 Attila-Károly Szabó1,2*
Attila-Károly Szabó1,2* Éva Várallyay3
Éva Várallyay3 Emese Demian3
Emese Demian3 Anna Hegyi3Zsuzsanna Nagyné Galbács3
Anna Hegyi3Zsuzsanna Nagyné Galbács3 József Kiss2János Bálint1
József Kiss2János Bálint1 Hugh D. Loxdale4
Hugh D. Loxdale4 Adalbert Balog1*
Adalbert Balog1*- 1Department of Horticulture, Faculty of Technical and Human Sciences, Sapientia Hungarian University of Transylvania, Târgu Mureş, Romania
- 2Institute of Plant Protection, Faculty of Agricultural and Environmental Sciences, Szent István University, Gödöllő, Hungary
- 3Molecular Plant Pathology Group, Department of Genomics, Agricultural Biotechnology Research Institute, Agricultural Research and Innovation Centre, Gödöllő, Hungary
- 4School of Biosciences, Cardiff University, Cardiff, United Kingdom
In the present study, we conducted field surveys to detect the population density of the most important invasive weed species and their associated virus vectoring aphids in crops grown under high input field (HIF) vs. low-input field (LIF) conditions, with and without fertilizers and pesticides. The most frequent invasive weed species were annual fleabane, Erigeron annua (L.), Canadian horseweed, Erigeron canadensis (L.) and Canadian goldenrod, Solidago canadensis (L.). These species were predominantly hosts of the aphids Brachycaudus helichrysi and Aulacorthum solani under both management systems. The 13% higher coverage of E. annua under LIF conditions resulted in a 30% higher B. helichrysi abundance and ∼85% higher A. solani abundance compared with HIF conditions. To reveal the incidence of virus infection in crop plants and invasive weeds, high-throughput sequencing of small RNAs was performed. Bioinformatics analysis combined with independent validation methods revealed the presence of six viruses, but with strikingly different patterns under LIF and HIF conditions. Their presence without symptoms in invasive weeds and crop plants supports the necessity of employing new approaches to those currently employed in invasive weed management. These findings also suggest that invasive weeds could serve as hosts for local aphid species and reservoirs for plant pathogenic viruses, both under low and high input management systems. In this light, as here demonstrated, viruses transmitted by local aphid species were found to differ between the management systems; hence, the importance of B. helichrysi and A. solani as virus vectors in particular clearly needs to be re-evaluated. Altogether, we accept that the present study is a pilot one and individual virus vectoring of aphids still needs to be directly tested. Even so, it represents one of the first contributions to this particular area, and thereby paves the way for further similar applied research in the future.
Introduction
Invasive weeds represent a serious threat to world crop production as global trade expands and climatic conditions shift (Agrow, 2015; Usda Forest Service, 2015). In this light, it has recently been suggested that the losses to crop yield caused by invasive weeds and their infesting aphids (Hemiptera: Aphididae), may increase substantially (by around 25%) within the EU over the next few years (Usda Forest Service, 2015; EPPO, 2017). Such weeds are particularly important because they not only serve as hosts for several local sap-feeding, virus-vectoring insects, especially aphids, but also because they may act as significant reservoirs of pathogenic plant viruses (Frey et al., 2003; Zimmermann et al., 2015).
Previous studies from Central Europe have reported 435 alien weed species from 82 plant families in the past 25 years (Anastasiu and Negrean, 2005). Most of these species, for example annual fleabane, Erigeron annua (L.), Canadian horseweed, Erigeron canadensis (L.), and goldenrod, Solidago canadensis L., occur in all types of habitat and are considered to be the most important and economically-relevant invasive weeds within the agro-ecosystem (Anastasiu and Negrean, 2005). Although weed control strategies involve apparently useful approaches, including physical (e.g. mulching, tilling, burning) and chemical and other control (e.g. use of high quality seeds, crop rotation, herbicide application) (Chitsaz and Nelson, 1983; Rand and Louda, 2004; Uchino et al., 2012; Mabuza et al., 2013), the net areas covered by invasive weed species continues to increase in Central European regions (Tunaitienė et al., 2015; Pacanoski, 2017).
An important factor worthy of consideration in relation to this topic is the effect of these weed species on local sap-feeding pest insect populations, especially aphids. Another factor is that invasive weeds acting as virus reservoirs pose a serious threat via local aphid species in terms of virus distribution and transmission to crop plants. From this standpoint, several virus vectoring aphid species whose host range naturally includes local plant species, mainly from the same family e.g. Asteraceae, have become even more important, feeding and reproducing on invasive weeds (Bell, 1983; Popkin et al., 2017). Direct and indirect interactions between weeds and crop plants in close proximity can influence each other’s susceptibility (“associational susceptibility”) and affects their herbivore abundances (Barbosa et al., 2009). However, aspects on natural habitat diversity (i.e. diverse habitat surrounded by natural landscape mosaics) and how management systems (low vs. high chemical input) influence associational susceptibility or resistance have rarely been included in such analyses (Steffan-Dewenter et al., 2001).
In accordance with these various factors, the aim of the present study was: (1) to assess the population density of the most important invasive weeds under differently managed agricultural systems (high-input fertilizers and chemical pesticides (high-input fields, HIF) vs. no chemical management (low-input fields, LIF); (2) to identify and compare population densities of the most frequent aphid species on the most frequent invasive weeds; and (3) survey and identify plant viruses using high-throughput small RNA sequencing of invasive weeds and surrounding crops under different management regimes.
Materials and Methods
Study Area, Focal Weed and Aphid Species
Field surveys were conducted over a 2-year period during the crop growing seasons of 2015 and the 2016 in Central and Eastern Transylvania, Romania. The methods used have also been described by Szabó et al. (2019). Assessments were run under two different input management regimes (low and high), carefully selected to take into consideration similar geographic and climate regions (see also / Supplementary Figure S1).
Low-Input, Traditionally Managed Fields (LIF)
This area belongs to a traditionally managed field (low-input) of the Old Saxon cultural region covering an area of ∼ 7,440 km2 at an altitude ranging from 230 to 800 m above sea level (a.s.l.) and characterized by a landscape mosaic of different land-cover types (28% forest, 24% pasture, and 37% arable land, mostly maize, potato, and alfalfa). The farming practices are predominantly small scale subsistence farming, with no chemical inputs. One consequence of this kind of land use is the exceptional biodiversity and natural value of the farming landscape (Akeroyd and Page, 2011). However, being not particularly viable economically, the abandonment of croplands in this region is frequent, resulting in the establishment and high abundance of invasive weeds (Zimmermann et al., 2015).
High-Input, Conventionally Managed Fields (HIF)
This study region contrasts the previously described region by growing large monocultures and farming landscapes with low levels of natural vegetation. The area of about 5,500 km2 at altitudes between 220 and 750 m a.s.l. has been, and continues to be, intensively treated with synthetic fertilizers and pesticides, major crops including maize, potato, and alfalfa (Supplementary Table S1). The studied fields inside the described region were situated at the same altitudinal range of about 250 m a.s.l. and under comparable bioclimatic conditions. The distance between the studied areas was roughly 200 km, with no direct connections (main roads and railways) between regions.
Previous assessments confirmed that three weed and two native aphid species are frequent in both regions, so these particular plant hosts and their infesting aphids were studied and sampled. The most important weed species found were the annual fleabane, Stenactis (=Erigeron) annua (L.), Canadian horseweed, Erigeron (=Conyza) canadensis (L.), and Canadian goldenrod, Solidago canadensis (L.). These species are known to grow in a diverse range of habitats and are considered important weeds in Europe, causing substantial crop losses following colonisation of new areas (Anastasiu and Negrean, 2005). Erigeron annua is often a dominant species within invasive weed communities, and has been reported from almost all European countries, its expansion increasing over recent years (Edwards et al., 2006; Tunaitienė et al., 2015; Pacanoski, 2017). Erigeron canadensis is an annual plant native throughout most of North and Central America. It is also widely naturalized in Eurasia (Nandula et al., 2006; Shah et al., 2014; Bajwa et al., 2016). Solidago canadensis is a perennial weed native to north-eastern and north-central America, but has established as an invasive throughout Europe (Abhilasha et al., 2008; Fenesi et al., 2015).
The two native aphid pests species examined in this study were the highly polyphagous leaf-curling plum aphid, Brachycaudus helichrysi (Kaltenbach) and the similarly polyphagous foxglove aphid, Aulacorthum solani (Kaltenbach) (Blackman and Eastop, 2000; Blackman, 2010). These are particularly important species, not only because of their wide host range, but also because of their diverse virus transmission. The host plant range of B. helichrysi includes members of the Asteraceae, e.g. Chrysanthemum, species of Prunus and also species of Solanum, Fragaria, Trifolium, Medicago, Citrus and maize, Zea mays (Tatchell et al., 1983; Powell et al., 1992; Isac et al., 1998; Popkin et al., 2017). Viruses transmitted by this aphid include Plum Pox virus, Potato virus Y and Beet mild yellowing virus (Isac et al., 1998). Host plants of A. solani include several crop plants including tomato, peppers, tobacco, celery, carrots, tulip bulbs, cucurbits, and legumes (Tatchell et al., 1983; Jandricic et al., 2014). The most important viruses transmitted by it are Potato viruses A, Y and X and Potato leaf roll virus, Cucumber mosaic virus, Soybean dwarf virus, Bean yellow mosaic virus and Turnip yellows virus (Jandricic et al., 2010, 2014).
Assessment of Invasive Weeds and Associated Aphids
Firstly, we selected two blocks of land approximately 3 km long and 1 km wide inside each management system (regions); these blocks were located 10 km distant from each other in LIF and in 15 km distant in HIF. Inside each block, we further established two transects (at least 1 km apart) of 10 m long × 1 m wide at an approximately equal distance (between 1 and 2 metre) to three major crops (maize, potato, and alfalfa) dominating (as crop) more than 95% of both management systems at field crop margins. In this way, each transect was surrounded by at least 8–10 ha of high-input, and at least 0.5–3 ha of low-input agricultural crops, including around weed populations dominated by maize, alfalfa and potato fields. The transect selections were made at the same period each year (between 5 and 8 May) and when all crop plants from both regions were at the same vegetation stage. The same transect and the same crop plant combinations were followed the next year. Each transect was carefully measured and located using GPS.
Secondly inside each transect, 10 × 1 m2 quadrats were placed. Each of these was further subdivided into 10 × 10 cm sub-quadrats (100 sub-quadrats inside each quadrat). Inside each of these sub-quadrats all plants (native and invasive) were counted and their coverage estimated (Andújar et al., 2010).
Thirdly, ten individual invasive weed plants from each sub-quadrat were randomly collected by hand and placed in plastic bags. The number of invasive plants collected for each species from each sub-quadrat mirrored the coverage of the species within the quadrat. We decided, subjectively, that at least eight plants be collected when the coverage of a given species in a sub-quadrat was at least 80% and comprised up to two plants if the coverage of the species was up to 20%. We decided upon these percentage coverage thresholds because in each quadrat there was one highly dominant invasive plant species (its coverage having at least 80%) and one species which had a coverage between 15 and 20%. Therefore, from each sub-quadrat, out of the 10 plants sampled, at least eight belonged to the dominant species and one or two to the second most dominant species.
Because plants contained aphid colonies, and the exact number of individual aphids was important, all plastic bags were labelled and kept at low temperature (∼0–4°C in a cool box), then returned to the laboratory, whereupon all samples were stored at −20°C, with aphids later counted and species identified (Blackman and Eastop, 2000; Blackman, 2010). In total, 100 plant samples were collected per transect and management system (400 samples per management system per collection data, and a total 1600 samples/year). The same methods of using quadrat dimensioning were followed in crop plants around each transect, i.e. equal distance (between 1 and 2 metre) to major crops (maize, potato, and alfalfa) dominating sites, except with regard to counting percentage plant coverages; here only plant material (the same number of samples as from weeds) were collected and stored for aphid assessment and virus detection.
Assessment began at the end of May and was repeated fortnightly five times during the summer until the end of the weed growing season, whereupon no more aphids were found. The whole procedure was repeated in the following year using the same collection methods within the same transects. All aphids were carefully counted under laboratory conditions, and the plant materials were used for virus identification employing metagenomic high-throughput sequencing (HTS) of small RNAs as an unbiased method, able to detect all viruses present in the sample (Roossinck et al., 2015).
Virus Detection From Invasive Weeds and Crop Plants
Leaf samples on which aphids had been found and counted were used for viral analyses from both weeds and crop plants (alfalfa, maize, and potato) and both management systems, with small RNA analyses performed in 2017. RNA was extracted using a phenol-chloroform method (White and Kaper, 1989). Briefly, frozen plant material was homogenized in an ice-cold mortar, suspended in 650 μl of extraction buffer (100 mM glycine, pH 9.0, 100 mM NaCl, 10 mM EDTA, 2% SDS and 1% sodium lauroylsarcosine) and mixed with an equal volume of water saturated phenol, and centrifuged for 5 min. The aqueous phase was treated with equal volumes of phenol, chloroform, and isoamyl-alcohol (25:24:1), and after subsequent treatment with chloroform: isoamyl-alcohol (24:1), was precipitated with 99.8% ethanol and then re-suspended in sterile water.
For small RNA HTS, small RNA was isolated from polyacrylamide gels involving RNA pools which were prepared by mixing equal amounts of RNA originating from different individuals, collection times (in the case of weeds) and from different species in the case of crops (Supplementary Table S2). This pooling strategy allowed detection of any virus present in any of the sampled individual plants at any time during the survey. As the crops investigated belonged to different families, and as such hosted very different viruses, we investigated their viral patterning as a collective pool. In contrast, the invasive weeds were all members of the Asteraceae; hence virus diagnostics were here performed separately for each species concerned.
These pools were used for small RNA library preparation (six libraries in total) using Truseq Small RNA Library Preparation Kit (Illumina, United States) and our modified protocol (Czotter et al., 2018). Samples were sequenced using HiScan2000 by UD Genomed (Debrecen, Hungary) 50 bp, single end. Fastq files of the sequenced libraries were deposited to the GEO and can be accessed through series accession number GSE132755.
Virus Diagnostics by RT-PCR and Northern Blot
Pooled RNA extracts or RNA extracts prepared from the individual crops were used as templates for cDNA synthesis by a RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, United States) with random primers according to the manufacturer’s instructions. The generated cDNA was then used for PCRs (primers amplifying viral parts are provided in Supplementary Table S6) performed with Phire Hot Start II DNA Polymerase (Thermo Fisher Scientific). Results were analysed using 1.2% agarose gel-electrophoresis, and PCR products were subsequently Sanger sequenced to prove them to be virus specific.
For Northern blot analysis, 4 μg of total RNA (the same pooled samples used for small RNA library preparation) were separated on a formaldehyde-containing 1.2% agarose gel and blotted to a Hybond-N membrane. Radioactively labelled random DNA probes were generated from cloned, purified PVX PCR product with a Decalabel DNA Labeling Kit (Thermo Scientific). Northern blots were hybridized with this probe in Church buffer (1% BSA (bovine serum albumin), 1 mM EDTA, 0.25 M Na2HPO4, 7% SDS, pH 7.2) at 65°C, washed according to the manufacturer’s instructions and exposed to X-ray film.
Data Analysis
For weed data, the mean coverage per 1 m2 sub-quadrat was determined by averaging the plant values from each 10 × 10 cm plot. Next, the inter-annual differences in coverage were tested using multivariate analysis of variance (MANOVA) and mean coverage values obtained for one 1 m2 quadrat (40 data/field type/collection dates) were considered. No significant difference in weed coverage were detected between years (P = 0.12). Therefore, data from the 2 years, collected on the same dates, were combined for the analyses. The weed frequency data were tested using Poisson-distributed errors residuals for normality of errors (Kolmogorov-Smirnov test) and for equality of variance (Levene’s test). Because residuals did not meet the assumption of normality, we used the non-parametric Kruskal-Wallis- and Mann-Whitney U test to compare variables. Weed species and management systems (HIF vs. LIF) were used as fixed factors and the average weed coverage in 1 m2 sub-transect as random factor.
We next determined how the cropping system differentially affected associational susceptibility to the two aphid species, B. helichrysi and A. solani. General linear modelling was used with mean aphid abundance on E. annua, E. canadensis and S. canadensis as response variable. Initial analyses indicated no difference between study years and aphid abundance averaged across study years (P < 0.23). The model included cropping system type (HIF vs. LIF), aphid species (B. helichrysi and A. solani), and their interaction as explanatory variables. Because aphid abundance is a discrete variable, Poisson-distributed errors were assessed. Aphid abundances on E. annua was normally distributed, so factorial ANOVA was used, followed by Tukey testing. Aphid abundance on E. canadensis and S. canadensis did not meet the assumption of normality, hence the Kruskal-Wallis test was used, followed by the Mann-Whitney U test.
Significant (P < 0.05) interactive effects (cropping system type × species) suggested that the effect of cropping system depended on aphid species. Aphid density analyses on crop plants were made considering only the abundance of B. helichrysi. As the data was normally distributed, ANOVA was used, followed by Tukey testing to compare abundances between treatments and crop plants. The density of A. solani was only high in potato, but there were no significant differences between treatments (T-test P = 0.78); therefore no other analyses at this species abundance on crops were made. All analyses were performed using R version 3.0.1 (R Core Team, 2013). Only a small number of other aphid species (e.g. Macrosiphum spp.) were detected, and we did not include them in the analyses.
Principal Components Analysis (PCA) was used to identify the proportion of variation in each PCA axis (Aphids density and treatments) explained by each virus distribution. We then used the average count of each virus reads numbers detected and log10 transformed from each weed and crop plant sample grouping as response variables, and used aphid abundance as component 1 (PCA axis1) and treatment (LIF vs. HIF) as component 2 (PCA axis 2) scores for each virus reads as independent variables. RNA and DNA viruses were analysed separately, and the only one insect virus detected (Helicoverpa zea nudivirus 2, HzNV-2) was not considered in our analyses. PCA covariance analyses were run using Community Analysis Package 4 (Pisces Conservation Ltd).
Virus diagnosis was determined by small RNA HTS. For bioinformatics analysis of the HTS results, we used CLC Genomic Workbench. Briefly: for trimming, quality control and QC reports, embedded protocols in CLC Genomic Workbench were employed. For virus diagnostics, we followed two strategies and used CLC Genomic Workbench: we built longer contigs from the non-redundant reads using assembler of CLC (de novo assembly) and compared the resultant contigs using BLAST to the NCBI Reference Genomes of plant viruses downloaded from GenBank. In parallel, we directly mapped contigs to Reference Genomes of those viruses which were represented at least with one contig in any of the libraries mapping to the reference tool of CLC Genomic workbench. Virus presence was recorded if at least two parameters were reached, i.e. virus specific contig was present and/or normalized redundant virus specific read count was >200, and/or coverage of the virus genome was >60%.
Results
Dominant Invasive Weed Species and Their Variations Between Management Systems
Three weed species, all considered invasive, were dominant during the 2 years field assessment. Erigeron annua was the most frequent, dominating both LIF (97.5%) and HIF treatment regimes (84.5%). Two other species were present at lower densities. S. canadensis was only present in LIF, with a coverage of 2.5%. No other invasive weeds were assessed under this management system. E. canadensis was only present under HIF with a coverage of 15%. Other weed species, mostly amaranth, Amaranthus spp. in HIF regimes with an average coverage of 0.5%, were observed at the end of the growing period of the above mentioned weed species. Dominance of E. annua was significant under both management systems (Figure 1).
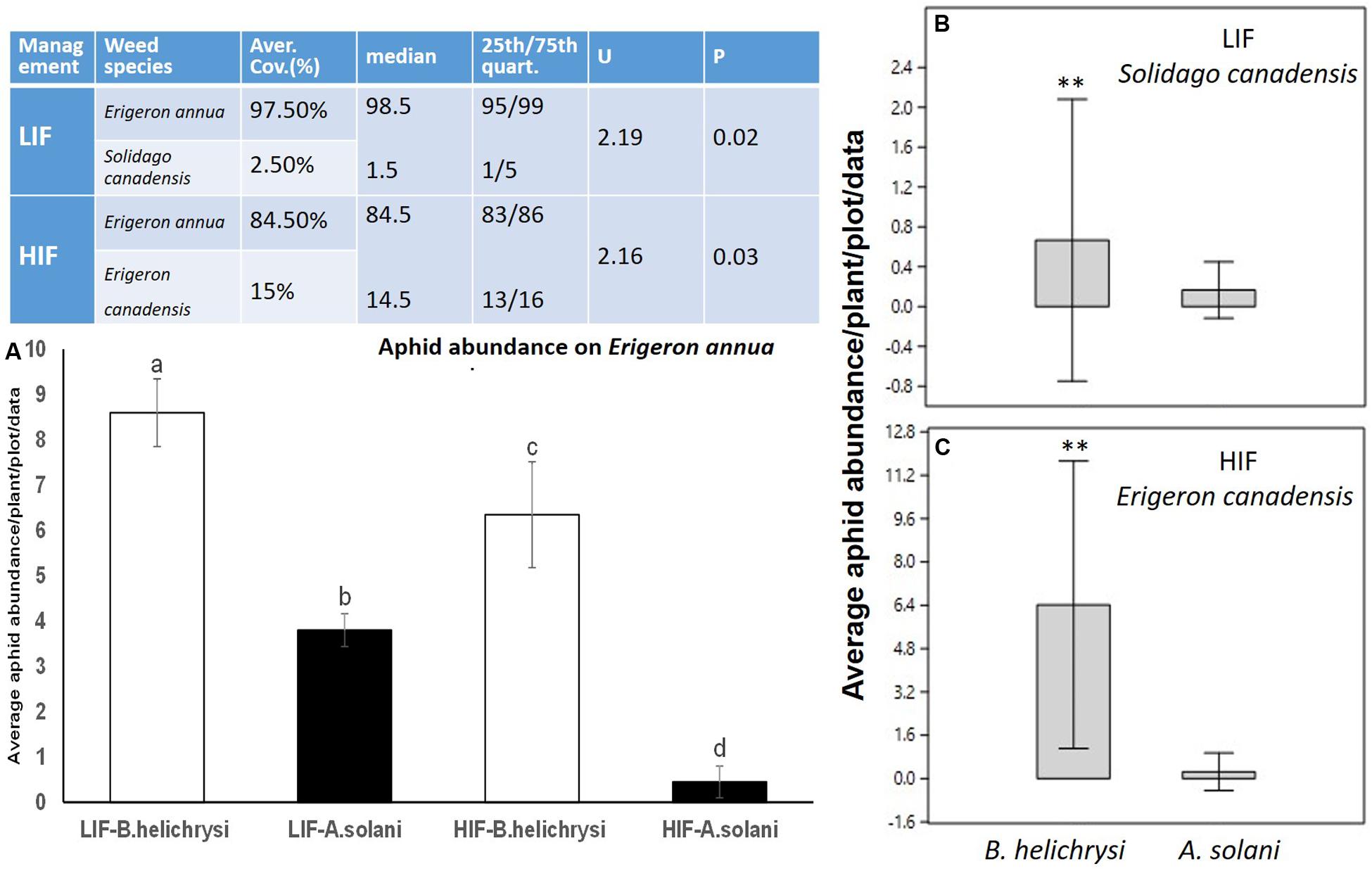
Figure 1. The coverage of the most frequent weed species between LIF and HIF regimes (Table) and the abundance of the most frequent aphid species between treatments (A) and weed plants (B,C). Weed data were compared using non-parametric Kruskal-Wallis- and Mann-Whitney U tests. Aphid abundance on E. annua was normally distributed, so factorial ANOVA was used, followed by Tukey testing. Aphid abundance on E. canadensis and S. canadensis did not meet the assumption of normality, hence the Kruskal-Wallis test was used, followed by the Mann-Whitney U test. Significance level = P < 0.05. ∗∗P < 0.01. Different letters (a–b) means statistical significant difference.
Aphids and Their Abundances on Invasive Weeds and Crop Plants
Two native aphid species were detected at high density on all three dominant invasive weeds. The most frequent was B. helichrysi, which infested the most frequent weed, E. annua, under both management regimes (LIF-B. helichrysi and LIF-A. solani F1–40 = 6.4, P < 0.001; LIF-B. helichrysi and HIF-A. solani F1–40 = 8.1, P < 0.001) (Figure 1A). The next most abundant species was A. solani, also present on E. annua plants under both management systems; its density was significantly higher under LIF compared with HIF (F1–40 = 8, P < 0.001) (Figure 1A). A higher density of B. helichrysi was observed on S. canadensis under LIF (U1–40 = 3.4, P < 0.01) (Figure 1B). Furthermore, the dominance of B. helichrysi on E. canadensis was observed under HIF conditions (U1–40 = 3.1, P < 0.01) (Figure 1C). A very low number of other aphid species were observed, i.e. about 12 individuals of Macrosiphum spp. were collected on S. canadensis. The abundance of B. helichrysi was higher in maize under LIF compared with HIF (F1–40 = 4.5, P < 0.01). No other differences between treatments were observed in alfalfa (F1–40 = 0.5, P < 0.89) and potato (F1–40 = 0.2, P < 0.91) (Figure 2).
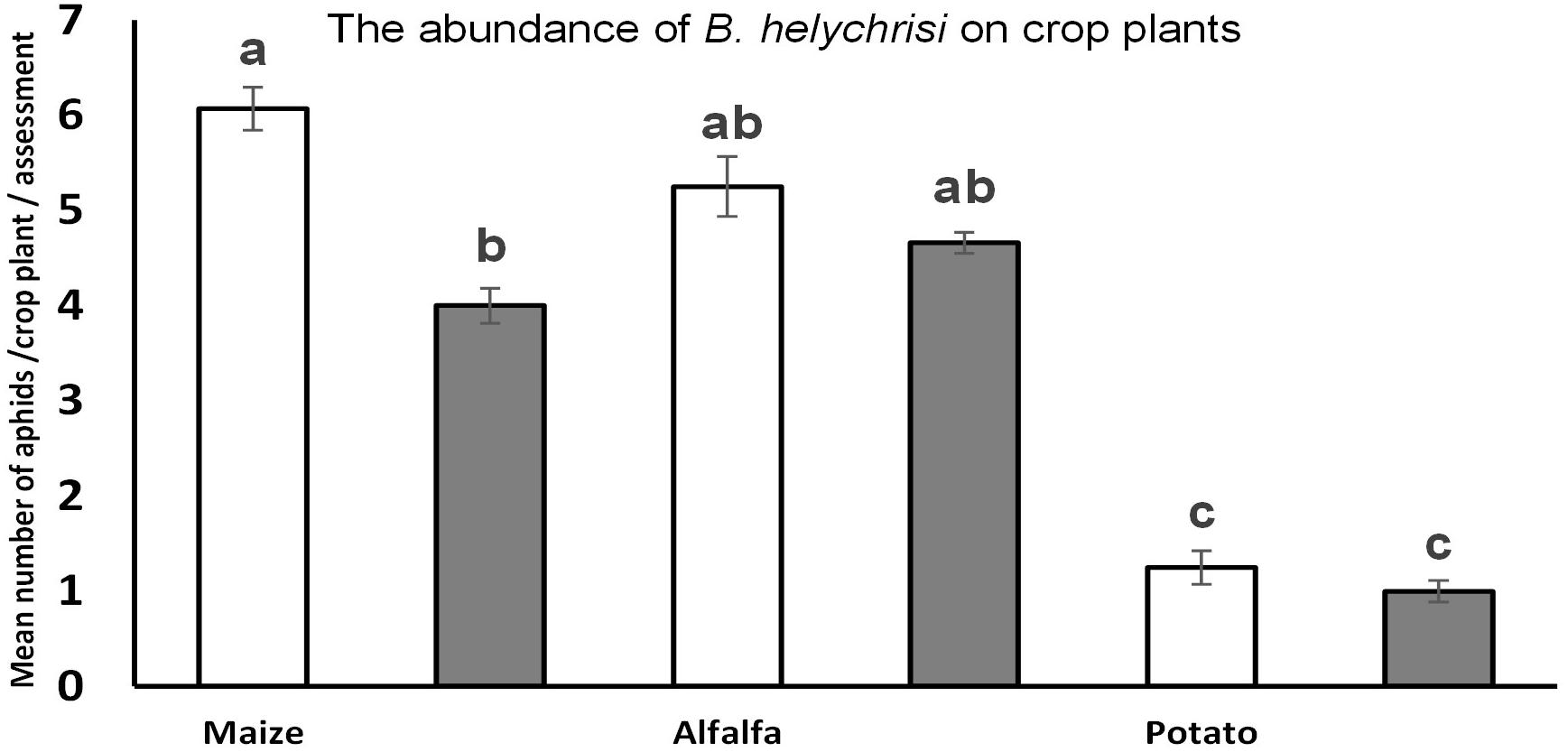
Figure 2. The abundance of B. helychrisi on crop plants under different treatments (HIF vs. LIF). ANOVA was used followed by Tukey testing to compare variables. Significance level = P < 0.05. Error bars represent standard errors. Different letters (a–b) means statistical significant difference.
Plant Viruses in Invasive Weeds and Crop Plants
Sequencing of the small RNA libraries resulted in 9.4–21 million raw reads (Supplementary Table S3). After trimming and quality control we obtained 9.2–20.4 million reads, which represented 2,019,811–7,192,941 individual sequences. Using overlapping stretches of these reads, we were able to construct de novo 3,553–19,038 longer contigs (Supplementary Table S3). Virus derived contigs were annotated following BLAST searches. Thereafter, we made direct mapping of the sequenced trimmed reads to each viral reference genome, which gave at least one hit according to contig annotation and counting all redundant and non-redundant reads derived from a given virus. To compare the number of reads in different libraries, normalized redundant reads (read/million read) were calculated. According to this approach the presence of 42 different viruses were detected.
Annotation can lead to false positive results in familiar viruses (Massart et al., 2019). Moreover, as we used pools for small RNA HTS, RNA from the non-infected plants could dilute the sample for virus specific reads – for example, a high, normalized, redundant read count could be counted even without the presence of the virus specific contig. Hence, the presence of the virus was counted only when the presence of the contig(s) coincided with a relatively high (>200) normalized redundant read and >>60% coverage of the genome.
Following this revised analytical approach, the number of viruses detected dropped to 16, differentiated as nine RNA and seven DNA viruses (Table 1, Figure 3, and Supplementary Table S4). The distribution of both RNA and DNA viruses varied greatly between crops and invasive weeds, both under LIF and HIF regimes. The most widespread virus detected was the insect baculovirus HzNV-2, which may have originated from larvae of the highly polyphagous corn earworm moth, Helicoverpa zea (Lepidoptera: Noctuidae) feeding in the fields sampled. Of plant viruses, PVY and TVCV were the most widespread, only lacking from library 6_SS and 2_TB, respectively. Crops grown under LIF contained nine viruses, while under HIF, only six were found. PVX and BLRV were present only in crops under HIF, while PVM and PBCoV were only present in LIF crops (Table 1, Figure 3, and Supplementary Table S4).
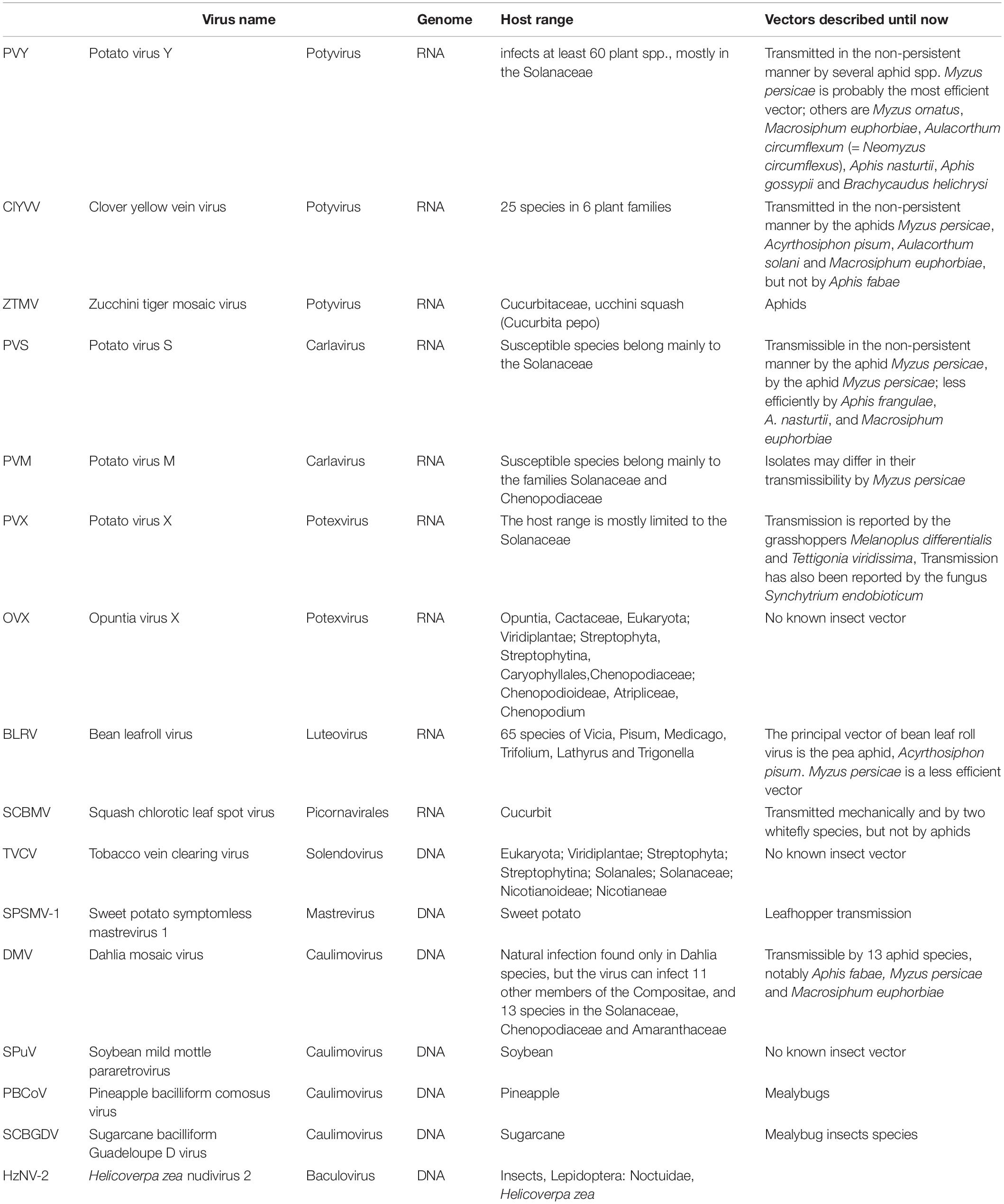
Table 1. The list of the 16 viruses detected in invasive weeds and associated crop plants using mild parameters in the bioinformatics analysis.
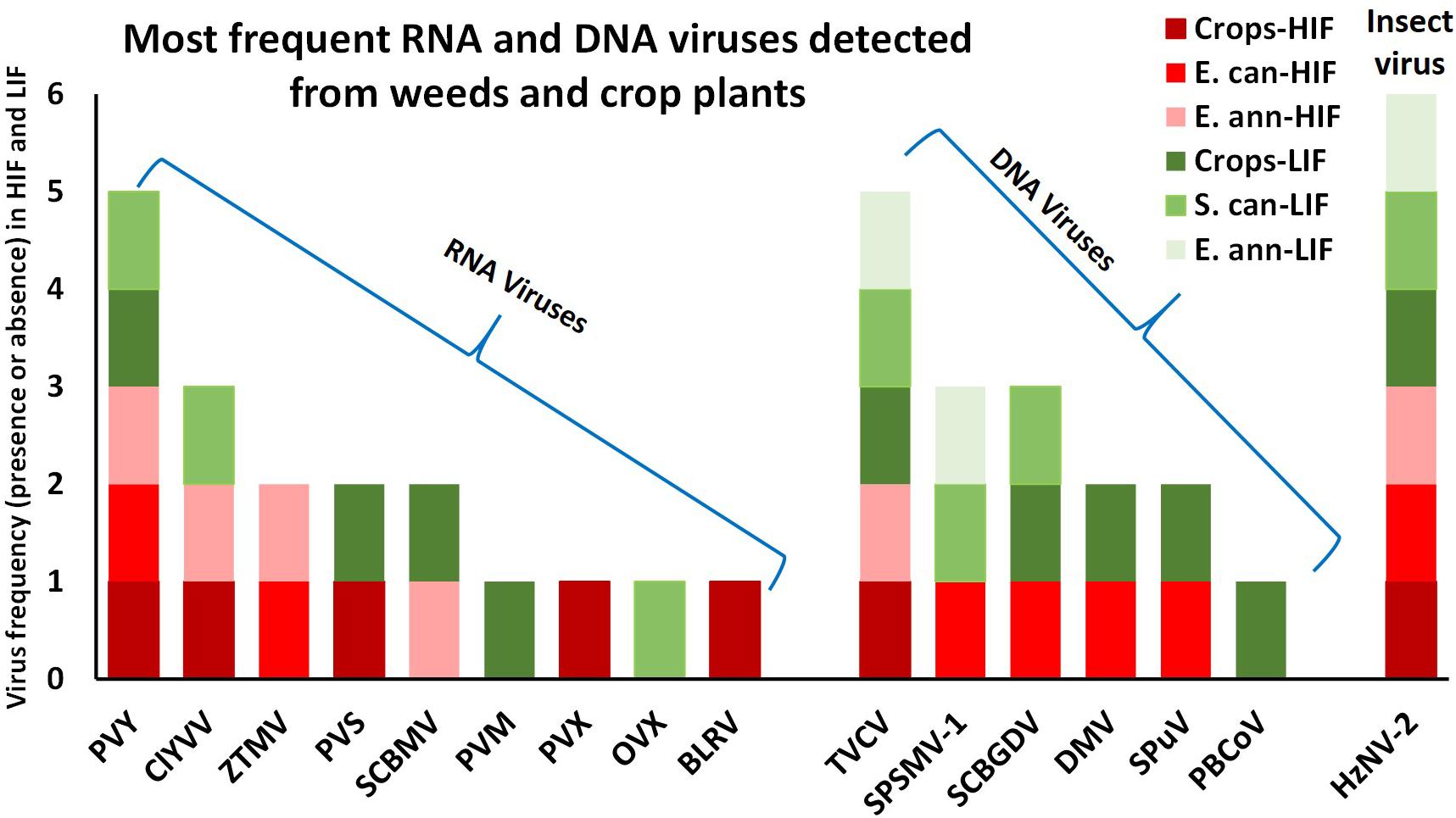
Figure 3. The presence of RNA and DNA viruses between crops and invasive weeds under low and high input field regimes. Red colours represents HIF, green colours represents LIF. E. ann - Erigeron annua, C. can - Conyza canadensis, S. can - Solidago canadensis. PVY, Potato virus Y; ClYVV, Clover yellow vein virus; ZTMV, Zucchini tiger mosaic virus; PVS, Potato virus S; SCBMV, Squash chlorotic leaf spot virus; PVM, Potato virus M; PVX, Potato virus X; OVX, Opuntia virus X; BLRV, Bean leafroll virus; TVCV, Tobacco vein clearing virus; SPSMV-1, Sweet potato symptomless mastrevirus 1; SCBGDV, Sugarcane bacilliform Guadeloupe D virus; DMV, Dahlia mosaic virus; SPuV, Soybean mild mottle pararetrovirus; PBCoV, Pineapple bacilliform comosus virus; HzNV-2,- Helicoverpa zea nudivirus 2.
The presence of potato infecting viruses were also investigated with independent methods, RT-PCR and Northern blot. Under HIF conditions, only PVX was detected, while under LIF, PVY, PVS, PVM were validated (Supplementary Table S5 and Supplementary Figure S2). Although we obtained many small RNA reads mapped to PVY and PVS, we could not validate their presence under HIF conditions (Supplementary Figure S2). It is possible that the antiviral silencing was very active in these circumstances, which explains why we detected the small RNAs, but not the entire viruses, the titre of which had declined. However, it is clear from the RT-PCR and Northern blot results that while viruses vectored by aphids (PVY, PVS, and PVM) are present under LIF conditions, PVX, which is mainly mechanically transmitted, is not, and instead was only detected in HIF regimes.
Several viruses detected during the present study have not been previously reported or have been rarely mentioned by the European and Mediterranean Plant Protection Organization, EPPO (Supplementary Table S6). In the case of the Opuntia virus X, its insect vector has not yet been described. No reports by EPPO concerning Tobacco vein clearing virus from Europe exist, whereas we detected this virus at high frequency in both crops and weeds under LIF and HIF regimes (Table 1 and Figure 3). This pararetrovirus was first described from a hybrid form of tobacco, Nicotiana edwardsonii (Lockhart et al., 2000), and found to be highly integrated into the host genome. There is still no additional TVCV deposited into GenBank. We made an effort in this respect and tried to amplify the virus in our samples. We obtained some product of the expected size, but subsequent Sanger sequence analyses showed that this arose from the host genome. We believe that such detection is a false positive and that we apparently had hits because some of the plant genomes host this type of retroviral element. The Sweet potato symptomless mastrevirus 1 is also absent from any EPPO alert lists, whereas we detected it in invasive weeds, but not in crops. The Pineapple bacilliform comosus virus has been reported from tropical areas, while we detected its presence at high read numbers in crops under LIF. The Sugarcane bacilliform Guadeloupe D virus, reported as frequent in the tropics, was detected in crops under LIF, but also in weeds, i.e. E. canadensis under HIF and S. canadensis under LIF. The Lepidoptera-infecting Helicoverpa zea nudivirus 2 has not been officially reported from Europe, and hence is not present in the EPPO listings. Its frequency was the highest in all crops and weeds under all management systems (Table 1 and Figure 3).
Using PCA to identify the proportion of variation in terms of aphid density and treatments explaining RNA and DNA virus distribution, we observed that aphid abundance was the most important factor governing RNA virus distribution (40%), whilst treatments (LIF vs. HIF) had only a smaller effect (26%) (Figure 4A). In contrast, DNA virus distribution was mostly determined by treatment (54%) rather than aphid distributions (29%) (Figure 4B).
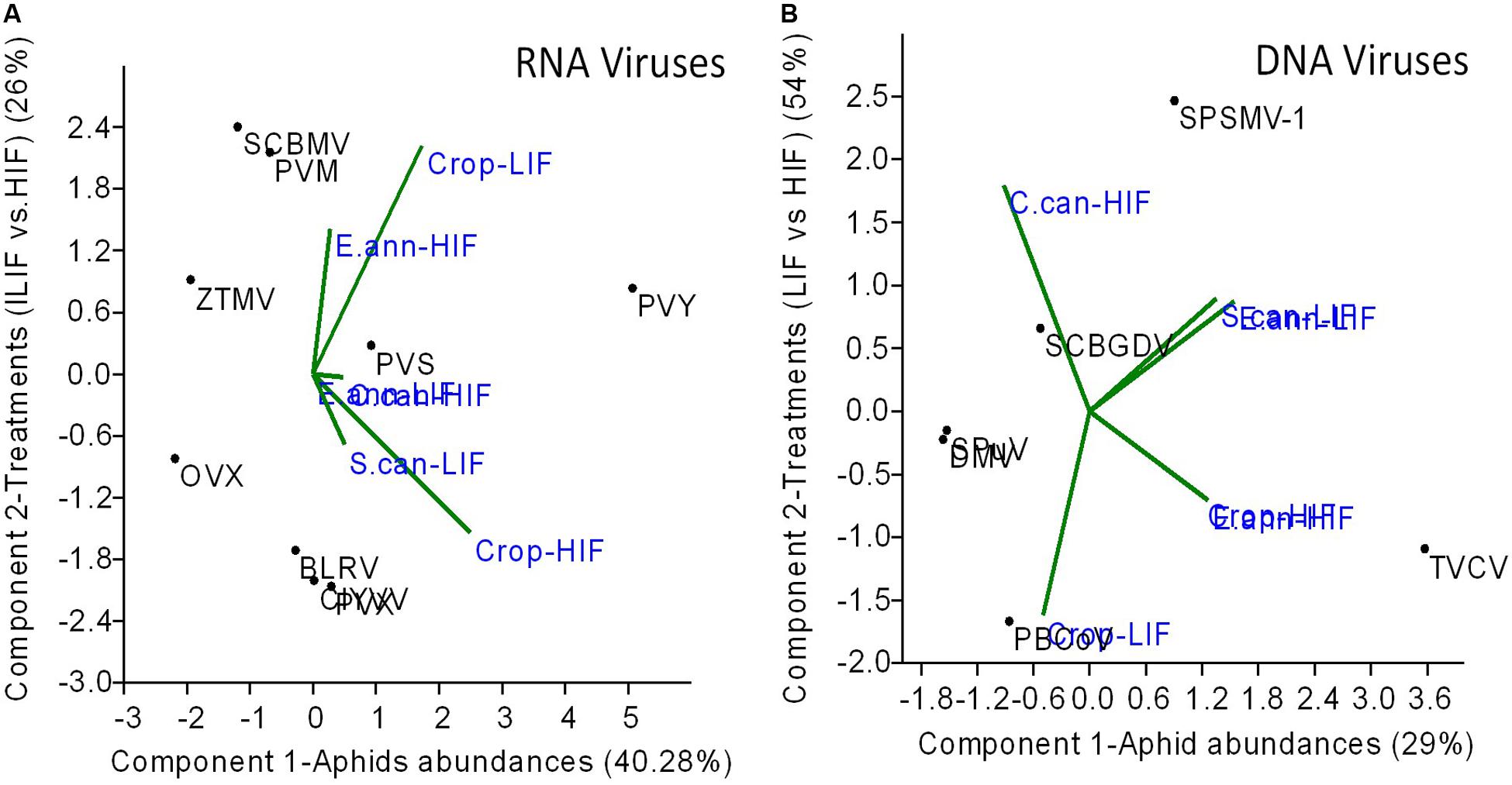
Figure 4. The effect of treatments (HIF vs. LIF and aphid B. helichrysi and A. solani) abundance on RNA (A) and DNA (B) virus distributions between crop plants and weeds. The only insect virus detected (HzNV-2) was not considered in the analyses. Principal Components Analysis (PCA) was used and the average count of each virus read from each weed and crop plant sample grouping were treated as response variables, and aphids abundance as component 1 (PCA axis1) and treatment (LIF vs. HIF) as component 2 (PCA axis 2) scores, related to each virus read as independent variables. RNA and DNA viruses were separately analysed.
Discussion
According to our present findings, it is clear that associational susceptibility exists between the most frequent invasive weed and crop plants under different crop management (LIF vs. HIF) regimes. As determined experimentally, a 13% higher coverage difference of E. annua in LIF further resulted in a significantly higher B. helichrysi abundance (about 30% more). The same trend was observed for A. solani, where a 13% higher coverage of E. annua resulted in an increase of about 85% for this aphid under LIF compared to HIF regimes (Figure 1). In our previous experiments testing colonization abilities of these particular aphids (Szabó et al., 2019), it was also shown that E. annua and E. canadensis are suitable weed hosts for both species but especially for B. helichrysi. Significant colonization of B. helichrysi from both weeds toward the crop plants maize, alfalfa, and potato was detected (Szabó et al., 2019). Therefore, virus transmission from invasive weeds to crop plants by these aphids is highly probable.
Several factors appear to influence associational susceptibility between invasive weeds and crop plants. The likelihood of detection of neighbouring plants, and the factors that can directly affect the survival of local aphid populations on these plants, seemingly include natural habitat diversity as well as lack of any chemical usage under the LIF regime. The short distance between weed populations and crop plants (here alfalfa, maize, and potato) and the high habitat diversity apparently may influence associational susceptibility of crop plants, and the presence of the two aphid species, especially B. helichrysi, can be explained in all three crop plants (Figure 2).
In addition, aphid abundance and the management systems used may directly influence plant virus distribution between weeds and crop plants, while RNA virus distribution (mostly aphidophag viruses transmitted through stylet-borne mechanisms) is probably more influenced by aphid density per se, and DNA virus distribution (transmitted less by aphids, probably more by mechanical means) between weeds and crops by contrast predominantly influenced by the management system involved (Figures 4A,B). While we made substantial efforts to extract and detect viruses from aphid stylets collected from weeds and crop plants during assessment and thereby confirm direct transmission, we were unsuccessful. This is undoubtedly due to two principal reasons: (1) The technique used may be fundamentally unsuited for virus detection from aphid stylets or (2) it may simply be that the titre of virus within the aphid stylets is very low, whereas in the plant itself, it is very much amplified, allowing detection. The previous colonization experiment (Szabó et al., 2019) clearly demonstrated that with both aphid species here studied, virus transmission definitely occurred during the insects’ movement from weeds to crop plants and the colonization of these.
Besides aphid-transmitted viruses, another important group of plant pathogenic viruses comprises those transmitted mechanically or indeed, by unknown means. In the case of PVX virus, as here presently found only in HIF regimes, if the tubers are infected, cultivators can readily disseminate it within and between potato crops. With PVM, it is usually present in herbaceous Solanaceous plants within the surrounding flora. We didn’t detect it in any of the Composite weeds sampled and tested, probably because they cannot host it. However, it was only present in the LIF regimes, suggesting that it could be transmitted by the sap sucking herbivorous insects like aphids feeding on these weeds. The same could happen with BLRV and PBCoV: since the Composite weeds do not host them, we hence failed to detect them in the libraries.
DNA viruses were detected in the crops within the LIF regimes, and contrastingly were present in weeds in both HIF and LIF, but here revealed a different pattern of infection (Supplementary Tables S2, S3 and Figure 3). This result is likely a consequence of the fact that weeds belonging to the family Compositae can host DNA viruses, which cannot be hosted by the crops investigated, such that the pattern of infection is further altered, a possibility that we naturally have had to take into consideration.
Lastly, several of the viruses detected in our study have not previously been reported from Europe, i.e. Opuntia virus X, Tobacco vein clearing virus, Sweet potato symptomless mastrevirus 1, Pineapple bacilliform comosus virus, Sugarcane bacilliform Guadeloupe D virus and the Helicoverpa zea nudivirus 2. Although our detection of these viruses by small RNA HTS seems credible, further studies involving a different approach (e.g. RT-PCR) need to be performed in order to add support to these findings. What is certain is that the presence of these viruses in both invasive weeds and crop plants without the production of pathological symptoms in the former raises the clear necessity of acknowledging the potential and indeed likely presence of such pathogenic disease reservoirs during invasive weed management scenarios.
Conclusion
Overall, we conclude from these findings that even if low-input management farming systems are widely studied (e.g. Akeroyd and Page, 2011; Fischer et al., 2012; Mikulcak et al., 2013) and are supposedly low-cost, effective systems (i.e. no or low management costs) with high biodiversity and cultural values (Hartel et al., 2013), the abandonment or absence of management may cause serious problems. This is mainly due to the likelihood of increased virus vectoring aphid densities, including of hitherto unknown or known but unrecorded viruses, which further affect local cultivated plants, and may in addition influence local wild flora and their associated wildlife (e.g. pollinators) in unpredicted ways. Damage produced by agricultural management of the agro-ecosystem, and indeed also in general environmental management, may overcome the costs of any environmentally-friendly weed control. Therefore, new weed management systems and assessment methods are necessary to evaluate the importance of weeds as virus reservoirs, even under low management regimes. From the standpoint of aphid control and associated virus transmission, the complete lack of any management needs to be seriously reconsidered, more especially the virus vectoring capacity of some aphid species like A. solani, which is highly polyphagous. Because of the unpredictable high local aphid density on invasive weeds, aphid migration onto several local crops (e.g. potato, considered as both a low-cost and low-input crop) may potentially cause — and indeed probably does often cause — unpredictable virus infections, including under low-input management regimes. From this we deduce that cultivation methods involving invasive weed and insect vector control need to be reconsidered, even when and if no other management approach is planned. To this end, further research is planned to quantitatively test viral infestation and detect virus spread via weed-aphid-crop plants.
Whilst this pilot present study is not definitive in that individual virus vectoring aphids were not directly tested, nevertheless it represents one of the first contributions to this particular topic area. As such, the study details the pioneering nature of this broad approach, one that shows considerable promise in attempts to understand the role of weed reservoirs in weed-crop aphid borne virus transmission, thereby paving the way for the next publications planned from our group in this fascinating area of applied research.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Ethics Statement
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. The manuscript does not contain any studies with human participants performed by any of the authors.
Author Contributions
A-KS, AB, and JK perceived and designed the experiment. ÉV, AH, and ED performed the small RNA HTS. A-KS, JB, A-KS, and AB performed the data collections. ZG and ED performed the RT-PCR. AH performed the Northern blot analysis. A-KS, AB, and HL performed the data analyses, and wrote the manuscript.
Funding
This research was supported by the Higher Education Institutional Excellence Program (1783-3/2018/FEKUTSTRAT) awarded by the Ministry of Human Capacities within the framework of plant breeding and plant protection researches of Szent István University and by the Institute of Research Programs of the Sapientia Hungarian University of Transylvania grant No. 21/2/12.06.2019.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are most grateful to all those who made the various scientific assessments as here described possible, especially local people.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2020.00684/full#supplementary-material
References
Abhilasha, D., Quintana, N., Vivanco, J., and Joshi, J. (2008). Do allelopathic compounds in invasive Solidago canadensis s.l. restrain the native European flora? J. Ecol. 96, 993–1001. doi: 10.1111/j.1365-2745.2008.01413.x
Agrow (2015). World Crop Protection News - Biopesticides Specials. Available online at: https://agrow.agribusinessintelligence.informa.com (accessed June 25, 2019).
Akeroyd, J., and Page, N. (2011). Conservation of high nature value (HNV) grassland in a farmed landscape in Transylvania, Romania. Contribu. Bot. 46, 57–71.
Anastasiu, P., and Negrean, G. (2005). Alien plants in Romania (I). Analele Stiintifice ale Universitatii “Al. I. Cuza” din Iasi Tomul LI, s. II a. Biol. Veget. 51, 87–96.
Andújar, D., Ribeiro, A., Carmona, R., Fernández-Quintanilla, C., and Dorado, J. (2010). An assessment of the accuracy and consistency of human perception of weed cover. Weed Res. 50, 638–647. doi: 10.1111/j.1365-3180.2010.00809.x
Bajwa, A. A., Sadia, S., Ali, H. H., Jabran, K., Peerzada, A. M., and Chauhan, B. S. (2016). Biology and management of two important Conyza weeds: a global review. Environ. Sci. Pollut. Res. 23, 24694–24710. doi: 10.1007/s11356-016-7794-7
Barbosa, P., Hines, J., Kaplan, I., Martinson, H., Szczepaniec, A., and Szendrei, Z. (2009). Associational resistance and associational susceptibility: having right or wrong neighbours. Annu. Rev. Ecol. Evol. Syst. 40, 1–20. doi: 10.1146/annurev.ecolsys.110308.120242
Bell, A. C. (1983). The life-history of the leaf-curling plum aphid Brachycaudus helichrysi in Northern Ireland and its ability to transmit potato virus YC(AB). Ann. Appl. Biol. 102, 1–6. doi: 10.1111/j.1744-7348.1983.tb02660.x
Blackman, R. (2010). Aphids - Aphidinae. Royal Entomological Society. Shrewsbury: Field Studies Council.
Blackman, R. L., and Eastop, V. F. (2000). Aphids on the World’s Crops: An Identification and Information Guide, 2nd Edn. Chichester: Wiley.
Chitsaz, M., and Nelson, D. C. (1983). Comparison of various weed control programs for potatoes. Am. Potato J. 60, 271–280. doi: 10.1007/BF02854277
Czotter, N., Molnár, J., Pesti, R., Demián, E., Baráth, D., Varga, T., et al. (2018). “Use of siRNAs for diagnosis of viruses associated to woody plants in nurseries and stock collections,” in Viral Metagenomics: Methods and Protocols, eds V. Pantaleo and M. Chiumenti (New York, NY: Springer), 115–130. doi: 10.1007/978-1-4939-7683-6_9
Edwards, P. J., Frey, D., Bailer, H., and Baltisberger, M. (2006). Genetic variation in native and invasive populations of Erigeron annuus as assessed by RAPD markers. Int. J. Plant Sci. 167, 93–101. doi: 10.1086/498729
EPPO (2017). Available online at: https://www.eppo.int/activities/iap_activities (accessed June 25, 2019).
Fenesi, A., Geréd, J., Meiners, S. J., Tóthmérész, B., Török, P., and Ruprecht, E. (2015). Does disturbance enhance the competitive effect of the invasive Solidago canadensis on the performance of two native grasses? Biol. Invas. 17, 3303–3315. doi: 10.1007/s10530-015-0954-8
Fischer, J., Hartel, T., and Kuemmerle, T. (2012). Conservation policy in traditional farming landscapes. Conserv. Lett. 5, 167–175. doi: 10.1111/j.1755-263X.2012.00227.x
Frey, D., Baltisberger, M., and Edwards, P. (2003). Cytology of Erigeron annuus s.l. and its consequences in Europe. Bot. Helvet. 113, 1–14. doi: 10.1007/s00035-003-0676-x
Hartel, T., Dorresteijn, I., Klein, C., Máthé, O., Moga, C. I., Öllerer, K., et al. (2013). Wood-pastures in a traditional rural region of Eastern Europe: characteristics, management and status. Biol. Conserv. 166, 267–275. doi: 10.1016/j.biocon.2013.06.020
Isac, M., Preda, S., and Marcu, M. (1998). Aphid species-vectors of plum pox virus. Acta Virol. 42, 233–234.
Jandricic, S. E., Mattson, N. S., Wraight, S. P., and Sanderson, J. P. (2014). Within-plant distribution of Aulacorthum solani (Hemiptera: Aphididae), on various greenhouse plants with implications for control. J. Econ. Entomol. 107, 697–707. doi: 10.1603/EC13428
Jandricic, S. E., Wraight, S. P., Bennett, K. C., and Sanderson, J. P. (2010). Developmental times and life table statistics of Aulacorthum solani (Hemiptera: Aphididae) at six constant temperatures, with recommendations on the application of temperature-dependent development models. Environ. Entomol. 39, 1631–1642. doi: 10.1603/EN09351
Lockhart, B. E., Menke, J., Dahal, G., and Olszewski, N. E. (2000). Characterization and genomic analysis of tobacco vein clearing virus, a plant pararetrovirus that is transmitted vertically and related to sequences integrated in the host genome. J. Gen. Virol. 81, 1579–1585. doi: 10.1099/0022-1317-81-6-1579
Mabuza, M. L., Sithole, M. M., Wale, E., Ortmann, G. F., and Darroch, M. A. G. (2013). Factors influencing the use of alternative land cultivation technologies in Swaziland: implications for smallholder farming on customary Swazi Nation land. Land Use Policy 33, 71–80. doi: 10.1016/j.landusepol.2012.12.009
Massart, S., Chiumenti, M., De Jonghe, K., Glover, R., Haegeman, A., Koloniuk, I., et al. (2019). Virus detection by high-throughput sequencing of small RNAs: large-scale performance testing of sequence analysis strategies. Phytopathology 109, 488–497. doi: 10.1094/phyto-02-18-0067-r
Mikulcak, F., Newig, J., Milcu, A. I., Hartel, T., and Fischer, J. (2013). Integrating rural development and biodiversity conservation in Central Romania. Environ. Conserv. 40, 129–137. doi: 10.1017/S0376892912000392
Nandula, V. K., Eubank, T. W., Poston, D. H., Koger, C. H., and Reddy, K. N. (2006). Factors affecting germination of horseweed (Conyza canadensis). Weed Sci. 54, 898–902. doi: 10.1614/WS-06-006R2.1
Pacanoski, Z. (2017). Current situation with invasive Erigeron annuus (L.) Pers. (daisy fleabane) in the Republic of Macedonia. EPPO Bull. 47, 118–124. doi: 10.1111/epp.12368
Popkin, M., Piffaretti, J., Clamens, A.-L., Qiao, G.-X., Chen, J., Vitalis, R., et al. (2017). Large-scale phylogeographic study of the cosmopolitan aphid pest Brachycaudus helichrysi reveals host plant associated lineages that evolved in allopatry. Biol. J. Linn. Soc. 120, 102–114. doi: 10.1111/bij.12869
Powell, G., Harrington, R., and Spiller, N. J. (1992). Stylet activities and potato virus Y vector efficiencies by the aphids Brachycaudus helichrysi and Drepanosiphum platanoidis. Entomol. Exp. Appl. 62, 293–300. doi: 10.1111/j.1570-7458.1992.tb00670.x
R Core Team (2013). A Language and Environment For Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rand, T. A., and Louda, S. M. (2004). Exotic weed invasion increases the susceptibility of native plants to attack by a biocontrol herbivore. Ecology 85, 1548–1554. doi: 10.1890/03-3067
Roossinck, M. J., Martin, D. P., and Roumagnac, P. (2015). Plant virus metagenomics: advances in virus discovery. Phytopathology 105, 716–727. doi: 10.1094/phyto-12-14-0356
Shah, M. A., Callaway, R. M., Shah, T., Houseman, G. R., Pal, R. W., Xiao, S., et al. (2014). Conyza canadensis suppresses plant diversity in its non-native ranges but not at home: a transcontinental comparison. New Phytol. 202, 1286–1296. doi: 10.1111/nph.12733
Steffan-Dewenter, I., Münzenberg, U., and Tscharntke, T. (2001). Pollination, seed set and seed predation on a landscape scale. Proc. R. Soc. B Biol. Sci. 268, 1685–1690. doi: 10.1098/rspb.2001.1737
Szabó, A.-K., Kiss, J., Bálint, J., Kõszeghi, S., Loxdale, H. D., and Balog, A. (2019). Low and high input agricultural fields have different effects on pest aphid abundance via different invasive alien weed species. NeoBiota 43, 27–45. doi: 10.3897/neobiota.43.31553
Tatchell, G. M., Parker, S. J., and Woiwod, I. P. (1983). “Synoptic monitoring of migrant insect pests in Great Britain and western Europe IV. Host plants and their distribution for pest aphids in Great Britain,” in Annual Report of Rothamsted Experimental Station, 1982, Part 2, eds I. P. Woiwod and L. R. Taylor (Harpenden: Rothamsted Experimental Station), 45–159.
Tunaitienė, V., Naugžemys, D., Patamsytė, J., and Žvingila, D. (2015). Gradient of genetic diversity of Erigeron annuus in the part of invasive European range. Bot. Lithuan. 21, 81–88. doi: 10.1515/botlit-2015-0011
Uchino, H., Iwama, K., Jitsuyama, Y., Ichiyama, K., Sugiura, E., Yudate, T., et al. (2012). Effect of interseeding cover crops and fertilization on weed suppression under an organic and rotational cropping system: 1. Stability of weed suppression over years and main crops of potato, maize and soybean. Field Crops Res. 127, 9–16. doi: 10.1016/j.fcr.2011.10.007
Usda Forest Service (2015). Invasive Species Activities. Available online at: https://www.fs.fed.us/invasivespecies/controlmgmt/activities.shtml (accessed March 17, 2020).
White, J. L., and Kaper, J. M. (1989). A simple method for detection of viral satellite RNAs in small plant tissue samples. J. Virol. Methods 23, 83–93. doi: 10.1016/0166-0934(89)90122-5
Keywords: invasive weeds, virus vector, aphids, crops, cropping systems, small RNA, HTS
Citation: Szabó A-K, Várallyay E, Demian E, Hegyi A, Galbács ZN, Kiss J, Bálint J, Loxdale HD and Balog A (2020) Local Aphid Species Infestation on Invasive Weeds Affects Virus Infection of Nearest Crops Under Different Management Systems – A Preliminary Study. Front. Plant Sci. 11:684. doi: 10.3389/fpls.2020.00684
Received: 26 August 2019; Accepted: 30 April 2020;
Published: 25 June 2020.
Edited by:
Calum Rae Wilson, University of Tasmania, AustraliaReviewed by:
Anna-Maria Botha-Oberholster, Stellenbosch University, South AfricaAri Mikko Hietala, Norwegian Institute of Bioeconomy Research (NIBIO), Norway
Copyright © 2020 Szabó, Várallyay, Demian, Hegyi, Galbács, Kiss, Bálint, Loxdale and Balog. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Attila-Károly Szabó, c3phYm9fYXRhQHlhaG9vLmNvbQ==; Adalbert Balog, YWRhbGJlcnQuYmFsb2dAbXMuc2FwaWVudGlhLnJv; YWRhbGJlcnQuYmFsb2dAZ21haWwuY29t