 Miriam Szurman-Zubrzycka1†
Miriam Szurman-Zubrzycka1† Brygida Baran1†
Brygida Baran1† Magdalena Stolarek-Januszkiewicz1
Magdalena Stolarek-Januszkiewicz1 Jolanta Kwaśniewska2
Jolanta Kwaśniewska2 Iwona Szarejko1
Iwona Szarejko1 Damian Gruszka1*
Damian Gruszka1*- 1Department of Genetics, Faculty of Biology and Environment Protection, The University of Silesia in Katowice, Katowice, Poland
- 2Department of Plant Anatomy and Cytology, Faculty of Biology and Environment Protection, The University of Silesia in Katowice, Katowice, Poland
Meiosis is a process of essential importance for sexual reproduction, as it leads to production of gametes. The recombination event (crossing-over) generates genetic variation by introducing new combination of alleles. The first step of crossing-over is introduction of a targeted double-strand break (DSB) in DNA. DMC1 (Disrupted Meiotic cDNA1) is a recombinase that is specific only for cells undergoing meiosis and takes part in repair of such DSBs by searching and invading homologous sequences that are subsequently used as a template for the repair process. Although role of the DMC1 gene has been validated in Arabidopsis thaliana, a functional analysis of its homolog in barley, a crop species of significant importance in agriculture, has never been performed. Here, we describe the identification of barley mutants carrying substitutions in the HvDMC1 gene. We performed mutational screening using TILLING (Targeting Induced Local Lesions IN Genomes) strategy and the barley TILLING population, HorTILLUS, developed after double-treatment of spring barley cultivar ‘Sebastian’ with sodium azide and N-methyl-N-nitrosourea. One of the identified alleles, dmc1.c, was found independently in two different M2 plants. The G2571A mutation identified in this allele leads to a substitution of the highly conserved amino acid (arginine-183 to lysine) in the DMC1 protein sequence. Two mutant lines carrying the same dmc1.c allele show similar disturbances during meiosis. The chromosomal aberrations included anaphase bridges and chromosome fragments in anaphase/telophase I and anaphase/telophase II, as well as micronuclei in tetrads. Moreover, atypical tetrads containing three or five cells were observed. A highly increased frequency of all chromosome aberrations during meiosis have been observed in the dmc1.c mutants compared to parental variety. The results indicated that DMC1 is required for the DSB repair, crossing-over and proper chromosome disjunction during meiosis in barley.
Introduction
Meiosis is a process of essential significance for sexual reproduction. During meiosis distribution of genetic material to gametes is associated with recombination which is achieved through crossing over and chromosome segregation. The recombination event (crossing-over) takes place during the first meiotic prophase between non-sister chromatids of homologous chromosomes. It leads to establishing physical links between homologous chromosomes, called chiasmata. Meiotic crossing-over shuffles genetic information, creates new combinations of alleles, and therefore generates genetic variations and drives evolution. The process of recombination during meiosis starts with a programmed DNA double-strand break (DSB). Meiotic DSBs are introduced through the catalytic action of the evolutionarily-conserved SPO11 (Sporulation Protein 11) protein complex which is an enzyme related to type II DNA topoisomerases (Keeney et al., 1997; Robert et al., 2016; Vrielynck et al., 2016). In general, DSBs can be repaired through two major pathways: homologous recombination (HR) and/or non-homologous end joining (NHEJ) (Ohnishi et al., 2009). Programmed DSBs during meiosis are eliminated by HR in the DSBR pathway (Double Strand Break Repair model). The model of DSB repair was first proposed by Szostak and coworkers in the 1980s (Szostak et al., 1983). The extensive studies of this process in Saccharomyces cerevisiae implemented only several alterations to the original model (reviewed in Andersen and Sekelsky, 2010). After introduction of DSB, the DNA ends are resected and long (about 1 kbp) 3′single-stranded overhangs are created, called 3′ssDNA tails (Ohnishi et al., 2009). RAD51 (Radiation sensitive 51) and DMC1 (Disrupted Meiotic cDNA1) recombinases attach to these tails and form nucleoprotein filaments that search for and invade homologous sequences either on a sister chromatid or on a homologous chromosome (Bishop et al., 1992; Shinohara et al., 1992). The latter case may lead to genetic recombination. After invasion on homologous sequence, the next step in the DSBR pathway is establishing the D-loop structure followed by formation of a double Holliday Junction (dHJ) intermediate. Then, the two strands at each HJ are nicked by specific enzymes and ligated. The resolution of dHJ can result in both, crossover and non-crossover repair products (COs and NCOs, respectively) (Andersen and Sekelsky, 2010). DMC1 and RAD51 belong to the same protein family of recombinases, involved in DNA repair through HR, which are related to the bacterial RecA (Bianco et al., 1998). They catalyze the process of pairing and invasion of 3′ssDNA tails formed at the DSB sites into homologous double-stranded DNA. Both of these proteins take part in the meiotic recombination events, however, DMC1 is specific only for cells undergoing meiosis, while RAD51 is ubiquitous and acts also in DSB repair in somatic cells. It is suggested that DMC1 promotes only the CO recombination with the homologous chromosome, which is unique to meiosis, and RAD51 plays its role mainly in sister chromatid exchange or the NCO recombination (Shinohara and Shinohara, 2004; Neale and Keeney, 2006). However, a recent work has shown that in the case of absence of the RAD51-mediated strand exchange activity, the DMC1 activity is sufficient to repair all DSBs during meiosis into both CO and NCO products and it does not affect meiotic crossing-over rates or patterns (Cloud et al., 2012; Da Ines et al., 2013; Singh et al., 2017).
In the plant kingdom, meiosis has been studied to the greatest degree in Arabidopsis thaliana (for review see Mercier et al., 2015). Cereals with large genomes and large chromosomes, such as barley (Hordeum vulgare), are characterized by highly skewed distribution of meiotic crossovers. Consequently, the large sub-centromeric regions, representing substantial proportions of the physical map, are seldom recombined (Higgins et al., 2012; Ramsay et al., 2014). Therefore, the molecular mechanisms underlying meiotic events may be distinct for model Arabidopsis with genome size of ∼135 Mbp (The Arabidopsis Genome Initiative [AGI], 2000) contained within five chromosomes and for barley with genome size of ∼5.3 Gbp contained within seven chromosomes (International Barley Genome Sequencing Consortium Mayer et al., 2012; Mascher et al., 2017). Our knowledge on the DMC1 function in plants comes mainly from studies performed in Arabidopsis. Moreover, its detailed function in DSB repair during meiosis is still extensively discussed. For example, some contradictory reports have appeared in rice (Oryza sativa L.): one, showing that OsDMC1 is required for homologous pairing (Deng and Wang, 2007), and the other, reporting that it is dispensable in this process (Wang et al., 2016), which is different from the role of DMC1 described in other species. These results imply that the function of DMC1 may be distinct in diverse organisms and a direct transfer of knowledge from related species may not be feasible. The recent findings in rice have been obtained studying rice insertion mutants (Wang et al., 2016). Although some in silico studies of DMC1 have been performed in monocot crops, including barley (Barakate et al., 2014), only very recently role of the barley homolog was analyzed in a spontaneous mutant (Colas et al., 2019).
Barley (Hordeum vulgare L.), ranking fourth in production and acreage, belongs to the most important cereal crops worldwide. Here, we present the identification of barley mutants in the DMC1 gene isolated using TILLING strategy in the HorTILLUS population derived from chemical mutagenesis of spring cultivar ‘Sebastian’. Cytological analysis of male meiocytes in the identified dmc1 mutants revealed various abnormalities during meiosis, in anaphase/telophase I and anaphase/telophase II, as well in tetrads. Our results indicate that DMC1 is involved in the DSB repair, crossing-over and chromosome disjunction during meiosis process in barley.
Materials and Methods
Plant Material
The HorTILLUS (Hordeum vulgare – TILLING – University of Silesia) population has been used for mutation detection in the HvDMC1 gene through TILLING approach. This population was developed after double treatment of spring barley cultivar ‘Sebastian’ with sodium azide and N-methyl-N-nitrosourea (Szurman-Zubrzycka et al., 2018). Each M2 plant of the HorTILLUS population originated from a different M1 plant. Eight-fold DNA pools from M2 HorTILLUS plants served as templates for mutational screening. The homozygous lines of the isolated dmc1 mutants were backcrossed with their parent variety and homozygous mutants selected from the F2 populations have been used for cytological analyses of meiosis. Barley cv. ‘Sebastian’ has been used as a wild type in this study.
Mutational Screening in HvDMC1 Using the TILLING Strategy
The sequence of the DMC1 gene in barley was identified and published by Klimyuk et al. (2000) in the NCBI database (Acc. no. AF234170.1). Its genomic and coding sequences consist of 5654 bp and 1035 bp, respectively. The HvDMC1 gene is composed of 14 exons and encodes a protein which is 344 amino acid in length (Figure 1). Our bioinformatics analysis revealed that HvDMC1 gene has no paralogs in barley genome (Supplementary Materials 1, 2). The DMC1 sequence is strongly conserved among various species representing the plant and animal kingdom. The bioinformatics tools: ClustalOmega1 and CODDLE (Codons Optimized to Discover Deleterious Lesions) were used to select fragment of the HvDMC1 gene for mutational screening. This in silico analysis enabled selection of the gene fragment which is highly conserved among homologous sequences from different plant species (Figure 2). Sequence encoding the Rad51 functional domain, which is characteristic for proteins involved in the DNA repair, was mapped in the HvDMC1 gene with the use of Pfam tool2. Based on these bioinformatics analyses, the 811 bp long fragment of the HvDMC1 gene containing exons 7 to 11, encoding a part of the Rad51 domain, was chosen as an amplicon for the TILLING screening. PCR reaction was optimized for specific primers labeled with IRDye-700 (forward) and IRDye-800 (reverse) (Supplementary Material 3). TILLING was performed on DNA of 5,376 M2 plants of the HorTILLUS population. The method of mutational screening applied in this study was performed according to the protocol described elsewhere (Szurman-Zubrzycka et al., 2017; Jost et al., 2019). Briefly, the eight-fold pools were used for PCR reaction with IRDye-700 and IRDye-800 labeled and unlabeled primers (Supplementary Material 3). The next step, formation of heteroduplexes, was performed at 95°C for 3 min for initial denaturation, and then at 70°C for 20 sec (×70 cycles, -0.1°C per cycle) for slow renaturation. Heteroduplexes appeared only in pools with mutations within the analyzed amplicon. After heteroduplex formation the samples were treated with 20 μl of 0.1× Celery Juice Extract (CJE) containing Cel I enzyme that specifically recognizes and cuts DNA mismatches. The enzymatic cleavage was performed at 45°C for 15 min. The products of cleavage were purified with 96% ethanol with 1% sodium acetate and then washed with 70% ethanol. After centrifugation the pellets were dried and dissolved in 3 μl of STOP buffer (containing 5% bromophenol blue-xylene, 40% formamide and 1% EDTA). Before loading on polyacrylamide gel the samples were denatured. The electrophoresis was carried out in LI-COR sequencers in denaturing 6% polyacrylamide gels in 1xTBE (Tris – Boric Acid – EDTA) running buffer at the following settings: 3000 V, 30 mA and 30 W. The lanes with additional bands indicating putative mutations in the analyzed bulks were selected for further analysis (Supplementary Material 4). For identification of single plants carrying the mutations, each sample from the selected bulk was then analyzed by mixing its DNA individually with DNA of the parent variety following the same method described for the eight-fold pools. The analyzed fragments from the identified plants were sequenced in order to confirm the presence of mutations.
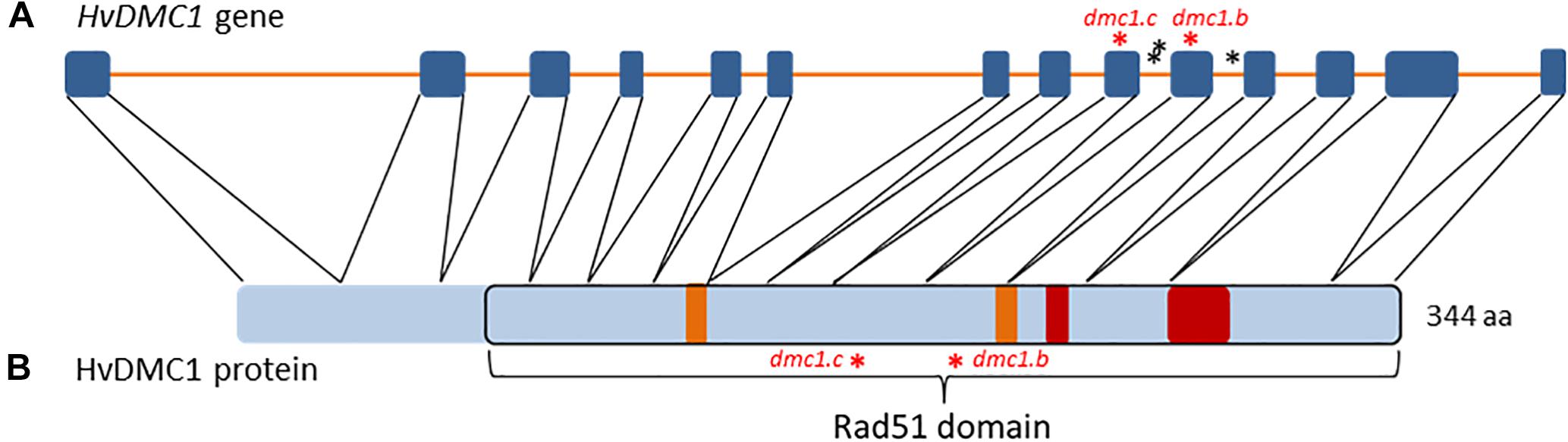
Figure 1. (A) The structure of the HvDMC1 gene. Blue boxes symbolize exons and orange lines introns. The identified mutations are indicated by asterisks (red – missense mutations, black – mutations in non-coding regions). (B) The structure of the HvDMC1 protein with localization of the Rad51 domain. Orange boxes symbolize the Walker A and Walker B functional motifs that are responsible for ATP binding, red boxes symbolize ssDNA-binding loops 1 and 2. The positions of amino-acid residues mutated in the dmc1.c and dmc1.b alleles are indicated by red asterisks.
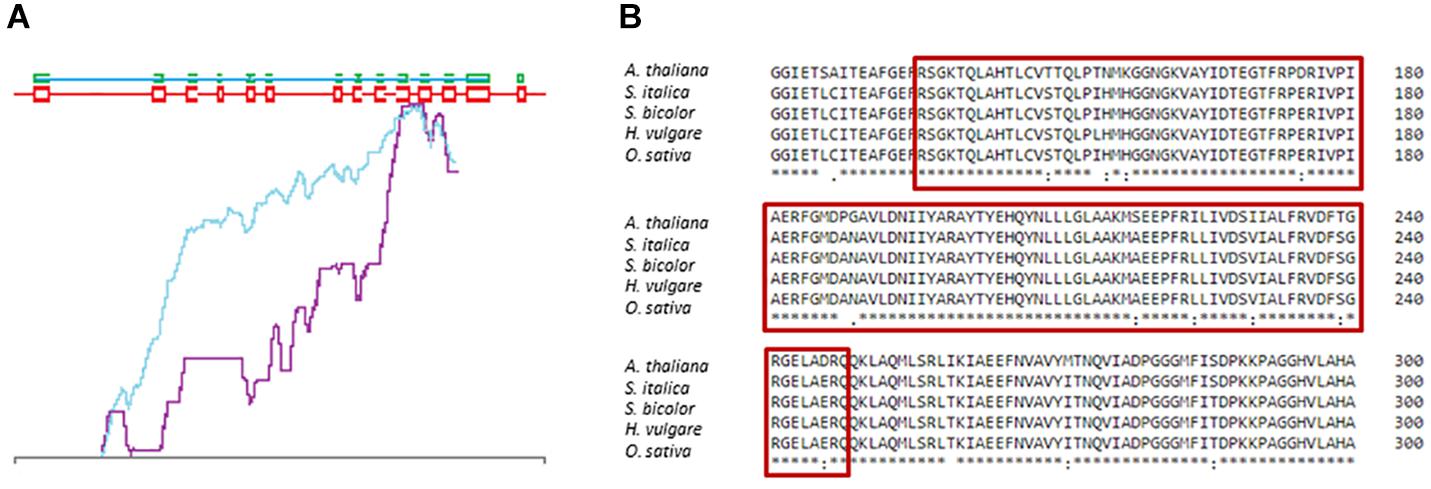
Figure 2. Selection of the HvDMC1 gene fragment for TILLING analysis (A) CODDLE analysis showing the most conserved fragment of the gene (blue line – score missense changes and truncations; violet – the PSSM difference). (B) Comparison of the DMC1 protein sequences from various plant species. The red frame shows conserved fragment of the protein which is encoded by the analyzed fragment of the HvDMC1 gene.
Preparation of Material for Microscopic Analyses of Meiosis
Plants of the dmc1 mutants as well as their parental cultivar ‘Sebastian’ were grown in a greenhouse at 22/20°C (day/night, respectively), under a photoperiod of 16 h/8 h and a light intensity of 400 μE/m2s for approximately 4 months, until their spikes reached length of 2.5–5 cm. Immature ears were harvested and immediately fixed in the methanol:acetic acid (3:1, v/v) overnight at room temperature.
To investigate the involvement of HvDMC1 in the DSB repair during meiosis, anthers from immature florets were used for preparation of male meiocyte spreads. Only cells in meiotic phases after crossing over were analyzed. Particular focus was given on cells in anaphase I/II and telophase I/II as well as tetrads, in which it was possible to observe micronuclei.
Cytogenetic slides were prepared using the Feulgen’s squash technique. Three anthers were isolated to prepare one slide. Cytological analyses were performed for each genotype in three repetitions with 15 slides per replica. The frequencies of anaphase/telophase I and anaphase/telophase II cells with chromosome aberrations were analyzed, on average, in 158 and 165 cells per slide, respectively. The frequencies of cells in tetrad stage with the micronuclei were estimated, based on analysis of, on average, 170 cells per slide. Preparations were examined with the Nikon ECLIPSE Ni bright field microscope. Images were captured by the Nikon DS.- Fi1c camera under 40× magnification.
Results
Mutation Identification and Characterization
After screening of 5,376 M2 plants of the HorTILLUS population, six independent mutations in the HvDMC1 gene were identified (Table 1). All identified mutations were confirmed by sequencing and all of them are G/C to A/T transitions. Based on the number of the identified mutations in the HvDMC1 gene (6), the length of amplicon (811 bp) and the number of M2 plants screened (5,376), the calculated mutation density in this gene was 1 mutation per 729 kbp.
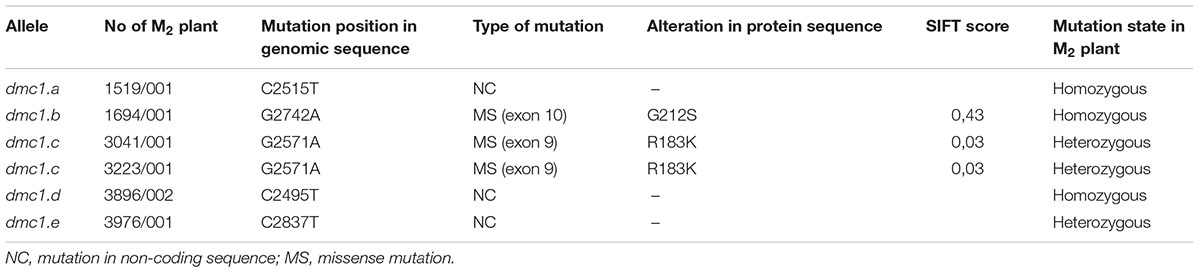
Table 1. Characteristics of mutations identified in the HvDMC1 gene.
Six mutations identified in the HvDMC1 gene gave five new alleles (dmc1.a – dmc1.e) (Table 1 and Figure 1). The same mutation G2571A (the dmc1.c allele) was induced and identified independently in two different M2 plants which originated from different M1 individuals: plant no. 3041/001 and plant no. 3223/001. To distinguish the origin of the mutated allele, it is hereafter named as dmc1-3041 or dmc1-3223 depending on the mutated line. Three mutations – dmc1.a, dmc1.d and dmc1.e occurred in non-coding, intron regions of the HvDMC1 gene. They were analyzed in silico and the positions of these mutations are neither in donor/acceptor sites of introns nor in polypyrimidine tracts or branch points, so they are probably not essential for splicing and do not have any impact on the encoded protein. Homozygous plants carrying these intronic mutations did not show any visible morphological changes when compared to ‘Sebastian’. Three other mutations – dmc1.b, dmc1.c-3041 and dmc1.c-3223, occurred in coding sequence (dmc1.b in exon 10, dmc1.c in exon 9) and they cause amino acids alterations at the protein level. The dmc1.b mutation changes glycine-212 to serine (G212S) and the dmc1.c mutation changes arginine-183 to lysine (R183K). Potentially, both of them can be used for functional analysis of the DMC1 gene in barley. The SIFT (Sorting Intolerant From Tolerant) tool was used to analyze in silico the influence of the identified mutations on protein activity and functioning. If the SIFT score is less than 0.05 the mutation is considered as deleterious for protein activity (Ng and Henikoff, 2003; Kumar et al., 2009). According to this bioinformatics analysis, the dmc1.b mutation is functionally neutral (SIFT score = 0.43), whereas the dmc1.c mutation is deleterious (SIFT score = 0.03). The multiple alignment of the DMC1 proteins from various species showed that the amino acid substituted in the dmc1.b mutant (glycine-212) is conserved among plant species, while the amino acid changed in the dmc1.c mutant (arginine-183) is conserved not only among plants, but also in Homo sapiens (Figure 3). This also suggests that the dmc1.c mutation may have more significant impact on the protein function, nevertheless for further investigation we have used all three mutant lines carrying missense mutations – dmc1.b, dmc1.c-3041 and dmc1.c-3223. We have developed homozygous mutant lines and used plants of the M4/M5 generation for backcross with their parent variety ‘Sebastian’ in order to reduce the number of putative background mutations. We selected homozygous mutant plants form the BC (backcross) F2 generations and used them for cytological analysis of meiosis.
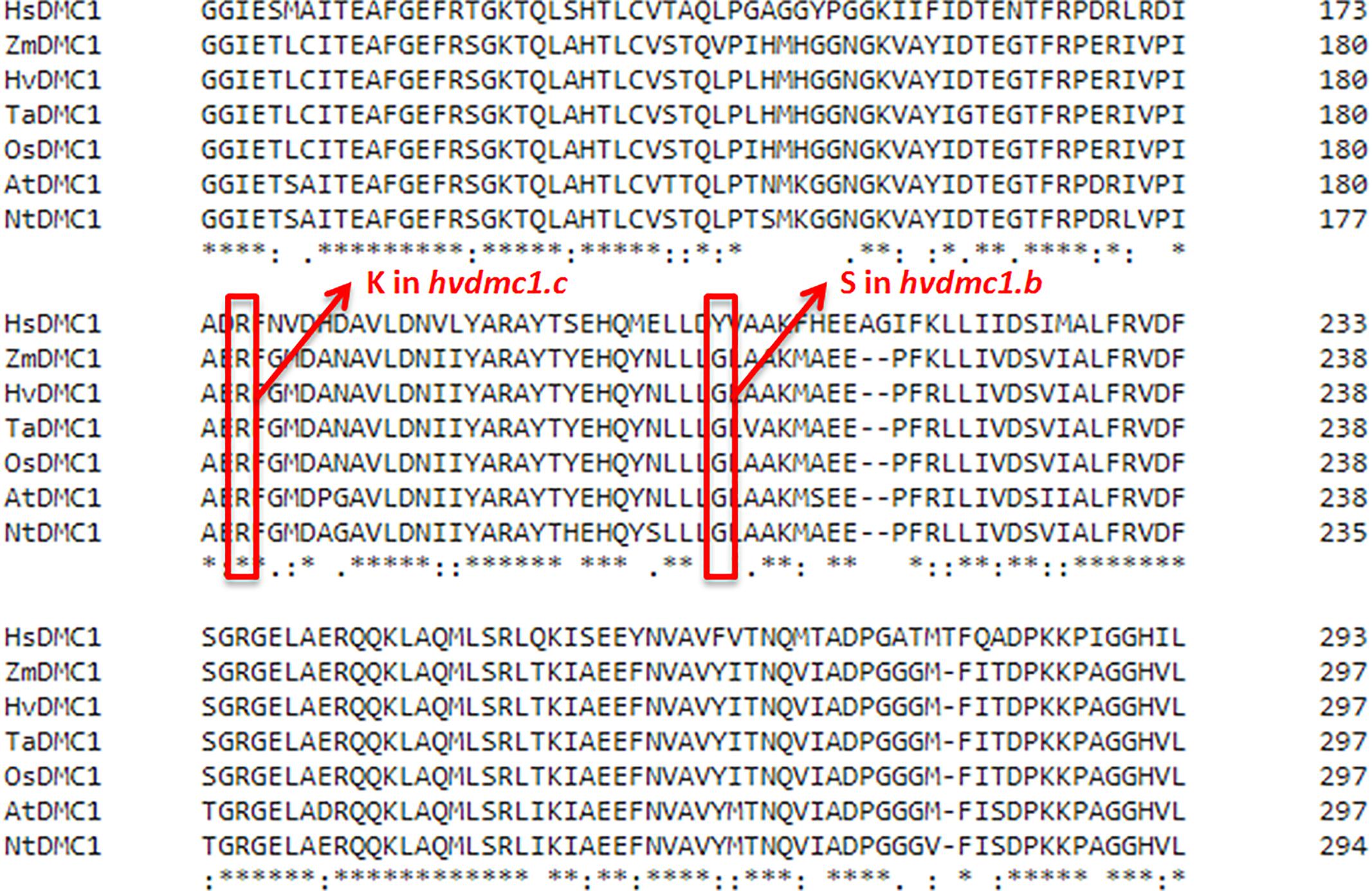
Figure 3. Multiple alignment of fragment of the DMC1 protein sequences from various species with positions of substituted amino acids in the dmc1.b and dmc1.c mutants indicated with red frames. Hs – Homo sapiens, Zm – Zea mays, Hv – Hordeum vulgare, Ta – Triticum aestivum, Os – Oryza sativa, At – Arabidopsis thaliana, Nt – Nicotiana tabacum.
Cytological Observations of Meiosis in the hvdmc1.b, hvdmc1.c-3041 and hvdmc1.c-3223 Mutants
The first observed phenotypic feature of all the identified dmc1 mutants selected for the cytological analysis was partial sterility of their spikes, which indicated some fertility disorders (Figure 4). Apart from that, the mutants did not show any evident morphological changes when compared to the wild type ‘Sebastian’. In all three dmc1 mutants we observed chromosome aberrations such as chromosomal bridges and chromosome fragments during anaphase and telophase I and II and micronuclei in tetrads (Figure 5). In the dmc1.c-3041 mutant an abnormal tetrads consisting of three or five haploid cells were observed (Figure 6). Additionally, we observed that both dmc1.c mutants showed disturbances in the formation of bivalents plate in metaphase I (Figure 7). However, this observation needs further investigations.

Figure 4. Spikes of cv. ‘Sebastian’ and the dmc1.b, dmc1.c-3041, and dmc1.c-3223 mutants displaying partial sterility. Red arrows indicate examples of places where no grains were developed.
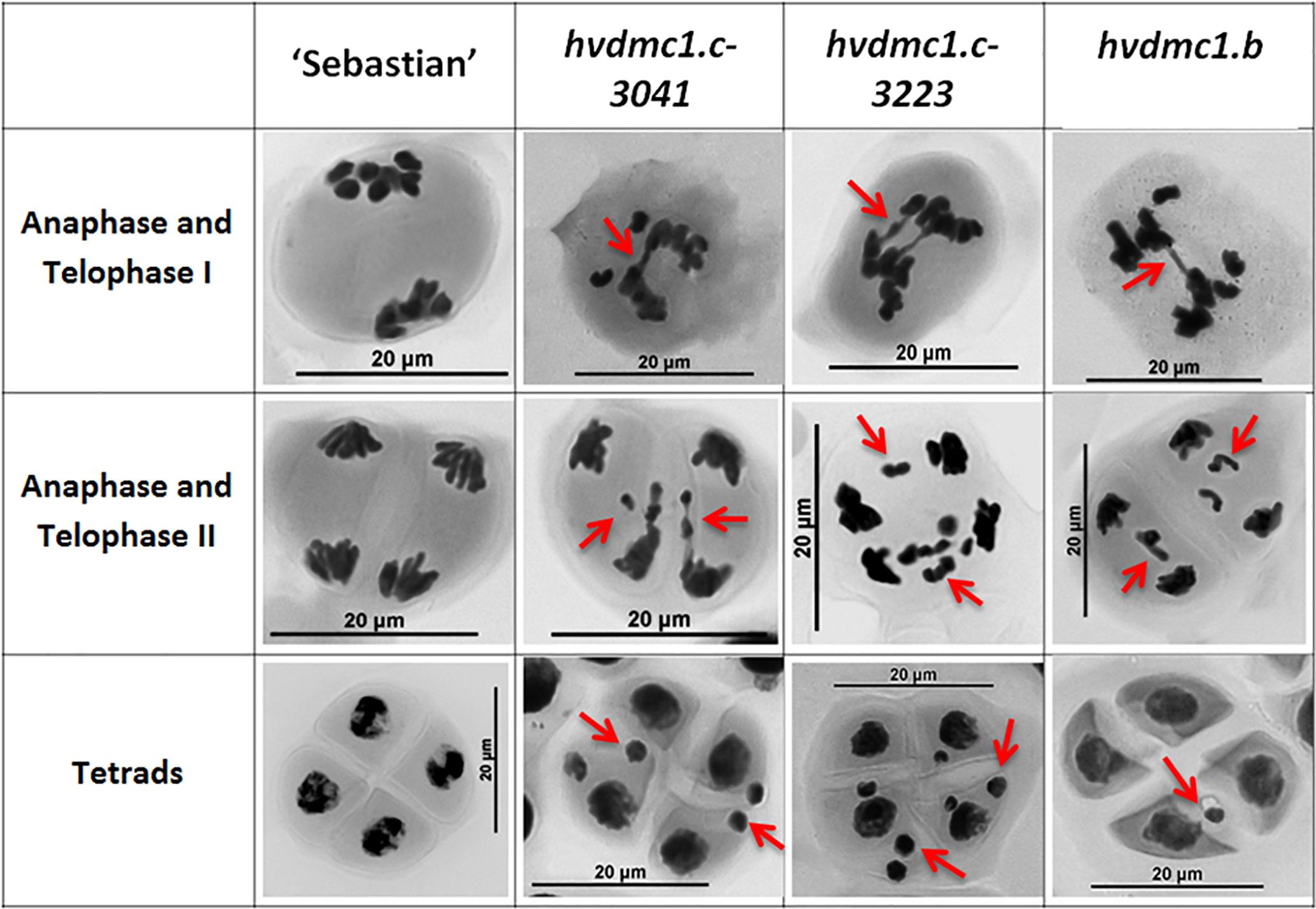
Figure 5. The summary panel with the examples of meiotic cells in different stages: anaphase/telophase I, anaphase/telophase II and in tetrads in ‘Sebastian’ and the dmc1.c-3041, dmc1.c-3223 and dmc1.b mutants. In the anaphase and telophase I the chromosome bridges, in the anaphase and telophase II the chromosome fragments, whereas in tetrad stage the micronuclei are indicated by red arrows.
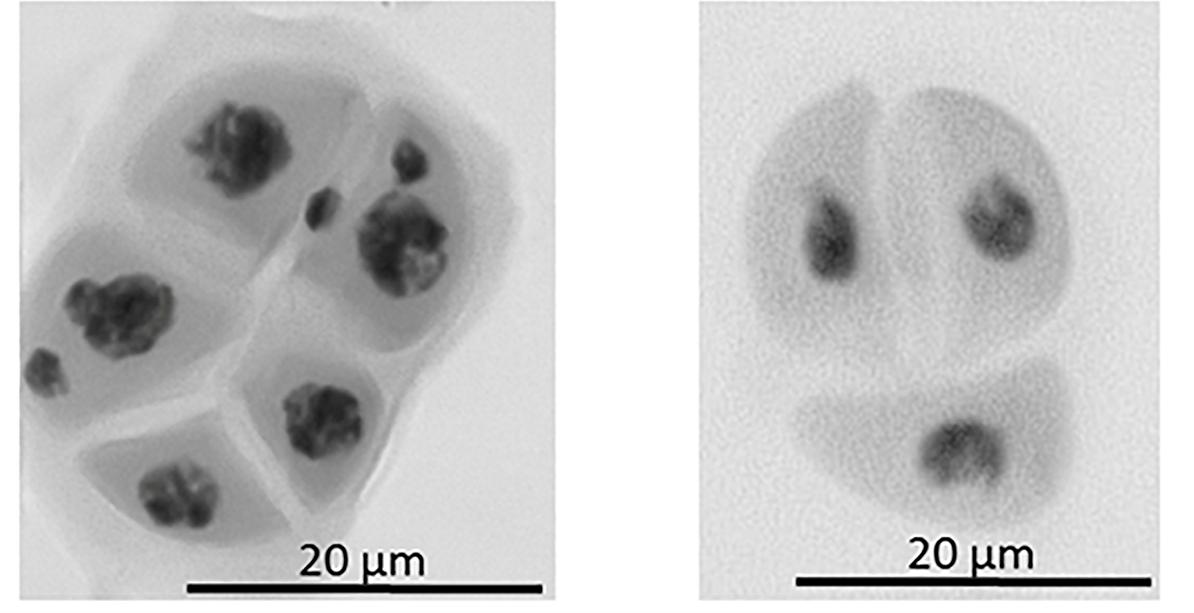
Figure 6. Example of abnormal tetrad formation in the dmc1.c-3041 mutant. On the left: tetrad composed of five cells, two of the cells show the presence of two micronuclei. On the right: tetrad composed of three cells, no micronuclei are present.
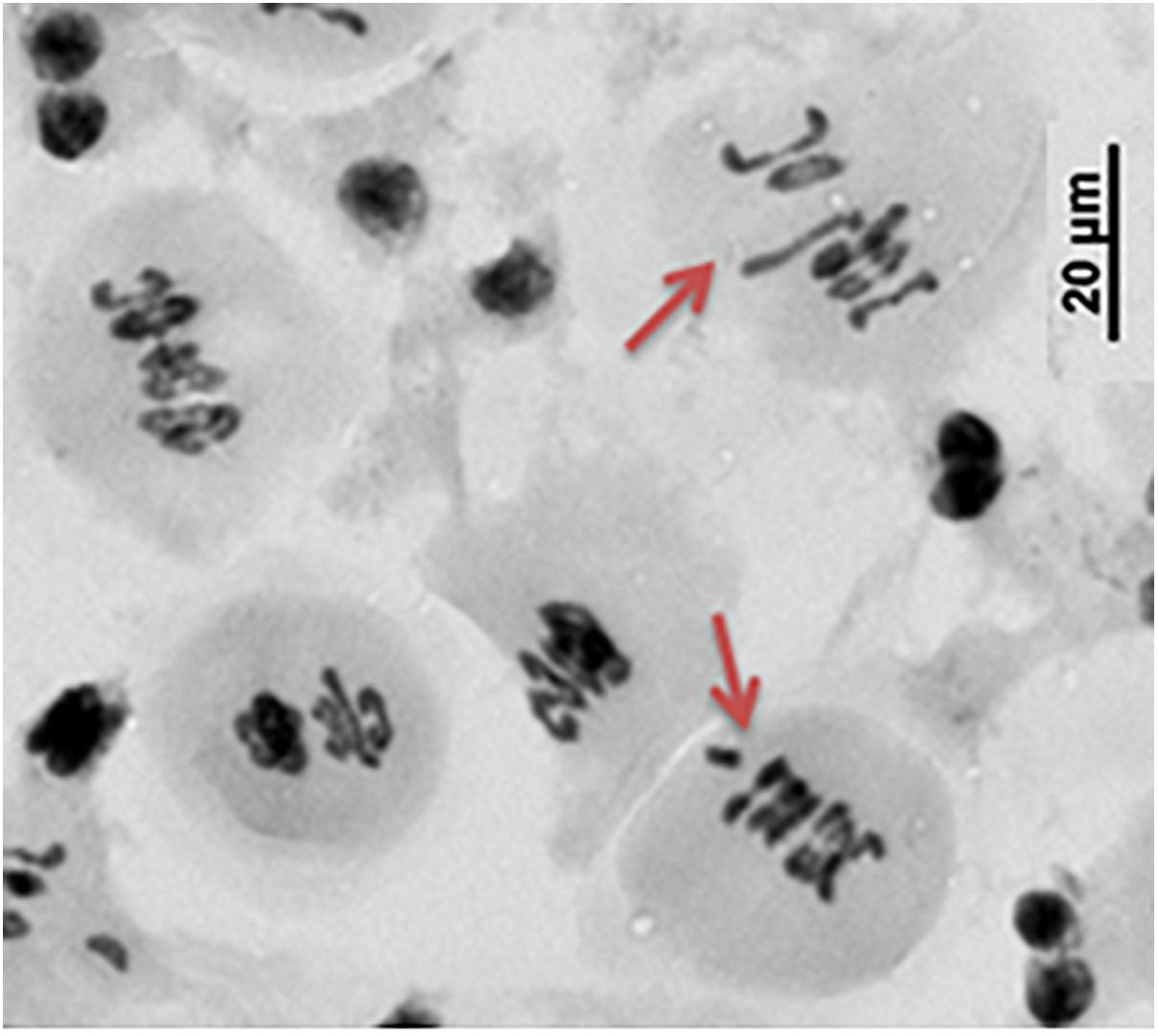
Figure 7. The abnormal chromosome assembling during metaphase I in the dmc1.c-3223 mutant.
In all analyzed meiosis phases, the dmc1.c-3041 and dmc1.c-3223 mutants displayed statistically significant differences in chromosomal aberration frequency compared to the wild type variety ‘Sebastian’ (Figures 8, 9). In anaphase/telophase I, the parent variety exhibited the chromosome aberration frequency at the level of 3%, whereas in the dmc1.c mutants the chromosome aberration frequency was significantly higher: 17% and 26% in dmc1.c-3041 and dmc1.c-3223, respectively (Figure 8A). In anaphase/telophase II, the frequency of cells with chromosome aberrations in the mutants was 19 and 40% (in the dmc1.c-3041 and dmc1.c-3223, respectively; Figure 8B) compared to 4% in ‘Sebastian’. The frequency of tetrads with micronuclei was also significantly higher in the mutants – 14 and 33% (in dmc1.c-3041 and dmc1.c-3223, respectively) than in the wild type, where it reached 1% (Figure 9).
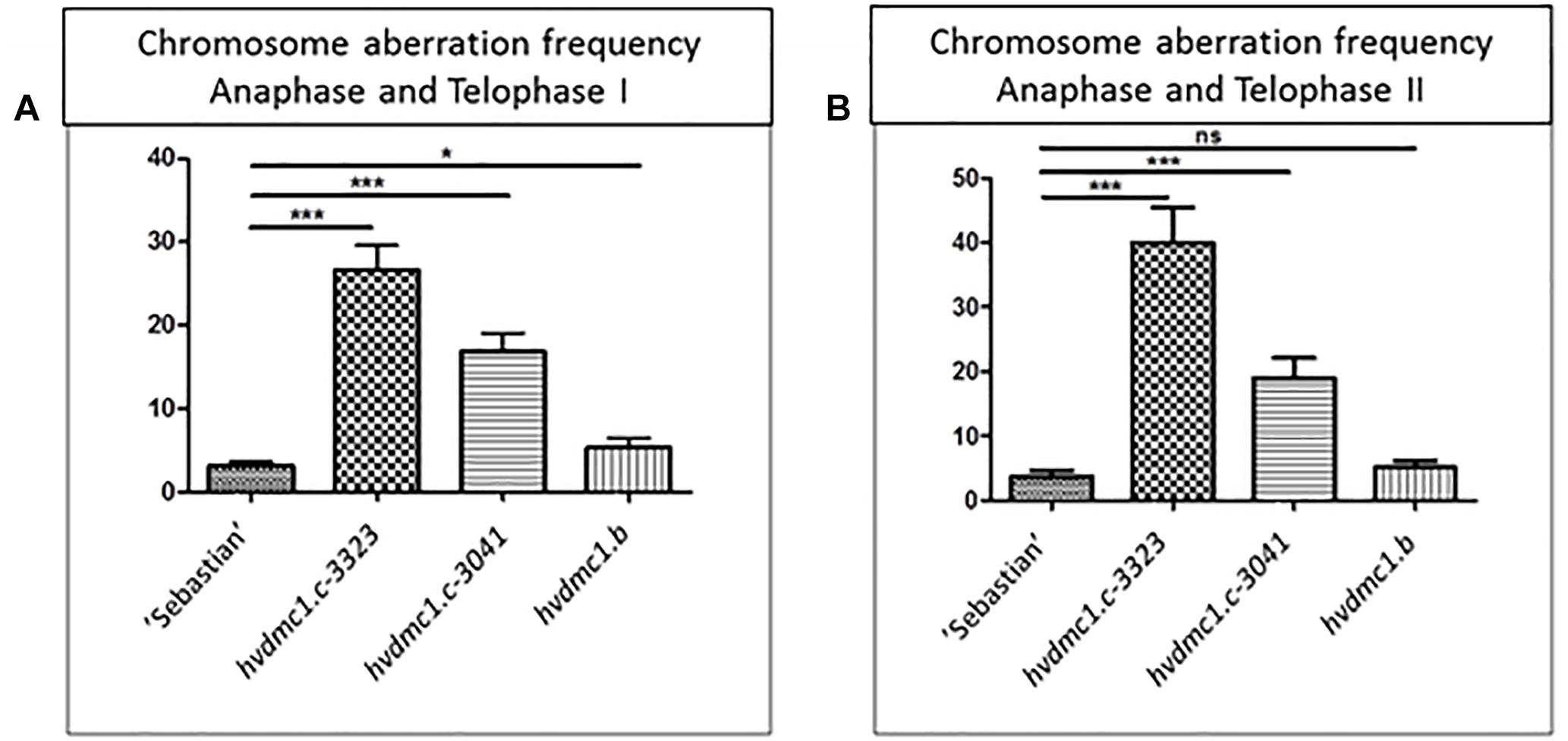
Figure 8. (A) Comparison of the chromosome aberration frequencies in anaphase and telophase I in ‘Sebastian’ (wt) and three dmc1 mutants. (B) Comparison of the chromosome aberration frequencies in anaphase and telophase II in ‘Sebastian’ and three dmc1 mutants. Stars indicate statistical significant differences (ANOVA; p < 0,05) between ‘Sebastian’ and the three dmc1 mutants, ns – differences between compared genotypes were not statistically significant.
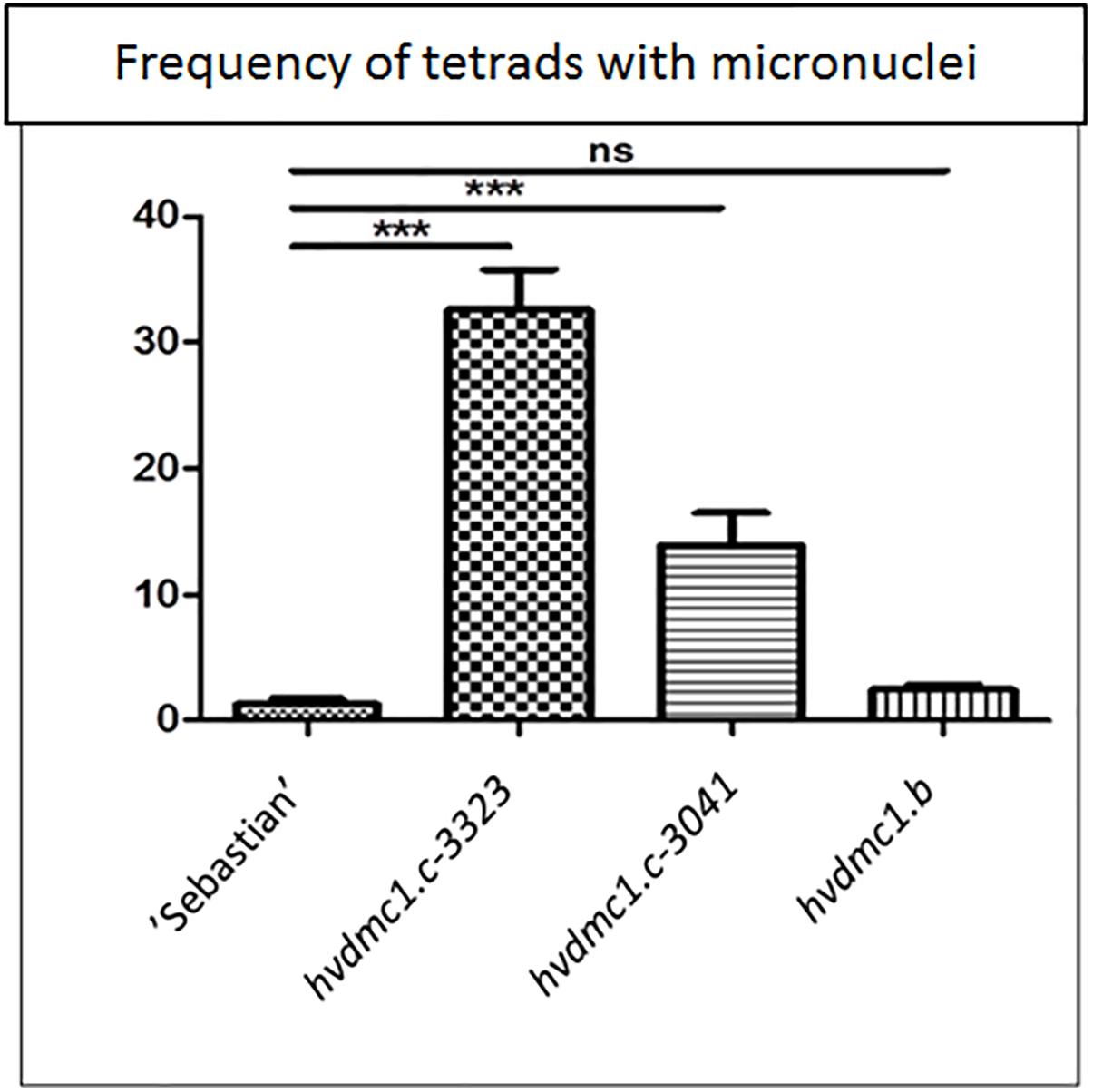
Figure 9. Comparison of the frequency of tetrads with micronuclei in ‘Sebastian’ (wt) and three dmc1 mutants. Stars indicate statistically significant differences (ANOVA; p < 0,05) between ‘Sebastian’ and the three dmc1 mutants, ns – differences between compared genotypes were not statistically significant.
The dmc1.b mutant also showed abnormalities during meiosis, but with much lower frequencies than the dmc1.c mutants. In anaphase and telophase I, the frequency of chromosomal aberrations in the dmc1.b mutant was 5.5%, about two times higher than in the wild type ‘Sebastian’. In anaphase and telophase II, the frequency of chromosome aberration (5%) observed in dmc1.b did not differ statistically from the parent variety ‘Sebastian’ (4%) (Figure 8). Analysis of micronuclei in tetrads has also shown no significant differences between this mutant and its parent variety (Figure 9).
Discussion
We have performed the analysis of meiosis in the dmc1.b mutant carrying missense mutation leading to the G212S substitution and both the dmc1.c lines (dmc1.c-3041 and dmc1.c-3223) carrying missense mutation causing the R183K change. Observation of meiosis in the mutants revealed that the dmc1.c lines showed differences in chromosomal aberrations frequency when compared to the wild type, whereas the dmc1.b mutant did not show significant disorders. This results are consistent with the in silico analysis of conservation of the substituted amino acid positions and with the SIFT values, which have shown that the dmc1.c mutation should have a more significant impact on protein function than dmc1.b.
Our dmc1.c mutants exhibited very high chromosome aberration frequencies in anaphase/telophase I/II and also very high number of tetrads with micronuclei. We suggest that these changes are the result of defects in DSB repair and anomaly in crossing-over, what strongly confirms that the DMC1 gene is involved in the DSB repair, recombination and chromosome disjunction during meiosis. Both dmc1.c mutants showed disturbances in the chromosome assembling during the metaphase I. Such abnormalities have previously been observed in other species, both in plants and animals. The DMC1-knock-out mice displayed aberrant chromosomal pairing or non-homologous chromosome pairing in spermatocytes (Habu et al., 1996). Our findings are also consistent with the observations of the atdmc1 mutants which exhibited abnormalities in the formation of bivalents and chiasmata (Da Ines et al., 2013). Moreover, in meiotic cells of the rice osdmc1a osdmc1b insertional double mutant, univalents and abnormal number of chromosomes in the metaphase plate during the second meiotic division were observed (Wang et al., 2016). In our study, one mutated line, dmc1.c-3041, formed irregular tetrads containing three or five haploid cells. Similar anomalies were observed in the rice osdmc1a osdmc1b double mutant (Wang et al., 2016). This type of anomaly may be the result of abnormal, uneven segregation of chromosomes to the opposite poles of the cell during meiotic divisions. The dmc1.b as well as both dmc1.c mutant lines show partial sterility of the spikes. Previously reported dmc1 mutants in different species showed sterility or partial sterility. For example, rice OsDMC1-RNAi lines, as well as the insertional osdmc1a osdmc1b double mutant grow normally during their vegetative phase, but they are characterized by total sterility (Deng and Wang, 2007; Wang et al., 2016). The insertional Arabidopsis mutant, atdmc1, produces viable seeds at very low ratio (1.5%) (Da Ines et al., 2013). The DMC1 knock-out mice displayed total sterility (Habu et al., 1996). This suggests that fertility disorders are a common feature of individuals lacking the DMC1 gene in different species. Our dmc1 mutants are not a knock-out type, they carry missense mutations in the analyzed gene and, as it was predicted in our analysis (Figure 1), the substituted amino-acid residues are located in the Rad51 domain, however outside the functional motifs of the DMC1 protein (such as the Walker A and B motifs, and loops 1 and 2), therefore the effect on sterility is not that strong as in other species.
In our study the mutational screening of 5,376 M2 plants from the HorTILLUS population revealed six independent G/C to A/T mutations within the HvDMC1 gene. Most mutations (88%) found to date in the HorTILLUS population represented this type of transition (calculated based on data of 32 genes TILLed; Szurman-Zubrzycka et al., 2018). Both mutagens used for creation of our TILLING population (MNU and NaN3) cause such DNA lesions. N-methyl-N-nitrosourea belongs to alkylating agents that are known to alkylate guanine and create O6-metG – the lesion with strong mutagenic property (Kleibl, 2002). O6-metG mispairs with thymine, which leads to its replacement by adenine in the subsequent replication cycle. If this methylation is induced in a non-transcribed (sense) DNA strand, it leads to G to A transition, whereas if it occurs in transcribed (antisense) DNA strand it results in C to T transition. TILLING populations which were developed after treatment with MNU for Glycine max and Oryza sativa showed 89.4 and 91.7% G/C to A/T transitions, respectively (Cooper et al., 2008; Suzuki et al., 2008). Sodium azide, the other mutagen used for establishing the HorTILLUS population, is mutagenic only for some plant species, among them barley and rice (reviewed in Gruszka et al., 2012). It was used as the only mutagen in other barley TILLING population - TILLMore developed for cultivar ‘Morex’, where it caused mainly G/C to A/T transitions (95.5%, Talamè et al., 2008; Sparla et al., 2014). The mutation density calculated based on mutations found in the HvDMC1 amplicon is 1 per 729 kbp. The average mutation density in the HorTILLUS population is 1 per 477 kbp, however it varies between gene fragments (Szurman-Zubrzycka et al., 2018). The value obtained for HvDMC1 is slightly lower, what could be caused by amplicon base content (G/C – 42%; A/T – 58%).
One very important aspect in terms of functional genetics is the presence of paralogs within the genome that can take over functions of gene of interest (functional redundancy). If there are two or more closely related genes, usually it is necessary to produce individuals with mutations in both or all of paralogs to perform functional analysis. Barley genome is one of the largest diploid genomes sequenced with a haploid genome size of more than 5 Gbp in seven large chromosomes (International Barley Genome Sequencing Consortium Mayer et al., 2012; Mascher et al., 2017). In order to check if there are any paralogs of HvDMC1 in barley genome we have screened its 2nd version that has been recently released (Mascher et al., 2017) with the use of the EnsemblPlants3 and the IPK Barley BLAST4 servers. Our analysis indicated that the HvDMC1 gene (HORVU5Hr1G040730) is located on chromosome 5 and has no paralogs in the genome (Supplementary Materials 1, 2), so our dmc1.b and dmc1.c mutants are good tools to study the function of this gene, because the risk of gene redundancy is very low.
An issue which is sometimes raised considering TILLING mutants is that observed phenotype may be caused by other, than analyzed, mutations in the genome (so called background mutations). Taking into consideration the size of barley genome and the overall mutation density found in the HorTILLUS population (ca. 1/500 kbp; Szurman-Zubrzycka et al., 2018), we can assume that each M2 plant carries more than 10,000 mutations. However, vast majority of them occur in non-coding regions, since genes (annotated coding sequences) make up 1.3% of barley genome (65.3 Mbp; Mascher et al., 2017). So, statistically, the number of mutations in genes equals to ca. 130 and, probably, most of them are silent and/or do not affect the protein function. Therefore, the probability of the presence of other deleterious mutation in a gene related to the same process of interest is very low. Nevertheless, in order to further decrease this probability, we performed backcrosses of the identified mutants with their parent variety ‘Sebastian’, which reduced the number of (putative) background mutations by half. The homozygous mutants selected from the F2 populations were phenotyped in this study. What is more, we observed that two different dmc1.c mutant lines (dmc1.c-3041 and dmc1.c-3223), that originated from different M1 plants and possess different mutational background, show similar defects during meiotic divisions. These two lines share only the mutation leading to the R183K substitution in DMC1, whereas any putative background mutations differ between them, which strongly suggests that the identified mutation is responsible for this phenotype.
Conclusion
The role of DMC1 has been validated mostly in model plant species, such as Arabidopsis thaliana and rice. Here, we described functional analysis of HvDMC1 in barley, which belongs to the most important cereal species worldwide. Our barley TILLING population, HorTILLUS, has been used for mutational screening in the HvDMC1 gene. We have identified and characterized a new allele, named dmc1.c, responsible for abnormalities during meiosis. Two mutated lines, from different M1 plants carrying the same mutation (G2571A that causes the R183K substitution), showed similar defects in this process, which strongly suggests that HvDMC1 is involved in the proper course of meiosis in barley. We conclude that DMC1 is required for DSB repair during meiosis, the process which has yet to be fully elucidated.
Data Availability
All datasets for this study are included in the manuscript and the Supplementary Files.
Author Contributions
IS and DG conceived the project and designed the experiments. DG supervised the project. MS-Z, BB, MS-J, and JK conducted the research. MS-Z, IS, and DG wrote the manuscript.
Funding
The authors gratefully acknowledge financial support of the research by the Food and Agriculture Organization (FAO) and International Atomic Energy Agency (IAEA) (Grant No. 15657 R0-R3) and by the Polish Ministry of Science and Higher Education (Grant Nos. 687/W-IAEA/2010/0, 773/W-IAEA/2010/0, 2214/FAO/IAEA/2011/0, and 2725/FAO/IAEA/2013/0).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2019.00761/full#supplementary-material
Footnotes
- ^www.ebi.ac.uk/Tools/msa/clustalo/
- ^http://pfam.sanger.ac.uk/
- ^http://plants.ensembl.org/index.html
- ^http://webblast.ipk-gatersleben.de/barley_ibsc/viroblast.php
References
Andersen, A., and Sekelsky, J. (2010). Meiotic versus mitotic recombination: two different routes for double-strand break repair. Bioessays 32, 1058–1066. doi: 10.1002/bies.201000087
Barakate, A., Higgins, J. D., Vivera, S., Stephens, J., Perry, R. M., Ramsay, L., et al. (2014). The synaptonemal complex protein ZYP1 is required for imposition of meiotic crossovers in barley. Plant Cell 26, 729–740. doi: 10.1105/tpc.113.121269
Bianco, P. R., Tracy, R. B., and Kowalczykowski, S. C. (1998). DNA strand exchange proteins: a biochemical and physical comparison. Front. Biosci. 3:D570–D603. doi: 10.2741/A304
Bishop, D. K., Park, D., Xu, L., and Kleckner, N. (1992). DMC1: a meiosis-specific yeast homolog of E. coli recA required for recombination, synaptonemal complex formation, and cell cycle progression. Cell 69, 439–456. doi: 10.1016/0092-8674(92)90446-J
Cloud, V., Chan, Y.-L., Grubb, J., Budke, B., and Bishop, D. K. (2012). Dmc1 catalyzes interhomolog joint molecule formation in meiosis with Rad51 and Mei5-Sae3 as accessory factors. Science 337, 1222–1225. doi: 10.1126/science.1219379
Colas, I., Barakate, A., Macaulay, M., Schreiber, M., Stephens, J., Vivera, S., et al. (2019). desynaptic5 carries a spontaneous semi-dominant mutation affecting Disrupted Meiotic cDNA 1 in barley. J. Exp. Bot. 70, 2683–2698. doi: 10.1093/jxb/erz080
Cooper, J. L., Till, B. J., Laport, R. G., Darlow, M. C., Kleffner, J. M., Jamai, A., et al. (2008). TILLING to detect induced mutations in soybean. BMC Plant Biol. 8:9. doi: 10.1186/1471-2229-8-9
Da Ines, O., Degroote, F., Goubely, C., Amiard, S., Gallego, M. E., and White, C. I. (2013). Meiotic recombination in Arabidopsis is catalysed by DMC1, with RAD51 playing a supporting role. PLoS Genet. 9:e1003787. doi: 10.1371/journal.pgen.1003787
Deng, Z., and Wang, T. (2007). OsDMC1 is required for homologous pairing in Oryza sativa. Plant Mol. Biol. 65, 31–42. doi: 10.1007/s11103-007-9195-2
Gruszka, D., Szarejko, I., and Maluszynski, M. (2012). “Sodium azide as a mutagen,” in Plant Mutation Breeding and Biotechnology. eds Q. Shu, B. Forster, and H. Nakagawa (Wallingford: CAB International Publishing House), 159–166. doi: 10.1079/9781780640853.0159
Habu, T., Taki, T., West, A., Nishimune, Y., and Morita, T. (1996). The mouse and human homologs of DMC1, the yeast meiosis-specific homologous recombination gene, have a common unique form of exon-skipped transcript in meiosis. Nucleic Acids Res. 24, 470–477. doi: 10.1093/nar/24.3.470
Higgins, J. D., Perry, R. M., Barakate, A., Ramsey, L., Waugh, R., Halpni, C., et al. (2012). Spatiotemporal asymmetry of the meiotic program underlies the predominantly distal distribution of meiotic crossovers in barley. Plant Cell 24, 4096–4109. doi: 10.1105/tpc.112.102483
International Barley Genome Sequencing Consortium Mayer, K. F., Waugh, R., Brown, J. W., Schulman, A., Langridge, P., et al. (2012). A physical, genetic and functional sequence assembly of the barley genome. Nature 491, 711–716. doi: 10.1038/nature11543
Jost, M., Szurman-Zubrzycka, M., Gajek, K., Szarejko, I., and Stein, N. (2019). “TILLING in barley,” in Barley. Mathods in Molecular Biology, Vol. 1900, ed. W. Harwood (New York, NY: Humana Press), 73–94. doi: 10.1007/978-1-4939-8944-7_6
Keeney, S., Giroux, C., and Kleckner, N. (1997). Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell 88, 375–384. doi: 10.1016/S0092-8674(00)81876-0
Kleibl, K. (2002). Molecular mechanisms of adaptive response to alkylating agents in Escherichia coli and some remarks on O6-methylguanine DNA-methyltransferase in other organisms. Mutat. Res. 512, 67–84. doi: 10.1016/S1383-5742(02)00025-X
Klimyuk, V., Garkoucha, T., Schwarzacher, T., and Jones, J. D. G. (2000). Hordeum vulgare DMC1 protein gene, complete cds. The NCBI database www.ncbi.nlm.nih.gov/nuccore/AF234170.1, GenBank accession no.: AF234170.1
Kumar, P., Henikoff, S., and Ng, P. (2009). Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protoc. 4, 1073–1081. doi: 10.1038/nprot.2009.86
Mascher, M., Gundlach, H., Himmelbach, A., Beier, S., Twardziok, S. O., Wicker, T., et al. (2017). A chromosome conformation capture ordered sequence of the barley genome. Nature 544, 427–433. doi: 10.1038/nature22043
Mercier, R., Mézard, C., Jenczewski, E., Macaisne, N., and Grelon, M. (2015). The molecular biology of meiosis in plants. Annu. Rev. Plant Biol. 66, 297–327. doi: 10.1146/annurev-arplant-050213-035923
Neale, M., and Keeney, S. (2006). Clarifying the mechanics of DNA strand exchange in meiotic recombination. Nature 442, 153–158. doi: 10.1038/nature04885
Ng, P., and Henikoff, S. (2003). SIFT: predicting amino acid changes that affect protein function. Nucleic Acids Res. 31, 3812–3814. doi: 10.1093/nar/gkg509
Ohnishi, T., Mori, E., and Takahashim, A. (2009). DNA double-strand breaks: their production, recognition, and repair in eukaryotes. Mutat. Res. 669, 8–12. doi: 10.1016/j.mrfmmm.2009.06.010
Ramsay, L., Colas, I., and Waugh, R. (2014). “Modulation of meiotic recombination,” in Biotechnological Approaches to Barley Improvement. Biotechnology in Agriculture and Forestry 69. eds J. Kumlehn and N. Stein (Berlin: Springer), 311–329. doi: 10.1007/978-3-662-44406-1_16
Robert, T., Nore, A., Brun, C., Maffre, C., Crimi, B., Bourbon, H.-M., et al. (2016). The TopoVIB-Like protein family is required for meiotic DNA double-strand break formation. Science 351, 943–949. doi: 10.1126/science.aad5309
Shinohara, A., Ogawa, H., and Ogawa, T. (1992). Rad51 protein involved in repair and recombination in S.cerevisiae is a RecA-like protein. Cell 69, 457–470. doi: 10.1016/0092-8674(92)90447-K
Shinohara, A., and Shinohara, M. (2004). Roles of RecA homologues Rad51 and Dmc1 during meiotic recombination. Cytogenet. Genome Res. 107, 201–207. doi: 10.1159/000080598
Singh, G., Da Ines, O., Gallego, M. E., and White, C. (2017). Analysis of the impact of the absence of RAD51 strand exchange activity in Arabidopsis meiosis. PLoS One 12:e0183006. doi: 10.1371/journal.pone.0183006
Sparla, F., Falini, G., Botticella, E., Pirone, C., Talamè, V., Bovina, R., et al. (2014). New starch phenotypes produced by TILLING in barley. PLoS One 9:e107779. doi: 10.1371/journal.pone.0107779
Suzuki, T., Eiguchi, M., Kumamaru, T., Satoh, H., Matsusaka, H., Moriguchi, K., et al. (2008). MNU-induced mutant pools and high performance TILLING enable finding of any gene mutation in rice. Mol. Genet. Genom. 279, 213–223. doi: 10.1007/s00438-007-0293-2
Szostak, J., Orr-Weaver, T., Rothstein, R., and Stahl, F. (1983). The double-strand-break repair model for recombination. Cell 33, 25–35. doi: 10.1016/0092-8674(83)90331-8
Szurman-Zubrzycka, M., Chmielewska, B., Gajewska, P., and Szarejko, I. (2017). “Mutation detection by analysis of DNA heteroduplexes in TILLING populations of diploid species,” in Biotechnologies for Plant Mutation Breeding. eds J. Jankowicz-Cieslak, T. Tai, J. Kumlehn, and B. J. Till (Cham: Springer International Publishing), 281–303. doi: 10.1007/978-3-319-45021-6_18
Szurman-Zubrzycka, M., Zbieszczyk, J., Marzec, M., Jelonek, J., Chmielewska, B., Kurowska, M., et al. (2018). HorTILLUS – a rich and renewable source of induced mutations for forward/reverse genetics and pre-breeding programs in barley (Hordeum vulgare L.). Front. Plant Sci. 9:216. doi: 10.3389/fpls.2018.00216
Talamè, V., Bovina, R., Sanguineti, M., Tuberosa, R., Lundqvist, U., and Salvi, S. (2008). TILLMore, a resource for the discovery of chemically induced mutations in barley. Plant Biotechnol. J. 6, 477–485. doi: 10.1111/j.1467-7652.2008.00341.x
The Arabidopsis Genome Initiative [AGI] (2000). Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408, 796–815. doi: 10.1038/35048692
Vrielynck, N., Chambon, A., Vezon, D., Pereira, L., Chelysheva, L., De Muyt, A., et al. (2016). A DNA topoisomerase VI-like complex initiates meiotic recombination. Science 351, 939–943. doi: 10.1126/science.aad5196
Keywords: barley, chromosome aberrations, crossing-over, DMC1, meiosis, TILLING
Citation: Szurman-Zubrzycka M, Baran B, Stolarek-Januszkiewicz M, Kwaśniewska J, Szarejko I and Gruszka D (2019) The dmc1 Mutant Allows an Insight Into the DNA Double-Strand Break Repair During Meiosis in Barley (Hordeum vulgare L.). Front. Plant Sci. 10:761. doi: 10.3389/fpls.2019.00761
Received: 06 March 2019; Accepted: 24 May 2019;
Published: 11 June 2019.
Edited by:
Ayako N. Sakamoto, National Institutes for Quantum and Radiological Science and Technology, JapanReviewed by:
Charles I. White, Centre National de la Recherche Scientifique (CNRS), FranceShuhei Nasuda, Kyoto University, Japan
Copyright © 2019 Szurman-Zubrzycka, Baran, Stolarek-Januszkiewicz, Kwaśniewska, Szarejko and Gruszka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Damian Gruszka, ZGFtaWFuLmdydXN6a2FAdXMuZWR1LnBs
†These authors have contributed equally to this work