 Angela Sierra-Almeida
Angela Sierra-Almeida Lohengrin A. Cavieres1,2
Lohengrin A. Cavieres1,2 León A. Bravo
León A. Bravo- 1Departamento de Botánica, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción, Concepción, Chile
- 2Instituto de Ecología y Biodiversidad, Santiago, Chile
- 3Departamento de Ciencias Agronómicas y Recursos Naturales, Facultad de Ciencias Agropecuarias y Forestales and Center of Plant, Soil Interactions and Natural Resources Biotechnology, Scientific and Technological Bioresource Nucleus, Universidad de La Frontera, Temuco, Chile
Although positive effects on growth and reproduction of Antarctic vascular plants have been reported under warmer temperatures, it could also increase the vulnerability of these plants to freezing. Thus, we assessed in situ whether warming decreases the freezing resistance of Colobanthus quitensis and Deschampsia antarctica, and we compared the level and mechanism of freezing resistance of these species in the field with previous reports conducted in lab conditions. We assessed the freezing resistance of C. quitensis and D. antarctica by determining their low temperature damage (LT50), ice nucleation temperature (NT) and freezing point (FP) in three sites of the King George Island. Plants were exposed during two growing seasons to a passive increase in the air temperature (+W). +W increased by 1K the mean air temperatures, but had smaller effects on freezing temperatures. Leaf temperature of both species was on average 1.7K warmer inside +W. Overall, warming decreased the freezing resistance of Antarctic species. The LT50 increased on average 2K for C. quitensis and 2.8K for D. antarctica. In contrast, NT and FP decreased on average c. 1K in leaves of warmed plants of both species. Our results showed an averaged LT50 of -15.3∘C for C. quitensis, and of -22.8∘C for D. antarctica, with freezing tolerance being the freezing resistance mechanism for both species. These results were partially consistent with previous reports, and likely explanations for such discrepancies were related with methodological differences among studies. Our work is the first study reporting the level and mechanisms of freezing resistance of Antarctic vascular plants measured in situ, and we demonstrated that although both plant species exhibited a great ability to cope with freezing temperatures during the growing season, their vulnerability to suffer freezing damage under a warming scenario increase although the magnitude of this response varied across sites and species. Hence, freezing damage should be considered when predicting changes in plant responses of C. quitensis and D. antarctica under future climate conditions of the Antarctic Peninsula.
Introduction
Antarctica is the coldest and windiest landmass on Earth (Robinson et al., 2003). Mean air temperatures in the coastal zone of the Antarctic Peninsula and adjacent islands (also called Maritime Antarctica) seldom exceed 0 or +5°C during the summer (Smith, 2003), with daily temperature ranges from -10 to +15°C for the same period (Convey, 2013). Thus, Antarctic plants are constantly dealing with low temperatures, even during the growing season (Convey, 1996; Convey et al., 2014). For this reason, low temperature stress seems to be part of the explanation for its lower plant species diversity compared to the Arctic (Convey, 2006). Only two vascular plants have been able to establish natural populations in the maritime Antarctica: The pearlwort Colobanthus quitensis (Kunth) Bartl. (Caryophyllaceae) and the hair grass Deschampsia antarctica Desv. (Poaceae) (Smith, 2003).
The constant low temperatures that characterize the growing period in the Maritime Antarctica are likely to be near the minimum thresholds for many physiological processes. This suggests that in the context of climate change, small increments in the temperature experienced by plants in this environment will have a relatively greater biological impact than the same increment experienced in a less extreme environment (Convey, 2001, 2006). During the last part of the past century air temperatures in the Antarctic Peninsula increased at a faster rate than the rest of Antarctica and the globe (Vaughan et al., 2003; Turner et al., 2014). This warmer climate caused longer growing seasons with higher temperatures, ice retreats and higher frequency of rains, which promoted the expansion and increase of population sizes and numbers of C. quitensis and D. antarctica along the Peninsula (Fowbert and Smith, 1994; Gerighausen et al., 2003; Torres-Mellado et al., 2011; Cannone et al., 2016). Apparently, warmer temperatures favored plant growth and reproduction by providing more favorable thermal conditions for different physiological processes (e.g., photosynthesis), but also by increasing plant nutrient availability via stimulation of soil microbial activity in N-cycling (Wasley et al., 2006; Yergeau et al., 2012). However, these positive effects of warmer temperatures in Antarctica could be negligible because warming can decrease plant freezing survival (Inouye, 2000).
The ability to survive freezing temperatures (i.e., freezing resistance) is highly related to the ambient temperature that plants experience (Beck et al., 2004; Bannister et al., 2005). Thus, warmer daytime temperatures due to climate change may decrease the ability of plant to survive freezing conditions. This is particularly important for high-latitude and -elevation plants to resist freezing temperatures, where it has been shown that warming turned even the most freezing-resistant species more vulnerable to damage by freezing (Loveys et al., 2006; Woldendorp et al., 2008; Rixen et al., 2012). For example, Marchand et al. (2006) reported a reduction of plant performance (i.e., leaf relative chlorophyll content, maximum efficiency of photosystem II, stomatal conductance) of four Arctic plant species after they were exposed to consecutive heat waves. Apparently, warmer conditions reduced cold acclimation of plants, resulting in damage after the exposure to the natural low temperatures of the Arctic. Likewise, Sierra-Almeida and Cavieres (2010) reported that in situ warmer temperatures decreased on average 4K the freezing resistance of seven alpine species of central Chilean Andes. Although Turner et al. (2016) reported that warming in the Antarctic Peninsula (AP) has stopped in the last decade; they warned that new warming episodes are likely to occur in the future. In addition, Lee et al. (2017) pointed out that the recently paused warming observed in the AP is a consequence of short-term natural climate variability and that a new warming phase will be observed across the AP. Thus, to assess whether warmer temperatures reduce the ability of Antarctic plants to resist freezing is crucial to predict their vulnerability to future warming events.
Chronically low temperatures of the Antarctica suggest that C. quitensis and D. antarctica are morphological and physiologically adapted to cope with these stressful conditions. Overall, plants have two physiological mechanisms of freezing resistance: avoidance and tolerance. Freezing avoidance (FA) prevents the ice formation through freezing point depression or by supercooling, meanwhile freezing tolerance (FT) is defined as the ability of plants to survive the extracellular freezing (Larcher, 2003). In the case of Antarctic plants, they have exhibited freezing avoidance and tolerance mechanisms (Bravo et al., 2001; Reyes-Bahamonde, 2013). Molecular and biochemical aspects of the freezing resistance have been studied in these species, especially in D. antarctica (e.g., Bravo and Griffith, 2005; Olave-Concha et al., 2005; Piotrowicz-Cieślak et al., 2005; Bravo et al., 2009; Zúñiga-Feest et al., 2009). However, studies dealing with the freezing temperature causing injury to these species are scarce (Bravo et al., 2001; Gianoli et al., 2004; Chew et al., 2012). Studies with laboratory grown plants have reported that the freezing resistance of C. quitensis fluctuates between -14 and -4.8°C, whilst for D. antarctica it fluctuates between -26.4 and -12°C. Discrepancies among those studies on the level of freezing resistance of Antarctic plants have been attributed to methodological issues. Particularly, the time and temperature that plants were maintained under greenhouse and/or growth chambers before freezing injury assays varied enormously among them. Therefore, in situ determinations are required to unveil the real level of freezing resistance of the Antarctic vascular plants and the mechanisms involved. In addition, manipulative field experiments are needed to assess the likely effect of warming in this important trait.
In this study, we conducted a field experiment in the King George Island, where we increased the air temperatures experienced by C. quitensis and D. antarctica during two growing seasons to assess their in situ vulnerability to freezing damage under different thermal conditions. Specifically, our aims were: (1) to assess the in situ level of the freezing resistance of C. quitensis and D. antarctica and whether warming decrease this ability (i.e., leaf NT, FP and/or LT50 of warmed plants should occur at higher temperatures than of unwarmed plants); and (2) to compare the level and mechanism of freezing resistance of these Antarctic plants species in the field with previous reports conducted under lab conditions.
Materials and Methods
Study Area
This study was carried out in the King George Island, South Shetland Archipelago, nearby the Henryk Arctowski Polish Station (62°09′ S, 58°28′ W). Plants were obtained from three sites, which differed in soil nutrients, plant cover and relative abundance of the Antarctic vascular species (Supplementary Table S1 and Figure 1). Site 1 (62° 9′43.33″S; 58°27′58.80″W) was located near the beach, about 90 m from the coast line thus receiving sea spray, plant cover is >90% and vegetation is dominated by D. antarctica. This site receives great inputs of guano and feces because of the activity of sea birds and mammals (see Supplementary Table S1 for soil nutrient contents). Site 2 (62° 9′49.15″S; 58°28′9.60″W) was located 200 m distant of the site 1, plant cover is 100% and it is dominated by a compact and continuous moss carpet where C. quitensis and D. antarctica are growing interspaced (Cavieres et al., 2018). This site seems to be favorable for plant growth because of well drained soils and nutrient availability (Figure 1 and Supplementary Table S1). Site 3 (62° 9′52.90″S; 58°28′21.31″W) is a typical fellfield located 550 m from the beach and at 30 m a.s.l. Plant cover is <10%, and the scarce vegetation is dominated by lichens and only isolated individuals of C. quitensis and D. antarctica species are present across a stony and rocky soil matrix (Figure 1).

FIGURE 1. Study area in the King George Island (South Shetland Archipelago) where experimental warming was installed. Photographs correspond to (A) Site 1, nearest the beach; (B) Site 2, and (C) Site 3; (D) showed several individuals of C. quitensis inside an OTC (see Materials and Methods sections for site descriptions and experimental design).
The growing season in the study area usually starts with the snowmelt in December and finishes in March with the first snowfall. During two growing periods we collected microclimatic data (83 and 55 days for the first and second growing season, respectively), period ever which the daily mean air temperature was 1.8°C, with mean maximum and minimum temperatures of 4.7 and -0.4°C, respectively, with night freezing temperatures occurring frequently during the entire growing seasons (Table 1). Precipitation occurs as rain in summer, with estimations that range from 350 to 750 mm (Green et al., 2007).
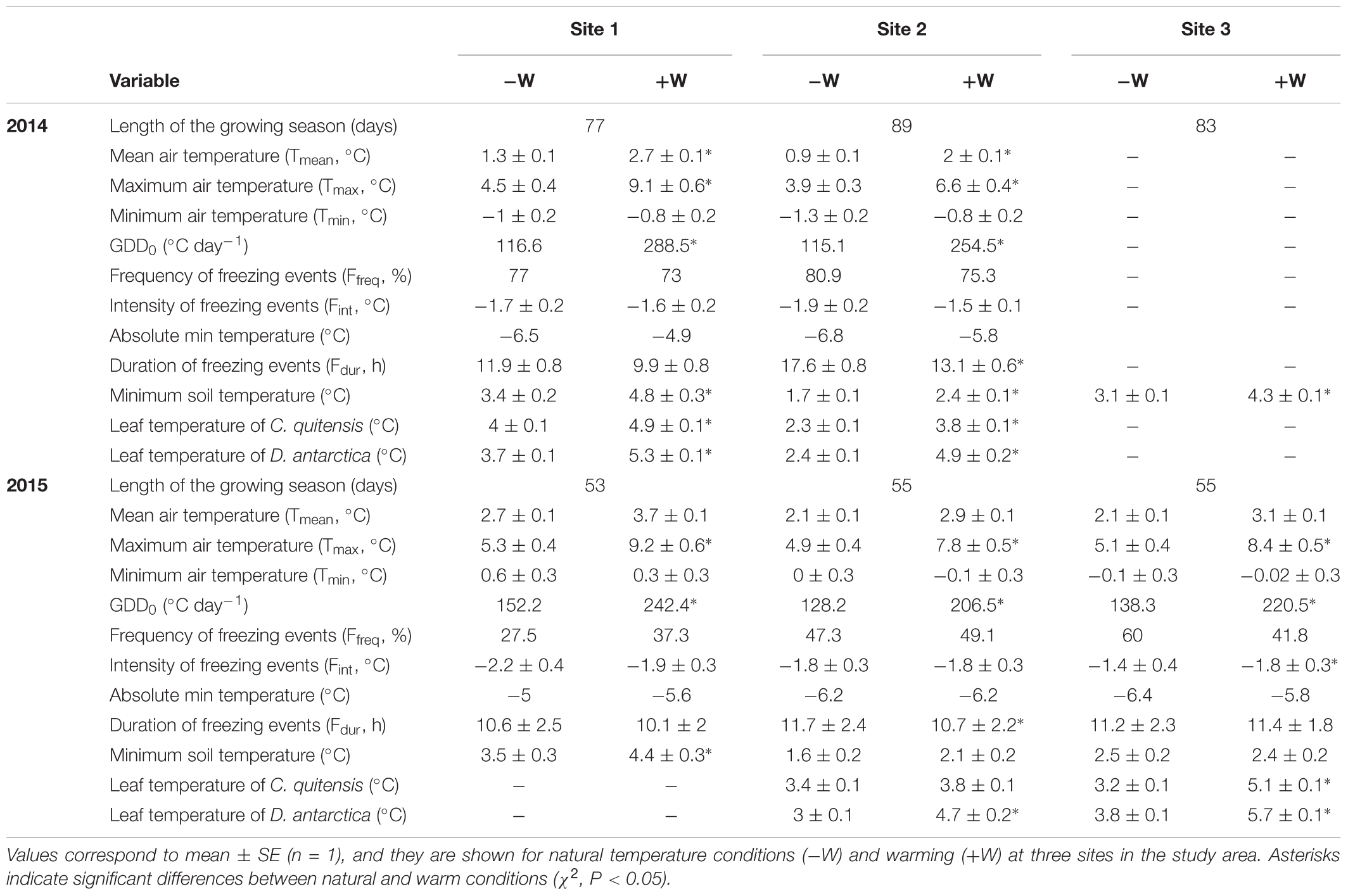
TABLE 1. Thermal conditions of Antarctic plants exposed to experimental warming during two growing seasons in the King George Island, Maritime Antarctica (Antarctic Peninsula).
Plant Species
Studied species were the pearlwort C. quitensis (Kunth) Bartl. (Caryophyllaceae) and the hair grass D. antarctica Desv. (Poaceae). C. quitensis is a long-lived perennial herb. It forms low, compact, discrete cushions with densely packed shoots and a log taproot (Greene and Holtom, 1971). Its geographical distribution comprises from Mexico and from the Andes mountains of Ecuador down to c. 68°S in the Maritime Antarctica (Moore, 1970). Despite its wide latitudinal distribution, this species inhabits sites with similar conditions, characterized by sparsely vegetated, sheltered, moist, and well-drained mineral soils (Smith, 2003). D. antarctica is a long-lived perennial herb that forms low, caespitose shallow-rooted tufts (Greene, 1964; Moore, 1979). D. antarctica distributes from central Chile and Argentina (33°S) to the Terra Firma Islands southwestern Antarctic Peninsula (68°S; Smith and Poncet, 1987). In Antarctica this species colonize habitats ranging from mineral to organic soils, from well drained to waterlogged areas, and from nutrient-deficient to highly nutrient-enriched habitats (Smith, 2003).
Experimental Design
In December 2013, on each site we selected seven plant individuals of C. quitensis and D. antarctica. On each individual, we placed a hexagonal Open Top Chamber (thereafter OTC), similar to those used in the International Tundra Experiment (ITEX). Each OTC was made with transparent Plexiglass®walls of 40 cm height, 115 cm in basal diameter, and reinforced with aluminum profiles. OTCs walls were punched with 25 holes of 1.5 cm diameter each to allow some wind to pass through and hence avoid an excessive increase in air temperature. OTCs were secured to the ground with ropes to avoid being moved and/or destroyed by the strong winds. Another seven individuals per species were randomly selected at 2 m distant from the nearest OTC. These individuals were growing under natural temperatures conditions. Hence, we obtained two experimental conditions with 7 replicates each: warmed (+W) and unwarmed control plants (-W) repeated in three sites. The spatial arrangement of both OTC and control plots was random, taking care that distance between OTC is enough to avoid any possible effects of OTCs on the neighboring control plots by affecting wind or snow deposition. Although the use of passive warming systems such as OTC has been controversial (e.g., Kennedy, 1995; De Boeck et al., 2012), some authors arguably consider that OTCs are a reasonable analog of regional warming for remote areas such as polar habitats (Hollister and Webber, 2000; Bokhorst et al., 2013).
Microclimatic conditions were monitored in warming and control plots during two growing seasons (Table 1). For this, a weather station was installed on each site (2 units HOBO®U-30 Station, Onset Computer Co., Bourne, MA, United States; 1 Em50 Data Logger, Decagon Devices Inc., Pullman, WA, United States). Air and soil temperature sensors were placed at 5 cm (n = 1) above and 5 cm below (n = 1) ground surface, and temperature conditions were recorded every hour. In addition, leaf temperature was measured for warmed and unwarmed plants of C. quitensis and D. antarctica. For this, each leaf temperature sensor was placed beneath a leaf (n = 1) and were connected to the weather station programmed to record temperature every hour. Missing leaf temperatures data in site 1 during 2015 were absent because sensors were destroyed. A similar situation occurred with air and leaf temperature sensors in site 3 during 2014. Due to logistic limitations to access the study area, weather stations were installed and uninstalled in the field for each growing season. Thus, records of microclimatic conditions in the Table 1 started in December 10th 2013 and finished in March 8th 2014 for the first growing season. For the second growing season records started in January 10th (sites 2 and 3) and 13th (site 1) and finished in March 4th 2015.
Air temperature data were used to estimate growing degree days (GDDs; McMaster and Wilhelm, 1997). GDDs were used as a measure of the accumulated amount of heat (in °C) above a base temperature to represent a cumulative index of the energy available to growing plants, according to the formula:
The daily GDDs were summed per each entire growing season. We used 0°C as a conservative base growing temperature (the temperature above which plants can perform metabolic functions, e.g., photosynthesis, cell elongation), because plants from cold climate generally vary in their absolute base growing temperature, and this value encompasses this variability (Körner, 2011).
Freezing Resistance Determinations
Plant Material Collection
We collected seven plant samples replicates for each species, experimental condition and site, excepting by the site 3 where we collected six replicates. Plant samples corresponded to complete individuals with at least seven modules (small rosettes or tillers). We collected all plant material between 11:00 AM and 12:00 PM. Plant samples were placed in plastic boxes with belowground organs wrapped in wet paper to prevent changes in tissue water content and mechanical damage. Samples were then transported to a field laboratory at the Polish Scientific Station, less than 10 min away from the study sites. We kept plant samples outdoor but protected from wind until freezing resistance determinations were performed within 24 h of collection, which were carried out between February 25th and March 5th 2015.
Low Temperature Damage
For each species, experimental condition and site, we estimated the freezing temperature producing 50% damage (LT50). For this, we selected and detached six rosettes/tillers from different plant samples, and they were separated into six subsamples. One subsample was used as control and stored at 2°C and darkness during 24 h. Remaining five subsamples were separately placed in a small plastic bag, which was then placed in a larger plastic bag with a weight to ensure that each subsample was submerged in a cryostat (F25-ME, Julabo Labortechnik GmbH, Germany) with antifreeze solution (Polycool Mix 25, PolyScience, IL, United States). Cryostat was cooled previously at five different target temperatures: -8, -12, -16, -20 and -25°C. All subsamples were transferred from outdoor to the cryostat and incubated during 2 h to reach homogeneous leaf temperatures. After that freezing treatment, subsamples were removed from the cryostat and placed back into cold room, under darkness and at 2°C during 24 h for thawing. In most studies dealing with plant freezing resistance, samples are cooled gradually to determine LT50 (2–5 K h-1; e.g., Hekneby et al., 2006; Ladinig et al., 2013; Briceño et al., 2014). However, given that cooling rates used by previous studies dealing with freezing resistance of Antarctic vascular plants varied from 1 to 17 K h-1 (Bravo et al., 2001; Gianoli et al., 2004; Chew et al., 2012), it makes impossible to find a consensus cooling rate for comparative purposes. Although sudden cooling can lead to increased tissue damage owing to anomalous water diffusion and ice crystal formation (Guy, 2003; Wisniewski et al., 2014), our procedure induced similar plant damage as a cooling rate of 16 K h-1 (see Pescador et al., 2018 for details of cooling rates assay), enabling to assess cooling directly as proxy for natural and immediate freezing exposure (Larcher et al., 2010).
Leaf damage was assessed as percentage of photoinactivation (PhI) of the photosystem II as described by Larcher (2000). For this, we measured the ratio of variable to maximum fluorescence (Fv/Fm) of dark-adapted leaf by using a chlorophyll fluorometer (MINI-PAM, Walz, Germany). LT50 corresponds to the temperature at which PhI reaches a 50% value in subsamples, and was determined by linear interpolation using the temperature of the highest PhI of <50% and the temperature of the lowest PhI of >50% (Bannister et al., 2005). PhI was chosen because it measures changes in photosynthetic performance that correlates very well with direct measurements of tissue damage (i.e., visual freezing injuries and vital stain; Boorse et al., 1998; Neuner and Buchner, 1999) and because is an easy, rapid and cheap method to work in areas with difficult logistic as it is the Antarctica.
Thermal Analyses
A small module (rosette or tiller) was removed from each of six-seven plant samples taken from each species, experimental condition and sites. Each module was attached to a thermocouple (Gauge 30 copper-constantan thermocouples; Cole Palmer Instruments, Vernon Hills, IL, United States), and immediately enclosed in a small, tightly closed cryotube. The cryotubes were placed in a cryostat (F25-ME, Julabo Labortechnik GmbH, Germany), and the temperature was decreased from 0 to -20°C, at a cooling rate of 2 K h-1. The temperature of individual module was monitored every second with a Personal Daq/56 multi-channel thermocouple USB data acquisition module (IOtech, Cleveland, OH, United States). The sudden rise in leaf temperature (exotherm) produced by the heat released during the extracellular freezing process was used to determine two variables: the ice nucleation temperature (NT), which corresponds to the lowest temperature before the exotherm, indicating the onset of ice crystal formation, and the freezing point (FP), the highest point of the exotherm, indicating the freezing of water in the apoplast, including symplastic water driven outward by the water potential difference caused by the apoplastic ice formation (Larcher, 2003). We chose this cooling rate because it is the same used by previous studies where thermal analyses were carried out (Bravo et al., 2001; Reyes-Bahamonde, 2013).
Statistical Analyses
Differences in air (i.e., mean, maximum, minimum, intensity, and duration of freezing events) and leaf temperatures between warming and control conditions were assessed by Chi square (χ2) tests. Differences in the effect of warming and site on LT50, NT and FP were assessed by using Factorial ANOVAs as well (See details in Supplementary Table S2). Differences between NT and LT50 in determining freezing resistance mechanisms for each species and experimental conditions were assessed with t-tests. Data were checked for normality before analyses.
Results
Microclimatic Conditions
Air, soil and leaf temperatures during both growing seasons were affected by warming (Table 1). Although minimum air temperatures (Tmin) and the intensity of freezing events (Fint) were similar between +W and -W conditions (average Tmin ranged from -13 to 0.6°C and average Fint was -1.8°C in both growing periods), the frequency (Ffreq) and duration of freezing events (Fdur) tended to be lower and of shorter duration inside +W plots (Table 1). The maximum air temperature (Tmax) was consistently higher inside +W plots. For example, in 2014 +W increased 4.6 and 2.7°C the air Tmax in the sites 1 and 2, respectively. This Tmax increase was of 3.9, 2.9, and 3.3°C in the sites 1, 2, and 3 in 2015. Regarding the growing degree days above 0°C (GDD0), in 2014 GDD0 were 59.6 and 54.8% greater in +W than in –W conditions in the sites 1 and 2, respectively (Table 1). In 2015, +W increased GDD0 on average 37% in the three sites (Table 1).
Freezing Resistance in the Field Under Warming Scenario
Antarctic plant species exhibited different ranges of freezing resistance in the field (Supplementary Table S3). Considering all sites together, average of ice Nucleation Temperature (NT) and Freezing Point (FP) of C. quitensis were -3.7 ± 0.1°C and -2.2 ± 0.1°C, respectively (Figure 2). Freezing temperature producing 50% photoinactivation (LT50) ranged from -12.4 ± 0.1°C in site 2 to -17.4 ± 0.5°C in site 1 (Figure 2; F2,32 = 172.5, P < 0.0001). For D. antarctica, NT ranged from -3.6 ± 0.3°C in site 1 to -5.4 ± 0.3°C in sites 2 and 3 (Figure 3; F2,34 = 24.1, P < 0.0001). Similarly, FP ranged from -2.5 ± 0.3°C in site 1 to -4 ± 0.6°C in site 3 (Figure 3; F2,32 = 12.2, P < 0.001). In contrast, LT50 decreased (more negative) from -20.4 ± 0.8°C in site 3 to -24 ± 0.5°C in sites 1 and 2 (Figure 3; F2,34 = 22.7, P < 0.0001). It seems noteworthy that NT were sharply higher than LT50 on C. quitensis and D. antarctica at any site, suggesting that both species are able to tolerate ice formation within their leaf tissues. Thus, the mechanism of freezing resistance did not vary with site and was freezing tolerance for both C. quitensis and D. antarctica (Supplementary Table S3).
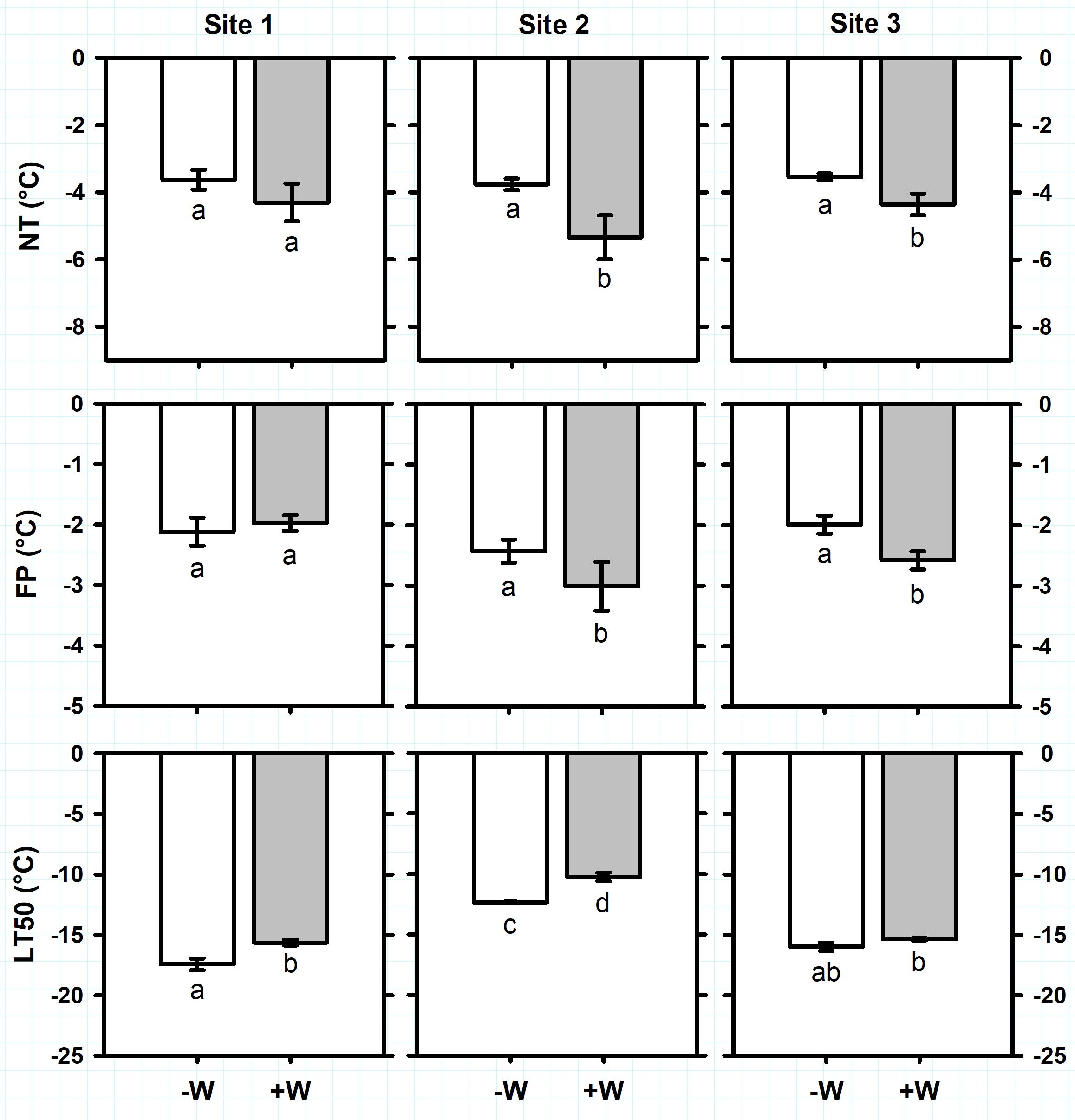
FIGURE 2. Freezing resistance of Colobanthus quitensis measured in plants growing in three sites in the King George Island. Freezing parameters measured were: NT, ice nucleation temperature (°C); FP, freezing point (°C); and LT50, freezing temperature producing 50% photoinactivation (°C). Values correspond to mean ± SE (n = 6–7). Treatments were: –W, plants under natural temperature conditions; +W, plants under warm temperature conditions. Less negative values indicate higher freezing resistance. Significant differences between natural and warm conditions are shown as different lowercases (P < 0.05).
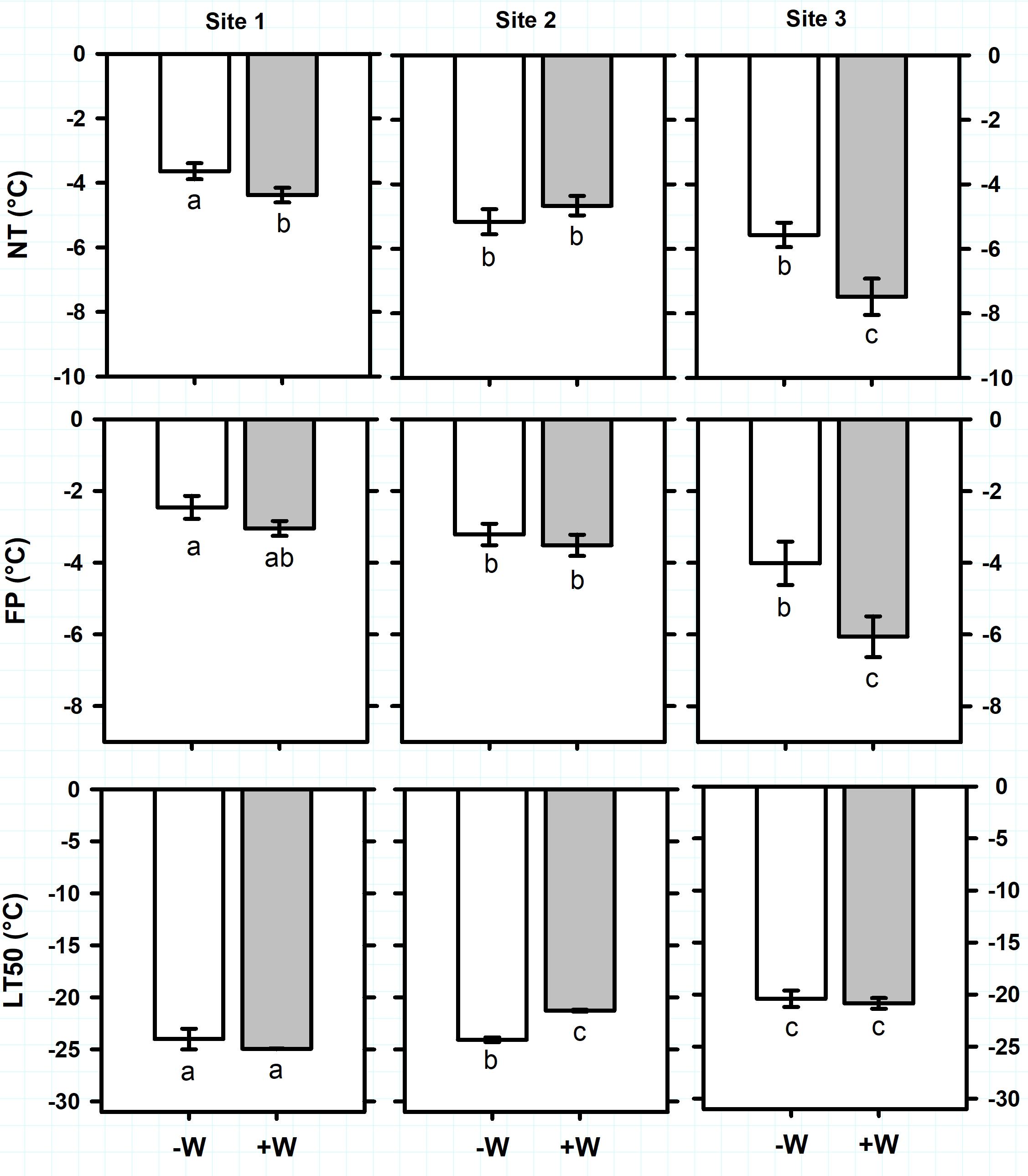
FIGURE 3. Freezing resistance of Deschampsia antarctica measured in plants growing in three sites in the King George Island. Freezing parameters measured were: NT, ice nucleation temperature (°C); FP, freezing point (°C); and LT50, freezing temperature producing 50% photoinactivation (°C). Values correspond to mean ± SE (n = 6–7). Treatments were: –W, plants under natural temperature conditions; +W, plants under warm temperature conditions. Less negative values indicate higher freezing resistance. Significant differences between natural and warm conditions are shown as different lowercases (P < 0.05).
Antarctic plants varied their vulnerability to suffer freezing damage with warming (Supplementary Table S3). For C. quitensis, warming (+W) increased (less negative temperature) on average 2K the LT50 of plants growing in sites 1 and 2 (Figure 2; F1,32 = 34, P < 0.0001). In contrast, +W decreased on average 1.2K the NT (F1,32 = 6.8, P = 0.014) and 0.6K the FP (F1,32 = 2.6, P = 0.012) of plants growing at sites 2 and 3 (Figure 2). For D. antarctica, +W effects on freezing resistance varied with site (Figure 3). For example, LT50 increased 2.8K with +W but only on plants from site 2 (F2,34 = 6.7, P = 0.003). In contrast, +W decreased 0.8 and 1.9 K the NT of plants growing in sites 1 and 3 (F1,34 = 5.4, P = 0.026), respectively, but no effects on NT were observed in plants at site 2 (Figure 3). Similarly, FP of D. antarctica inside +W occurred at temperatures 2K more negative than control plants but only at site 3 (F1,34 = 8.4, P = 0.007). Despite the opposite effects of warming on NT and LT50 of Antarctic plants, both C. quitensis and D. antarctica were always classified as freezing tolerant species (Supplementary Table S3).
Discussion
Sufficient levels of resistance to freezing temperatures during the summer is key for the survival, growth and reproduction of C. quitensis and D. antarctica in the Maritime Antarctica (Cavieres et al., 2016). Paradoxically, the regional warming that promote the growth and reproduction of these species (Cannone et al., 2016) could reduce their survival ability making even the best cold adapted plants more susceptible to damage by freezing temperatures (Woldendorp et al., 2008; Ball et al., 2011). As far as we are aware, our work is the first study reporting the level and mechanisms of freezing resistance of Antarctic vascular plants measured in situ, and we demonstrated that both plant species exhibited a great ability to cope with freezing temperatures during the growing season. Nonetheless, increases in the temperatures experienced by plants during the growing season changed this functional trait suggesting increases in their vulnerability to suffer freezing damage under warmer temperature scenarios.
Overall, warmer conditions decreased the freezing resistance of both Antarctic species. That is, LT50 occurred at higher (less negative) temperatures in warmed plants of C. quitensis and D. antarctica (Supplementary Table S3 and Figures 2, 3). The LT50 increased on average 2K for C. quitensis and 2.8K for D. antarctica, despite mean air temperatures increased only by 1K with OTCs. This suggests that further increases in ambient temperature, as those projected in future climate scenarios could lead to greater changes in this plant functional trait. Whilst LT50 increases in warmer conditions were observed at all sites for C. quitensis, for D. antarctica it showed site specific responses (Supplementary Tables S2, S3). These results highlight two aspects that have to be considered. First, C. quitensis would be more vulnerable to freezing damage than D. antarctica with warmer conditions. Several studies have reported that the Antarctic vascular plants respond differently to warmer conditions (Xiong et al., 2000; Cannone et al., 2016). For example, whereas in D. antarctica no effects of warming on leaf carbon gain and plant growth has been observed, in C. quitensis warmer temperatures promoted both plant traits (Sáez et al., 2018). Our microclimatic data showed that although freezing temperatures events were frequent and of long duration during the Antarctic summer, the intensity of those events were relatively mild (average of -2°C, with absolute records c. -7°C). Previous studies on the climate of the study area have reported summer freezing events of -5 and -7.8°C (Cygan, 1981; Araźny et al., 2013). However, air temperatures suddenly decrease in autumn, with some records of -13°C in April (Araźny et al., 2013). If we considered that snow duration and cover are highly unpredictable in the area because of topography and especially of wind speed (Angiel et al., 2010), Antarctic plants could be frequently exposed to such freezing temperatures during the summer- autumn transition. In that scenario, D. antarctica has a temperature safety margin of seldom 10K because warmed plants exhibited an averaged LT50 of -22.4°C. However, this safety margin doesn’t exist for C. quitensis, which exhibited an averaged LT50 of -13.8°C. Although, both species have the ability to cope with summer freezing events in the area, if ambient temperatures continue to rise, as some authors propose (Lee et al., 2017), they might have negative consequences for plant survival of C. quitensis but not for D. antarctica.
Secondly, a site-dependent response of D. antarctica to warming was observed, where warming increased LT50 only in plants from site 2. This site is dominated by moss carpets and presents a permanent water-saturated but well drained substrate, abundant in organic matter and N content of 18–40 ppm (Kozeretska et al., 2010; Supplementary Table S1). According to substrate preferences of this species, moss carpets is where this species is more abundant and frequent (Casanova-Katny and Cavieres, 2012; Park et al., 2013), and where individual plants grow bigger (Casanova-Katny and Cavieres, 2012; Cavieres et al., 2018). Given that this site presents a greater availability of resources (i.e., water and nutrients) compared to the other sites, and where the presence of neighbors (moss carpets) can ameliorates the harsh climatic conditions (see Casanova-Katny and Cavieres, 2012; Cavieres et al., 2018), the site-dependent LT50 response to warming of D. antarctica could be attributed to the tradeoff between plant growth and stress resistance, where warmer temperatures are favoring plant allocation to growth. The absence of better soils (site 3), the presence of sea spray and animal disturbance (site 1) and the absence of moss carpets (sites 1 and 3) generate that plants on these sites are constantly dealing with stress, even under warmer conditions.
Contrary to our expectations, NT and FP occurred at lower (more negative) temperatures in leaves of warmed plants of both species. FP and NT depend on specific properties of the plant tissues and may vary according to the cell sap concentration and/or the accumulation of water-binding substances inside the cell (Sakai and Larcher, 1987). NT decreases in plant tissues with small cell sizes, relative low water content, and/or little or no intercellular space for nucleation (Sakai and Larcher, 1987). Sáez et al. (2018) reported that in situ warmer temperatures induced changes in morpho-anatomical leaf traits of C. quitensis and D. antarctica that might relate with the changes in freezing resistance reported here, but further studies are needed to unveil their relation and consequences for the plant freezing resistance. In addition, several studies have reported that water-soluble carbohydrates depress FP, and their accumulation is positively related to abiotic stress survival, which is also the case of these two Antarctic plant species (Bravo et al., 2001; Pastorczyk et al., 2014). We expected that lower FP values contributed to increase the freezing resistance of Antarctic plants, by decreasing their LT50. However, this was not the case. This could be related with the fact that +W plants were exposed to warmed temperatures during days but similar cold temperatures as –W plants during the nights. This has two implications. First, plants under +W may be exposed to more frequent freeze/thaw events than plants under -W. This may cause recurrent xylem embolism and cell dehydration (Pearce, 2001), which may induce the warmed plants to keep some freezing avoidance capability such as lower NT and FP than unwarmed plants. It has been observed that the degree of frost hardening may be a function of the number of freezing events (Beck et al., 2004). Second, warmer days imply better conditions for CO2 assimilation in the +W treatment (e.g., Sáez et al., 2018) but similar respiration rates during the night on both +W or -W. Then, there is a higher probability that carbohydrates synthesized exceeded carbohydrates respired in +W than in -W treatment with the consequent higher accumulation of non-structural carbohydrate in +W, which can act as compatible solutes reducing the FP, but these putative explanations remain to be elucidated.
Previous studies have measured the freezing resistance of C. quitensis and/or D. antarctica of plants grown under controlled conditions in the lab. However, there were discrepancies in the level (i.e., LT50 values) and mechanism of freezing resistance (i.e., freezing avoidance or tolerance), as well as in their capacity for cold acclimation. For instance, Bravo et al. (2001) reported that C. quitensis avoided freezing by supercooling, that non-acclimated plants of C. quitensis experienced freezing injury at -4.8°C, when ice nucleation was induced by silver iodine, and its LT50 decreased only 1K after cold acclimation at 4°C for 21 days under the same measurement condition. In contrast, LT50 of C. quitensis decreased from -7 to -15°C with a similar cold-acclimation period without using ice nucleator according to Gianoli et al. (2004) and Reyes-Bahamonde (2013), and they classified C. quitensis as a freezing tolerant species (Table 2). In the case of D. antarctica, all previous studies classified it as a meanly freezing tolerant plant (Table 3). According to Bravo et al. (2001), cold-acclimation decreased LT50 from -12 to -26.6°C. However, Chew et al. (2012) found that LT50 of D. antarctica decreased from -12°C in non-acclimated to -17°C in cold-acclimated plants, whilst Reyes-Bahamonde (2013) found that this species exhibited a LT50 of -16.5 and -18.4°C in non- and cold-acclimated plants, respectively (Table 3).
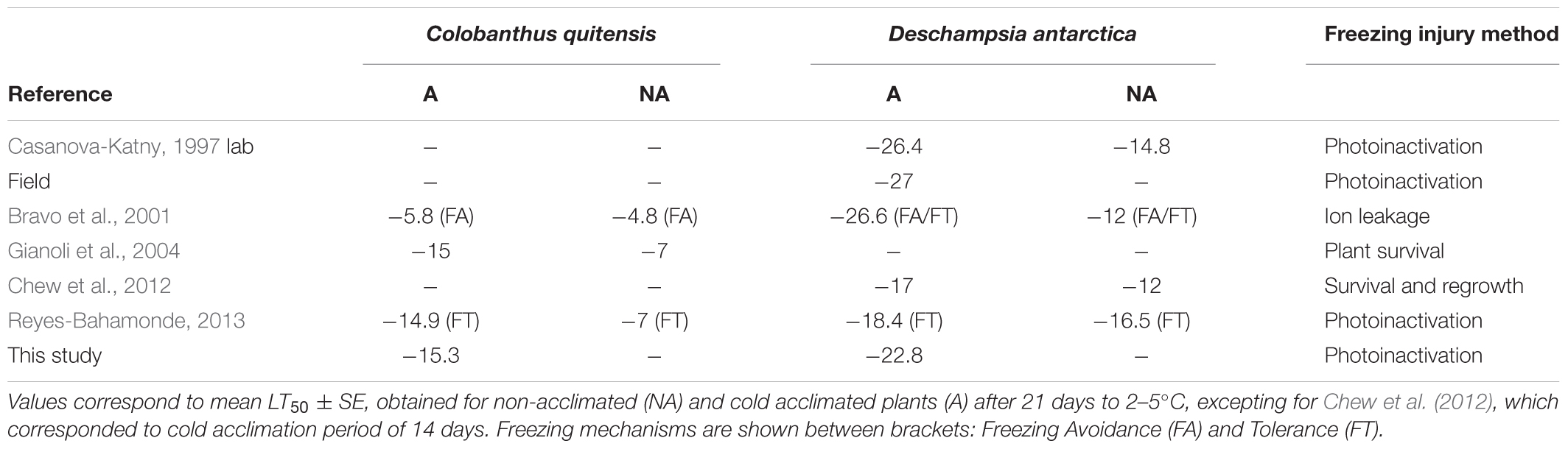
TABLE 2. Previous studies where freezing resistance of Antarctic plants has been reported.
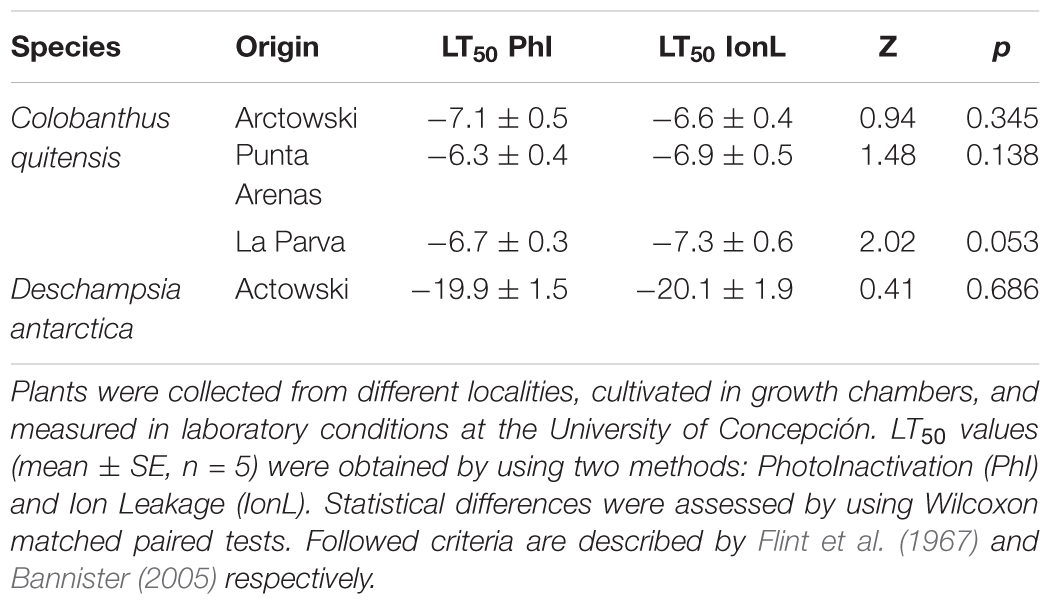
TABLE 3. A comparison of two criteria for LT50 determinations in Colobanthus quitensis and Deschampsia antarctica.
Considering all sites together our results showed an average LT50 of -15.3 and -22.8°C for C. quitensis and D. antarctica, respectively, and that both species exhibited freezing tolerance as the mechanism of freezing resistance. In the case of C. quitensis our LT50 are similar to those previously reported for cold-acclimated plants, which is reasonable considering temperature conditions recorded in the field when the determinations were carried out. For D. antarctica, however, LT50 were relatively different from previous reports. Likely explanations for the discrepancies in the level and mechanism of freezing resistance of Antarctic vascular plants found here with those previously reported arise from methodological differences among studies. For example the method used to assess freezing injury can lead to important differences. Bravo et al. (2001) estimated LT50 of C. quitensis and D. antarctica from electrolyte leakage by freezing-induced cell lysis, whereas Reyes-Bahamonde (2013) LT50 estimations were based on photoinactivation. On the contrary, Gianoli et al. (2004) calculated LT50 of C. quitensis with plant survival percentage, and Chew et al. (2012) LT50 estimations of D. antarctica were based on tiller survival from re-growth. It has been reported that photoinactivation method agrees very well with the results obtained by methods that directly measure plant tissue damage (i.e., survival, visual assessment of freezing injuries and vital staining; Boorse et al., 1998; Neuner and Buchner, 1999). Our estimations coincided with similar LT50 reported for C. quitensis with those methods (see Table 2 for references). The electrolyte leakage method can lead to confusing results, because on one hand, it can overestimate leaf damage given that cellular solutes other than electrolytes may be induced by freezing, and on the other hand, coriaceous leaves do not release electrolytes readily, which can lead to spurious estimates of LT50 using this method (Boorse et al., 1998; Bannister, 2007). However, Antarctic vascular plants exhibited similar LT50 regardless they were calculated by ion leakage or photoinactivation methods as it is shown in the Table 3. In the case of D. antarctica, our LT50 values were intermediate compared to previous studies. Nevertheless, this result have to be taken with caution as some replicates did not reach the 50 percent damage (i.e., six replicates in the site 1). Thus, average LT50 in the field could be even more negative than we reported. This point is consistent with Casanova-Katny (1997) who found that the LT50 of D. antarctica was below -27°C in the field.
As mentioned, the ambient temperatures experienced by plants affect their ability to resist freezing temperatures (Beck et al., 2004). Thus, a second methodological aspect that differed among studies and that could underlie the discrepancies in the freezing resistance of Antarctic species is the residence time and temperature used on growth chambers. For example, Bravo et al. (2001) and Gianoli et al. (2004) collected adult plants from Antarctica that were vegetative propagated at 15°C for a couple of years before the freezing determinations. Chew et al. (2012) obtained adult plants from seeds collected in the field, while Reyes-Bahamonde (2013) used plants grown at constant 11°C during 2 months after their collection in the field. However, although Bravo et al. (2001) and Gianoli et al. (2004) used similar plants residence time and growth temperature for determinations, they found different LT50 for C. quitensis. A similar situation occurred for D. antarctica (Table 2). Probably, multiple factors could influence the level of freezing resistance. Such discrepancies reinforce the importance of in situ determinations as we did here.
Finally, our results clearly showed that warmer temperatures affect the freezing resistance of Antarctic vascular plants. These results are in line with previous studies conducted in alpine and arctic plant where similar plant responses to warming (decreases in freezing resistance with warming) were reported (e.g., Loveys et al., 2006; Marchand et al., 2006; Woldendorp et al., 2008; Sierra-Almeida and Cavieres, 2010; Rixen et al., 2012). The ability to withstand freezing temperatures is a key feature of species inhabiting cold climates, hence if new warming phases occur in the Antarctic Peninsula due to climate change, the survival of C. quitensis could be threatened. Whilst D. antarctica seems to be unaffected by warming on this trait, other aspects of its biology could be altered by warming (e.g., increases in respiration). Nevertheless, more research is needed to unveil the likely consequences of global warming on plants from cold biomes where in situ determinations of plant freezing resistance are crucial to understand the physiological mechanisms underlying plant adaptations to current and future climatic scenario for the Antarctic in particular.
Author Contributions
AS-A conceived and designed the study, conducted the data collection, analyzed the data, and edited the manuscript. LC contributed to the study conception, designed and installed experimental setup and climatic sensors, and edited the manuscript. LB designed and installed experimental setup and climatic sensors, and contributed to the data analysis and manuscript approval.
Funding
This work was supported by PIA-Conicyt under grant ART-1102; MINECOM F ICM under grant P02-005; and CONICYT under PFB-23 supporting the IEB, and Fondecyt 11150710 (AS-A) 1151173 and NEXER-UFRO (LB).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Oscar Viel icebreaker crew, the Chilean Army, and Arctowski Station crew for their help and logistic support during 51th Antarctic Scientific Expedition (ECA) of the Chilean Antarctic Institute (INACH). We thank Ana Sanhueza for her help with climatic data analyses and Claudia Reyes for methodological comparisons for LT50 determinations.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01456/full#supplementary-material
References
Angiel, P. J., Potocki, M., and Biszczuk-Jakubowska, J. (2010). Weather condition characteristics at the H. Arctowski Station (South Shetlands, Antarctica) for 2006, in comparison with multi-year research results. Miscellanea Geogr. 14, 79–89.
Araźny, A., Kejna, M., and Sobota, I. (2013). Ground temperature at the Henryk Arctowski Station (King George Island, Antarctic) – case study from the period January 2012 to February 2013. Bull. Geogr. Phys. Geogr. Series 6, 59–80. doi: 10.2478/bgeo-2013-0004
Ball, M. C., Harris-Pascal, D., Egerton, J. J. G., and Lenné, T. (2011). “The paradoxical increase in freezing injury in a warming climate: frost as a driver of change in cold climate vegetation,” in Temperature Adaptation in a Changing Climate: Nature at Risk, eds K. B. Storey and K. K. Tanino (Wallingford:CAB International), 179–185.
Bannister, P. (2005). Frost resistance of the New Zealand narrow-leaved snow tussock grass, Chionochloa rigida. N. Zeal. J. Bot. 43, 425–430. doi: 10.1080/0028825X.2005.9512965
Bannister, P. (2007). A touch of frost? Cold hardiness of plants in the Southern Hemisphere. N. Zeal. J. Bot. 45, 1–33. doi: 10.1080/00288250709509700
Bannister, P., Maegli, T., Dickinson, K., Halloy, S., Knight, A., Lord, J., et al. (2005). Will loss of snow cover during climatic warming expose New Zealand alpine plants to increased frost damage? Oecologia 144, 245–256.
Beck, E. H., Heim, R., and Hansen, J. (2004). Plant resistance to cold stress: mechanisms and environmental signals triggering frost hardening and dehardening. J. Biosci. 29, 449–459. doi: 10.1007/BF02712118
Bokhorst, S., Huiskes, A. D., Aerts, R., Convey, P., Cooper, E. J., Dalen, L., et al. (2013). Variable temperature effects of open top chambers at polar and alpine sites explained by irradiance and snow depth. Glob. Change Biol. 19, 64–74. doi: 10.1111/gcb.12028
Boorse, G. C., Bosma, T. L., Meyer, A. C., Ewers, F. W., and Davis, S. D. (1998). Comparative methods of estimating freezing temperatures and freezing injury in leaves of chaparral Shrubs. Int. J. Plant Sci. 159, 513–521. doi: 10.1086/297568
Bravo, L. A., Bascuñán-Godoy, L., Pérez-Torres, E., and Corcuera, L. J. (2009). “Cold hardiness in Antarctic vascular plants,” in Plant Cold Hardiness: From the Laboratory to the Field, eds L. Gusta, M. Wisnewski, and K. Tanino (Wallingford: CAB International), 198–213. doi: 10.1079/9781845935139.0198
Bravo, L. A., and Griffith, M. (2005). Characterization of antifreeze activity in Antarctic plants. J. Exp. Bot. 56, 1089–1096. doi: 10.1093/jxb/eri112
Bravo, L. A., Ulloa, N., Zúñiga, G. E., Casanova, A., Corcuera, L. J., and Alberdi, M. (2001). Cold resistance in Antarctic Angiosperms. Physiol. Plant. 111, 55–65. doi: 10.1186/1471-2229-12-114
Briceño, V. F., Harris-Pascal, D., Nicotra, A. B., Williams, E., and Ball, M. C. (2014). Variation in snow cover drives differences in frost resistance in seedlings of the alpine herb Aciphylla glacialis. Environ. Exp. Bot. 106, 174–181. doi: 10.1016/j.envexpbot.2014.02.011
Cannone, N., Guglielmin, M., Convey, P., Worland, M. R., and Favero Longo, S. E. (2016). Vascular plant changes in extreme environments: effects of multiple drivers. Clim. Chang. 134, 651–665. doi: 10.1007/s10584-015-1551-7
Casanova-Katny, M. A. (1997). Eficiencia Fotoquímica del PSII en Deschampsia antarctica (Desv.): una Gramínea Tolerante a la Congelación. Master Thesis: Valdivia, Universidad Austral de Chile.
Casanova-Katny, M. A., and Cavieres, L. A. (2012). Antarctic moss carpets facilitate growth of Deschampsia antarctica but not its survival. Pol. Biol. 35, 1869–1878. doi: 10.1007/s00300-012-1229-9
Cavieres, L. A., Sáez, P., Sanhueza, C., Sierra-Almeida, A., Rabert, C., Corcuera, L. J., et al. (2016). Ecophysiological traits of Antarctic vascular plants: their importance in the responses to climate change. Plant Ecol. 217, 343–358. doi: 10.1007/s11258-016-0585-x
Cavieres, L. A., Vivas, M., Mihoc, M. A. K., Osses, D. A., Ortiz-Gutiérrez, J. M., Sáez, P., et al. (2018). The importance of facilitative interactions on the performance of Colobanthus quitensis in an Antarctic tundra. J. Veg. Sci. 29, 236–244. doi: 10.1111/jvs.12616
Chew, O., Lelean, S., John, U. P., and Spangenberg, G. C. (2012). Cold acclimation induces rapid and dynamic changes in freeze tolerance mechanisms in the cryophile Deschampsia antarctica E. Desv. Plant Cell. Environ. 35, 829–837. doi: 10.1111/j.1365-3040.2011.02456.x
Convey, P. (1996). The influence of environmental characteristics on life history attributes of Antarctic terrestrial biota. Biol. Rev. 71, 191–225. doi: 10.1111/j.1469-185X.1996.tb00747.x
Convey, P. (2001). “Terrestrial ecosystem response to climate changes in the Antarctic,” in Fingerprints of Climate Change - Adapted Behaviour and Shifting Species Ranges, eds G. R. Walther, C. A. Burga, and P. J. Edwards (New York, NY: Kluwer), 17–42.
Convey, P. (2006). Antarctic terrestrial ecosystems: responses to environmental changes. Polarforsch 75, 101–111.
Convey, P. (2013). “Antarctic ecosystems,” in Encyclopedia of Biodiversity, 2nd Edn, ed. S. A. Levin (San Diego: Elsevier), 179–188. doi: 10.1016/B978-0-12-384719-5.00264-1
Convey, P., Chown, S. L., Clarke, A., Barnes, D. K., Bokhorst, S., Cummings, V., et al. (2014). The spatial structure of Antarctic biodiversity. Ecol. Monogr. 84, 203–244. doi: 10.1890/12-2216.1
Cygan, B. (1981). Characteristics of meteorological conditions at the Arctowski Station during the summer season of 1979-1980. Pol. Polar Res. 2, 35–46.
De Boeck, H., De Groote, T., and Nijs, I. (2012). Leaf temperatures in glasshouses and open-top chambers. New Phytol. 194, 1155–1164. doi: 10.1111/j.1469-8137.2012.04117.x
Flint, H. L., Boyce, B. R., and Beattie, D. J. (1967). Index of injury is a useful expression of freezing injury to plant tissues as determined by the electrolytic method. Can. J. Plant Sci. 47, 229–230. doi: 10.4141/cjps67-043
Fowbert, J. A., and Smith, R. I. L. (1994). Rapid population increases in native vascular plants in the Argentine Island. Antarctic Peninsula. Arct. Antarct. Alp. Res. 26, 290–296. doi: 10.2307/1551941
Gerighausen, U., Bräutigam, K., Mustafa, O., and Peter, H. U. (2003). “Expansion of vascular plants on an Antarctic Island: a consequence of climate change?,” in Antarctic Biology in a Global Context, eds A. H. L. Huiskes, W. W. C. Gieskes, J. Rosema, R. M. L. Schorno, S. M. Van der Vies, and W. J. Wolff (Leiden: Backhuys), 79–83.
Gianoli, E., Inostroza, P., Zúñiga-Feest, A., Reyes-Díaz, M., Cavieres, L. A., Bravo, L. A., et al. (2004). Ecotypic differentiation in morphology and cold resistance in populations of Colobanthus quitensis (Caryophyllaceae) from the Andes of Central Chile and the Maritime Antarctic. Arct. Antarct. Alp. Res. 36, 484–489. doi: 10.1657/1523-0430 (2004)036[0484:EDIMAC]2.0.CO;2
Green, T. G. A., Schroeter, B., and Sancho, L. G. (2007). “Plant Life in Antarctica,” in Functional Plant Ecology, 2nd Edn, eds F. I. Pugnaire and F. Valladares (Boca Raton, FL: CRC Press), 389–455.
Greene, D. M., and Holtom, A. (1971). Studies in Colobanthus quitensis (Kunth) Bartl. and Deschampsia antarctica Desv. III. Distribution, habitats and performance in the Antarctic botanical zone. Brit. Antarct. Surv. B 26, 1–29.
Guy, C. (2003). Freezing tolerance of plants: current understanding and selected emerging concepts. Can. J. Bot. 8, 1216–1223. doi: 10.1139/b03-130
Hekneby, M., Antolín, M. C., and Sánchez-Díaz, M. (2006). Frost resistance and biochemical changes during cold acclimation in different annual legumes. Environ. Exp. Bot. 55, 305–314. doi: 10.1016/j.envexpbot.2004.11.010
Hollister, R. D., and Webber, P. J. (2000). Biotic validation of small open-top chambers in a tundra ecosystem. Glob. Change Biol. 6, 835–842. doi: 10.1046/j.1365-2486.2000.00363.x
Inouye, D. W. (2000). The ecological and evolutionary significance of frost in the context of climate change. Ecol. Lett. 3, 457–463. doi: 10.1046/j.1461-0248.2000.00165.x
Kennedy, A. D. (1995). Simulated climate-change- are passive greenhouses a valid microcosm for testing biological effects of environmental perturbations. Glob. Change Biol. 1, 29–42. doi: 10.1111/j.1365-2486.1995.tb00004.x
Körner, C. (2011). Coldest places on earth with angiosperm plant life. Alp. Bot. 121, 11–22. doi: 10.1007/s00035-011-0089-1
Kozeretska, I. A., Parnikoza, I. Y., Mustafa, O., Tyschenko, O. V., Korsun, S. G., and Convey, P. (2010). Development of Antarctic herb tundra vegetation near Arctowski station, King George Island. Polar Sci. 3, 254–261. doi: 10.1016/j.polar.2009.10.001
Ladinig, U., Hacker, J., Neuner, G., and Wagner, J. (2013). How endangered is sexual reproduction of high-mountain plants by summer frosts? Frost resistance, frequency of frost events and risk assessment. Oecologia 171, 743–760. doi: 10.1007/s00442-012-2581-8
Larcher, W. (2000). Temperature stress and survival ability of Mediterranean sclerophyllous plants. Plant Biosys. 134, 279–295. doi: 10.1080/11263500012331350455
Larcher, W. (2003). Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups. Berlin: Springer.
Larcher, W., Kainmüller, C., and Wagner, J. (2010). Survival types of high mountain plants under extreme temperatures. Flora 205, 3–18. doi: 10.1016/j.flora.2008.12.005
Lee, J. R., Raymond, B., Bracegirdle, T. J., Chades, I., Fuller, R. A., Shaw, J. D., et al. (2017). Climate change drives expansion of Antarctic ice-free habitat. Nature 547, 49–54. doi: 10.1038/nature22996
Loveys, B. R., Egerton, J. J. G., and Ball, M. C. (2006). Higher daytime leaf temperatures contribute to lower freeze tolerance under elevated CO2. Plant Cell Environ. 29, 1077–1086. doi: 10.1111/j.1365-3040.2005.01482.x
Marchand, F. L., Kockelbergh, F., van de Vijver, B., Beyens, L., and Nijs, I. (2006). Are heat and cold resistance of arctic species affected by successive extreme temperature events? New Phytol. 170, 291–300.
McMaster, G. S., and Wilhelm, W. W. (1997). Growing degree-days: one equation, two interpretations. Agric. For. Meteorol. 87, 291–300. doi: 10.1016/S0168-1923(97)00027-0
Moore, D. M. (1970). Studies in Colobanthus quitensis (Kunth) Bartl. and Deschampsia antarctica Desv.: II. Taxonomy, distribution and relationships. Br. Antarct. Surv. Bull. 23, 63–80.
Moore, D. M. (1979). “Southern ocean heaths (incl. Magellanic Moorland),” in Ecosystems of the World: Heathland and Related Shrublands, ed. R. L. Specht (Amsterdam: Elsevier), 490–497.
Neuner, G., and Buchner, O. (1999). Assessment of foliar frost damage: a comparison of in vivo chlorophyll fluorescence with other viability tests. J. Appl. Bot. 73, 50–54.
Olave-Concha, N., Bravo, L. A., Ruiz-Lara, S., and Corcuera, L. J. (2005). Differential accumulation of dehydrin-like proteins by abiotic stresses | in Deschampsia antarctica Desv. Pol. Biol. 28, 506–513. doi: 10.1007/s00300-005-0718-5
Park, J. S., Ahn, I. Y., and Lee, E. J. (2013). Spatial distribution patterns of the Antarctic Hair grass Deschampsia antarctica in relation to environmental variables on Barton Peninsula, King George Island. Arct. Antarct. Alp. Res. 45, 563–574. doi: 10.1657/1938-4246-45.4.563
Pastorczyk, M., Giełwanowska, I., and Lahuta, L. B. (2014). Changes in soluble carbohydrates in polar Caryophyllaceae and Poaceae plants in response to chilling. Acta Physiol. Plant. 36, 1771–1780. doi: 10.1007/s11738-014-1551-7
Pescador, D. S., Sánchez, A. M., Luzuriaga, A. L., Sierra-Almeida, A., and Escudero, A. (2018). Winter is coming: plant freezing resistance as a key functional trait for the assembly of annual Mediterranean communities. Ann. Bot. 121, 335–344. doi: 10.1093/aob/mcx166
Piotrowicz-Cieślak, A. I., Gielwanowska, I., Bochenek, A., Loro, P., and Górecki, R. J. (2005). Carbohydrates in Colobanthus quitensis and Deschampsia antarctica. Acta Soc. Bot. Pol. 74, 209–217. doi: 10.5586/asbp.2005.027
Reyes-Bahamonde, C. (2013). Consecuencias del aumento de la temperatura y la sequía en la Resistencia al Congelamiento de Deschampsia antarctica Desv. (Poaceae) and Colobanthus quitensis (Kunth.) Bartl. (Caryophyllaceae). U.G. Thesis, Universidad de Concepción, Concepción (Chile).
Rixen, C., Dawes, M. A., Wipf, S., and Hagedorn, F. (2012). Evidence of enhanced freezing damage in treeline plants during six years of CO2 enrichment and soil warming. Oikos 121, 1532–1543. doi: 10.1111/j.1600-0706.2011.20031.x
Robinson, S. A., Wasley, J., and Tobin, A. K. (2003). Living on the edge – plants and global change in continental and maritime Antarctica. Glob. Change Biol. 9, 1681–1717. doi: 10.1046/j.1365-2486.2003.00693.x
Sáez, P. L., Cavieres, L. A., Galmés, J., Gil-Pelegrin, E., Peguero-Pina, J. J., Sancho-Knapik, D., et al. (2018). In situ warming in the Antarctic: effects on growth and photosynthesis in Antarctic vascular plants. New Phytol. 218, 1406–1418. doi: 10.1111/nph.15124
Sakai, A., and Larcher, W. (1987). Frost Survival of Plants. Responses and Adaptation to Freezing Stress. Berlin: Springer. doi: 10.1007/978-3-642-71745-1
Sierra-Almeida, A., and Cavieres, L. A. (2010). Summer freezing resistance decreased in high-elevation plants exposed to experimental warming in the central Chilean Andes. Oecologia 163, 267–276. doi: 10.1007/s00442-010-1592-6
Smith, R. I. L. (2003). “The enigma of Colobanthus quitensis and Deschampsia antarctica in Antarctica,” in Antarctic Biology in a Global Context, eds A. H. L. Huiskes, W. W. C. Gieskes, J. Rosema, R. M. L. Schorno, S. M. Van der Vies, and W. J. Wolff (Leiden: Backhuys), 234–239.
Smith, R. I. L., and Poncet, S. (1987). Deschampsia antarctica and Colobanthus quitensis in the Terra Firma Islands. Br. Antarct. Sur. Bull. 74, 31–35.
Torres-Mellado, G. A., Jaña, R., and Casanova-Katny, M. A. (2011). Antarctic hairgrass expansion in the South Shetland archipelago and Antarctic Peninsula revisited. Polar Biol. 34, 1679–1688. doi: 10.1007/s00300-011-1099-6
Turner, J., Barrand, N., Bracegirdle, T., Convey, P., Hodgson, D., Jarvis, M., et al. (2014). Antarctic climate change and the environment: an update. Polar Rec. 50, 237–259. doi: 10.1039/c6pp90004f
Turner, J., Lu, H., White, I., King, J. C., Phillips, T., Hosking, J. S., et al. (2016). Absence of 21st century warming on Antarctic Peninsula consistent with natural variability. Nature 535, 411–423. doi: 10.1038/nature18645
Vaughan, D., Marshall, G., Connelley, W., Parkinson, C., Mulvaney, R., Hodgson, D., et al. (2003). Recent rapid regional climate warming on the Antarctic Peninsula. Clim. Chang. 60, 243–274. doi: 10.1023/A:1026021217991
Wasley, J., Robinson, S. A., Lovelock, C. E., and Popp, M. (2006). Climate change manipulations show Antarctic flora is more strongly affected by elevated nutrients than water. Glob. Change Biol. 12, 1800–1812. doi: 10.1111/j.1365-2486.2006.01209.x
Wisniewski, M., Gusta, L., and Neuner, G. (2014). Adaptative mechanisms of freeze avoidance in plants: a brief update. Environ. Exp. Bot. 99, 133–140. doi: 10.1016/j.envexpbot.2013.11.011
Woldendorp, G., Hill, M. J., Doran, R., and Ball, M. C. (2008). Frost in a future climate: modelling interactive effects of warmer temperatures and rising atmospheric [CO2] on the incidence and severity of frost damage in a temperate evergreen (Eucalyptus pauciflora). Glob. Change Biol. 14, 294–308. doi: 10.1111/j.1365-2486.2007.01499.x
Xiong, F. S., Mueller, E. C., and Day, T. A. (2000). Photosynthetic and respiratory acclimation and growth response of Antarctic vascular plants to contrasting temperatures regimes. Am. J. Bot. 87, 700–710. doi: 10.2307/2656856
Yergeau, E., Bokhorst, S., Kang, S., Zhou, J., Greer, C. W., Aerts, R., et al. (2012). Shifts in soil microorganisms in response to warming are consistent across a range of Antarctic environments. Int. Soc. Microb. Ecol. 6, 692–702. doi: 10.1038/ismej.2011.124
Zúñiga-Feest, A., Bascuñán-Godoy, L., Reyes-Diaz, M., Bravo, L. A., and Corcuera, L. J. (2009). Is survival after ice encasement related with sugar distribution in organs of the Antarctic plants Deschampsia antarctica Desv. (Poaceae) and Colobanthus quitensis (Kunth) Bartl. (Caryophyllaceae)? Polar Biol. 32, 583–591. doi: 10.1007/s00300-008-0553-6
Keywords: Antarctica, climate change, Colobanthus quitensis, Deschampsia antarctica, freezing events, LT50, photoinactivation, warming
Citation: Sierra-Almeida A, Cavieres LA and Bravo LA (2018) Warmer Temperatures Affect the in situ Freezing Resistance of the Antarctic Vascular Plants. Front. Plant Sci. 9:1456. doi: 10.3389/fpls.2018.01456
Received: 01 June 2018; Accepted: 12 September 2018;
Published: 08 October 2018.
Edited by:
Hans J. De Boeck, University of Antwerp, BelgiumReviewed by:
Charles L. Guy, University of Florida, United StatesAud Helen Halbritter, University of Bergen, Norway
Copyright © 2018 Sierra-Almeida, Cavieres and Bravo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Angela Sierra-Almeida, YW5nZWxhc2llcnJhQHVkZWMuY2w=