectaries of F. raddeana. Macro and SEM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Part of outer tepal showing depressed, cup-shaped nectary (N), and slightly convex cells of surrounding area.
Scanning electron microscope analysis revealed that the inner surface of nectaries of F. imperialis and F. eduardii were flat, whereas the surrounding area and the rest of the tepals were slightly undulate. In F. raddeana, the area within the nectary was similar to the remainder of the tepal area and was also slightly undulate (Figure 10B). Subepidermal nectary parenchyma consisted of 2–4 layers. Staining with Lugol’s iodine solution revealed the presence of numerous starch grains in the plastids of epidermal and subepidermal cells (Figure 4B).
During anthesis, the entire nectary area was coated with nectar. It was easily accessible in the form of large droplets. Nectar was replenished on its removal. Flowers of this subgenus produced, on average, 133.3 ± 107.5 μl of nectar of concentration 26.2 ± 23.1%. The highest concentration and the smallest volume were recorded for F. raddeana (50.1 ± 15.7% and 8.7 ± 1.4 μl, respectively). The lowest concentration was recorded for F. eduardii (5 ± 8.1%). Flowers of F. imperialis produced the largest volume of nectar recorded for the subgenus Petilium (204.8 ± 94.7 μl) (Table 1).
Subgenus Rhinopetalum
This subgenus was represented by three species (Table 1). The typical nectaries were deeply depressed spurs, bearing two densely papillose ridges (Figure 11A). The ridges were adpressed, protecting access to the nectaries. In F. stenanthera, there were two additional papillose ridges adjacent to the nectary area (Figure 11B). Nectaries were visible on the reverse side of the tepals as dark “horns.” These projections differed from species to species; in F. gibbosa, one of the tepal “horns” was always significantly larger than the other. In F. stenanthera, all projections were of the same size. In F. bucharica, they were of similar size – more prominent at the bud stage, becoming flatter in mature flowers. In this species, nectaries were uniformly green, like the background, but the nectaries at the top were darkly spotted, located just above the entrance to the nectary. In F. gibbosa, the area surrounding the nectaries was dark brown, but the ridges were paler. Nectaries of F. stenanthera had both green and brown elements (Figure 11A). The surrounding area was mostly brown, and the sides were greenish or dark yellow. The deeply depressed area within the spur of the nectaries of Rhinopetalum was densely clothed with short cilia. Nectaries were flat and glabrous (Figure 11B).
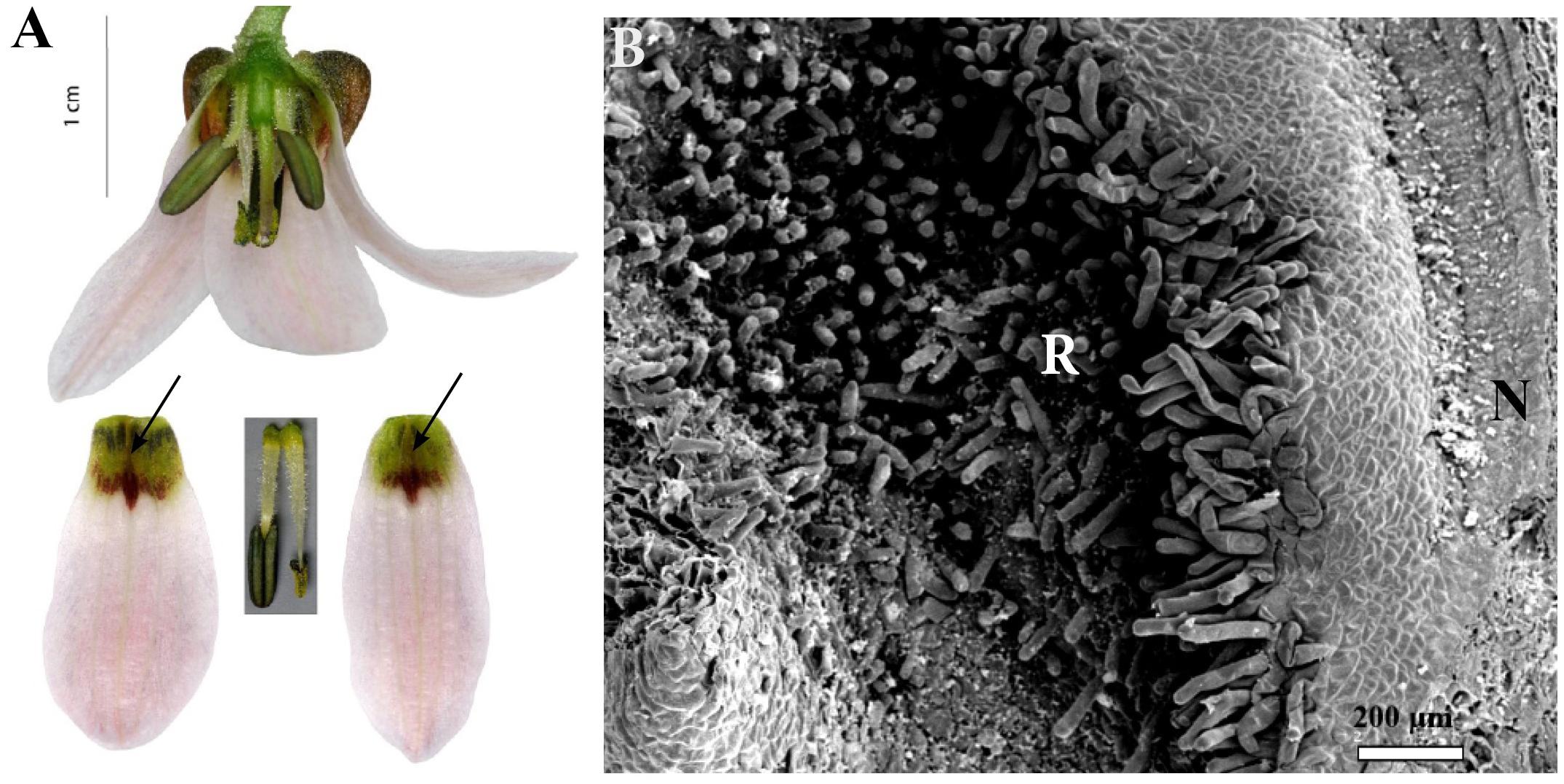
FIGURE 11. Flowers and nectaries of F. stenanthera. Macro and SEM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Part of outer tepal showing deeply depressed spurs nectary (N), protected by densely papillose ridges (R).
It was difficult to measure the size of nectaries for representatives of Rhinopetalum, since, they were hidden inside the spur, and the borders of the nectary were not obvious. Moreover, in F. gibbosa, one of the nectaries was significantly larger. The approximate mean size of the nectary for members of this genus was 7.9 ± 3.6 mm2. The largest nectaries were recorded for F. stenanthera (11.6 ± 1.4 mm2), and the smallest nectaries were recorded for F. gibbosa (7.7 ± 0.8 mm2). Nectaries of all species were located very close to the base of the perianth, the mean value of the distance to the perianth being 0.6 ± 0.3 mm (Table 1).
Scanning electron microscope analysis revealed that the surface consisted of three distinct parts. The area of nectar secretion was represented by a depressed groove having a flat surface. It was surrounded by slightly elevated walls and had a slightly undulate surface. The rest of the spur was densely coated with short cilia. The remainder of the tepal was slightly undulated (Figure 11B).
Subepidermal nectary parenchyma consisted of four or more layers, and the nectar-bearing area occupied a relatively narrow region located at the center of the tepals.
During anthesis, the entire nectary area was coated with nectar, although it was not visible and was protected by the papillose ridges. Nectar was replenished on its removal. Flowers of this subgenus produced, on average, 0.5 ± 0.8 μl of nectar of concentration 46.6 ± 14.8%. On average, F. bucharica produced 0.3 ± 0 μl of nectar of concentration 52.7 ± 1.8%, with F. stenanthera producing 0.6 ± 0.8 μl and having the lower concentration of 45.5 ± 15.9% (Table 1).
Subgenus Theresia
The nectaries of F. persica were slightly depressed and triangular in shape. They occupied an area measuring 3.5 ± 0.4 mm2 and were located 2 ± 0.3 mm above the base of the perianth (Figure 12A and Table 1). The green nectaries contrasted sharply with the surrounding dark purple background. However, several nectary cells were pigmented.
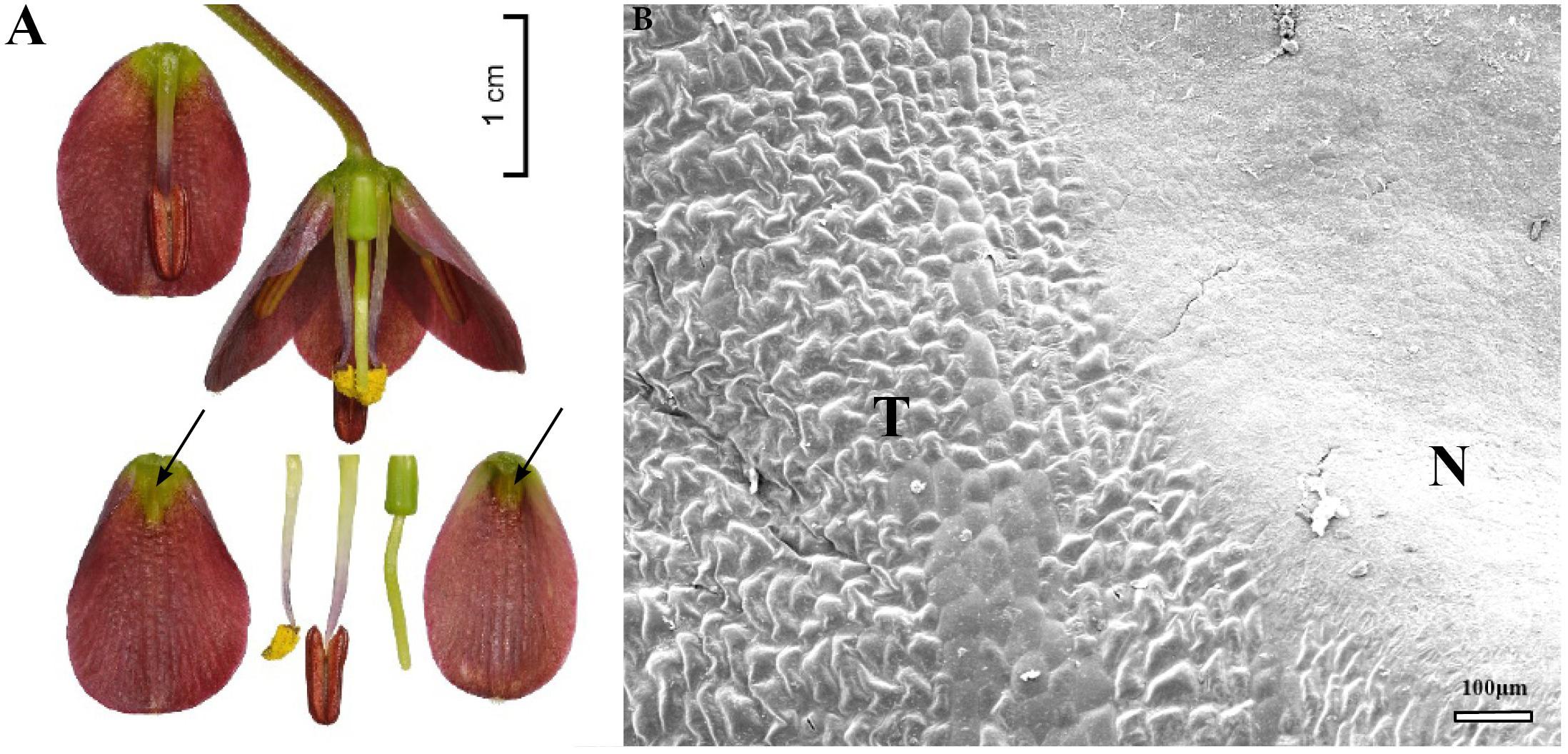
FIGURE 12. Flowers and nectaries of F. persica. Macro and SEM images. (A) Flowers and tepals, nectaries marked with arrows. (B) Part of outer tepal showing the flat nectary area (N) and slightly convex cells of surrounding area (T).
Scanning electron microscope analysis revealed that the inner surface of nectaries was flat, whereas the surrounding area and the remainder of the tepal bore slightly convex cells (Figure 12B). Subepidermal nectary parenchyma consisted of 3–5 layers.
During anthesis, the entire nectary area was coated with nectar, which extended beyond the nectary. Nectar from the inner tepals was easily accessible and lacked projections for protection, but nectar from the outer tepals was concealed behind the inner tepals. Nectar was replenished on its removal.
Fritillaria persica produced, on average, 4.3 ± 4.5 μl of nectar of concentration of 46.5 ± 18.7% (Table 1).
Other Species
We also studied the nectaries of F. grandiflora and F. olgae, which are not classified into any subgenus (Rix, 2001). However, both of them presumably belong to the subgenus Fritillaria (Rix, 1974; Kiani et al., 2017). Fritillaria grandiflora had darkly colored, round nectaries, surrounded by tessellated tepals. Fritillaria olgae had ovate to triangular, darkly colored nectaries that contrasted with the green tepals. Nectaries of F. grandiflora measured 14.7 ± 1 mm2 and were located 6.4 ± 0.6 mm from the base of the perianth, for F. olgae, these values were 19.6 ± 1.9 mm2 and 2.5 ± 0.2 mm, respectively (Table 1).
Scanning electron microscope analysis revealed that the inner surface of the nectary was flat, whereas the cells of the surrounding area and the remainder of the tepal were slightly convex. Subepidermal nectary parenchyma consisted of 3–5 layers.
During anthesis, the entire nectary area was coated with nectar. Unless collected, large droplets of nectar were found at the edges of tepals in F. olgae. Nectar was replenished on its removal. Fritillaria grandiflora produced, on average, 42.3 ± 1.3 μl nectar of concentration 23.8 ± 2.8%. Fritillaria olgae produced, on average, 74.4 ± 33.2 μl of concentration 29.5 ± 14% (Table 1).
Discussion
The present study reports SEM and LM analyses and descriptions of the nectaries for 56 species of Fritillaria contained in seven subgenera, including 29 species which were studied for the first time. This study is also the first to examine nectary surface of members of subgenera Japonica, Korolkowia, Liliorhiza, and Theresia. The most likely area used as a landing site for insect pollinators was imaged under SEM.
We also conducted SEM studies of the relevant area, which might be considered as the probable landing site for insect pollinators.
The nectaries of Fritillaria are positioned adaxially on each of the six perianth segments (Figures 1, 2). However, several other nectary features, such as size, shape, or color, were generally variable among species. Such variation in shape and position of the nectaries was previously described by Rix and Rast (1975), Bakhshi Khaniki and Persson (1997), and Kiani et al. (2017), who studied the nectaries of 31 taxa from four subgenera using SEM and LM.
Despite differences in the appearance of nectaries, their morphology and positon were similar within the different subgenera. However, our studies echoed the conclusions of Bakhshi Khaniki and Persson (1997) that morphology and position of nectaries might be important diagnostic features in the taxonomy of the genus, as the nectaries of different subgenera vary greatly. In contrast, nectary ultrastructure, which is similar among all the species studied, does not provide any taxonomic information. The only exception is the subgenus Petilium, which is distinguished from other subgenera by the presence of starch in nectary cells.
We examined the nectaries for the occurrence of callose, which may push the projections into the cytoplasm and facilitate deposition of wall material (Offler et al., 2003), but we did not observe it in the cell walls of any species, although it was previously detected as the component of wall ingrowths in F. meleagris by Stpiczyńska et al. (2012). This difference might be species-specific (F. meleagris was not included in this study) or dependent on the flowers’ development stage. Clearly, this needs further studies. However, the outer epidermal cells and/or nectary cells in all species contained lipid droplets. The presence of a lipid layer on the plant surface may provide a way to reduce water loss. However, the role of lipid within nectary cells is in need of investigation (Kamińska and Stpiczyńska, 2011).
The present study also provides information about nectar sugar composition for 34 species and the quality and volume of this reward for additional 46 species. Since the nectar properties, like concentration and amount available for floral visitors, are highly variable, it cannot serve as a taxonomic tool. Also, contrary to Rix and Rast (1975), our study indicates that the fructose/glucose ratio varies greatly within and between the species studied; therefore, it does not provide useful taxonomic information.
Subgenus Fritillaria
Usually, nectaries are more or less flat, surrounded by an area with slightly convex cells. Only in F. pyrenaica did we find the area around the nectary to be comprised of dense conical papillae, a feature that had not been previously described for any other Fritillaria species. In several species like F. davisii, F. uva-vulpis, and F. michailovskyi, there are ‘warts’ on the tepals or on the border of the nectary (Bakhshi Khaniki and Persson, 1997).
Rix and Rast (1975) studied nectar properties for several members of the subgenus Fritillaria. We obtained similar results for F. bithynica, F. elwesii, and F. pyrenaica. In comparison with the other species studied by Rix and Rast (1975), F. pallidiflora, F. crassifolia, F. michailovskyi, F. acmopetala, and F. pontica had glucose-dominant nectar in this study. Fritillaria amana produced sucrose-dominant nectar (H:S in the ratio of 2:3). Differences in the results obtained might be related to the method used (gas liquid chromatography vs. HPLC). However, it is evident that many complex factors affect nectar properties such as time of collection, weather conditions while sampling, or variation in the nectar properties depending on the development of the inflorescence (Willmer, 2011).
Subgenus Japonica
Scanning electron microscope analyses and studies of nectar properties were conducted for the first time for species in the subgenus Japonica. The structure of the nectary and tepal surface was similar to that of other Fritillaria species studied. However, in the middle of the nectar-bearing area of F. ayakoana (Figures 6B,D) was a small, upwardly curved ridge or protuberance, a feature previously described for F. kaiensis and F. japonica (Naruhashi et al., 1997). Similar ridges also occur in the closely related subgenus Rhinopetalum (Bakhshi Khaniki and Persson, 1997). We conducted the first nectar sugar analysis for F. amabilis (the volume of F. ayakoana nectar was too small to be collected in the field). It was similar to that found in other fritillary species investigated, that is, hexose-dominant, with a relatively high sugar concentration (41%). However, the amount of nectar produced was considerably lower than the mean value for the genus, and again it was similar to the values detected in members of the closely related subgenus Rhinopetalum.
Subgenus Korolkowia
Scanning electron microscope analyses and studies of nectar properties were conducted for the first time for F. sewerzowii. The tips of F. sewerzowii nectaries are visible from outside the flower; however, the main part is concealed within the very narrow, bell-shaped perianth (Figure 7A).
The surrounding papillose ridges only partly restrict access to the nectaries, in contrast to the ridges present in the members of Rhinopetalum, which almost completely cover the nectary. In this study, nectary area and floral reward were studied for the first time. The nectary area, unlike that of most fritillaries, is not flat, but it is slightly undulate and wrinkled (Figure 7B). Surprisingly, flowers of this species produced nectar without any trace of sucrose, a result which was previously only described for F. imperialis (Rix and Rast, 1975).
Subgenus Liliorhiza
Nectaries and nectar in this subgenus were studied for the first time. SEM analysis revealed that the nectaries of F. liliacea (Figure 8B) were surrounded by a row of elevated cells with grooved surfaces, similar to those found in F. camschatcensis (Figure 8D). Flowers of most members of this subgenus produced copious nectar (mean value 48–49 μl) of medium sugar concentration (mean value 31%); only in the flowers of F. camschatcensis were traces of viscous nectar found. Nectar was hexose-dominant, although, the nectar of F. gentneri also contained a substantial amount of sucrose (20%).
Subgenus Petilium
In this study, the nectaries of the subgenus Petilium differed from the other species studied. They are elliptical (Bakhshi Khaniki and Persson, 1997) and depressed. Anatomical studies revealed large accumulations of starch (Figure 4B), which was not previously reported for Fritillaria nectaries. Large amounts of starch, found, for example, in bee-pollinated Anemopaegma album (Bignoniaceae), are thought to be responsible for the secretion of large amounts of sugar during the peak secretory period (Dafni and Vereecken, 2016; Guimaraes et al., 2016). Furthermore, the white, glistening appearance of nectaries within the subgenus Petilium may also result from the presence of starch; since, the flat upper epidermis may act as a thin film reflector responsible for its glossiness. It may further serve as a filter to backscattered light as the starch bodies located in the parenchyma layers have strong light-scattering properties, as described for Ranunculus spp. (van der Kooi et al., 2017). Moreover, the convex-shaped nectaries always appear full of nectar, even when they are empty.
An earlier study (Rix and Rast, 1975) considered F. imperialis to be distinct within Fritillaria, as sucrose was absent from its nectar. This study also found that sucrose was absence in the nectar of F. eduardii. In contrast, the nectar of the very closely related F. raddeana contains sucrose and can be described as balanced, based on the ratios of the sugars it contains (fructose, glucose, and sucrose in the ratio 4:3:5). There are large differences in the rate of nectar production within the subgenus Petilium. Both F. imperialis and F. eduardii produce large volumes (204 μl per flower) of dilute nectar (9%), whereas F. raddeana produces small volumes (7 μl per flower) of highly concentrated nectar (51%). Theoretically, nectar volumes are under strong selection pressures (e.g., balancing the costs and benefits of nectar production to the plant) and genetic control. However, differences in the volume/sugar concentration of even closely related species have previously been published (Davis et al., 1994).
Subgenus Rhinopetalum
As previously described, the nectaries of this subgenus are furrowed or lobed (Bakhshi Khaniki and Persson, 1997). The aperture of the nectary spur is densely surrounded with short papillae. However, the surface of the nectary area is flat and smooth (Bakhshi Khaniki and Persson, 1997). The presence of the papillae may protect the small quantity of nectar from evaporation or crystallization (Nicolson and Thornburg, 2007). Floral features are influence by ecological factors, like habitat type (Petanidou et al., 2006), and the members of Rhinopetalum are normally found in more arid habitats, like semideserts, than is normal within Fritillaria (Rix and Zarrei, 2007a,b; Kiani et al., 2017). This study provides the first record of a hexose-rich species (F. stenanthera) within the subgenus.
Subgenus Theresia
In both this study and that of Bakhshi Khaniki and Persson (1997), nectaries of F. persica were bright and green and contrasted strongly against a dark background (Figure 12A). However, Kiani et al. (2017) showed that the appearance of the nectary area as distinct depends on the flower color which is highly variable (pale green, pale yellow, bright yellow, orange, or dark purple), and in some populations, the nectaries of F. persica may be difficult to differentiate. Nectaries of the outer tepals are shielded by the inner tepals and are, therefore, probably not easily accessible to visiting insects. SEM analysis revealed that like those of most Fritillaria species, the nectaries were uniform and flat and surrounded by an area occupied by slightly convex cells of the tepals (Figure 12B). Flowers of F. persica produce rather a small amount of nectar, but as a single specimen usually produces several dozen flowers, the overall reward is relatively plentiful. Nectar is strongly hexose-dominant.
Other Species
Scanning electron microscope analyses and studies of nectar properties were conducted for the first time for F. grandiflora and F. olgae. The nectaries of F. grandiflora and F. olgae were similar to those found in flowers of the subgenus Fritillaria. Also, SEM analysis revealed the typical Fritillaria pattern, comprising a flat and uniform nectary area surrounded by an area bearing convex cells. Both these features might indicate affinities to the subgenus Fritillaria. However, F. olgae nectar was sucrose-dominant (H:S in the ratio 2:3), such as it is generally found in passerine-pollinated species in the subgenus Petilium.
Ecological Context
The flowers of Fritillaria are very diverse – not only in color, shape, and appearance but also in the array of floral rewards like nectar sugar concentration and composition or reward location (Bakhshi Khaniki and Persson, 1997). Fritillaria have a wide geographical distribution and occupy a variety of different habitats (Hanson et al., 2009; Kiani et al., 2017). Recent DNA studies show a strong geographic relationship within Fritillaria (Day et al., 2014), even among morphologically divergent species. Species rich areas are normally associated with highly variable habitats and/or more recent oscillating climates and microclimates, resulting in numerous range changes, periods of isolation, and recombination (Myers et al., 2000; Kiani et al., 2017). However, some elements of this remarkable diversity might also be the result of a relatively rapid coevolution with their pollinators, as several species, which are distantly related have similar-looking nectaries (convergence): like, for example, F. pudica and F. carica or F. purdyi and F. crassifolia, respectively (Rix and Strange, 2014). As many fritillaries are native to remote, difficult to access, or uninhabited areas (Kiani et al., 2017), information regarding their reproduction is limited. Data concerning pollination system or Fritillaria flower visitors are only available for six species (Hedström, 1983; Búrquez, 1989; Peters et al., 1995; Minagi et al., 2005; Pendergrass and Robinson, 2005; Zox and Gold, 2008; Zych and Stpiczyńska, 2012; Zych et al., 2014).
In temperate habitats of the northern hemisphere, where Fritillaria species grow, most plants are insect-pollinated and are characterized by lack of specialization of their flowers, thus, attracting a large range of insects (Galetto et al., 1998). This is generally the case for Fritillaria, where the nectaries are variable and most are easily accessible, therefore, are likely to be visited by a range of different floral visitors. Our microscopical studies revealed that the structures of nectaries of putatively insect pollinated species are similar. Most Fritillaria species studied had a relatively flat nectary area surrounded by slightly convex cells, important for insect pollination, providing extra perch during flower manipulation by insects, thus, increasing foraging efficiency (Whitney et al., 2011; Ojeda et al., 2012). Conical papillae, found on the nectaries of F. pyrenaica, cause the thin film of the nectar to glisten. In F. davisii, the tepal surface and the area adjacent to the nectary was covered with papillae, arranged in rows along the length of the tepal. This may act as a physical nectar guide and a tactile cue, orientating insects toward both the reward and the reproductive parts of the flower. As the floral reward is easily accessible, and the corolla is normally wide open, insects can easily locate and exploit this resource.
Although data from the literature is scare, bees were seen by authors, visiting Fritillaria flowers. These animals frequently seek out flowers with medium nectar volumes of medium sugar concentration, often located toward the base of the flower (Willmer, 2011), criteria common in Fritillaria flowers. Many Fritillaria species have hexose-rich nectar, which according to floral syndrome theory is preferred by short-tongue bees (Chalcoff et al., 2006). However, bee-pollinated plants show a wide range of nectar sugar compositions, as would be expected in the nectar of flowers pollinated by such a large group (Stiles and Freeman, 1993). Pollination by bees is the most common pollinating interaction, and it would be fair to expect that melittophily is the most common syndrome in Fritillaria.
Other types of entomogamy are also present in Fritillaria, for example, F. camschatcensis is fly-pollinated (Zox and Gold, 2008). The checkered pattern found on many flowers of Fritillaria might encourage increased visitation by carrion-flies or wasps, with a strong preference for mottled petals. These groups of animals often visit large, tubular flowers with wide-open corollas and dull red, purple, brown, or greenish petals (Willmer, 2011). Several other Fritillaria species, such as F. graeca, F. montana, and F. davisii fall into this category. These species produce relatively small volumes of nectar and sometimes emit a disagreeable odor. In F. camschatcensis and F. davisii, traces of viscous and almost solid nectar form a thick film over the nectary. This would be difficult for pollinators to access. Such presentation of nectar may act as a phenotypic filter, preventing insects other than flies, with have a cushion-like labium, to gather floral rewards (Stpiczyńska et al., 2014).
To date, there is no data on pollinators or floral visitors to members of the subgenus Rhinopetalum. They have unusual nectaries concealed in sac-like structures, covered with trichomes, which are not easily accessible. Densely papillose ridges of the nectary apertures potentially exclude feeding animals with relatively short proboscises (Stolar and Davis, 2010) and/or reduce evaporation. All three species examined in this subgenus, produce small volumes of nectar with relatively high sugar concentration. Pale pink or white flowers and nectar concealed in grooves covered with fine hairs are the normal characteristics associated with butterfly pollination, which also occurs in Lilium martagon, another species with similar nectaries (Brantjes and Bos, 1980).
Pollinator availability is low for winter or early spring flowering plants as low temperatures impede insect pollinator activity. By contrast, birds, which might be considered ‘alternative pollinators,’ are warm-blooded and more reliable at low temperatures, especially where cold and/or rainy weather conditions might be frequent (Fang et al., 2012). Although several studies indicate that frequent pollinator shifts have occurred during angiosperm speciation events, it may be the case that a large proportion of these events occur relatively late within specific pollination systems (Serrano-Serrano et al., 2017). This might also be the case for Fritillaria. Moreover, evidence indicates that the switch from entomophily to ornithophily occurred at least twice during the history of the genus, once for each of the two main clades.
Two very closely related Asian members of the subgenus Petilium, F. imperialis and F. eduardii, fulfill many of the criteria that characterize ornithophilous flowers. They show diurnal anthesis, have scarlet or orange flowers, and lack nectar guides. Their pale anthers and style extend beyond the large corolla, and these robust reproductive elements are able to withstand visits by large pollinators. Although birds do not display innate preference for red (Bené, 1945; Stiles, 1976; Micheneau et al., 2006; Handelman and Kohn, 2014), flowers that are visited by these animals often have red colouration (Goldsmith and Goldsmith, 1979; McDade, 1983; Delph and Lively, 1989). This might suggest that some new characters in bird-pollinated flowers have evolved to discourage visits by illegitimate flower visitors, in this instance insects (Cronk and Ojeda, 2008; Lunau et al., 2011). In F. imperialis and F. eduardii, the pollen is pale, which makes it less attractive to insects and less prone to potential pollen theft (Wilmsen et al., 2017).
Analysis of nectary morphology revealed the absence of collenchyma, this could have helped the flower to withstand contact with a hard beak, as it occurs in several ornithophilous flowers (Stpiczyńska et al., 2004, 2005, 2009). The starch grains, found in all members of the subgenus Petilium, might be regarded as a derived strategy to support the intensive secretion of large amounts of sugar during peak nectary activity (De la Barrera and Nobel, 2004; Heil, 2011; Stpiczyńska et al., 2012). Our studies reveal that this kind of energy storing in members of Petilium had two possible results. Fritillaria raddeana produces small volumes of highly concentrated nectar, whereas F. eduardii and F. imperialis, on the other hand, produce large volumes of very dilute nectar. In fact, in F. eduardii, the concentration of nectar sugar does not even reach 10%. The results for these two species match the data available in the literature, which state that bird-pollinated flowers produce nectar whose low sugar concentration averages 20–25% (w/w) (Nicolson, 2002). This indicates that the attraction of potential bird-pollinators might be important to the various nectar features related to pollination. Moreover, the nectar of F. imperialis and F. eduardii is hexose-rich, and it lacks even traces of sucrose. As nectar originates from sucrose-rich phloem sap, the proportion of monosaccharides in the final nectar depends on the activity of invertases in the nectary wall. Hydrolysis of sucrose increases the osmolality of the nectar, and the resulting water influx can convert a 30% sucrose nectar into a 20% hexose nectar with a great (1.6 fold) increase in volume. As passerine birds are the largest bird pollinators, they require large amounts of energy and water (Nicolson, 2002).
Different components of nectar respond in different ways to various environmental factors like elevation. Relative sucrose concentration declines in response to increasing elevation, but the percentage of fructose intensifies (Stiles and Freeman, 1993). Usually, this process is gradual, suggesting the response is physiological, possibly temperature related, rather than a reduction in the selection of sucrose-rich nectar. Fritillaria imperialis grows on rocky slopes at about 1000–3000 m (Tekşen and Aytaç, 2008; Kiani et al., 2017), whereas F. eduardii grows at 1200–2100 m and F. raddeana grows at 1000 m (Kiani et al., 2017), and the average sucrose concentrations reflect this; the decline in the case of Petilium is not gradual. Moreover, species of certain plant families have nectars of relatively consistent sucrose composition (Willmer, 2011), which is also not reflected for Petilium. The concentration and composition of nectar varies greatly within this subgenus (F. imperialis and F. eduardii vs. F. raddeana). Nevertheless, the higher hexose content in the nectar of highland plants might originally have facilitated the switch to nectarivory by passerine birds (Stiles, 1978; Stiles and Freeman, 1993), and this may play a significant role in members of Petilium. Physiological constraints related to nectar production at higher altitudes may have led to sucrose elimination. Both F. eduardii and F. imperialis have pendulous, orange or reddish flowers, held on top of a thick stem, which provides a suitable perch for foraging birds. This might potentially lead to further pressure to reduce nectar concentration, which makes flowers less attractive to insect visitors and more attractive to birds, indicating that the nectar properties of F. imperialis and F. eduardii are the result of double selective pressure.
A second shift to ornithophily occurred in the distinct branch, consisting of mostly American species. Two species, sometimes co-occurring F. recurva and F. gentneri, also fulfill many of the criteria characteristic of ornithophilous flowers, that is, diurnal anthesis, scarlet flowers lacking nectar guides, and production of copious amounts of rather dilute nectar. However, the flowers of both species are held on a thin, pendulous inflorescence not suitable for perching while feeding but would suit hummingbird pollination (Willmer, 2011). The stamens of F. recurva are extended beyond the corolla tube, and, thus, allow contact between the reproductive elements and larger flower visitors. Fritillaria gentneri is a naturally occurring hybrid between F. affinis and F. recurva, and it possesses many intermediate flower features, but, without extended stamens. Flowers of F. gentneri and F. recurva are also visited under natural conditions by andrenids and halictids (Pendergrass and Robinson, 2005). It is likely that they are pollinated both by insects and birds, and we did not find many characters that might discourage illegitimate visitors. Moreover, F. affinis is postulated as an insect pollinated species.
Scanning electron microscope analysis revealed that the flat nectary area is surrounded by an area comprising slightly convex tepals cells. This might provide tactile cues for insect pollinators. Bees, for example, prefer such a surface for landing (Whitney et al., 2011). It might also help them to maintain their grip and stay inside the flowers while obtaining nectar from the flat nectary area. The epidermal wall was no thicker than in other species, nor more collenchymatous; it did not seem to provide any extra support and/or protective function, such as preventing damage to the nectary area while coming into contact with the hard beaks (Stpiczyńska et al., 2004, 2005, 2009).
As previously mentioned, nectar of F. gentneri and F. recurva was more copious and of lower concentration when compared to other closely related species of the subgenus Liliorhiza. However, the relative proportion of sugars is similar for all members of the subgenus Liliorhiza studied here, and it is characterized by high hexose concentration. Also, nectar of hummingbird-pollinated species is hexose-rich, which is unusual for hummingbird-pollinated flowers. Generally, hummingbirds visit flowers that have sucrose-dominant nectar (Cronk and Ojeda, 2008), also this matches the birds’ recorded preferences in taste tests (Baker, 1975). The intestinal walls of hummingbirds contain a sucrase enzyme, which helps them to tolerate sucrose-rich solutions (del Rio and Karasov, 1990). However, data on hummingbird preferences are often conflicted (del Rio and Karasov, 1990; Lotz and Nicolson, 1996; Willmer, 2011), as these birds freely take hexose-rich nectar when other sources are unavailable (Willmer, 2011). Many flowers visited by hummingbirds are not distinctly adapted to hummingbird-pollination. However, the capacity of hummingbirds to easily extract nectar from open melittophilous flowers, may account for the many shifts toward ornithophily. Moreover, hummingbirds are inquisitive and they investigate many flower types and designs (Wilson et al., 2007), and their spatial memory helps them to return to rewarding plants (Healy and Hurly, 2003). Plants can benefit from these visits, as hummingbirds efficiently transfer pollen even with flowers of a poor morphological fit (Wilson et al., 2007).
Characters found in putative insect-pollinated species of Fritillaria, such as rapid nectar replenishment and large, brightly colored corollas, may be regarded as preadaptations for bird-pollination (Wilson et al., 2007). This is, especially, evident in F. olgae, a species that produces copious, but rather dilute, sucrose-rich nectar (Castellanos et al., 2003).
Flower features determine which animals or group of animals will be attracted. Moreover, the character and location of the reward can significantly influence the species that are attracted. The relationship between the characteristics of Fritillaria nectar and nectaries and their diversity may guide two evolutionary processes: selection of the biotic environment for floral features (sympatric congeners and types of pollinators) and the degree of floral response to this selection (its integration and precision). The attractiveness of these features, on the other hand, ensures that the pollinators attracted to a particular species are affected by these characters. Specialization along this path could result in coevolutionary pollinator attraction or pollinator switches (Armbruster and Muchhala, 2009). From our Fritillaria study, the foundation for these switches might be the quantity and quality of the reward offered to flower visitors.
Based on our Fritillaria data, such shifts seem unlikely to generate reproductive isolation that would allow sufficient divergence of populations by pollinator selection. Therefore, it is probable that bird-pollinated species of this genus like, for example, F. imperialis or F. recurva and F. gentneri are, and will continue to be, an intermediate phase during which both ancestral and new rewards and advertisements are present, and both ancestral and new pollinators visit the same flower (Armbruster and Muchhala, 2009). There is, still, a considerable need for further studies of Fritillaria pollination system in natural habitats and the genetic basis of character shifts in relation to their pollination system.
Author Contributions
KR and MZ conceived the study and wrote the draft version of the paper. KR, LH, and PK assembled field data. KR, AB, MC, and AG performed the nectar analysis. KR, MZ, JS, MS, and LH analyzed the data. LH and KR assembled photographic documentation. All authors contributed to the final version.
Funding
The study was financially supported by research grant no. 4786 2013/11/N/NZ8/00611 from the Polish National Science Centre (to KR).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Paweł Pstrokoński, Agata Roguz, Piotr Woźniak, Susanne Sontag, Agnes Dellinger, Heidemarie Halbritter, Justyna Ryniewicz, Mateusz Skłodowski, Anna Szaciłło, and Joanna Rusecka for their help in the collection of field data and laboratory analyses; Colin Everett and Paul Cumbleton for plant material; Jan Kryciński for drawings; and Dorota Szubierajska for their help in the maintenance of experimental plants.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01246/full#supplementary-material
Footnotes
- ^ The photographs of flowers and nectaries are representative species for the Fritillaria subgenera.
References
Armbruster, S. W., and Muchhala, N. (2009). Associations between floral specialization and species diversity: cause, effect, or correlation? Evol. Ecol. 23, 159–179. doi: 10.1007/s10682-008-9259-z
Baker, H. G. (1975). Sugar concentrations in nectars from hummingbird flowers. Biotropica 7, 37–41. doi: 10.1007/BF00379522
Bakhshi Khaniki, G., and Persson, K. (1997). Nectary morphology in South West Asian Fritillaria (Liliaceae). Nord. J. Bot. 17, 579–612. doi: 10.1111/j.1756-1051.1997.tb00355.x
Beck, C. (1953). Fritillaries, A Gardener’s Introduction to the Genus Fritillaria. London: Faber & Faber.
Bené, F. (1945). The role of learning in the feeding behavior of black-chinned hummingbirds. Condor 47, 3–22. doi: 10.2307/1364444
Brantjes, N. B. M., and Bos, J. J. (1980). Hawkmoth behaviour and flower adaptation reducing self pollination in two Liliiflorae. New Phytol. 84, 139–143. doi: 10.1111/j.1469-8137.1980.tb00756.x
Búrquez, A. (1989). Blue tits, Parus caeruleus, as pollinators of the crown imperial, Fritillaria imperialis, in Britain. Oikos 55, 335–340. doi: 10.2307/3565592
Castellanos, M. C., Wilson, P., and Thomson, J. D. (2003). Pollen transfer by hummingbirds and bumblebees, and the divergence of pollination modes in Penstemon. Evolution 57, 2742–2752. doi: 10.1111/j.0014-3820.2003.tb01516.x
Chalcoff, V. R., Aizen, M. A., and Galetto, L. (2006). Nectar concentration and composition of 26 species from the temperate forest of South America. Ann. Bot. 97, 413–421. doi: 10.1093/aob/mcj043
Cronk, Q., and Ojeda, I. (2008). Bird-pollinated flowers in an evolutionary and molecular context. J. Exp. Bot. 59, 715–727. doi: 10.1093/jxb/ern009
Dafni, A., and Vereecken, N. J. (2016). Pollination biology from micro-morphological adaptations to community ecology of plant–pollinator interactions. Plant Biol. 18, 3–8. doi: 10.1111/plb.12411
Davis, A. R., Sawhney, V. K., Fowke, L. C., and Low, N. H. (1994). Floral nectar secretion and ploidy in Brassica rapa and B. napus (Brassicaceae). I. Nectary size and nectar carbohydrate production and composition. Apidologie 25, 602–614. doi: 10.1051/apido:19940611
Day, P. D., Berger, M., Hill, L., Fay, M. F., Leitch, A. R., Leitch, I. J., et al. (2014). Evolutionary relationships in the medicinally important genus Fritillaria L. (Liliaceae). Mol. Phylogenet. Evol. 80, 11–19. doi: 10.1016/j.ympev.2014.07.024
De la Barrera, E., and Nobel, P. S. (2004). Nectar: properties, floral aspects, and speculations on origin. Trends Plant Sci. 9, 65–69. doi: 10.1016/j.tplants.2003.12.003
del Rio, M. C., and Karasov, W. K. (1990). Digestion strategies in nectar- and fruit-eating birds and the sugar composition of plant rewards. Am. Nat. 136, 618–637. doi: 10.1086/285119
Delph, L. F., and Lively, M. (1989). The evolution of floral color change: pollinator attraction versus physiological constraints in Fuchsia excorticata. Evolution 43, 1252–1261. doi: 10.1111/j.1558-5646.1989.tb02572.x
Fang, Q., Chen, Y. Z., and Huang, S. Q. (2012). Generalist passerine pollination of a winter-flowering fruit tree in central China. Ann. Bot. 109, 379–384. doi: 10.1093/aob/mcr293
Galetto, L., Bernardello, G., and Sosa, C. A. (1998). The relationship between floral nectar composition and visitors in Lycium (Solanaceae) from Argentina and Chile: what does it reflect? Flora 193, 303–314. doi: 10.1016/S0367-2530(17)30851-4
Goldsmith, T. H., and Goldsmith, K. M. (1979). Discrimination of colors by the black-chinned hummingbird, Archilochus alexandri. J. Comp. Physiol. 130, 209–220. doi: 10.1007/BF00614607
Guimaraes, E., Nogueira, A., and Machado, S. R. (2016). Floral nectar production and nectary structure of a bee-pollinated shrub from Neotropical savanna. Plant Biol. 18, 26–36. doi: 10.1111/plb.12370
Handelman, C., and Kohn, J. R. (2014). Hummingbird color preference within a natural hybrid population of Mimulus aurantiacus (Phrymaceae). Plant Species Biol. 29, 65–72. doi: 10.1111/j.1442-1984.2012.00393.x
Hanson, T., Brooks, T. M., Da Fonseca, G. A., Hoffmann, M., Lamoreux, J. F., Machlis, G., et al. (2009). Warfare in biodiversity hotspots. Conserv. Biol. 23, 578–587. doi: 10.1111/j.1523-1739.2009.01166.x
Healy, S. D., and Hurly, T. A. (2003). Cognitive ecology: foraging in hummingbirds as a model system. Adv. Study Behav. 32, 325–359. doi: 10.1098/rsbl.2017.0610
Heil, M. (2011). Nectar: generation, regulation and ecological functions. Trends Plant Sci. 16, 191–200. doi: 10.1016/j.tplants.2011.01.003
Kamińska, M., and Stpiczyńska, M. (2011). The structure of the spur nectary in Dendrobium finisterrae Schltr. (Dendrobiinae, Orchidaceae). Acta Agrobot. 64, 19–26. doi: 10.5586/aa.2011.003
Kawano, S., Masuda, J., and Hayashi, K. (2008). Life-history monographs of Japanese plants. 10: Fritillaria koidzumiana Ohwi (Liliaceae). Plant Species Biol. 23, 51–57. doi: 10.1111/j.1442-1984.2008.00208.x
Kiani, M., Mohammadi, S., Babaei, A., Sefidkon, F., Naghavi, M. R., Ranjbar, M., et al. (2017). Iran supports a great share of biodiversity and floristic endemism for Fritillaria spp. (Liliaceae): a review. Plant Divers. 39, 245–262. doi: 10.1016/j.pld.2017.09.002
King, D. (2001). The Genus Fritillaria, Bulbs of North America. Portland, OR: Timber Press & North American Rock Garden Society.
Leon, C., Fay, M. F., and Rix, M. (2009). Fritillaria yuminensis. Curtis’s Bot. Mag. 26, 21–32. doi: 10.1111/j.1467-8748.2009.01633.x
Lotz, C. N., and Nicolson, S. W. (1996). Sugar preferences of a nectarivorus passerine bird, the lesser double- collared sunbird (Nectarinia chalybea). Funct. Ecol. 10, 360–365. doi: 10.2307/2390284
Lunau, K., Papiorek, S., Eltz, T., and Sazima, M. (2011). Avoidance of achromatic colours by bees provides a private niche for hummingbirds. J. Exp. Biol. 214, 1607–1612. doi: 10.1242/jeb.052688
Mabberley, D. J. (2008). Mabberley’s Plant-Book : A Portable Dictionary of Plants, their Classification and Uses, Utilizing Kubitzki’s The families and Genera of Vascular Plants (1990-) and Current Botanical Literature; Arranged According to the Principles of Molecular Systematics. Cambridge: Cambridge University Press.
McDade, L. A. (1983). Long-tailed hermit hummingbird visits to inflorescence color morphs of Heliconia irrasa. AOS 73, 309–313. doi: 10.2307/1367077
Micheneau, C., Fournel, J., and Pailler, T. (2006). Bird pollination in an angraecoid orchid on Reunion Island (Mascarene Archipelago, Indian Ocean). Ann. Bot. 97, 965–974. doi: 10.1093/aob/mcl056
Minagi, K., Yoshiya, T., and Haramm, T. (2005). Notes on flower visitors and fruit set in Fritillaria ayakoana Naruhashi (Monocotyledoneae: Liliaceae). Bull. Shimane Nat. Mus. Mt Sanbe 3, 57–60.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Naruhashi, N., Sato, T., and Noda, S. (1997). Comparative anatomy of flowers in the Fritillaria Japonica complex (Liliaceae). J. Phytogeog. Taxon. 45,1–12.
Naruhashi, N., Takata, Y., and Negoro, H. (2006). Pollinators and dispersing insects of seeds in Fritillaria koidzumiana (Liliaceae). J. Phytogeog. Taxon. 54, 57–63.
Nicolson, S., and Thornburg, R. (2007). “Nectar chemistry,” in Nectaries and Nectar, eds S. W. Nicolson, M. Nepi, and E. Pacini (Berlin: Springer), 215–264. doi: 10.1007/978-1-4020-5937-7_5
Nicolson, S. W. (2002). Pollination by passerine birds: why are the nectars so dilute? Comp. Biochem. Physiol. B 131, 645–652. doi: 10.1016/S1096-4959(02)00014-3
Offler, C. E., McCurdy, D. W., Patrick, J. W., and Talbot, M. J. (2003). Transfer cells: cells specialized for a special purpose. Annu. Rev. Plant Biol. 54, 431–454. doi: 10.1146/annurev.arplant.54.031902.134812
Ojeda, I., Santos-Guerra, A., Caujapé-Castells, J., Jaén-Molina, R., Marrero,Á, and Cronk, Q. C. B. (2012). Comparative micromorphology of petals in Macaronesian Lotus (Leguminosae) reveals a loss of papillose conical cells during the evolution of bird pollination. Int. J. Plant Sci. 173, 365–374. doi: 10.1086/664713
Pendergrass, K., and Robinson. (2005). A Recovery Plan for Fritillaria gentneri (Gentner’s fritillary). Washington, D.C: U.S. Fish and Wildlife Service.
Petanidou, T., Van Laere, A., Ellis, W. N., and Smets, E. (2006). What shapes amino acid and sugar composition in Mediterranean floral nectars? Oikos 115, 155–169. doi: 10.1111/j.2006.0030-1299.14487.x
Peters, W. S., Michael, P., Gerhard, G., and Peters, D. S. (1995). Pollination of the crown imperial Fritillaria imperialis by great tits Parus major. J. Ornithol. 136, 207–212. doi: 10.1007/BF01651242
Rix, E. M. (1974). Notes on Fritillaria (Liliaceae) in the Eastern Mediterranean Region, I & II. Kew Bull. 29, 633–654. doi: 10.2307/4108130
Rix, E. M. (1984). “Fritillaria L,” in Flora of Turkey and the East Aegean Islands, eds P. H. Davis and R. R. Mill (Edinburgh: Univeristy Press), 284–302.
Rix, E. M. (2001). Fritillaria A Revised Classification. Edinburgh: The Fritillaria Group of the Alpine Garden Society.
Rix, E. M., and Rast, D. (1975). Nectar sugars and subgeneric classification in Fritillaria. Biochem. Syst. Ecol. 2, 207–209. doi: 10.1016/0305-1978(75)90063-0
Rix, M., and Strange, K. (2014). Fritillaria sororum Liliaceae. Curtis’s Bot. Mag. 31, 214–222. doi: 10.1111/curt.12075
Rix, M., and Zarrei, M. (2007a). Fritillaria gibbosa. Curtis’s Bot. Mag. 24, 50–53. doi: 10.1111/j.1467-8748.2007.00560.x
Rix, M., and Zarrei, M. (2007b). Fritillaria karelinii. Curtis’s Bot. Mag. 24, 46–49. doi: 10.1111/j.1467-8748.2007.00559.x
Rønsted, N., Law, S., Thornton, H., Fay, M. F., and Chase, M. W. (2005). Molecular phylogenetic evidence for the monophyly of Fritillaria and Lilium (Liliaceae; Liliales) and the infrageneric classification of Fritillaria. Mol. Phylogenet. Evol. 35, 509–527. doi: 10.1016/j.ympev.2004.12.023
Santana, D. (1984). Morphological and Anatomical Observations on the Bulbs, and Tepals of Fritillaria (Liliaceae) Section Liliorhiza (Kellogg) Watson, (their) Taxonomic Implications with a Synopsis and the Reproductive Biology of the Section. Berkeley, CA: University of California.
Serrano-Serrano, M. L., Rolland, J., Clark, J. L., Salamin, N., and Perret, M. (2017). Hummingbird pollination and the diversification of angiosperms: an old and successful association in Gesneriaceae. Proc. Biol. Sci. 284:20162816. doi: 10.1098/rspb.2016.2816
Stiles, F. G. (1976). Taste preferences, color preferences, and flower choice in hummingbirds. Condor 78, 10–26. doi: 10.2307/1366912
Stiles, F. G. (1978). Ecological and evolutionary implications of bird pollination. Am. Zool. 18, 718–727. doi: 10.1093/icb/18.4.715
Stiles, F. G., and Freeman, C. E. (1993). Patterns in floral nectar characteristics of some bird-visited plant species from Costa Rica. Biotropica 25, 191–205. doi: 10.2307/2389183
Stolar, J., and Davis, A. R. (2010). Floral nectary structure, nectar production, and carbohydrate composition in the Lilium asiatic hybrid ‘Trésor’. Botany 88, 185–205. doi: 10.1139/B09-109
Stpiczyńska, M., Davies, K. L., and Gregg, A. (2004). Nectary structure and nectar secretion in Maxillaria coccinea (Jacq.) L.O. Williams ex Hodge (Orchidaceae). Ann. Bot. 93, 87–95. doi: 10.1093/aob/mch008
Stpiczyńska, M., Davies, K. L., and Gregg, A. (2005). Comparative account of nectary structure in Hexisea imbricata (Lindl.) Rchb.f. (Orchidaceae). Ann. Bot. 95, 749–756. doi: 10.1093/aob/mci081
Stpiczyńska, M., Davies, K. L., and Gregg, A. (2009). Nectary structure of Ornithidium sophronitis Rchb.f. (Orchidaceae: Maxillariinae). Acta Agrobot. 62, 3–12. doi: 10.5586/aa.2009.021
Stpiczyńska, M., Nepi, M., and Zych, M. (2012). Secretion and composition of nectar and the structure of perigonal nectaries in Fritillaria meleagris L. (Liliaceae). Plant Syst. Evol. 298, 997–1013. doi: 10.1007/s00606-012-0609-5
Stpiczyńska, M., Nepi, M., and Zych, M. (2014). Nectaries and male-biased nectar production in protandrous flowers of a perennial umbellifer Angelica sylvestris L. (Apiaceae). Plant Syst. Evol. 301, 1099–1113. doi: 10.1007/s00606-014-1152-3
Tamura, M. N. (1998). “Liliaceae,” in The Families and Genera of Vascular Plants. III. Flowering Plants. Monocotyledons, Lilianae (except Orchidaceae), ed. K. Klaus (Berlin: Springer), 343–353.
Tekşen, M., and Aytaç, Z. (2008). Fritillaria mughlae (Liliaceae), a new species from Turkey. Ann. Bot. Fenn. 45, 141–147. doi: 10.5735/085.045.0209
Tekşen, M., and Aytaç, Z. (2011). The revision of the genus Fritillaria L. (Liliaceae) in the Mediterranean region (Turkey). Turk J. Bot. 35, 447–478.
Tomović, G., Vukojičić, S., Niketić, M., Zlatković, B., and Stevanović, V. (2007). Fritillaria (Liliaceae) in Serbia: distribution, habitats and some taxonomic notes. Phytol. Balc. 13, 359–370.
van der Kooi, C. J., Elzenga, J. T., Dijksterhuis, J., and Stavenga, D. G. (2017). Functional optics of glossy buttercup flowers. J. R. Soc. Interface 14, 1–9. doi: 10.1098/rsif.2016.0933
White, G. (1789). The Natural History & Antiquities of Selborne. New York, NY: J. M. Dent & Sons Ltd.
Whitney, H. M., Bennett, K. M., Dorling, M., Sandbach, L., Prince, D., Chittka, L., et al. (2011). Why do so many petals have conical epidermal cells? Ann. Bot. 108, 609–616. doi: 10.1093/aob/mcr065
Willmer, P. (2011). Pollination and Floral Ecology. Princeton, NJ: Princeton University Press. doi: 10.1515/9781400838943
Wilmsen, S., Gottlieb, R., Junker, R. R., and Lunau, K. (2017). Bumblebees require visual pollen stimuli to initiate and multimodal stimuli to complete a full behavioral sequence in close-range flower orientation. Ecol. Evol. 7, 1384–1393. doi: 10.1002/ece3.2768
Wilson, P., Wolfe, A. D., Armbruster, W. S., and Thomson, J. D. (2007). Constrained lability in floral evolution: counting convergent origins of hummingbird pollination in Penstemon and Keckiella. New Phytol. 176, 883–890. doi: 10.1111/j.1469-8137.2007.02219.x
Wykes, G. (1952). An investigation of the sugars present in the nectar of flowers of various species. New Phytol. 51, 210–215. doi: 10.1111/j.1469-8137.1952.tb06127.x
Zaharof, E. (1986). Variation and taxonomy of Fritillaria graeca (Liliaceae) in Greece. Plant Syst. Evol. 154, 41–46. doi: 10.1007/BF00984867
Zox, H., and Gold, W. (2008). Ecology of black lily (Fritillaria camschatcensis) a Washington State sensitive species. Douglasia (Fall) 32, 17–22.
Zych, M., Goldstein, J., Roguz, K., and Stpiczyńska, M. (2013). The most effective pollinator revisited: pollen dynamics in a spring-flowering herb. Arthropod Plant Interact. 7, 315–322. doi: 10.1007/s11829-013-9246-3
Zych, M., and Stpiczyńska, M. (2012). Neither protogynous nor obligatory out-crossed: pollination biology and breeding system of the European Red List Fritillaria meleagris L. (Liliaceae). Plant Biol. 14, 285–294. doi: 10.1111/j.1438-8677.2011.00510.x
Keywords: Fritillaria, nectar, pollinator shift, ornithogamy, nectary surface, SEM
Citation: Roguz K, Bajguz A, Goębiewska A, Chmur M, Hill L, Kalinowski P, Schönenberger J, Stpiczyńska M and Zych M (2018) Functional Diversity of Nectary Structure and Nectar Composition in the Genus Fritillaria (Liliaceae). Front. Plant Sci. 9:1246. doi: 10.3389/fpls.2018.01246
Received: 30 March 2018; Accepted: 06 August 2018;
Published: 24 September 2018.
Edited by:
Robert W. Thornburg, Iowa State University, United StatesReviewed by:
Juan De Dios Alché, Consejo Superior de Investigaciones Científicas (CSIC), SpainHester Sheehan, University of Cambridge, United Kingdom
Copyright © 2018 Roguz, Bajguz, Gołębiewska, Chmur, Hill, Kalinowski, Schönenberger, Stpiczyńska and Zych. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Katarzyna Roguz, ay5yb2d1ekBiaW9sLnV3LmVkdS5wbA== Marcin Zych, bXp5Y2hAYmlvbC51dy5lZHUucGw=