 Petia Simeonova Nikolova1,2*
Petia Simeonova Nikolova1,2* Taryn L. Bauerle3
Taryn L. Bauerle3 Karl-Heinz Häberle2
Karl-Heinz Häberle2 Helmut Blaschke2
Helmut Blaschke2 Ivano Brunner4Rainer Matyssek2
Ivano Brunner4Rainer Matyssek2- 1Forest Resources and Management, Swiss Federal Research Institute WSL, Birmensdorf, Switzerland
- 2Ecophysiology of Plants, TUM School of Life Sciences, Technische Universität München, Freising, Germany
- 3School of Integrative Plant Science, Cornell University, Ithaca, NY, United States
- 4Forest Soils and Biogeochemistry, Swiss Federal Research Institute WSL, Birmensdorf, Switzerland
Trees adjust multiple structural and functional organ-specific characteristics, “traits”, to cope with diverse soil conditions. Studies on traits are widely used to uncover ecological species adaptability to varying environments. However, fine-root traits are rarely studied for methodological reasons. We analyzed the adaptability of the fine-root systems of European beech and Norway spruce to extreme drought within species-specific tree groups at Kranzberger Forst (Germany), focusing on the seasonality of morphological, physiological, and biochemical key traits in view of carbon (C) and nitrogen dynamics. We hypothesized that fine roots of both species adjust to seasonal drought: with beech representing a “fast” (i.e. with fast C turnover), and spruce a “slow” (i.e. with long-term C retention) ecological strategy. We identified three functional fine-root categories, based on root function (absorptive or transport fine roots), and mycorrhizal status of the absorptive fine-roots (mycorrhizal or non-mycorrhizal). Solely the non-mycorrhizal absorptive roots adjusted in a species-specific manner supporting fine-root ecological strategy hypothesis. During drought, beech produced thin ephemeral (absorptive non-mycorrhizal) fine roots with high specific fine-root area and high respiratory activity, representing fast C turnover and enabling effective resource exploitation. These adjustments reflect a “fast” ecological strategy. Conversely, spruce absorptive fine roots did not respond to the soil moisture deficit by growth but instead increased root suberization. Drastically lowered respiratory activity of this functional category facilitated C retention and structural persistence during drought, indicating a “slow” ecological strategy in spruce. Absorptive mycorrhizal fine roots maintained respiration throughout the drought event in both tree species, but in spruce this was the only fine-root category with high respiration. This suggests, that spruce relies heavily on mycorrhizal associations as a method of drought resistance. Accumulation of non-structural carbohydrates and high C concentrations were observed in the transport fine roots of both species, indicating drought-induced osmotic protection of these roots. Thus, functional classification enabled us to determine that fine-root branches of each species are not tied to one sole ecological strategy. The suggested approach helps to better understand the complex interplay between structure and function belowground.
Introduction
Plants and especially long-lived trees have evolved a variety of structural and functional characteristics (“traits”) both above- and belowground to optimize the use of heterogeneous spatiotemporal resources (Lavorel and Garnier, 2002; Rennenberg et al., 2006; Freschet et al., 2018). Analyses of plant traits make it possible to decipher species-specific trade-offs in adapting to resource limited site conditions (Iversen et al., 2017; McCormack et al., 2017; Brunner et al., 2019). Belowground traits include root morphology and physiology along with mycorrhizal associations in relation to seasonal and soil variation (Laliberté, 2017). Although more than 300 root traits have been identified across individual studies (Iversen et al., 2017), they remain underrepresented in global trait databases (Ma et al., 2018). Frequently, the lack of root trait data is a direct result of methodological sampling difficulty (Joslin et al., 2000; Pregitzer, 2002; Brunner et al., 2015), challenges in integrating outcomes from varying environments, diverse measurement techniques and complex species-specific stress responses (Iversen et al., 2017).
Fine roots, commonly defined as <2 mm in diameter (Böhm, 1979), are the most physiologically active plant components of a root system. Shifts in fine-root diameter can serve as a proxy for root water/nutrient uptake capacity (Zobel et al., 2006; Tobner et al., 2013). However, studies often fail to determine diameter thresholds that indicate changes in root function but, instead, tend to rely on arbitrary thresholds. The thinnest fine-root fraction (e.g., < 1 mm) can better reflect belowground adjustments to resource availability (Leuschner et al., 2001; Zobel et al., 2007). Such thin fine-root laterals, typical of many deciduous tree species, sometimes referred to as fibrous or feeder roots (Sutton and Tinus, 1983), do not undergo secondary growth, are short-lived, and display high N concentration and respiration rate (RR). These laterals are ephemeral, turning over at rates similar to deciduous leaves (Eissenstat et al., 2013). Thin highly ephemeral roots typify a “fast-strategy” and represent one extreme of the whole-plant economic spectrum (Reich, 2014, but see also Withington et al., 2006). While, coarse, slow-growing fine-roots exemplifies a “slow-strategy” (Wang et al., 2016; Leuschner and Meier, 2018). Perennial plants with fine-roots of similar diameter may differ in form and function, which makes diameter-based root trait and biomass data difficult to interpret (Iversen et al., 2017). Therefore, an alternative classification based on defined functional groups provides an improved alternative when comparing across species and sites (Freschet and Roumet, 2017).
The “fast-slow” plant economics spectrum defined by Reich (2014) depicts a range of adaptive organ-specific strategies as basic ecological features of plant life forms. According to Reich (2014), plant organs, i.e. roots and shoots of an individual species, should conform to a resource use strategy with implications for whole-plant performance and community assembly (but see Tobner et al., 2013). Species with rapid resource turnover, so-called “fast” species (Reich, 2014), produce short-lived organs for rapid resource capture and translocation. “Slow” species, however, have long-lived organs but prolonged retention of resources. In the context of roots, “slow” species should possess long-lived fine-roots, low specific fine-root length (SRL, among other related traits) and strongly rely on ectomycorrhizal associations (ECM) (Agerer, 2001; Brundrett, 2002; Bergmann et al., 2020).
We aimed to determine if fine-root strategies of adult beech and spruce trees follow similar (“fast” vs. “slow”) patterns as those found in the leaf economics spectrum. This study utilized the prolonged, extraordinarily hot and dry, summer conditions that prevailed over wide regions of Western and Central Europe in 2003 (Rebetez et al., 2006) including the study site, Kranzberger Forst (Freising, Germany) (Raspe et al., 2004). Fine-root production and fine-root recovery rate were unaffected in beech during drought in 2003 (Nikolova et al., 2009). In contrast, fine root production and biomass recovery rate decreased by almost a factor of six in spruce during the drought year. We therefore hypothesized that beech and spruce represent belowground “fast” and “slow” plant strategies with corresponding fine-root traits. To this end, we examined the fine-root carbon (C) and nitrogen (N) status, morphological parameters, [fine root diameter (D), specific fine-root area (SRA)], and physiological parameters, [fine-root RR and concentration of non-structural carbohydrates (NSC)] in response to seasonal drought. Fine-root samples were classified into three categories, based on fine-root function and mycorrhizal abundance. This classification enabled (1) the quantification of seasonal progressive drought responses within functionally defined fine-root classes, and (2) an in situ comparison of fine-root traits between beech and spruce to distinguish underlying mechanisms of belowground drought adaptation.
Material and Methods
Site Conditions and Climate
The study was conducted at a mixed European beech-Norway spruce (Fagus sylvatica L./Picea abies [L.] H. Karst) stand at Kranzberger Forst near Freising, Germany in 2003 (Matyssek et al., 2010). The site is composed of two groups of approximately 70-year-old beech trees, each surrounded by spruce trees, which were taller although younger by about 20 years (Pretzsch et al., 2010; Häberle et al., 2012). Rooting depth was about 1 m in a Luvisol (FAO classification) which had developed from Loess over Tertiary sediments and limited to approximately 1 mby a compacted hardpan layer. Litter layer depth was about 5 cm under the spruce canopy and 3 cm under the beech. The C:N ratio within the upper 10 cm of soil ranged between 14 and 17, with the highest values occurring under beech (Schuhbäck, 2004). Soil nutrients and water were non-limiting during average growth years.
The study site is classified as temperate based on a 30-year record (1971–2000) of mean daily air temperature (T) and annual precipitation (P) (7.8°C and 786 mm, respectively), with periods of snow cover between December and February (Nikolova et al., 2009). During the 2003 growing season, extreme weather conditions were recorded at Kranzberger Forst (Nikolova et al., 2009): mean T was 3.2 °C higher, and P was 30% lower relative to the long-term seasonal averages. In the same year, a drought period occurred from August through September (Figure 1), imposing tree water limitations during the late summer. Soil moisture differed between the two tree species starting in the spring of 2003 (Nikolova et al., 2009), a direct result of the ability of spruce to take up and transpire water before beech flushed its leaves (Beier, 1998). Available soil water was completely depleted in 2003 under spruce by mid-July, under beech by mid-August, respectively. This resulted in a longer period of exhausted soil water availability for spruce (i.e., 75 d in spruce vs. 45 d in beech). Additionally, soil temperature (T(0)) was monitored at 0 cm soil depth, i.e. at the border of the humus layer and mineral soil.
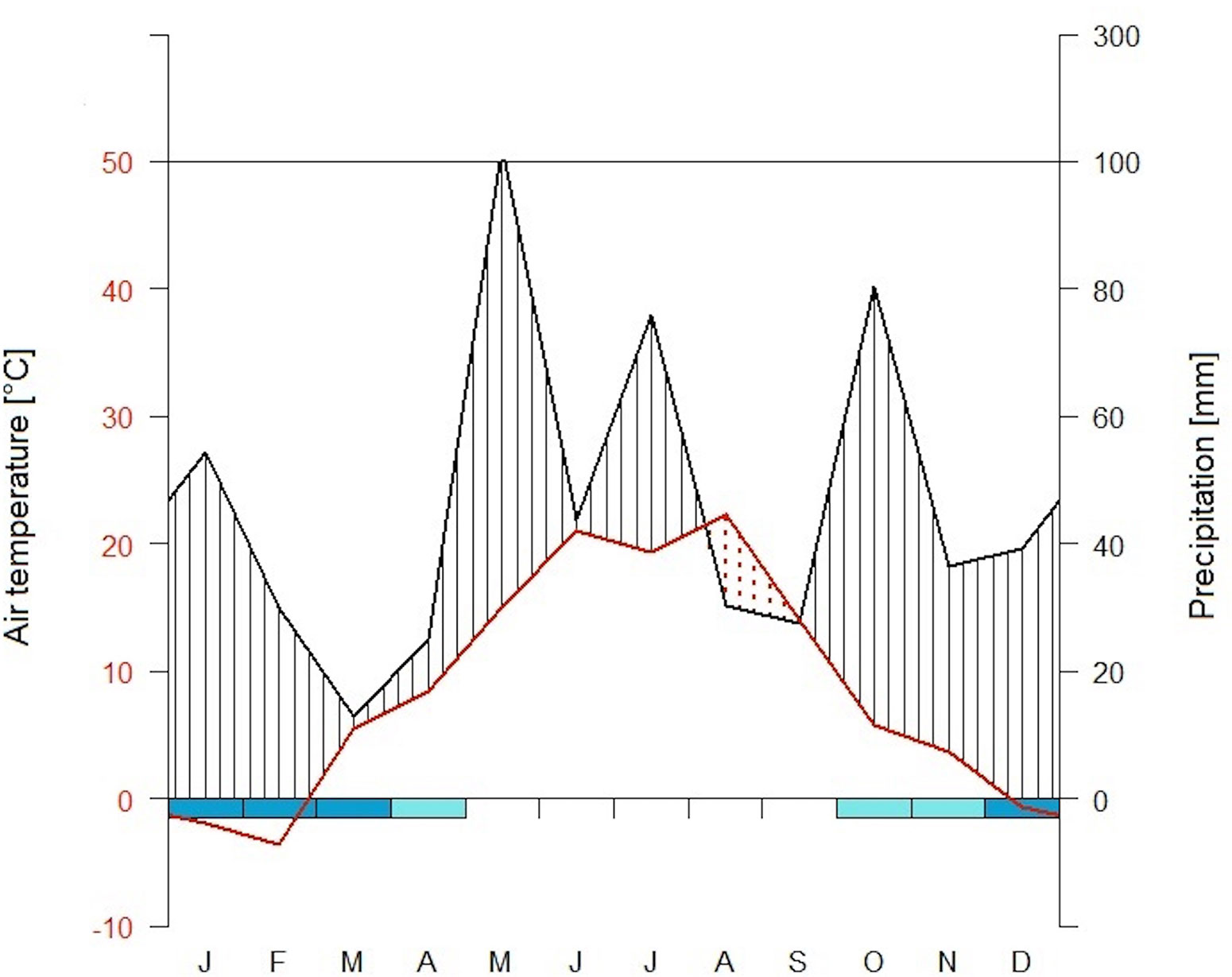
Figure 1 Climate diagram at Kranzberger Forst during the study year of 2003. Scales of air temperature T (red line) and precipitation P (black line) according to Walter and Lieth (1960), i.e. 10°C correspond to 20 mm, respectively. Arid period (when P < 2×T) is filled in dotted red vertical lines, wet periods are filled in black lines. Cold months (when absolute daily minimums ≤ 0°C) are shown in dark blue, probable frost months (when absolute monthly minimums ≤ 0°C) are shown in light blue [using the R package climatol (Guijarro, 2020)].
Fine-Root Categories and Sampling
Measurements of fine-root parameters were organized in four sampling campaigns: spring (April, May), early summer (June, July) with exhausted soil water only under spruce, late summer (August) with exhausted soil water under beech and spruce, and autumn (October, November) when available soil water partially recovered under both tree species (Nikolova et al., 2009).
To characterize fine-root trait differences between beech and spruce, three functional fine-root categories were distinguished among sampled fine-root branches that reflect the commonly used < 2 mm in diameter classification, further called “rootlets” based on (1) root function (absorptive or transport fine roots), and (2) mycorrhizal status of the absorptive fine roots (mycorrhizal or non-mycorrhizal) (Figure S1):
● Absorptive foraging fine roots (FR): fast-growing, non-mycorrhizal fine roots with primary xylem, primarily serving for soil exploration (Guo et al., 2008; Zadworny and Eissenstat, 2011);
● Transport fine roots (TR): non-mycorrhizal fine roots with secondary xylem which fulfill the role of water transport and starch and nutrient storage (McCormack et al., 2015);
● Absorptive mycorrhizal fine roots (MR): intensely branched fibrous fine roots, enlarging the plant absorptive surface by related symbionts (Agerer, 2001; McCormack et al., 2015).
In the experimental forest site, 7–10 sampling positions were randomly selected and marked within both beech and spruce groups. At each sampling position, one rootlet was entirely extracted from the topsoil, i.e. from the humus layer and the upper 10 cm of the mineral soil. Rootlets were subdivided into the three fine-root categories and dried to a constant weight, DW (g) (i.e., DWFR, DWTR, and DWMR) during the four sampling campaigns in 2003.
Dry masses were used to calculate the proportion of functional fine-root categories to individual measured parameters (e.g. RR, C, N, NSC) for an individual rootlet for each sampling campaign and tree species.
where Xi is the parameter X calculated for the rootlet i; XFR, XTR and XMR are parameter levels each as derived from sub-samples of the fine-root categories FR, TR, and MR; DWiFR, DWiTR, and DWiMR are dry masses (g) each of fine-root categories within a rootlet i, with DWi as total rootlet dry mass.
In some cases the transition from absorptive to transport fine-roots occurs gradually and may vary across species (McCormack et al., 2015). In a small pre-experiment, we determined fine root anatomy for our species to identify functional breakpoints. To this end, serial transverse sections (50 μm, from distal to proximal) were obtained in May and August on 7–10 fine-root segments of each tree species using a cryomicrotome (Frigocut, Reichert-Jung, Heidelberg, Germany). After staining with safranin and astrablue, the fine-root cross sections were mounted on glass slides and examined with a Trinocular Phase Contrast microscope (Leitz ARISTOPLAN, Leitz Meßtechnik GmbH, Germany) equipped with a digital color camera system (KAPPA model CF 20/4 DX; Kappa GmbH, Gleichen, Germany) and Kappa ImageBase 2.2 software. Anatomical study was, however, not in focus of the present investigation.
Measurement of Fine-Root RR
The fine-root RR (nmol CO2 g−1 s−1) was measured by differential infrared gas analysis, IRGA (CIRAS-2, PP-Systems, UK) in combination with an open-chamber system (PLC Conifer, PP-Systems, UK). The window size of the conifer cuvette (70 x 50 mm) allowed RR measurement of fine-root sub-samples with a fresh weight ≤ 0.6 g. Corresponding sub-samples were taken from each fine-root category, cleaned by brushing off soil particles and dead root ramifications, and transferred into mesh bags each (60 x 40 mm, mesh size of 50 µm) to protect the analyzer from contamination. Empty bags were also run in preceding tests to ensure the absence of air contamination. Each mesh bag with its root sub-sample was immediately placed in the IRGA cuvette, operated with air humidity fixed to 90%, flow rate of 0.2 l min−1, and incoming CO2 concentration of 400 µl l−1. No CO2 contamination was detected in empty cuvettes with concentrations between 400 and 1,000 µl l−1 (also see Burton and Pregitzer, 2002). Cuvette temperature (Tc) was set according to the measurement protocol (see below, ca. 7, 15, and 22°C). Root respiration was recorded upon stabilization, within 3–5 min after closing the cuvette. Since fine roots were not rinsed before measurements, microbial respiration included in the measurement RR, but was likely negligible (i.e. < 5% of RR, according to Burton and Pregitzer, 2003). Rates of microbial respiration per mass unit of soil debris are orders of magnitude lower than those of respiring, mass-related root tissue (Zak et al., 1999).
The relationship between fine-root development (see methodological details in Nikolova et al., 2006) and weather conditions (data not shown) permitted data pooling in four sampling campaigns in 2003, i.e., of April 15, April 24 and May 9 (spring), June 27 and July 4 (early summer), August 22 and 24 (late summer), and October 28, 30, and November 3 (autumn). Each time, RR was assessed for each fine-root category and tree species. Three Tc levels (7, 15, and 22°C) were applied to determine the temperature response of RR. To this end, extracted rootlets were covered with wet paper, transported to the lab in plastic bags and stored at approximately 12°C until measurement, within 3 h after sampling. Preliminary tests found RR remained stable within that time period. On each sampling date, three to six RR replicates were measured per root category, tree species and Tc level. A fresh root sub-sample was inserted at each temperature change. The RR response to Tc was examined for each fine-root category and sampling date by exponential regressions using van’t Hoff equation:
where ρ and θ are model coefficients, and RR is the RR of the respective fine-root category in beech or spruce. By means of Eq. 2, RR of each fine-root category was normalized to Tc = 10°C (i.e., RR10). RR10 was then calculated for each excavated rootlet and sampling date (Eq. 1). In addition, respiratory Q10 were determined for each fine-root category:
with Q10 as the RR response to 10°C temperature change, and θ as model coefficient. Short-term Q10 per each sampling date was distinguished from seasonal long-term Q10 (Burton and Pregitzer, 2003). A Q10 of about 1.0 reflects low temperature dependence of RR and thus low metabolic activity of roots (e.g., dormancy), whereas high Q10 indicates highly active metabolism. RR upon determination for each fine-root category (nmol CO2 g−1 s−1) was scaled to the entire rootlet (Eq. 1). In September 2004, a RR reference measurement was taken at a temperature of 10°C in the three beech and spruce fine-root categories.
Fine-Root Morphology
Four to six root samples per fine-root category of both beech and spruce were optically scanned after RR analysis (Scanner STD4800, Regent Instruments Inc., Canada). The scanner had an optical resolution of 300 dpi and pixel size of 0.085 mm which allowed measurements of root diameters > 0.18 mm (Biernacki and Lovett-Doust, 2002). Stored digital images where processed in batch mode using WinRHIZOTM Pro analysis software (Regent Instruments Inc., Canada) to assess the SRA (cm2 g−1) and D (mm) of each fine-root category from each of the four sampling campaigns in 2003.
Carbon and N Analysis
Scanned samples were then analyzed for C and N on 4–6 root sub-samples per fine-root category. Samples were dried at 65°C, milled and analyzed by combustion in an elemental analyzer (Leco, CHN1000, USA). Individual beech and spruce rootlet C and N concentrations were calculated according to Eq. 1. In September 2004, the fine-root samples used as RR reference were additionally analyzed for C and N content.
Non-Structural Carbohydrate Analysis
In each sampling campaign, four to six rootlets per tree species were harvested and prepared for sugar and starch concentration analyses. Cut rootlets were covered with wet paper and transported in a plastic bag to the lab to avoid root damage and desiccation. Sub-samples from the three fine-root categories were carefully cleaned of soil particles and dead root ramifications by light brushing, frozen in liquid N, and stored at −80°C until analysis. The root samples were then ground manually in liquid N. Lyophilized, sugars (glucose, fructose, sucrose), and starch (hydrolyzed to glucose) were extracted according to Fleischmann et al. (2009) before measuring concentrations by HPLC. In spruce samples, pinitol was additionally identified (using expertise by M. Popp, University of Vienna, Austria). Total sugar concentration (TSC, mg g−1) was additionally calculated as the sum of all analyzed sugars, inclusive pinitol in case of spruce. Individual beech and spruce rootlet non-structural carbohydrates concentrations were calculated according to Eq. 1.
Data Analysis
Seasonal effects (i.e., between the sampling campaigns) on fine-root parameters (i.e. SRA, D, non-structural carbohydrates, N and C concentrations, and C:N) were tested for each tree species by a factorial two-way ANOVA including fixed factors “category” and “sampling campaign”. The interaction term “category” × “sampling campaign” was also included to test whether the responses of different fine-root categories depend on the sampling timing. Model residuals were tested for normality (Shapiro-Wilk test) and variance homogeneity (Levene test). For multiple comparisons, subsequent Tukey-HSD post hoc tests (HSD.test function of the agricolae package in R; Mendiburu, 2020) were performed. The temperature effect on RR was assessed in each fine-root category by an exponential fit (Eq. 2), based on datasets measured at three temperature levels over four sampling campaigns in 2003. Effect of N on RR was analyzed with individual beech and spruce rootlets by standardized major axis regression models (smatr R package; Warton et al., 2012), using RR10 for standardization. Non-parametric U-test (Mann-Whitney U test) was used to test for significant differences between two sample units. Statistical evaluation was performed with SPSS (version 13.0, SPSS INC., Chicago, IL, USA) and R (version 3.4.4; R Development Core Team, 2018). In all analyses, differences at p < 0.05 were considered significant.
Results
Fine-Root Categories
Foraging roots (FR) constituted between 17–35% of the total beech rootlet biomass with a maximum of 35% in late summer and a minimum of 17% in autumn (Figure 2; Table S1). Transport fine roots (TR) contributed between 20-42% of the total rootlet biomass, with a minimum of 20% in late summer and a maximum of 42% in autumn, while mycorrhizal absorptive fine roots (MR) were consistently approximately 40% of the total rootlet biomass throughout the entire study period.
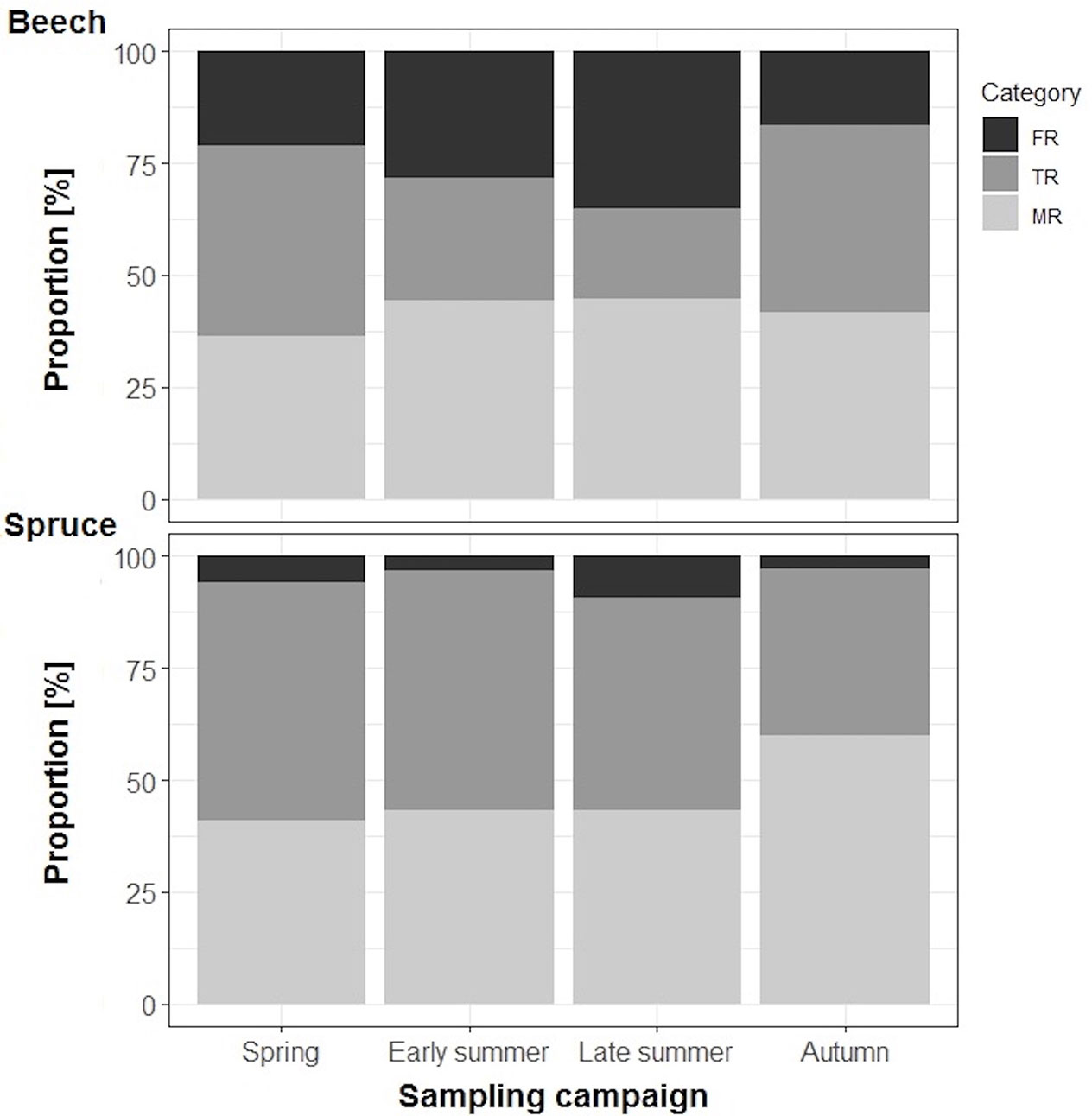
Figure 2 Average proportions (%) of three fine-root categories in the biomass of individual beech (n=7–8) and spruce (n=8–10) rootlets: FR = absorptive foraging roots with primary xylem, TR = transport roots with secondary xylem, MR = absorptive mycorrhizal fine roots.
For spruce, FR contributed substantially less to the total rootlet biomass compared to beech and ranged between 10% in late summer and 3% in autumn (Figure 2; Table S2). Transport fine roots accounted for 37–53% of the total spruce fine-root biomass with a maximum of 53% in spring and a minimum of 37% in autumn. Absorptive mycorrhizal fine-roots contributed approximately 40% in spring through late summer and reached a maximum of 60% in autumn.
In spring, FR in both beech and spruce had whitish surface, but later, in early summer, spruce FR started to change color to yellow and brown (Figure S1). At the same time, beech FR remained whitish but appeared thinner and started to branch (Figure S2). In late summer of 2003, anatomical differences were apparent between beech and spruce FR: in beech, the living primary cortex was white, while in spruce, the primary cortex turned brown and shriveled (i.e., likely died), and the root surface suberized during drought (Figure S1).
Fine-Root Morphology
In beech, SRA and D differed between fine-root categories depending on the season of the sampling campaign (Table 2). Beech FR SRA increased significantly from 167 cm2 g−1 in spring to a maximum of 533 cm2 g−1 in autumn. Beech SRA of the TR and MR categories were highest in early summer (78 and 520 cm2 g−1, respectively), but reached lowest levels in late summer (58 and 388 cm2 g−1); however this change was significant only for MR (Table 1). In beech, MR had generally the thinnest diameters (0.61–0.67 mm) and TR, had, in contrast, the largest diameters (1.40–1.62 mm). Interestingly, FR had diameters that were similar to TR at the beginning of the growing season (1.72 mm), but became thinner in early summer when their diameters were similar to those of MR (0.56 mm; Table 1).
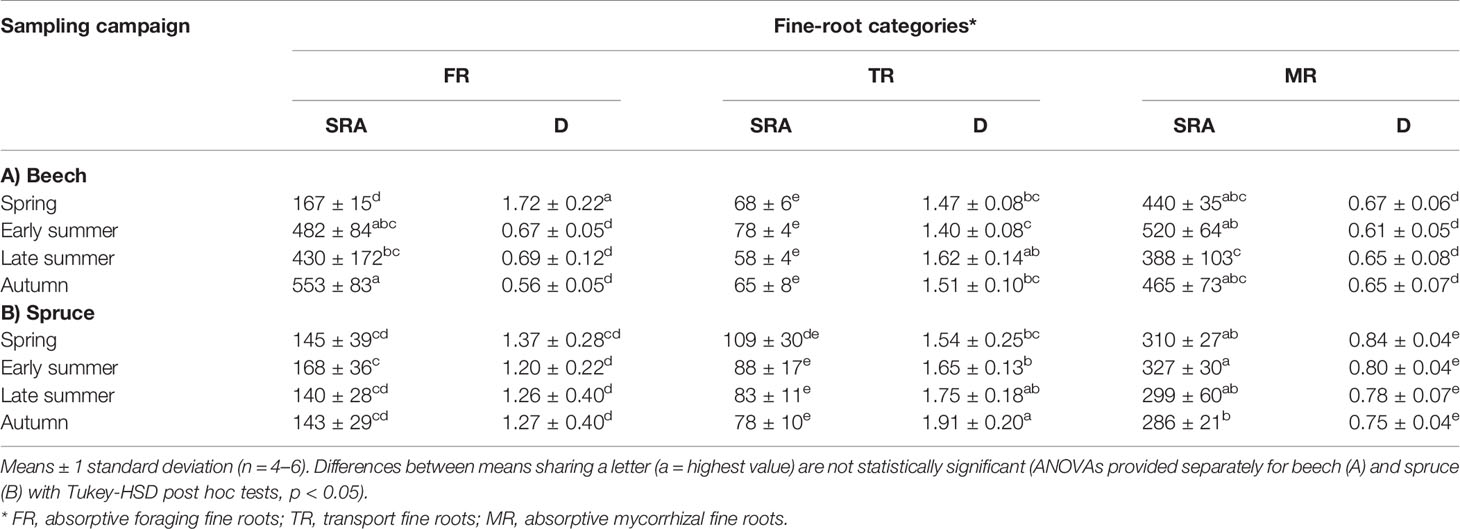
Table 1 Specific fine-root area (SRA, cm2 g−1) and diameter (D, mm) in (A) beech and (B) spruce fine-root categories, assessed at four sampling campaigns during 2003.
In spruce, SRA differed between fine-root categories and sampling campaigns with both factors independent from each other (Table 2). Spruce FR, in contrast to beech, had low seasonal variation of SRA (Table 1), reaching maximal levels in early summer (168 cm2 g−1), and minimal in late summer (140 cm2 g−1). TR had highest SRA in spring (109 cm2 g−1) and lowest in autumn (78 cm2 g−1). However, the changes in SRA of FR and TR were not significant relative to the other samplings. Spruce MR had the highest SRA among spruce roots with a maximum in SRA in early summer (327 cm2 g−1), and a minimum in autumn (286 cm2 g−1). Spruce D varied significantly between fine-root categories depending on the season of the sampling campaign (Table 2). Remarkably, no general seasonal adjustment of D was observed in spruce. In spruce, the diameter of FR roots was generally larger compared to beech FR, but in contrast to beech did not change with season (1.20–1.37 mm; Table 1). In spruce, TR had the largest diameter gradually increasing from 1.54 mm in spring to maximum of 1.91 mm in autumn. The MR category had the thinnest diameter (0.75–0.84 mm); however, the MR were coarser compared to the same category in beech.
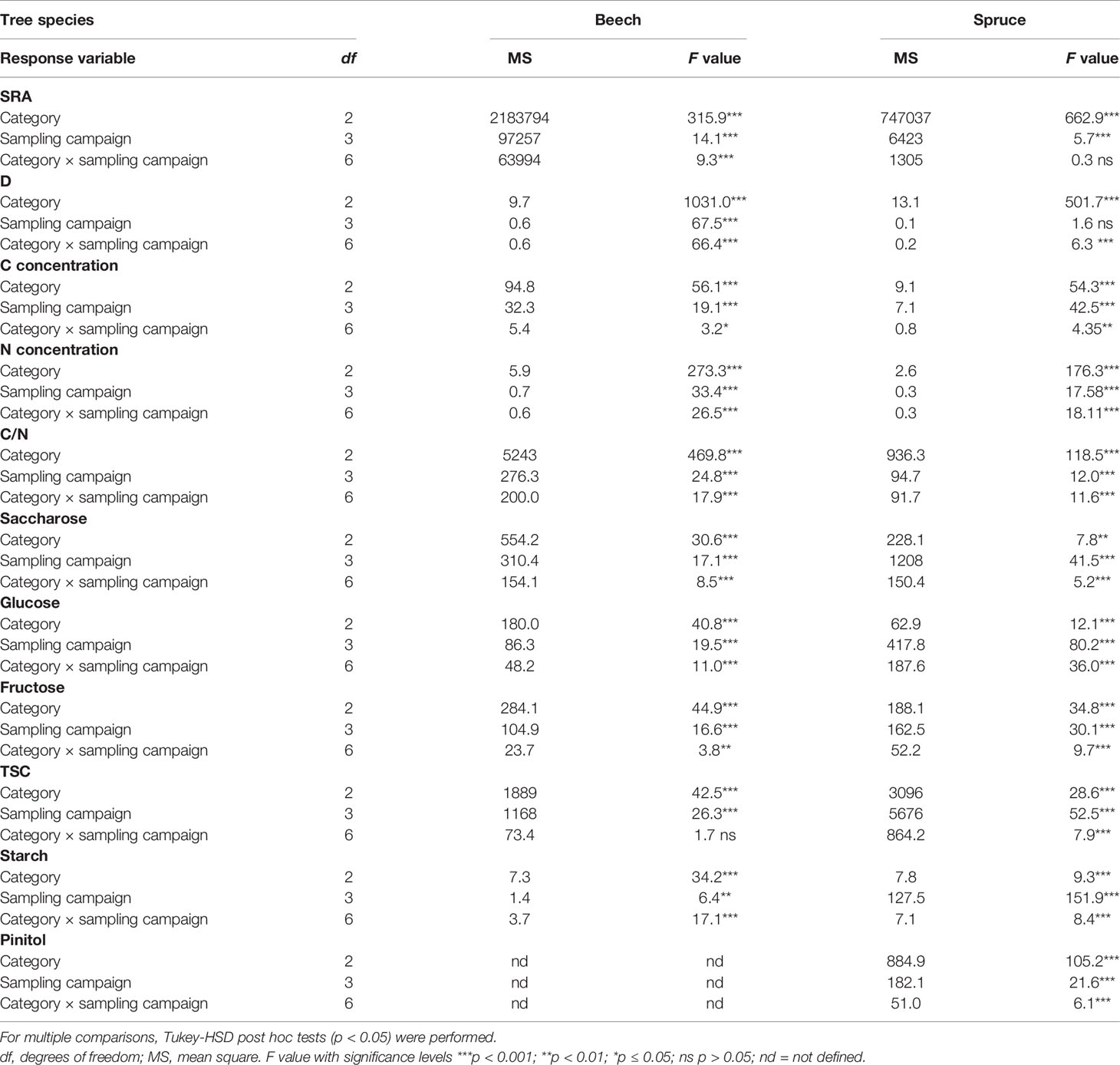
Table 2 ANOVAs outcome for key fine-root parameters, assessed at four sampling campaigns during 2003 in fine-root categories of beech and spruce. Main factors are “category” and “sampling campaign” as well as their interaction term. Total number of samples for beech was 135, and for spruce 167.
Fine-Root Respiration
In beech, FR had the highest RR10 and temperature sensitivity (Figure 3; Table 3): RR10 ranged between 17.06 nmol CO2 g−1 s−1 in spring and 9.46 in autumn, and Q10 was highest in early summer (i.e., 2.20) but lowest in spring and late summer (i.e., around 1.4). In contrast, TR had the lowest RR10 (4.14–2.40 nmol CO2 g−1 s−1) and temperature sensitivity: Q10 even dropped to 1.09 in the extremely dry month of August (i.e., late summer). On average, MR had RR10 levels of 4.5 CO2 g−1 s−1 (late summer) to 8.9 nmol CO2 g−1 s−1 (spring), with the lowest Q10 of 1.16 in late summer.
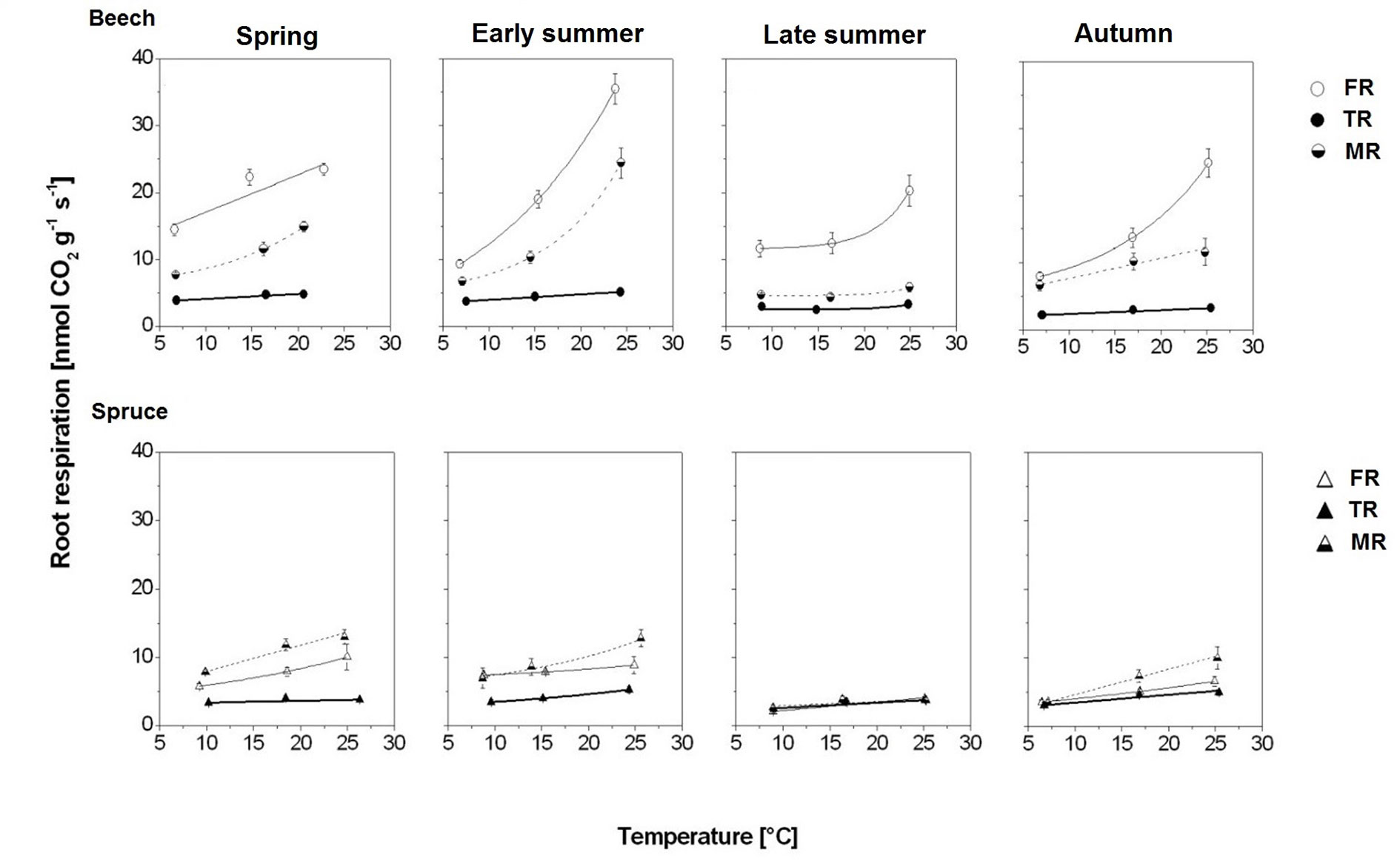
Figure 3 Seasonal response of root respiration rate (RR) to temperature in beech and spruce per fine-root category, given as exponential response functions of absorptive foraging fine roots FR (solid thin line), transport fine roots TR (solid bold line), and absorptive mycorrhizal fine roots MR (dashed line); means ± 1 standard error (n = 4–6). Model statistics are provided in Table 3.
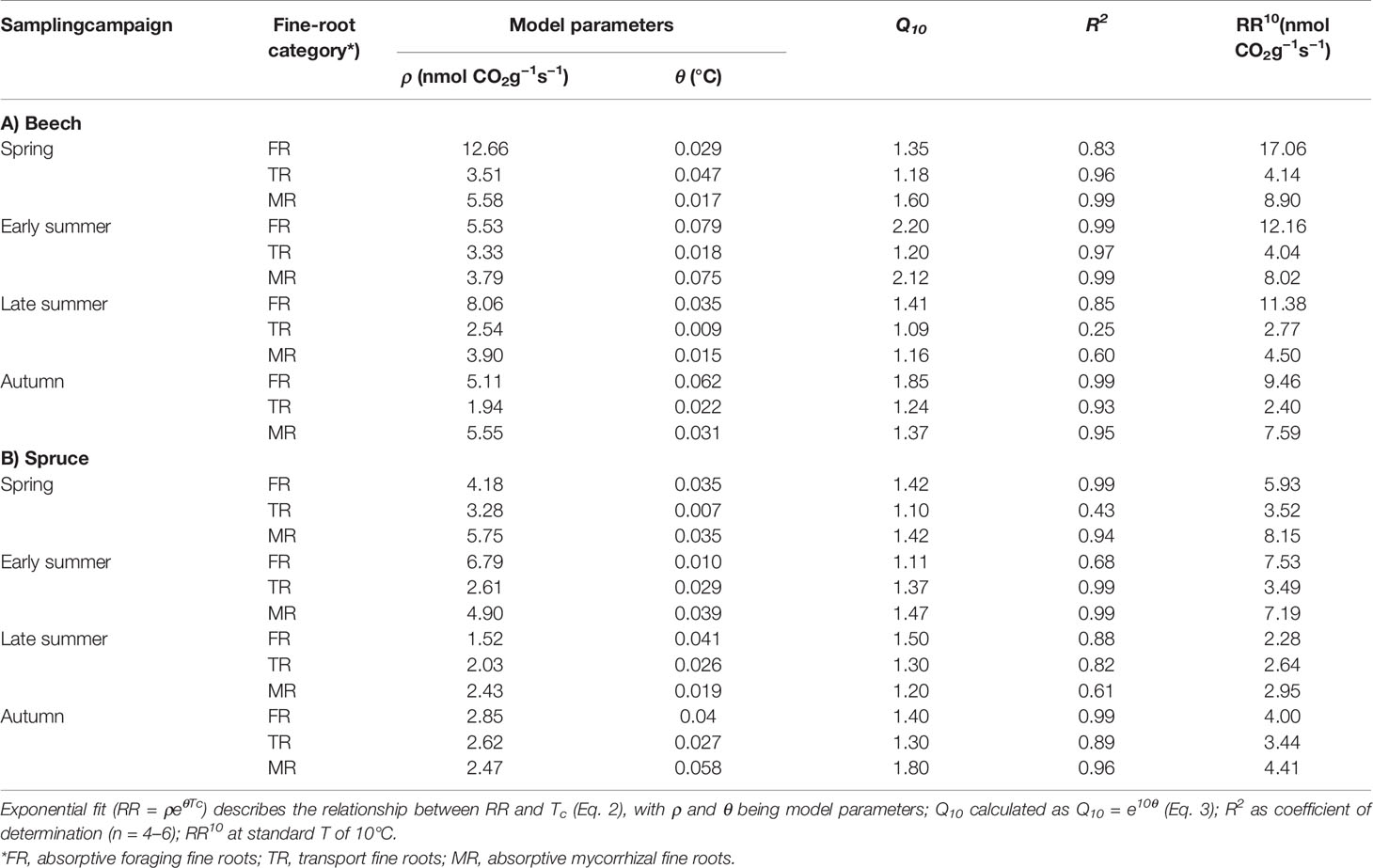
Table 3 Root respiration rate (RR) in response to temperature per fine-root category of beech (A) and spruce (B) at four sampling campaigns during 2003.
Compared to beech, all fine-root categories of spruce had lower respiration levels and less variation in RR10 throughout the entire study period. Spruce FR had 2–3 times lower RR10 but comparable Q10 except early summer when Q10 in spruce FR was extremely low (i.e., 1.11; Figure 3; Table 3). Spruce TR had the lowest RR10 and Q10, ranging between 2.6 and 3.5 nmol CO2 g−1 s−1, and 1.09 and 1.37, respectively. The highest RR10 and temperature sensitivity were found in spruce MR (Figure 3; Table 3): RR10 maximum in spring (8.15 nmol CO2 g−1 s−1), and Q10 maximum of 1.80 after precipitation returned in autumn (Figure 1).
Rootlet RR at mean daily soil temperature (RRT(0)) and at a standard temperature of 10°C (RR10) were higher in beech than spruce over the entire study period (Figure S3). In late summer, rootlet RRT(0) decreased significantly in both tree species despite higher soil temperature (Ts). Beech rootlets had the highest RR10 in spring (9.90 nmol CO2 g−1 s−1). For the rest of the season, rootlet RR were similar in both species with the seasonal RR10 minimum occurring in late summer. In autumn 2004, rootlet RR10 was, in both tree species, 30–50% higher compared to autumn 2003, with higher increases in spruce.
Carbon and N Status
Carbon and N concentrations varied among transport versus absorptive fine-root categories and sampling periods (Table 2). In beech, FR and MR had the lowest C concentration in spring (400 mg g−1 to 420 mg g−1) that increased over the course of the season up to nearly 430–450 mg g−1 (Table 4). Transport root C concentration was generally higher than in the other two categories, and similarly increased across the entire sampling period (i.e., from 467 mg g−1 to 476 mg g−1). During late summer, FR reached maximal N concentrations among all categories (27.2 mg g−1); however, these patterns shifted by autumn where FR and MR had similar N concentrations (nearly 19 mg g−1). This similar N level was the result in a drop in N in the FR roots at the end of vegetation period. Across the entire study period, TR had the lowest N concentration among all categories, ranging from 6.8 mg g−1 in early summer to 10.1 mg g−1 in autumn. The C:N ratio was lowest in FR (a minimum of 16.6 mg g−1 in late summer and a maximum of 29.5 mg g−1 in spring), and highest in TR (a minimum of 47.2 mg g−1 in autumn a maximum of 69.5 mg g−1 in early summer), with lowest seasonal variation in MR category (23–32 mg g−1). When calculated per rootlet, C:N of beech rootlets were the lowest in late summer, a result of the high proportion of high N containing FR roots that were present at this time (Table 4, Figure S4).
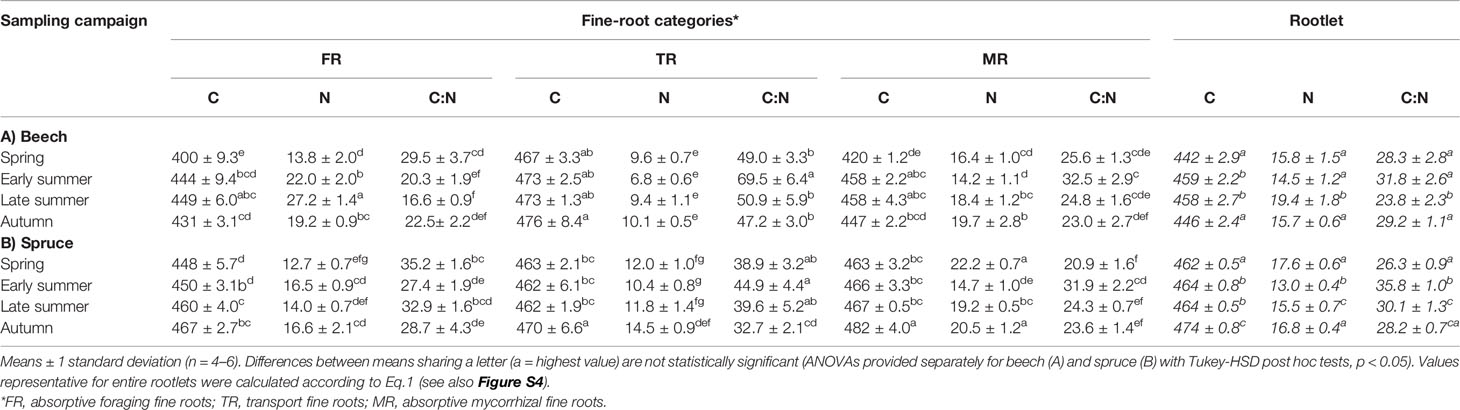
Table 4 Concentration of carbon (C, mg g−1) and nitrogen (N, mg g−1), and the C:N ratio in the fine-root categories of (A) beech and (B) spruce at four sampling campaigns during 2003.
In spruce, C was more stable across all fine-root categories and months (Table 4) with lowest levels in FR during spring (448 mg g−1) and highest in MR during autumn (482 mg g−1). In contrast to beech, TR was not the C-richest category in spruce, but was similar to MR (C differences between both categories were not significant). Absorptive mycorrhizal roots had generally the highest N levels, with the exception of early summer when N decreased substantially (14.7 mg g−1) to the N-levels of FR (i.e., nearly 16 mg g−1). Similar to beech, TR had the lowest N concentration among the fine-root categories, especially in early and late summer (10–12 mg g−1). In contrast to beech, spruce rootlets had the highest C:N in early summer, which was due to the reduced N at the beginning of the drought period (Table 3; Figure S4). In autumn 2004, N content in spruce rootlets was nearly 30% higher compared to autumn 2003, indicating N limitation in spruce at the end of the growing season in 2003 (Figure S4).
Relationship Between RR and N Concentration
In both tree species, RR10 was positively correlated to the corresponding N concentration of the rootlets (Figure 4). In beech, RR10 peaked in response to N in spring (β = 7.27, p < 0.001), compared to early spring in spruce (β = 12.18, p < 0.001; Table 5). Beech RR10 responded positively to N across all sampling dates (coefficient β between 4.97 and 7.27), while spruce responded weakly (late summer: β = 0.61, p = 0.001), or not at all (autumn: β = 3.65, p = 0.478; Table 5).
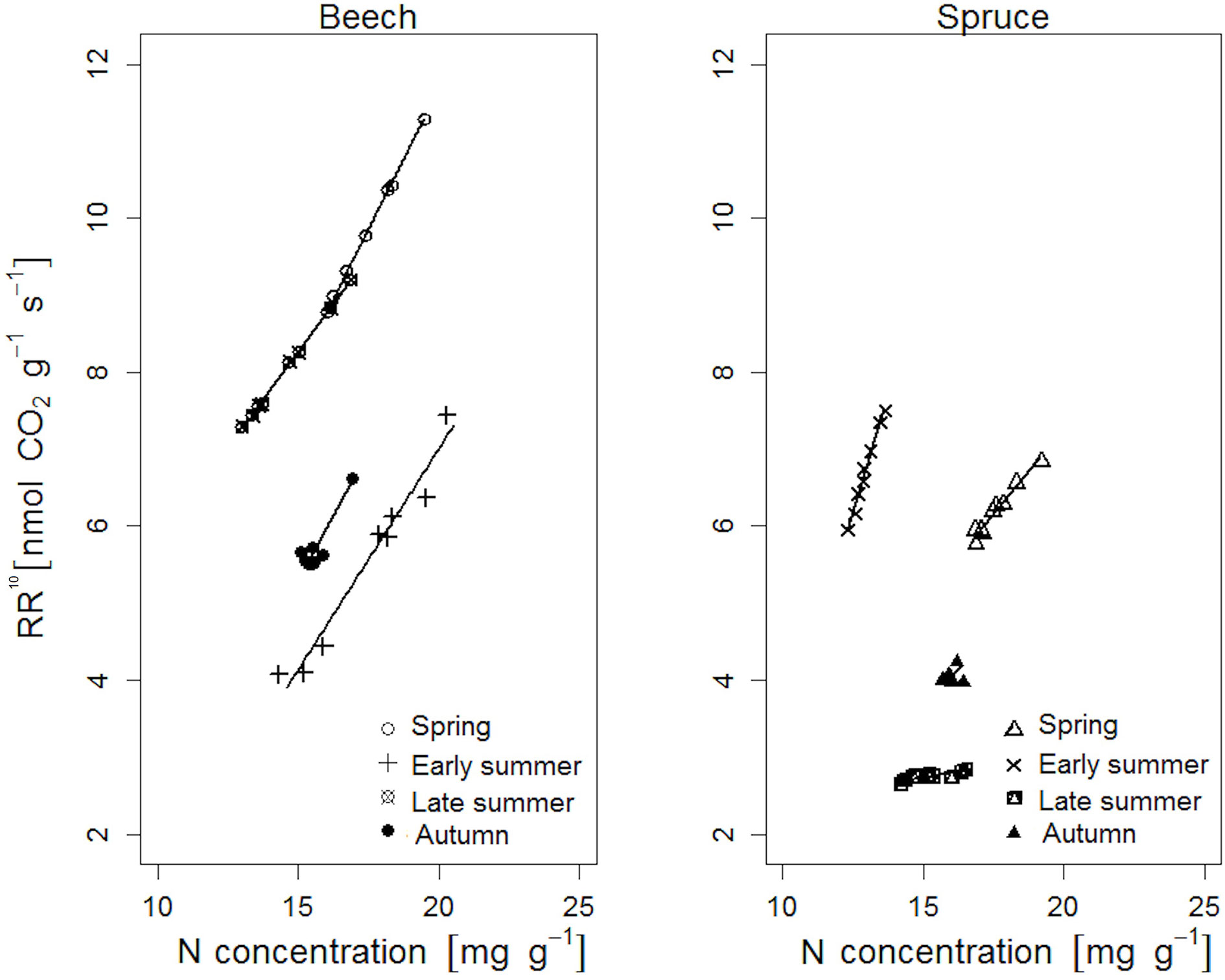
Figure 4 Relationship between root respiration rate at a standard temperature of 10°C (RR10) and nitrogen concentration of individual beech and spruce rootlets. Lines represent a linear fit (regression statistics are provided in Table 5).
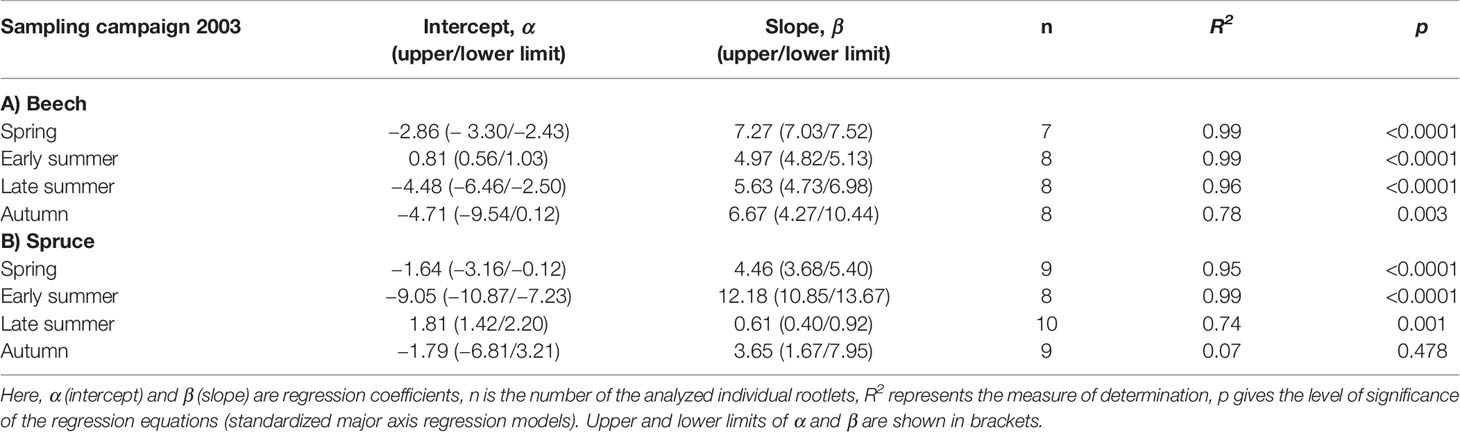
Table 5 Linear fit (RR10 = α + βN) between root respiration rate (RR) at a standard temperature of 10°C (RR10; nmol CO2 g−1 s−1) and nitrogen concentration (N, mg g−1) in individual rootlets of (A) beech and (B) spruce at four sampling campaigns during 2003.
Non-Structural Carbohydrates
All studied non-structural carbohydrates varied among fine-root categories and sampling periods (Table 2). Interestingly, the seasonal responses of NSC in beech were similar among the studied fine-root categories, i.e. were independent on the sampling timing. During spring and early summer 2003, beech FR had the lowest TSC concentration 38 mg g−1 (Table 6; Figure S5), which increased by 40–60% during late summer the second half of the growing season as a result of an increase in sucrose and fructose (in late summer), and glucose concentrations (in autumn). Transport fine-roots of beech had lowest TSC also in spring (35.2 mg g−1), but peaked in the second half of the growing season also due to enhanced fructose and glucose concentration. Absorptive mycorrhizal fine-roots of beech had low TSC during spring and early summer (approximately 20 mg g−1), but levels doubled during late summer and autumn similar to FR. The three fine-root categories of beech showed however different patterns of starch allocation during the study period (Table 6): highest concentration was detected in FR in spring (3.26 mg g−1) and steadily decreased through the rest of the growing season (Figure S5). In TR starch was extremely low in spring and then completely exhausted in early summer. Interestingly, the starch levels in TR recovered in late summer and in autumn; this seasonal change was however not significant. Absorptive mycorrhizal roots had oscillating starch levels, with minimal levels in late summer, but accumulated starch in autumn (1.86 mg g−1).
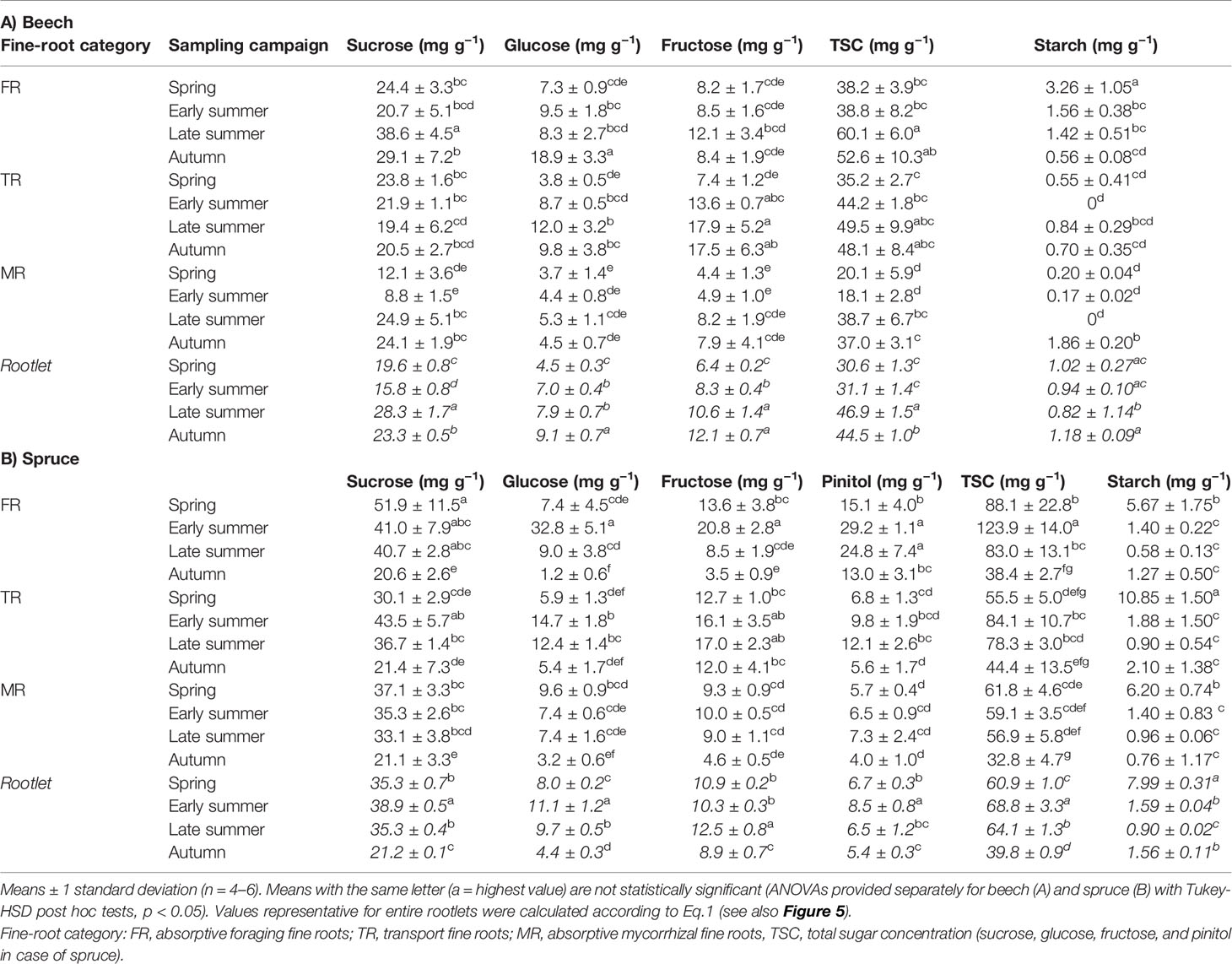
Table 6 Non-structural carbohydrate concentrations in fine-root categories of (A) beech and (B) spruce in 2003.
Spruce fine-root categories had more variable seasonal patterns in NSC fractions compared to beech (Table 2 and Table 6). Foraging non-mycorrhizal roots had highest TSC in early summer (123.9 mg g−1), and lowest in autumn (38.4 mg g−1), with glucose, fructose and pinitol influencing the seasonal variation (Figure S5). Total sugar concentration in TR of spruce also had the highest levels in early summer (84.1 mg g−1), but minimum in autumn (44.4 mg g−1), although the seasonal variation, driven mainly by sucrose, was less pronounced than in FR. In MR, TSC had highest levels in spring (61.8 mg g−1), and lowest in autumn (32.8 mg g−1) due to a simultaneous decrease in the concentration of all analyzed sugars. Starch concentrations in fine-root categories of spruce were highly variable at the beginning of the study (6–11 mg g−1 in spring), but then decreased five-fold and leveled out at similar concentrations for the remainder of the study (1–2 mg g−1; Table 6). Interestingly, TR had in spring starch levels of 10.85 mg g−1 that were approximately two-fold greater than FR or MR roots but, then decreased five-to-ten-fold to levels typical for the other two categories.
At the beginning of the growing season, beech rootlets had two times lower TSC relative to spruce (Figure 5). During late summer and autumn, however, TSC of beech rootlets increased by 50%. In spruce, TSC was only slightly enhanced in early summer, remained stable into late summer, but decreased in autumn. In both species, the seasonal dynamics of TSC was mainly driven by sucrose which contributed 55–65% to TSC. In general, the higher sucrose levels during spring and summer period as well as the permanent presence of pinitol resulted in the higher TSC in spruce relative to beech fine-roots. Rootlet starch concentration in beech did not show pronounced seasonal dynamics (Table 6, Figure 5). Despite higher levels of starch in spruce during spring, by the end of the growing season starch levels were similar in both beech and spruce (i.e., about 1.0–1.5 mg g−1) (Table 6; Figure 5).
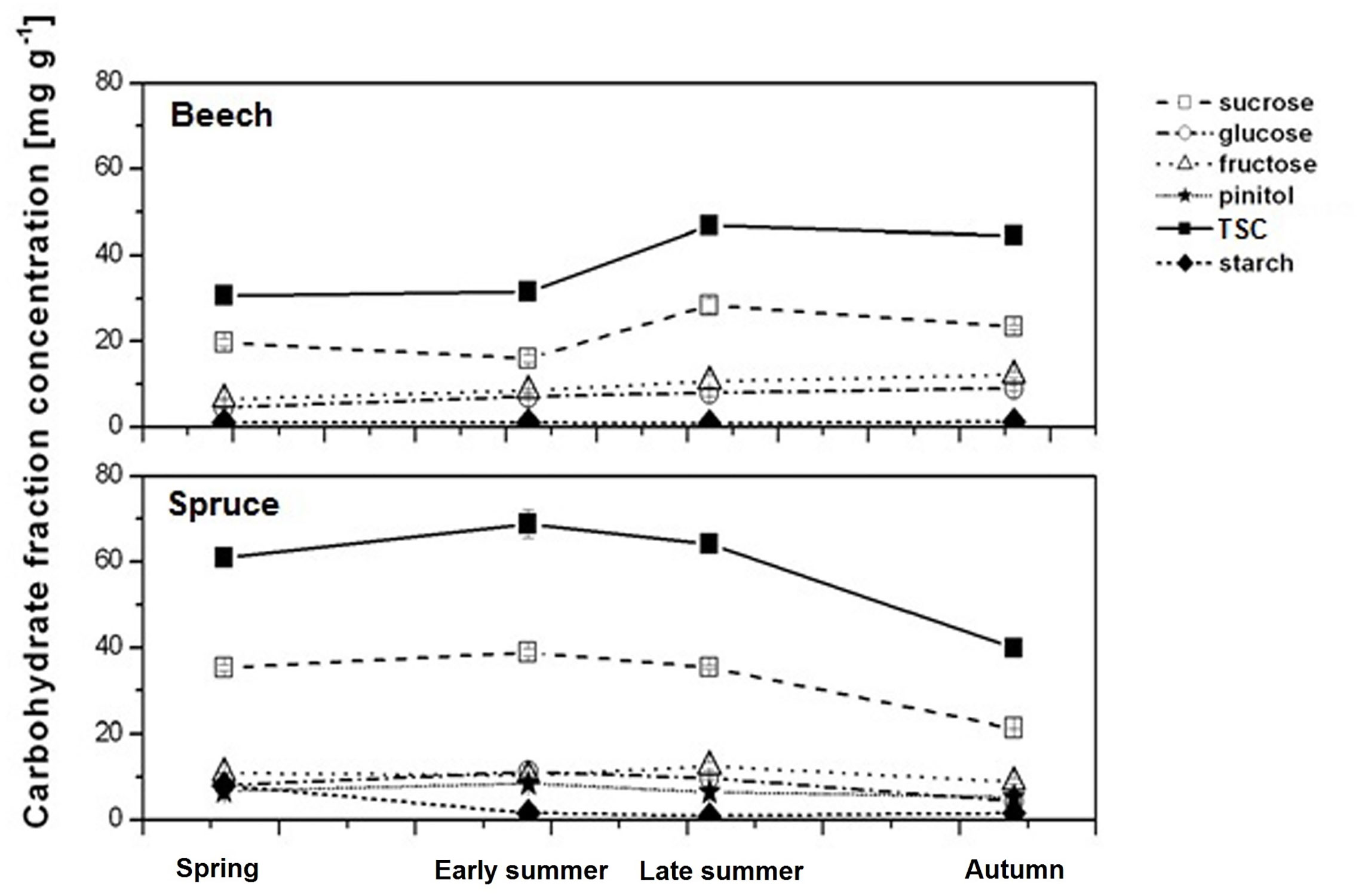
Figure 5 Concentrations of the carbohydrate fractions (sucrose, glucose, fructose, pinitol) and the total sugar concentration TSC (i.e. integral of sucrose, glucose, fructose, and, in case of spruce, pinitol) together with starch concentration in individual rootlets of beech and spruce during 2003 (means ± 95% confidence interval, n = 7–10).
Discussion
Our study presents a novel look at beech and spruce fine-root ecological strategies (cf. Weemstra et al., 2017) by directly comparing key morphological and physiological traits of three functionally defined fine-root categories. We hypothesized that the fine-roots of both species would adjust to seasonal drought differently with beech following a “fast”, and spruce a “slow” ecological strategy. However, we determined that each tree species employs diverse ecological strategies dependent on the functional fine-root categories investigated (Figure 6).
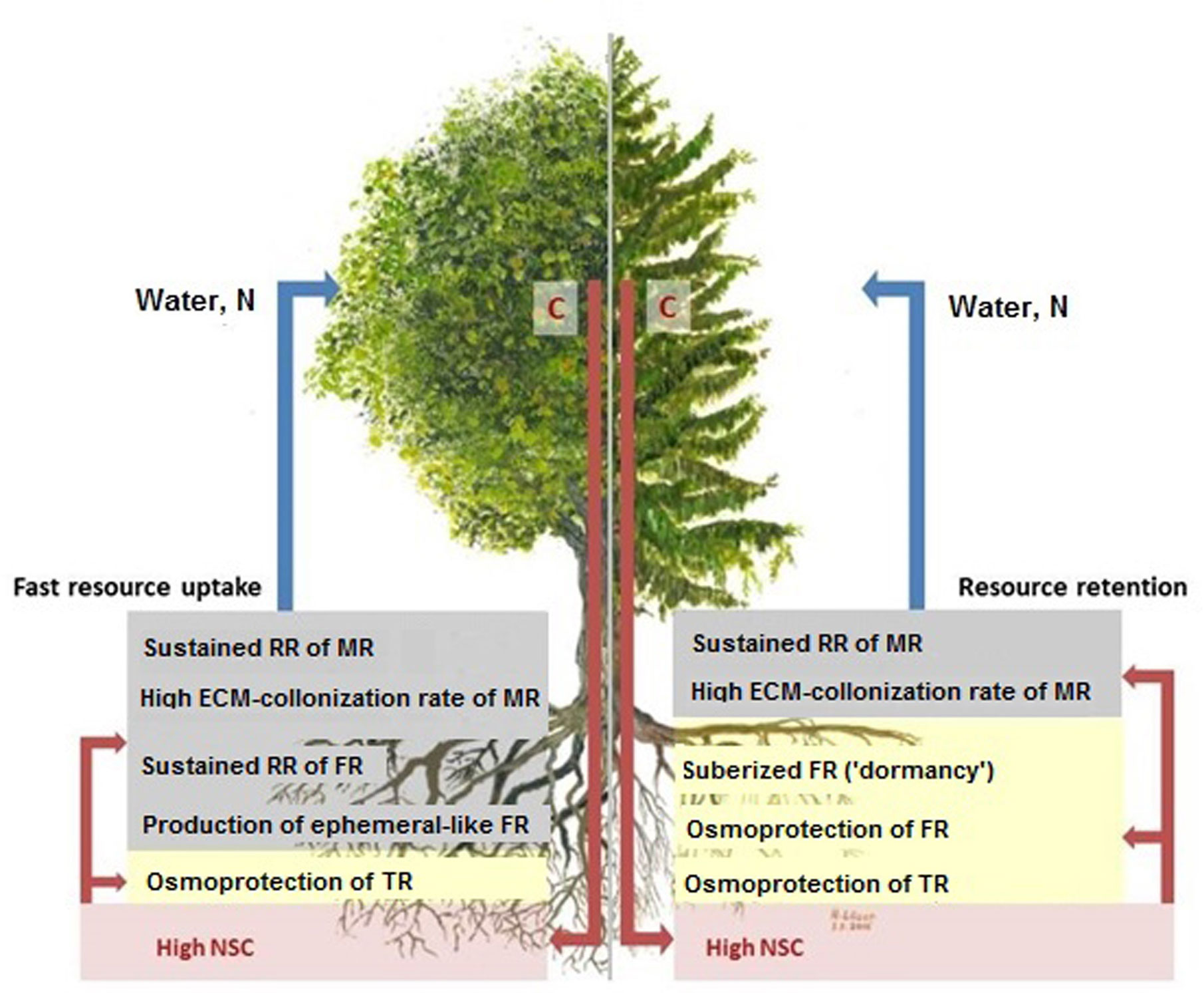
Figure 6 Scheme of the two different ecological strategies of beech (left) and spruce (right) fine-roots under seasonal drought. Red arrows show the transport of C (assimilates, NSC) from leaves into root structures and functions, blue arrows show the returns of soil resources (e.g., water, N) from roots to leaves. The fine-root traits associated with “fast” ecological strategy are indicated in grey, those associated with “slow” ecological strategy are shown in yellow. The NSCs (red lettering) are crucial in linking and controlling above and belowground processes. ECM-colonization data according to Nikolova, 2007. Abbreviations see text.
Morphological Traits
The non-mycorrhizal absorptive roots adjusted in a species-specific manner supporting fine-root ecological strategy hypothesis. In beech, SRA of FR had the highest seasonal variation among all categories that more than doubled from spring through autumn. Enhanced SRA of FR was accompanied by a distinct reduction in mean diameter to the levels typical of mycorrhizal MR. Such a diameter decline may be associated with optimized uptake of scarce resources (Zobel et al., 2007; Ma et al., 2018). During drought, thin ephemeral roots are “shed” by species which are capable of root regrowth upon soil rewetting (Meier and Leuschner, 2008). The ability to “shed” roots and to initiate new root growth when available resources are present, represents an effective adaptation to drought, as long as the trees can afford the C demand (Brunner et al., 2015). This “shed-regrowth” mechanism as detected here in FR allows beech to regain fine-root biomass after drought (Leuschner et al., 2001). While substantial fine-root growth after drought comes at a C cost, the ability to grow new absorptive fine-roots with high SRA is essential for continued water uptake, demonstrating a “fast” ecological strategy for FR (Figure 6).
In contrast, spruce FR showed no seasonal variation of SRA. Instead, spruce FR became pigmented from white to brown during the dry midsummer (Blaschke et al., 2006; Nikolova et al., 2006). Also during this time, the roots temporarily stopped growing which was not the case in previous years (i.e. 1999, 2000, and 2002) when a sufficient water supply was present (Nikolova et al., 2010). The high degree of suberization of the FR can reduce radial hydraulic conductivity and may act as a physical barrier to the movement of water/nutrients into the plant (Steudle, 2000). Development of more suberized and lignified roots in drying soils has been described in plants with limited access to water, e.g. desert plants (Nobel and Huang, 1992) as well as in Quercus ilex and Vitis vinifera (Brunner et al., 2015 and the references therein) and Pinus halepensis (Leshem, 1970) and may serve as a mechanism to decrease water loss from roots. Orlov (1957) similarly observed color progression in absorbing roots of spruce under natural conditions and related this to the senescence of the cortex. The present study is, to our knowledge, the first study to report a shift in color/suberization in spruce fine-roots in response to drought. Such adjustment of FR may reflect an earlier senescence of the spruce fine-roots in order to enhance C retention during harsh drought, thus supporting the “slow” ecological strategy (Figure 6).
Physiological Traits
Beech had higher RR than spruce across all root types. In beech, this high RR was driven by FR, despite this category containing roots of a greater diameter compared to MR (e.g., in spring). This finding conflicts with other studies that report a decrease in RR with root diameter (e.g., Pregitzer et al., 1998; Di Iorio et al., 2016). Absorptive roots, like FR, represent primary roots with active cell divisions within the apical meristem, and a determinate growth pattern (Dubrovsky, 1997; Heimsch and Seago, 2008). Thus, the stimulation of root respiration and growth during the summer dry periods could be a result of phytohormonal control (Chapman et al., 2003; Perrot-Rechenmann, 2014) likely leading to temporary dysfunction of the primary-root apical meristem but stimulation of lateral-root production. Dubrovsky (1997) suggested the purpose of such dysfunction is to allow for the formation of lateral roots which quickly elongate (i.e. respire) and then slow down and finally cease elongation only a few days after emergence. In cactus, Dubrovsky (1997) related this adjustment to temporary water availability in arid environments (see also Chapman et al., 2003). In the present study, similar mechanisms may have induced a formation of lateral-like ephemeral FR roots with high SRA in beech. These young FR maintained the high RR of the beech rootlets during the rest of the growing season, thus representing a “fast” ecological strategy (Figure 6). Consequently, the formation of ephemeral root parts may represent a competitive advantage of beech, enabling rapid proliferation when resources are available (Nikolova, 2007; Bauerle, in prep.) or may play a role in hydraulic redistribution when present (Caldwell et al., 1998; Bauerle et al., 2008; Zapater et al., 2011).
During spring, spruce had the highest RR and Q10 in FR. During late summer, when soils were extremely dry, spruce FR possessed the lowest RR but had the highest metabolic activity (as reflected by high Q10) among all three fine-root categories. The suberized exodermis protects the FR from desiccation (Cruz et al., 1992; Steudle, 2000), however at the price of C expenditure for respiration. Since the proportion of FR in the rootlet biomass was low (<10%) in spruce, this fine-root category did not drive the RR of rootlets for this species.
Overall, in both tree species, MR had medial RR, with the exception of spruce in spring and late summer, when MR had the highest respiration among all root categories. This could be explained by the relatively high percentage of non-mycorrhizal root tips (ca. 45%; Nikolova, 2007), which are fast-growing (Brundrett, 2002) and were produced in new root flushes during spring. In contrast, in late summer, the respiration activity of MR was likely needed to support the high fungal colonization rate (over 90%; Nikolova, 2007). In spruce rootlets, MR is likely driving the respiration activity during drought, thereby following a “fast” ecological strategy (Figure 6).
Not surprisingly, respiration was low in TR in both tree species, as such aged root sections (cf. Solly et al., 2018) with secondary growth serve water/nutrient transport rather than resource uptake (Lobet et al., 2014) indicating a “slow” ecological strategy. The lowest Q10 was also measured in TR (around 1.10), as RR approached the level of maintenance respiration (RR10 around 2.5 nmol CO2 g−1s−1, in both tree species), in the absence of high temperature sensitivity (Burton et al., 1998).
Carbon and N Concentrations
In beech rootlets, C concentration was the highest during early and late summer. This corresponds to the seasonal C trend reported for beech fine-roots from adult coppice forests in Italy’s Prealps (Terzaghi et al., 2013). In contrast, C concentration of spruce rootlets consistently increased through the growing season, reaching higher levels than beech rootlets by the end of the growing season. In both tree species, C dynamics of the rootlets seemed to depend on the proportion of TR, which was the root category with the highest C concentration.
Beech rootlets only had significantly higher N concentration compared to spruce in late summer, which possibly resulted from the enhanced proportion of ephemeral roots with highest N. In rootlets, our results did not support the traits described by Reich (2014) for the foliar economic spectrum, that species with “fast” strategies, i.e. beech, will have higher N concentration compared to the “slow”-species, i.e. spruce. In contrast, our findings are in line with Weemstra et al. (2017) who found that root traits do not necessarily correlate with leaf traits, in particular in species with more conservative root traits (i.e. with thick roots and long root lifespan) such as (evergreens) conifers. Concerning these tree species, the discrepancy in leaf and root traits may result from the confounding effect of mycorrhiza on the seasonality in water and soil resources uptake.
According to Gordon and Jackson (2000) the average C:N ratio of fine roots <2 mm is 43:1 across a broad range of ecosystems and biomes. In beech fine-roots, Terzaghi et al. (2013) reported a C:N ratio from 40:1 to 90:1. Both studies reported on values which are much higher than the C:N ratio from our study. This discrepancy occurred as a result of the higher N concentration we found in beech and spruce rootlets (13–19 mg g−1). This higher N concentration of rootlets in the trees from Kranzberger Forst is, however, not surprising, as it reflects the high levels of N deposition measured at that time in similar forests in Bavaria (20–25 kg ha−1 a−1; Raspe et al., 2018).
In our study, N concentration was a significant predictor for root RR of the rootlets (see also Burton et al., 1998; Ceccon et al., 2016), but the seasonal dynamics in these relationships was species-specific, in particular during the harsh drought in late summer. In beech, the N status varied largely within the rootlets, indicating, in this tree species, an adaptable fine-root system to a patchy soil environment. In contrast, spruce rootlets were more uniform and likely showed a temporal ‘dormancy’ during late summer, in response to increased soil moisture deficits. The differential N–RR relationship outlines contrasting coping strategies for beech and spruce in the presence of drought: Fast mobilization and use of internal C stores for new fine-root growth to ensure sustained resource uptake in beech, but reduced fine-root growth and uptake via suberization of FR in spruce to prevent resource loss.
Non-Structural Carbohydrates
Non-structural carbohydrates are crucial in mitigating drought stress in plants (O’Brien et al., 2014; Hartmann and Trumbore, 2016; Hartmann et al., 2018). During drought, there is a distinct trade-off between growth and reserve accumulation, eventually leading to a decline of NSC concentrations in above-ground organs (McDowell, 2011) with simultaneously enhanced reallocation to roots (Brunner et al., 2015). Stored carbohydrates are important particularly in deciduous species that need to rely on the stored reserves to initiate leaf and root growth (Chapin et al., 1990; Landhäusser et al., 2012). In beech rootlets from our study, TSC increased through the growing season. This dynamic was largely driven by the sucrose content of the FR, which had the highest TSC reserves during late summer, to meet the C demand for respiration and regrowth. Physiologically active FR allow an increased water uptake and transfer to aboveground organs (Badri and Vivanco, 2009; Karst et al., 2016). TSC also remained high in transport fine-roots. The observed TSC accumulation in TR may lower the osmotic potential (Smirnoff, 1995; Brunner et al., 2015; Hommel et al., 2016), allowing for prolonged functionality of the aging roots responsible for transport and storage under drought. Such enhanced accumulation of assimilates reveals a “slow” ecological strategy of transport fine-roots in beech (Figure 6). In MR, sucrose, glucose and fructose concentrations almost doubled during the second half of the growing season, probably to maintain the high ectomycorrhizal colonization under drought (Shi et al., 2002).
In spruce rootlets, lowest levels of TSC were detected late in the growing season, corresponding to the enhanced proportion of MR with depleted sugars and starch concentration in autumn. In spruce FR and TR, highest TSC concentrations were found during the dry late summer, which is in line with temporally increased soluble sugars in roots in response to drought (McDowell, 2011; Müller et al., 2016). Thus, in both fine-root categories (FR and TR) a “slow” ecological strategy to drought adaptation was detected (Figure 6). Such temporal sugar accumulation may result from photosynthesis exceeding water-limited growth demands (Körner, 2003). Drought-induced TSC accumulation may be a common mechanism for survival during periods of stress in spruce. The high root concentrations of glucose, fructose, and pinitol may not only decrease the root water potential, facilitating water absorption from dry soil (Lambers et al., 1998) but also aid in sustaining and extending the mycelial network (Ekblad et al., 2013). For example, sustaining roots with a mycorrhizal association during drought requires sucrose to be hydrolyzed to glucose and fructose, and then consumed by both the fungal partners and root cortical cells (Nehls and Hampp, 2000). Although such use of sucrose is relevant for maintaining the respiration of MR in both tree species, spruce in particular, appears to rely more heavily on its mycorrhizae to withstand drought (Paradiso et al., 2019 and references therein). Under recurring prolonged episodes of drought, the persistence of drought-adapted ECM fungi can aid in tree survival, where trees with functional associations tolerating soil water potentials as low as −3 to −5.5 MPa (Smith and Read, 2008).
In this study, fine-root starch concentrations in both tree species were 10–100-fold lower than previously reported values for adult trees under natural growing conditions (e.g., Brunner et al., 2002; Barbaroux et al., 2003; Terzaghi et al., 2016; Rosinger et al., 2020). Soil water availability differed between the two tree species starting in the spring of 2003 (Nikolova et al., 2009), a direct result of the ability of spruce to take up and transpire water before beech flushed its leaves (Beier, 1998). This resulted in a longer period of exhausted soil water availability for spruce (i.e., 75 d in spruce vs. 45 d in beech). Experimental studies show starch concentration in roots to strongly decrease with the strength of stress (Thomas et al., 2002; Braun et al., 2004), which may result in higher mortality of the non-mycorrhizal fine roots, and/or in reduced fungal diversity of the mycorrhizal roots (Shi et al., 2002; Pena et al., 2010). In beech, the low starch concentration may have predetermined the short lifespan (Marshall and Waring, 1985; Nikolova, 2007) and, thus, the ephemeral character of beech fine roots during exceptional drought. In spruce, fine roots with reduced growth and low starch reserves may eventually represent a starch degradation into glucose, needed to support the mycorrhizal network. This would be reasonable as fungi use sugar alcohols such as arabitol and mannitol to enhance their osmotic strength during drought (Shi et al., 2002), and to enlarge their mycelium to proliferate into deeper and wetter mineral soil horizons (Ekblad et al., 2013). The heavy seed production that occurred for both tree species during 2003 (Dietrich et al., in prep.) could also have induced a major reallocation of the mobile C-pool into reproductive organs (Körner, 2003), additionally declining the stored starch in roots.
Specifics of the Study
The present study was a part of a larger project, where a comprehensive investigation on the belowground effects of an experimentally enhanced ozone regime was conducted from 2002 through 2004 (Matyssek et al., 2010). While this study was not explicitly set-up to study species specific drought effects we were able to capture this response through several of our samples. The systematic measurement of root RR and other related parameters was planned only for 2003 (cv. Nikolova, 2007), and only short methodological checks on these parameters were done during the non-limited years 2002 and 2004. Nevertheless, other parameters recorded systematically from 2002 to 2004 such as the “autotrophic” and “heterotrophic” soil RR, the fine-root production and the fine-root recovery rate (Nikolova et al., 2009), as well as the total soil RR, the standing fine-root biomass, the amount of annually produced fine-root biomass and its δ13C signature (Nikolova et al., 2010) depicted contrasting responses of both species during 2003 with spruce being more effected by extreme drought compared to beech. This result is not surprising considering the different growth habits including leaf physiology, branching architecture (funnel-like in beech), rooting depth (shallow-rooted in spruce) of deciduous vs. evergreen species (Nikolova et al., 2009). But the underlying mechanisms of such species-specific adjustments to seasonal drought still remained unclear. Despite the limitation of not having reference data from “normal” conditions, the present study makes use 1/100 (1 in 100 years) drought event to analyze the belowground adaptability of these two most important Central European tree species (Ellenberg, 1996). Our investigation presents novel seasonal data on a broad range of fine-root traits and suggests possible mechanisms of adaptability, which should be tested in further experiments on adult forest trees.
Conclusions
We found for fine-roots in beech and spruce, that each species is not tied to one sole ecological strategy in coping with drought. In beech, the youngest absorptive FR followed the “fast” strategy, i.e. short-lived roots, enlarged specific fine-root area, and high root RR. High ECM colonization typical of mycorrhizal roots also represents a “fast” ecological strategy. These adjustments indicated enhanced C turnover, which facilitate effective acquisition of available belowground resources and rapid translocation of resources to aboveground organs. Transport fine-roots with developed secondary xylem, however, followed a “slow” strategy, as NSC increased during drought, possibly preventing resource efflux and root desiccation. Overall, during seasonal drought, beech fine-root traits largely reflected a “fast” strategy, particularly the youngest absorptive fine-roots.
In contrast, fine-root traits of spruce reflected largely a “slow” strategy. Suberized foraging fine-roots and the transport fine-roots had larger diameters and higher NSC levels facilitating long-term C retention during drought. Such adjustments can protect spruce fine-roots against desiccation and lower C required for respiration. However, absorptive mycorrhizal fine-roots were more indicative of the “fast” strategy. Therefore, the resource acquisition in spruce during drought seems to largely rely on mycorrhizal fungi.
The present study took advantage of the naturally occurring severe drought during the summer of 2003. However, the question remains as to what extent do beech and spruce trees recover from successive years of drought. Such scenarios are realistic considering the recent droughts of 2018 and 2019 in Central Europe. Our findings indicate that beech trees could likely suffer from C starvation during successive drought events since root regeneration can deplete the C reserves under drought-impaired photosynthesis. Spruce’s survival lies heavily in the ability of mycorrhizal communities to survive recurring drought and continue to contribute to water/nutrient uptake and plant vitality. If instead the mycorrhizal association becomes a competitor during drought (Kariman et al., 2018) and therefore a dwindling C supply in weakened trees then the fate of spruce is also likely dire.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
PN, IB, RM, K-HH, and HB conceptualized the main research questions. PN collected data and performed the data analyses. PN wrote the manuscript, RM, TB, K-HH, HB, and IB revised the manuscript.
Funding
This work was funded by the Deutsche Forschungsgemeinschaft (DFG), through SFB 607 “Growth and Parasite Defence—Competition for Resources in Economic Plants from Agronomy and Forestry”.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer IB declared a past co-authorship with one of the authors [IB] to the handling editor.
Acknowledgments
We thank Ivan Nikolov for his skillful support of the fine-root analyses, Thomas Feuerbach for his precise technical assistance, and Marc Goebel for his valuable contribution to editing the final version of this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2020.01211/full#supplementary-material
References
Agerer, R. (2001). Exploration types of ectomycorrhizae. Mycorrhiza 11, 107–114. doi: 10.1007/s005720100108
Badri, D. V., Vivanco, J. M. (2009). Regulation and function of root exudates. Plant Cell Environ. 32, 666–681. doi: 10.1111/j.1365-3040.2008.01926.x
Barbaroux, C., Bréda, N., Dufrêne, E. (2003). Distribution of above-ground and below-ground carbohydrate reserves in adult trees of two contrasting broad-leaved species (Quercus petraea and Fagus sylvatica). New Phytol. 157, 605–615. doi: 10.1046/j.1469-8137.2003.00681.x
Bauerle, T. L., Richards, J. H., Smart, D. R., Eissenstat, D. M. (2008). Importance of internal hydraulic redistribution for prolonging the lifespan of roots in dry soil. Plant Cell. Environ. 31, 177–186. doi: 10.1111/j.1365-3040.2007.01749.x
Beier, C. (1998). Water and element fluxes calculated in a sandy forest soil taking spatial variability into account. For. Ecol. Manage. 101, 269–280. doi: 10.1016/S0378-1127(97)00142-4
Bergmann, J., Weigelt, A., van der Plas, F., Laughlin, D. C., Kuyper, T. W., Guerrero-Ramirez, N., et al. (2020). The fungal collaboration gradient dominates the root economics space in plants. Sci. Adv. 6. doi: 10.1126/sciadv.aba3756
Biernacki, M., Lovett-Doust, J. (2002). Developmental shifts in watermelon growth and reproduction caused by the squash bug, Anasa tristis. New Phytol. 155, 265–273. doi: 10.1046/j.1469-8137.2002.00462.x
Blaschke, H., Nikolova, P., Raidl, S., Matyssek, R. (2006). “Ectomycorrhiza: exploration types, hyphal network, rhizomorphs, and their response to chemical and physical properties of the soil environment,” in COST 631. Handbook of Methods used in Rhizosphere Research. Eds. Luster, J., Finlay, R.(WSL Birmensdorf: Eidg. Forschungsanstalt WSL), 380–381.
Braun, S., Zugmaier, U., Thomas, V., Flückiger, W. (2004). Carbohydrate concentrations in different plant parts of young beech and spruce along a gradient of ozone pollution. Atmos. Environ. 38, 2399–2407. doi: 10.1016/j.atmosenv.2003.12.037
Brundrett, M. C. (2002). Coevolution of roots and mycorrhizas of land plants. New Phytol. 154, 275–304. doi: 10.1046/j.1469-8137.2002.00397.x
Brunner, I., Brodbeck, S., Walthert, L. (2002). Fine root chemistry, starch concentration, and ‘vitality’ of subalpine conifer forests in relation to soil pH. For. Ecol. Manage. 165, 75–84. doi: 10.1016/S0378-1127(01)00633-8
Brunner, I., Herzog, C., Dawes, M. A., Arend, M., Sperisen, C. (2015). How tree roots respond to drought. Front. Plant Sci. 6:547:547. doi: 10.3389/fpls.2015.00547
Brunner, I., Herzog, C., Galiano, L., Gessler, A. (2019). Plasticity of fine-root traits under long-term irrigation of a water-limited Scots pine forest. Front. Plant Sci. 10:701:701. doi: 10.3389/fpls.2019.00701
Burton, A. J., Pregitzer, K. S. (2002). Measurement carbon dioxide concentration does not affect root respiration of nine tree species in the field. Tree Physiol. 22, 67–72. doi: 10.1093/treephys/22.1.67
Burton, A. J., Pregitzer, K. S. (2003). Field measurements of root respiration indicate little to no seasonal temperature acclimation for sugar maple and red pine. Tree Physiol. 23, 273–280. doi: 10.1093/treephys/23.4.273
Burton, A. J., Pregitzer, K. S., Zogg, G. P., Zak, D. R. (1998). Drought reduces root respiration in sugar maple forests. Ecol. Appl. 8, 771–778. doi: 10.1890/1051-0761(1998)008[0771:DRRRIS]2.0.CO;2
Caldwell, M. M., Dawson, T. E., Richards, J. H. (1998). Hydraulic lift: consequences of water efflux from the roots of plants. Oecologia 113, 151–161. doi: 10.1007/s004420050363
Ceccon, C., Tagliavini, M., Schmitt, A. O., Eissenstat, D. M. (2016). Untangling the effects of root age and tissue nitrogen on root respiration in Populus tremuloides at different nitrogen supply. Tree Physiol. 36, 618–627. doi: 10.1093/treephys/tpw022
Chapin, III, F. S., Schulze, E., Mooney, H. A. (1990). The ecology and economics of storage in plants. Annu. Rev. Ecol. Syst. 21, 423–447. doi: 10.1146/annurev.es.21.110190.002231
Chapman, K., Groot, E., Nichol, S., Rost, T. L. (2003). Primary root growth and the pattern of root apical meristem organization are coupled. J. Plant Growth Regul. 21, 287–295. doi: 10.1007/s00344-002-0036-x
Cruz, R. T., Jordan, W. R., Drew, M. C. (1992). Structural changes and associated reduction of hydraulic conductance in roots of Sorghum bicolor L. following exposure to water deficit. Plant Physiol. 99, 203–212. doi: 10.1104/pp.99.1.203
Di Iorio, A., Giacomuzzi, V., Chiatante, D. (2016). Acclimation of fine root respiration to soil warming involves starch deposition in very fine and fine roots: a case study in Fagus sylvatica saplings. Physiol. Plant. 156, 294–310. doi: 10.1111/ppl.12363
Dubrovsky, J. G. (1997). Determinate primary-root growth in seedlings of Sonoran desert Cactaceae; its organization, cellular basis, and ecological significance. Planta 203, 85–92. doi: 10.1007/s00050168
Eissenstat, D., McCormack, M. L., Du, Q. (2013). “Global change and root lifespan,” in Plant Roots: The Hidden Half, vol. 27 . Eds. Eshel, A., Beeckman, T. (Boca Raton, FL: CRC Press), 1–27. 13.
Ekblad, A., Wallander, H., Godbold, D. L., Cruz, C., Johnson, D., Baldrian, P., et al. (2013). The production and turnover of extramatrical mycelium of ectomycorrhizal fungi in forest soils: role in carbon cycling. Plant Soil 366, 1–27. doi: 10.1007/s11104-013-1630-3
Ellenberg, H. (1996). Vegetation Mitteleuropas mit den Alpen in ökologischer, dynamischer und historischer Sicht (Stuttgart: Ulmer).
Fleischmann, F., Winkler, J. B., Oßwald, W. (2009). Effects of ozone and Phytophthora citricola on non-structural carbohydrates of European beech (Fagus sylvatica) saplings. Plant Soil 323, 75–84. doi: 10.1007/s11104-009-9927-y
Freschet, G. T., Roumet, C. (2017). Sampling roots to capture plant and soil functions. Funct. Ecol. 31, 1506–1518. doi: 10.1111/1365-2435.12883
Freschet, G. T., Violle, C., Bourget, M. Y., Scherer-Lorenzen, M., Fort, F. (2018). Allocation, morphology, physiology, architecture: the multiple facets of plant above-and below-ground responses to resource stress. New Phytol. 219, 1338–1352. doi: 10.1111/nph.15225
Gordon, W. S., Jackson, R. B. (2000). Nutrient concentrations in fine roots. Ecology 81, 275–280. doi: 10.1890/0012-9658(2000)081[0275:NCIFR]2.0.CO;2
Guijarro, J. A. (2020). Package climatol. Available at: https://cran.r-project.org/web/packages/climatol/climatol.pdf (Accessed June, 2020).
Guo, D., Mitchell, R. J., Withington, J. M., Fan, P. P., Hendricks, J. J. (2008). Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: root branch order predominates. J. Ecol. 96, 737–745. doi: 10.1111/j.1365-2745.2008.01385.x
Häberle, K. H., Weigt, R., Nikolova, P., Reiter, I., Cermak, J., Wieser, G., et al. (2012). “Case study “Kranzberger Forst”: Growth and defence in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.),” in Growth and Defence in Plants. Eds. Matyssek, R., Schnyder, H., Oßwald, W., Ernst, D., Munch, J., Pretzsch, H. (Berlin, Heidelberg: Springer), 243–271.
Hartmann, H., Trumbore, S. (2016). Understanding the roles of nonstructural carbohydrates in forest trees–from what we can measure to what we want to know. New Phytol. 211, 386–403. doi: 10.1111/nph.13955
Hartmann, H., Adams, H. D., Hammond, W. M., Hoch, G., Landhäusser, S. M., Wiley, E., et al. (2018). Identifying differences in carbohydrate dynamics of seedlings and mature trees to improve carbon allocation in models for trees and forests. Environ. Exp. Bot. 152, 7–18. doi: 10.1016/j.envexpbot.2018.03.011
Heimsch, C., Seago, J. J.L. (2008). Organization of the root apical meristem in angiosperms. Am. J. Bot. 95, 1–21. doi: 10.3732/ajb.95.1.1
Hommel, R., Siegwolf, R., Zavadlav, S., Arend, M., Schaub, M., Galiano, L., et al. (2016). Impact of interspecific competition and drought on the allocation of new assimilates in trees. Plant Biol. 18, 785–796. doi: 10.1111/plb.12461
Iversen, C. M., McCormack, M. L., Powell, A. S., Blackwood, C. B., Freschet, G. T., Kattge, J., et al. (2017). A global Fine-Root Ecology Database to address below-ground challenges in plant ecology. New Phytol. 215, 15–26. doi: 10.1111/nph.14486
Joslin, J. D., Wolfe, M. H., Hanson, P. J. (2000). Effects of altered water regimes on forest root systems. New. Phytol. 147, 117–129. doi: 10.1046/j.1469-8137.2000.00692.x
Kariman, K., Barker, S. J., Tibbett, M. (2018). Structural plasticity in root-fungal symbioses: diverse interactions lead to improved plant fitness. PeerJ 6, 1–38. doi: 10.7717/peerj.6030
Karst, J., Gaster, J., Wiley, E., Landhäusser, S. M. (2016). Stress differentially causes roots of tree seedlings to exude carbon. Tree Physiol. 37, 154–164. doi: 10.1093/treephys/tpw090
Körner, C. (2003). Carbon limitation in trees. J. Ecol. 91, 4–17. doi: 10.1046/j.1365-2745.2003.00742.x
Laliberté, E. (2017). Below-ground frontiers in trait-based plant ecology. New Phytol. 213, 1597–1603. doi: 10.1111/nph.14247
Lambers, H., Scheurwater, I., Mata, C., Nagel, O. W. (1998). “Root respiration of fast- and slow-growing plants, as dependent on genotype and nitrogen supply: a major clue to the functioning of slow-growing plants,” in Inherent Variation in Plant Growth, Physiological Mechanisms and Ecological Consequences. Eds. Lambers, H., Poorter, H., Van Vuren, M. (Leiden: Backhuys Publishers), 139–157.
Landhäusser, S. M., Pinno, B. D., Lieffers, V. J., Chow, P. S. (2012). Partitioning of carbon allocation to reserves or growth determines future performance of aspen seedlings. For. Ecol. Manage. 275, 43–51. doi: 10.1016/j.foreco.2012.03.010
Lavorel, S., Garnier, E. (2002). Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Funct. Ecol. 16, 545–556. doi: 10.1046/j.1365-2435.2002.00664.x
Leshem, B. (1970). Resting roots of Pinus halepensis: structure and function, and reaction to water stress. Bot. Gaz. 131, 99–104. doi: 10.1086/336518
Leuschner, C., Meier, I. C. (2018). The ecology of Central European tree species: Trait spectra, functional trade-offs, and ecological classification of adult trees. Perspect. Plant Ecol. 33, 89–103. doi: 10.1016/j.ppees.2018.05.003
Leuschner, C., Hertel, D., Coners, H., Büttner, V. (2001). Root competition between beech and oak: a hypothesis. Oecologia 126, 276–284. doi: 10.1007/s004420000507
Lobet, G., Couvreur, V., Meunier, F., Javaux, M., Draye, X. (2014). Plant water uptake in drying soils. Plant Physiol. 164, 1619–1627. doi: 10.1104/pp.113.233486
Ma, Z., Guo, D., Xu, X., Lu, M., Bardgett, R. D., Eissenstat, D. M., et al. (2018). Evolutionary history resolves global organization of root functional traits. Nature 555, 94–97. doi: 10.1038/nature25783
Marshall, J. D., Waring, R. H. (1985). Predicting fine root production and turnover by monitoring root starch and soil temperature. Can. J. For. Res. 15, 791–800. doi: 10.1139/x85-129
Matyssek, R., Wieser, G., Ceulemans, R., Rennenberg, H., Pretzsch, H., Haberer, K., et al. (2010). Enhanced ozone strongly reduces carbon sink strength of adult beech (Fagus sylvatica) - Resume from the free-air fumigation study at Kranzberg Forest. Environ. Pollut. 158, 2527–2532. doi: 10.1016/j.envpol.2010.05.009
McCormack, M. L., Dickie, I. A., Eissenstat, D. M., Fahey, T. J., Fernandez, C. W., Guo, D., et al. (2015). Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 207, 505–518. doi: 10.1111/nph.13363
McCormack, M. L., Guo, D., Iversen, C. M., Chen, W., Eissenstat, D. M., Fernandez, C. W., et al. (2017). Building a better foundation: improving root-trait measurements to understand and model plant and ecosystem processes. New Phytol. 215, 27–37. doi: 10.1111/nph.14459
McDowell, N. G. (2011). Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 155, 1051–1059. doi: 10.1104/pp.110.170704
Meier, I. C., Leuschner, C. (2008). Belowground drought response of European beech: fine root biomass and carbon partitioning in 14 mature stands across a precipitation gradient. Global Change Biol. 14, 2081–2095. doi: 10.1111/j.1365-2486.2008.01634.x
Mendiburu, F. (2020). Package agricolae. Available at: https://cran.r-project.org/web/packages/agricolae/agricolae.pdf (Accessed June, 2020).
Müller, K., Kramer, S., Haslwimmer, H., Marhan, S., Scheunemann, N., Butenschön, O., et al. (2016). Carbon transfer from maize roots and litter into bacteria and fungi depends on soil depth and time. Soil Biol. Biochem. 93, 79–89. doi: 10.1016/j.soilbio.2015.10.015
Nehls, U., Hampp, R. (2000). Carbon allocation in ectomycorrhizas. Physiol. Mol. Plant P. 57, 95–100. doi: 10.1006/pmpp.2000.0285
Nikolova, P. S., Blaschke, H., Matyssek, R. (2006). “Digital in situ recording of fine-root architecture and growth dynamics,” in in COST 631. Handbook of methods used in rhizosphere research. Eds. Luster, J., Finlay, R.(WSL Birmensdorf: Eidg. Forschungsanstalt WSL), 170–171.
Nikolova, P. S., Raspe, S., Andersen, C. P., Mainiero, R., Blaschke, H., Matyssek, R., et al. (2009). Effects of the extreme drought in 2003 on soil respiration in a mixed forest. Eur. J. For. Res. 128, 87–98. doi: 10.1007/s10342-008-0218-6
Nikolova, P. S., Andersen, C. P., Blaschke, H., Matyssek, R., Häberle, K. H. (2010). Belowground effects of enhanced tropospheric ozone and drought in a beech/spruce forest (Fagus sylvatica L./Picea abies [L.] Karst). Environ. Pollut. 158, 1071–1078. doi: 10.1016/j.envpol.2009.07.036
Nikolova, P. S. (2007). Below-ground competitiveness of adult beech and spruce trees: resource investments versus returns. [dissertation thesis] (Freising: Technische Universität München).
Nobel, P. S., Huang, B. (1992). Hydraulic and structural changes for lateral roots of two desert succulents in response to soil drying and rewetting. Int. J. Plant Sci. 153, 163–170. doi: 10.1086/297073
O’Brien, M. J., Leuzinger, S., Philipson, C. D., Tay, J., Hector, A. (2014). Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nat. Clim. Change 4, 710–714. doi: 10.1038/nclimate2281
Orlov, A. J. (1957). Observations on absorbing roots of spruce (Picea excelsa Link) in natural conditions. Bot. Z. USSR 42, 1172–1181.
Paradiso, E., Jevon, F., Matthes, J. (2019). Fine root respiration is more strongly correlated with root traits than tree species identity. Ecosphere 10, 11. doi: 10.1002/ecs2.2944
Pena, R., Offermann, C., Simon, J., Naumann, P. S., Geßler, A., Holst, J., et al. (2010). Girdling affects ectomycorrhizal fungal (EMF) diversity and reveals functional differences in EMF community composition in a beech forest. Appl. Environ. Microb. 76, 1831–1841. doi: 10.1128/AEM.01703-09
Perrot-Rechenmann, C. (2014). “Auxin signaling in plants,” in Molecular Biology, The Plant Sciences 2. Ed. Howell, S. H. (New York: Springer Science+Business Media), 245–268.
Pregitzer, K. S., Laskowski, M. J., Burton, A. J., Lessard, V. C., Zak, D. R. (1998). Variation in sugar maple root respiration with root diameter and soil depth. Tree Physiol. 18, 665–670. doi: 10.1093/treephys/18.10.665
Pregitzer, K. S. (2002). Fine roots of trees–a new perspective. New Phytol. 154, 267–270. doi: 10.1046/j.1469-8137.2002.00413_1.x
Pretzsch, H., Dieler, J., Matyssek, R., Wipfler, P. (2010). Tree and stand growth of mature Norway spruce and European beech under long-term ozone fumigation. Environ. Pollut. 158, 1061–1070. doi: 10.1016/j.envpol.2009.07.035
R Development Core Team (2018). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing).
Raspe, S., Grimmeisen, W., Schultze, B. (2004). Der Sommer 2003 grub dem Wald das Wasser ab. LWF aktuell 43, 4–6.
Raspe, S., Dietrich, H. P., Köhler, D., Schubert, A., Stiegler, J. (2018). Stickstoff im Überfluss. LWF aktuell 117, 21–24.
Rebetez, M., Mayer, H., Dupont, O., Schindler, D., Gartner, K., Kropp, J. P., et al. (2006). Heat and drought 2003 in Europe: a climate synthesis. Ann. For. Sci. 63, 569–577. doi: 10.1051/forest:2006043
Reich, P. B. (2014). The world-wide ‘fast–slow’ plant economics spectrum: a traits manifesto. J. Ecol. 102, 275–301. doi: 10.1111/1365-2745.12211
Rennenberg, H., Loreto, F., Polle, A., Brilli, F., Fares, S., Beniwal, R. S., et al. (2006). Physiological responses of forest trees to heat and drought. Plant Biol. 8, 556–571. doi: 10.1055/s-2006-924084
Rosinger, C., Sandén, H., Godbold, D. L. (2020). Non-structural carbohydrate concentrations of Fagus sylvatica and Pinus sylvestris fine roots are linked to ectomycorrhizal enzymatic activity during spring reactivation. Mycorrhiza 30, 197–210. doi: 10.1007/s00572-020-00939-x
Schuhbäck, T. (2004). Nährelementenstatus und Bodenzustand an der Bestandesgrenze Buche-Fichte im Mischbestand Kranzberger Forst. [masters thesis] (Freising: Technische Universität München).
Shi, L., Guttenberger, M., Kottke, I., Hampp, R. (2002). The effect of drought on mycorrhizas of beech (Fagus sylvatica L.): changes in community structure, and the content of carbohydrates and nitrogen storage bodies of the fungi. Mycorrhiza 12, 303–311. doi: 10.1007/s00572-002-0197-2
Smirnoff, N. (1995). Environment and Plant Metabolism: flexibility and acclimation (Oxford: Bios Scientific Publ).
Solly, E. F., Brunner, I., Helmisaari, H.-S., Herzog, C., Leppälammi-Kujansuu, J., Schöning, I., et al. (2018). Unravelling the age of fine roots of temperate and boreal forests. Nat. Commun. 9, 3006. doi: 10.1038/s41467-018-05460-6
Steudle, E. (2000). Water uptake by plant roots: an integration of views. Plant Soil. 226, 45–56. doi: 10.1023/A:1026439226716
Sutton, R. F., Tinus, R. W. (1983). Root and root system terminolgy. For. Sci. 29, 124–137. doi: 10.1093/forestscience/29.s1.a0001
Terzaghi, M., Montagnoli, A., Di Iorio, A., Scippa, G. S., Chiatante, D. (2013). Fine-root carbon and nitrogen concentration of European beech (Fagus sylvatica L.) in Italy Prealps: possible implications of coppice conversion to high forest. Front. Plant Sci. 4:2013.00192:192. doi: 10.3389/fpls.2013.00192
Terzaghi, M., Di Iorio, A., Montagnoli, A., Baesso, B., Scippa, G. S., Chiatante, D. (2016). Forest canopy reduction stimulates xylem production and lowers carbon concentration in fine roots of European beech. For. Ecol. Manage. 379, 81–90. doi: 10.1016/j.foreco.2016.08.010
Thomas, V. F. D., Hiltbrunner, E., Braun, S., Flückiger, W. (2002). Changes in root starch contents of mature beech (Fagus sylvatica L.) along an ozone and nitrogen gradient in Switzerland. Phyton 42, 223–228.
Tobner, C. M., Paquette, A., Messier, C. (2013). Interspecific coordination and intraspecific plasticity of fine root traits in North American temperate tree species. Front. Plant Sci. 4:242:242. doi: 10.3389/fpls.2013.00242
Wang, P., Diao, F., Yin, L., Huo, C. (2016). Absorptive roots trait plasticity explains the variation of root foraging strategies in Cunninghamia lanceolata. Environ. Exp. Bot. 129, 127–135. doi: 10.1016/j.envexpbot.2016.01.001
Warton, D., II, Duursma, R. A., Falster, D. S., Taskinen, S. (2012). smatr 3 - an R package for estimation and inference about allometric lines. Methods Ecol. Evol. 3, 257–259. doi: 10.1111/j.2041-210X.2011.00153.x
Weemstra, M., Sterck, F. J., Visser, E. J., Kuyper, T. W., Goudzwaard, L., Mommer, L. (2017). Fine-root trait plasticity of beech (Fagus sylvatica) and spruce (Picea abies) forests on two contrasting soils. Plant Soil 415, 175–188. doi: 10.1007/s11104-016-3148-y
Withington, J. M., Reich, P. B., Oleksyn, J., Eissenstat, D. M. (2006). Comparisons of structure and life span in roots and leaves among temperate trees. Ecol. Monogr. 76, 381–397. doi: 10.1890/0012-9615(2006)076[0381:COSALS]2.0.CO;2
Zadworny, M., Eissenstat, D. M. (2011). Contrasting the morphology, anatomy and fungal colonization of new pioneer and fibrous roots. New Phytol. 190, 213–221. doi: 10.1111/j.1469-8137.2010.03598.x
Zak, D. R., Holmes, W. E., MacDonald, N. W., Pregitzer, K. S. (1999). Soil temperature, matric potential, and the kinetics of microbial respiration and nitrogen mineralization. Soil Sci. Soc Am. J. 63, 575–584. doi: 10.2136/sssaj1999.03615995006300030021x
Zapater, M., Hossann, C., Bréda, N., Bréchet, C., Bonal, D., Granier, A. (2011). Evidence of hydraulic lift in a young beech and oak mixed forest using 18O soil water labelling. Trees 25, 885. doi: 10.1007/s00468-011-0563-9
Zobel, R. W., Alloush, G. A., Belesky, D. P. (2006). Differential root morphology response to no versus high phosphorus, in three hydroponically grown forage chicory cultivars. Environ. Exp. Bot. 57, 201–208. doi: 10.1016/j.envexpbot.2005.05.002
Keywords: ecological strategy, extreme drought, Fagus sylvatica, fine-root classification, fine-root traits, morphology, non-structural carbohydrates, Picea abies
Citation: Nikolova PS, Bauerle TL, Häberle K-H, Blaschke H, Brunner I and Matyssek R (2020) Fine-Root Traits Reveal Contrasting Ecological Strategies in European Beech and Norway Spruce During Extreme Drought. Front. Plant Sci. 11:1211. doi: 10.3389/fpls.2020.01211
Received: 09 April 2020; Accepted: 24 July 2020;
Published: 13 August 2020.
Edited by:
Boris Rewald, University of Natural Resources and Life Sciences Vienna, AustriaReviewed by:
Mattia Terzaghi, University of Salerno, ItalyBenjamin Delory, Leuphana University, Germany
Isabella Børja, Norwegian Institute of Bioeconomy Research (NIBIO), Norway
Copyright © 2020 Nikolova, Bauerle, Häberle, Blaschke, Brunner and Matyssek. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Petia Simeonova Nikolova, cGV0aWEubmlrb2xvdmFAd3NsLmNo