 María Carmen Martí
María Carmen Martí Ana Jiménez
Ana Jiménez Francisca Sevilla
Francisca Sevilla- Abiotic Stress, Production and Quality Laboratory, Department of Stress Biology and Plant Pathology, Centre of Edaphology and Applied Biology of Segura, Spanish National Research Council, Murcia, Spain
Plants are sessile organisms presenting different adaptation mechanisms that allow their survival under adverse situations. Among them, reactive oxygen and nitrogen species (ROS, RNS) and H2S are emerging as components not only of cell development and differentiation but of signaling pathways involved in the response to both biotic and abiotic attacks. The study of the posttranslational modifications (PTMs) of proteins produced by those signaling molecules is revealing a modulation on specific targets that are involved in many metabolic pathways in the different cell compartments. These modifications are able to translate the imbalance of the redox state caused by exposure to the stress situation in a cascade of responses that finally allow the plant to cope with the adverse condition. In this review we give a generalized vision of the production of ROS, RNS, and H2S in plant mitochondria. We focus on how the principal mitochondrial processes mainly the electron transport chain, the tricarboxylic acid cycle and photorespiration are affected by PTMs on cysteine residues that are produced by the previously mentioned signaling molecules in the respiratory organelle. These PTMs include S-oxidation, S-glutathionylation, S-nitrosation, and persulfidation under normal and stress conditions. We pay special attention to the mitochondrial Thioredoxin/Peroxiredoxin system in terms of its oxidation-reduction posttranslational targets and its response to environmental stress.
Introduction
Reactive oxygen species (ROS) are considered unavoidable byproducts of aerobic metabolism. Mitochondria play a pivotal role in plant cells providing the energy in the form of adenosine triphosphate (ATP) during oxidative phosphorylation, but also providing intermediates of the tricarboxylic acid cycle (TCA) for major biosynthetic pathways. Mitochondria are also one of the main cellular ROS source but also a source and a target of nitric oxide (NO) and derived reactive nitrogen species (RNS) (Foyer and Noctor, 2013; Lázaro et al., 2013; Albert et al., 2017; Jayawardhane et al., 2020). Balanced ROS and RNS production and scavenging is an essential characteristic of mitochondrial function for ensuring cellular growth and plant maintenance. Local changes of ROS and RNS homeostasis mediate downstream signaling events via interactions with individual proteins and signaling. Similar to ROS and RNS, a new player in mitochondrial signaling is H2S (Filipovic and Jovanović, 2017). Proteins sense redox changes and transmit information by posttranslational modifications (PTMs) of cysteine sensitive thiols, which are oxidized to proteins disulfide, trisulfides, sulfenic, sulfinic or sulfonic acid derivates, as well as S-glutathionylated and S-nitrosated forms. Additionally, H2S does not spontaneously react with reduced thiol groups but it can react with oxidized ones leading to protein persulfidation (Sevilla et al., 2015a; Wang et al., 2018; Huang et al., 2019).
In the process of ROS elimination and redox signal integration, mitochondria contain antioxidants enzymes such as Mn-superoxide dismutase (Mn-SOD) and ascorbate dependent peroxidase (APX) (Jiménez et al., 1997; Chew et al., 2003) as well as redox sensitive proteins, which are classified into two classes: redox sensors, including peroxiredoxin (PRX) and glutathione peroxidase-like (GPXL) and redox transmitters, containing thioredoxin (TRX) and glutaredoxin (GRX). However, in contrast to that occurring in plastids, cytosol, and nucleus, the TRX/PRX and glutathione/GRX systems are not very well studied in mitochondria. The scarce information about TRX/PRX system in mitochondria is striking, taking into account that posttranscriptional/translational regulation is a likely determinant of the mitochondrial stress response as well as the role of this redox system as sensor and initiator of signaling cascades in a variety of processes and stress conditions (Sevilla et al., 2015a; Zannini et al., 2018).
This review is focused on summarizing the information about ROS, RNS and H2S dependent mitochondrial PTMs, as regulatory mechanisms of plant mitochondria function under normal and stress conditions. We focus on the main enzymes from the major biological processes occurring in the mitochondria such us the ETC, the TCA and photorespiration and on how PTMs affecting cysteine residues (S-oxidation, S-glutathionylation, S-nitrosation, and persulfidation) exert their regulatory mechanisms on plant mitochondrial enzymes. We pay special attention to the mitochondrial TRX/PRX system in terms of its posttranslational control and its response to environmental stress.
Mitochondrial ROS, RNS, and H2S Generation
ROS Generation in Mitochondria
Oxygen is essential for the aerobic life and due to its chemical feature as a molecule containing two unpaired electrons it can suffer monovalent reduction generating ROS with a bimodal action. Their beneficial aspects are related to the effect on the cellular redox state and the role in signaling cascades while their detrimental aspect is related to an overproduction or insufficient endogenous antioxidant defenses which can damage all kind of cellular structures leading, in the worst case scenario, to cell death (Mhamdi and Van Breusegem, 2018). As signaling molecules, ROS have a special importance during developmental and physiological processes in plants. A redox compartmentalization exists for a proper function of ROS-dependent signaling pathways and ROS production, which depend on the reaction with other reactive species and interaction with antioxidants and scavengers (Waszczak et al., 2018; Foyer et al., 2020). Mitochondria generate energy in form of ATP by oxidative phosphorylation during glucose metabolism, a process coupled with mitochondrial respiration through a generated transmembrane potential via mitochondrial complexes I to IV of the electron transport chain (ETC), with the final result of the four-electron reduction of O2 to H2O. ROS are generated as byproducts after electron leaking at complex I and III (Hernández et al., 1993; Noctor et al., 2007) and under specific conditions at complex II during the reverse electron transport (Turrens, 2003). Thus, ROS such as superoxide anion (O2·−), hydrogen peroxide (H2O2) and hydroxyl radical (HO.) are generated after one, two and three-electron reduction, respectively. A role for complex II in H2O2 production has been reported to be even more important that complex I with a special influence under stress conditions, and also ubiquinone pool might serve as ROS site in plant mitochondria (Umbach et al., 2005; Gleason et al., 2011). A high proportion of NADH/NAD+ and/or ADP/ATP can increase the electron transport contributing to a high membrane potential, which is correlated to a higher reduction of the ETC components, increasing the probability of electron leakage to O2 and thus increasing ROS production. In the presence of transition metal ions, the more reactive HO· is formed. The O2·− generated in the mitochondrial matrix is rapidly dismutated to H2O2 by Mn-SOD contributing to the ROS production in the organelle. This H2O2 is scavenged by APX (Jiménez et al., 1997; Jiménez et al., 1998) and PRXIIF (Barranco-Medina et al., 2007). The plant ETC contains other enzymes which do not contribute to the proton gradient and act as security oxidation valves limiting ROS production: type II NAD(P)H-dehydrogenases can avoid the electron transport from complex I and II to ubiquinone while alternative oxidase (AOX) couples the ubiquinol oxidation to the O2 reduction to water and dissipation of energy in form of heat (Rasmusson and Wallström, 2010; Del-Saz et al., 2017). The increased and/or decreased expression of AOX along with the inhibition of its activity, have been considered as important factors in the establishment of the mitochondrial generation of ROS in signal transduction pathways (Vishwakarma et al., 2015) with the induction of AOX described as a mitochondrial retrograde signaling model (Maxwell et al., 1999; Lázaro et al., 2013). Evidences obtained with respiratory inhibitors and/or mutants have shown that fully functional mitochondria are also determinant for optimal photosynthesis. That dependency was linked to both cytochrome c oxidase (COX) and AOX pathway as well as to uncoupled protein (UCP) activity, which may modify the oxidation rate of ETC substrates by an increase of ubiquinol oxidation when the electrochemical proton gradient decreases across the inner membrane (Millar et al., 2011). Together with AOX, the terminal enzyme of ascorbate (ASC) biosynthesis, L-galactone-1,4-lactone dehydrogenase (GLDH), has been described to be a dual-function enzyme as structural assembly factor of complex I and as associated to the cytochrome respiratory pathway (Schimmeyer et al., 2016; Rodríguez-Ruiz et al., 2017) allowing the synthesis of ASC when coupled as an alternative electron donor for the respiratory chain. In this way, this protein may have an specific role under conditions of possible inhibition of components of the TCA as occurring under oxidative stress (Dumont and Rivoal, 2019). Interestingly, the last step of ASC biosynthesis does not produce H2O2 as it happens in animals where the activity is carried out by an oxidase instead of a dehydrogenase, altering the cellular redox state (reviewed by Fenech et al., 2019).
RNS Generation in Mitochondria
Nitric oxide (NO) refers to its nitrosyl radical (·NO). In many species, including plants, NO is a key signalling molecule that plays a role in a large number of biological processes (Kolbert et al., 2019) due to its chemistry. Reactive nitrogen species (RNS) are a family of molecules derived from NO, such as S-nitrosothiols (SNOs), S-nitrosoglutathione (GSNO), peroxynitrite (ONOO-), nitrogen dioxide (NO2), and nitro-fatty acids (NO2-FA), among others (Romero-Puertas et al., 2008; Martínez-Ruiz et al., 2013; Mata-Pérez et al., 2017).
In mitochondria both, an oxidative and a reductive pathway have been described for the NO generation. Regarding the oxidative route involving an L-Arg-dependent NOS-like activity, a mitochondrial Arabidopsis NO synthase 1 (AtNOS1) implicated in mitochondrial biogenesis was identified (Guo and Crawford, 2005). However, this enzyme was later on characterized as a small GTPase and not a NO synthase, being renamed as AtNOA1 (nitric oxide associated 1). In contrast to animals, a NOS-like enzyme has not been identified in plants (Astier et al., 2018; Kolbert et al., 2019). The reductive route of NO generation is dependent on nitrite and occurs in the inner membrane probably via cytochrome c oxidase (CCO) and/or reductase, generating ATP under hypoxia (Jayawardhane et al., 2020). Using different inhibitors, the involvement of the respiratory chain mainly complex III, CCO, and AOX was later reported although with clear results for CCO again under hypoxia (Gupta and Igamberdiev, 2016). The NO generated, can accept or loose an electron to generate nitrosyl anion (NO-) or the nitrosonium cation (NO+) or other RNS including higher oxides such as NO2 and N2O3, or it can react immediately with superoxide originated from ETC, to form peroxynitrite (ONOO−) (Leitner et al., 2009). NO can also react with GSH to form GSNO, considered as an in vivo reservoir of NO.
H2S Generation in Mitochondria
Sulfur is an essential macronutrient, taken up as sulfate and assimilated into cysteine. Volatile H2S is an inflammable gas considered as a pollutant but also as a new signaling molecule in both plants and animals (Filipovic and Jovanović, 2017). In plants it has been involved in multiple processes including response to abiotic stresses, regulation of photosynthesis, stomatal closure or autophagy (Aroca et al., 2018; Zhou et al., 2020). Opposite to ROS and RNS, H2S does not spontaneously react with reduced thiol groups, but it can react with oxidized ones leading to protein S-sulfhydration (now named persulfidation) (Paul and Snyder, 2012; Dumont and Rivoal, 2019). The increasing interest on H2S resides, among others, in that when administered exogenously, it has a positive effect in regulating plants adaptation (Zhang et al., 2009; Jin et al., 2017; Aroca et al., 2018), although the mechanism is not fully understood.
In plant systems, H2S is generated mainly in chloroplasts via the photosynthetic sulfate-assimilation pathway (García et al., 2015) although in mitochondria, the enzyme cyanoalanine synthase c1 (CAS-C1) also generates sulfide (Álvarez et al., 2012). Other enzymes described in mitochondria are certain desulfydrases which decomposes D/L-cysteine into pyruvate, H2S, and NH3 (Riemenschneider et al., 2005).
Post-Traslational Modifications Affecting Cysteine Residues: S-Oxidation, S-Glutathionylation, S-Nitrosation, and Persulfidation
S-Oxidation
The –SH group of cysteine has a pKa of about 8.3 in free amino acids. However, in oxidoreductases proteins, such as TRXs and glutaredoxins (GRXs) this pKa is lower, because it is affected by the surrounding micro-environment (Mailloux et al., 2014). Therefore, the –SH group of this type of cysteine residues will appear predominantly as more reactive thiolate and will react with close -SH groups to form a disulfide bridge (Meyer et al., 2012; Sevilla et al., 2015a). Additionally, –SH cysteine residues will suffer different oxidation states in response to redox signals mediates by ROS (Figure 1).
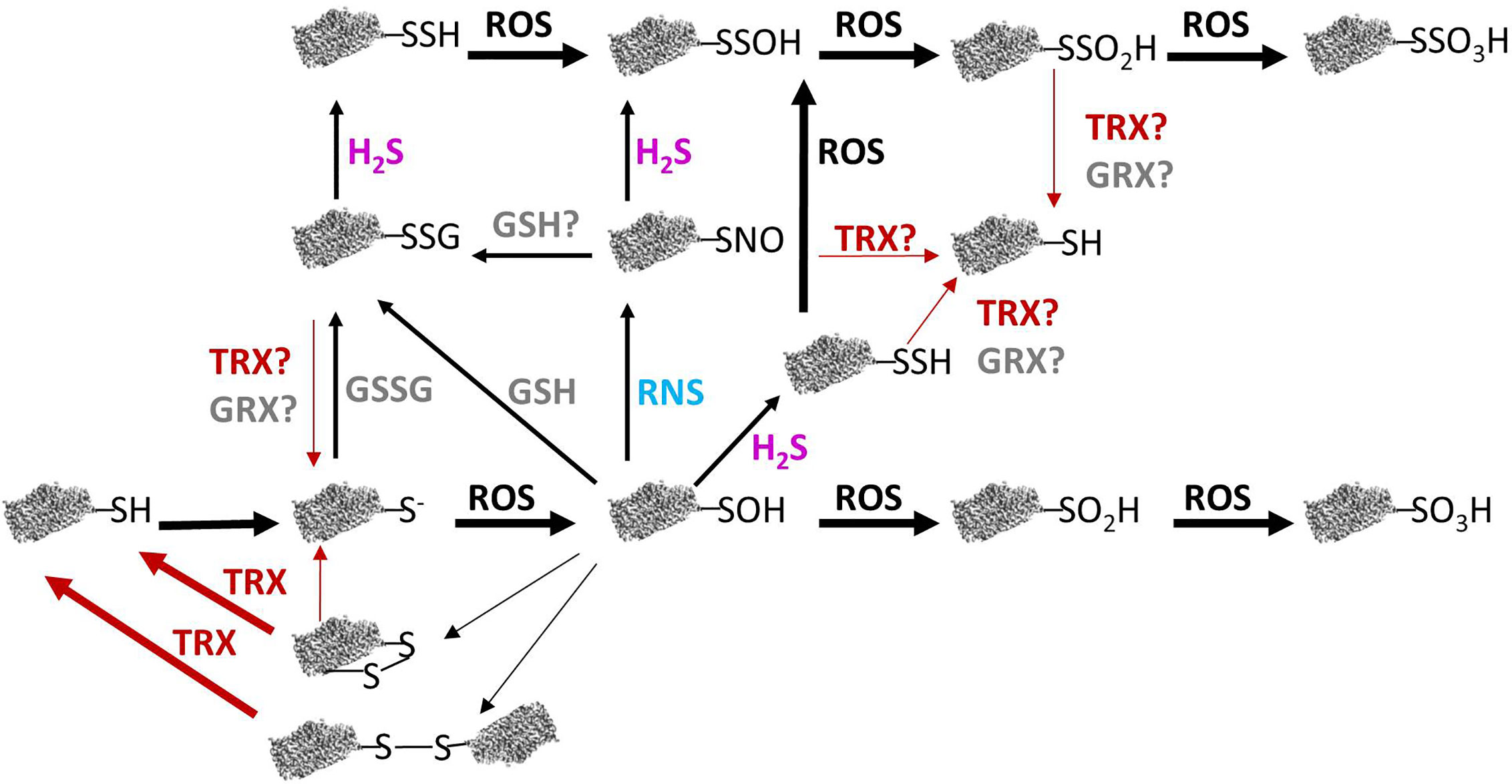
Figure 1 Relation between redox modifications of protein cysteine thiols and their reversibility in plant mitochondria. Reactive oxygen species (ROS)-mediated oxidation of protein thiols leads to different redox modifications. Reactive nitrogen species (RNS) can produce S-nitrosation of cysteine residues and GSH and GSSG provoke S-glutathionylation whereas H2S can lead to persulfidation. Some of these modifications may be reversed by TRX or GRX (see text for details). GRX, glutaredoxin; GSH, reduced glutathione; GSSG, oxidized glutathione; H2S, hydrogen sulfide; RNS, reactive nitrogen species; ROS, reactive oxygen species; R-S−, thiolate anion; R-SH, reduced thiol; R-SNO, S-nitrosation; R-SOH, sulfenic acid; RO2H, sulfinic acid; RO3H, sulfonic acid; R-SSG, S-glutathionylation; R-SSH, persulfidation; R-SSOH, persulfenic acid; R-SSO2H, persulfonic acid; R-SSO3H, persulfinic acid; R-SS-R, disulfide bond; TRX, thioredoxin.
The key player ROS that provokes oxidative reactions is H2O2, due to three main characteristics: reactivity with cysteine, prolonged half-life relative to other ROS, and capacity to diffuse through membranes. The significance of the oxidation produced by H2O2 on cysteine residues depends on the levels of H2O2. At high levels of H2O2, proteins can be oxidized irreversibly and lost their ability to function. Thus, H2O2 levels are tightly controlled by antioxidant systems to avoid overoxidation of functional proteins. In this sense, enzymes as catalase, multiple Cys-based peroxidases including PRXs, GPX, and those enzymes of ASC-GSH cycle, exert a fine control of H2O2 cellular levels (Dietz, 2011; Foyer and Noctor, 2011; Lázaro et al., 2013).
As the levels of H2O2 rise, it oxidizes the thiolate anion of protein cysteine residues to the sulfenic form (Cys–SOH). The sulfenic form can form an intra-or intermolecular disulfide bond by reaction with a neighboring thiolate or can be reduced and reverted to the thiolate form. A typical example of this kind of protein is the PRX, which by the action of TRXs and/or GRXs can be converted back to the reduced thiol group (Dietz, 2011; Sevilla et al., 2015a). Cysteine residues involved in dithiol-disulfide redox exchange frequently are located at catalytic sites of enzymes or are cysteines involved in metal binding and usually show a high degree of conservation. This dithiol-disulfide modification is transient and can act as a molecular switch altering protein activity, conformation, subcellular localization, and/or binding properties (Van Ruyskensvelde et al., 2018). In fact, protein cysteine sulfenylation is considered to play a key role in sensing and redox signalling (Huang et al., 2019). Alternatively, Cys–SOH can react with low-molecular weight thiols, such as GSH, to form S-glutathionylated disulfides. If the levels of H2O2 increases, the sulfenic form can suffer two more oxidation steps and become sulfinic (SO2H) or sulfonic (SO3H) species. The sulfinic form can be reduced by sulfiredoxin (SRX) through an ATP-dependent reaction (Iglesias-Baena et al., 2011), however the hyperoxidized sulfonic form, to date, has been shown to be irreversible (Figure 1).
S-Glutathionylation
The presence of GSH in plant mitochondria (Jiménez et al., 1997; Fernández-García et al., 2009; Foyer and Noctor, 2011) allows the posttranslational modification (PTM) of proteins via reversible S-glutathionylation of protein cysteine residues. S-glutathionylation can occur by spontaneous disulfide bond formation between the cysteine sulfenic form and GSH or a thiolate form derived from a reduced cysteine with GSSG, being the extend of the reactions dependent on the GSH/GSSG ratio in mitochondria, which is usually high in normal conditions (Dumont and Rivoal, 2019) (Figure 1). S-glutathionylation can be considered as a defense mechanism against the overoxidation of cysteine residues during oxidative stress that decreases the GSH/GSSG ratio (Zechmann, 2014; Sevilla et al., 2015b). After glutathionylation, disulfide bonds may be formed with another protein thiol and both forms can be reversed by GSH, GRX, or TRX as it has been described in animal systems (Beer et al., 2004) (Figure 1). In plants, several TRXs h present a de-glutathionylation activity although with a lower efficiency than GRXs as demonstrated for Arabidopsis cytosolic glyceraldehyde-3-phosphate dehydrogenase (Bedhomme et al., 2012), explaining the absence of GRX in plant mitochondria with de-glutathionylation activity (Zannini et al., 2018). In fact, mitochondrial TRX h undergoes glutationylation and the thiolation alters its redox potential as identified by MS (Gelhaye et al., 2004).
S-Nitrosation
The presence of RNS in mitochondria and the PTMs that they produce, allow mitochondria to be involved in many biological processes. RNS exert their role mainly by protein tyrosine nitration, metal nitrosylation and S-nitrosation (Romero-Puertas et al., 2008; Camejo et al., 2013; Corpas et al., 2013; Camejo et al., 2015; Lamotte et al., 2015). Tyrosine nitration is mediated by ONOO- and NO2 which are generated respectively, by reaction of O2.- and O2 with NO. During this process, a 3-nitrotyrosine group is produced by addition of a nitro group at the ortho position of the phenolic group of tyrosine (Ischiropoulos et al., 1992). During metal nitrosylation, NO binds to transition metals in metalloproteins (Keilin and Hartree, 1937). Finally, the most important pathway by which RNS execute their action is by protein S-nitrosylation. S-nitrosylation can occur by different mechanisms depending of the NO source, all of them resulting in the addition of a NO group to a cysteine residue in the target protein (Figure 1). During S-nitrosation, the S-nitrosothiol can be formed through NO reaction with thiyl radicals formed from one electron oxidation of thiolates via NO2 or NO2 and NO can react to form N2O3, which reacts with the cysteine thiolate (Hess et al., 2005). Additionally, S-nitrosation can occur via trans-nitrosylation, by which an S-nitrosylated group transfers its NO to another thiolate group or to GSH to form the NO reservoir GSNO, which in turn can also transfer its NO to S-nitrosylate proteins or its GSH to S-glutationylate proteins. In fact, GSNO is a mobile NO pool which effectively transduces NO signalling. To do that, GNSO levels are tightly controlled by its production and degradation. Previously we mentioned that GSNO is formed by the transfer of NO to GSH. GSNO degradation can be via non-enzymatic and via GSNO reductase (GSNOR) enzymatic activity (Lindermayr, 2018). S-nitrosylation and therefore, GSNOR, can regulate a large amount of cellular functions and signaling events due to the capacity of alter the activity, stability, conformation, interactions with other molecules or subcellular localization of the S-nitrosated proteins, playing an essential role to protect cells under nitrosative stress (Sevilla et al., 2015a; Romero-Puertas and Sandalio, 2016; Corpas et al., 2019a).
Persulfidation
H2S can exert its function through persulfidation, a PTM affecting the thiol group of cysteine (-SH) in proteins to be modified into a persulfide group (-SSH) (Gotor et al., 2019; Sandalio et al., 2019). H2S reacts with either disulfides or sulfenic acids to yield persulfides which are highly reactive to ROS, being oxidized to perthiosulfenic acids (–SSOH), perthiosulfinic (–SSO2H), and perthiosulfonic (–SSO3H) acids (Figure 1). Oxidized persulfides, can be then reduced back to their thiol forms by TRXs or GRXs (Wedmann et al., 2016; Zhou et al., 2020), so the presence in plant mitochondria of TRXo1 (Martí et al., 2009) may be a key component of the action of this PTM in the organelle, allowing the recycling and reusing of H2S by the cell.
Mitochondrial Targets of S-Oxidation, S-Glutathionylation, S-Nitrosylation, and Persufidation
In this review, we focus on the cysteine S-oxidation described for mitochondrial TRX targets and other cysteine PTMs that affect those proteins and/or proteins that are key players in the TCA, the ETC and photorespiration (Figures 2, 3).
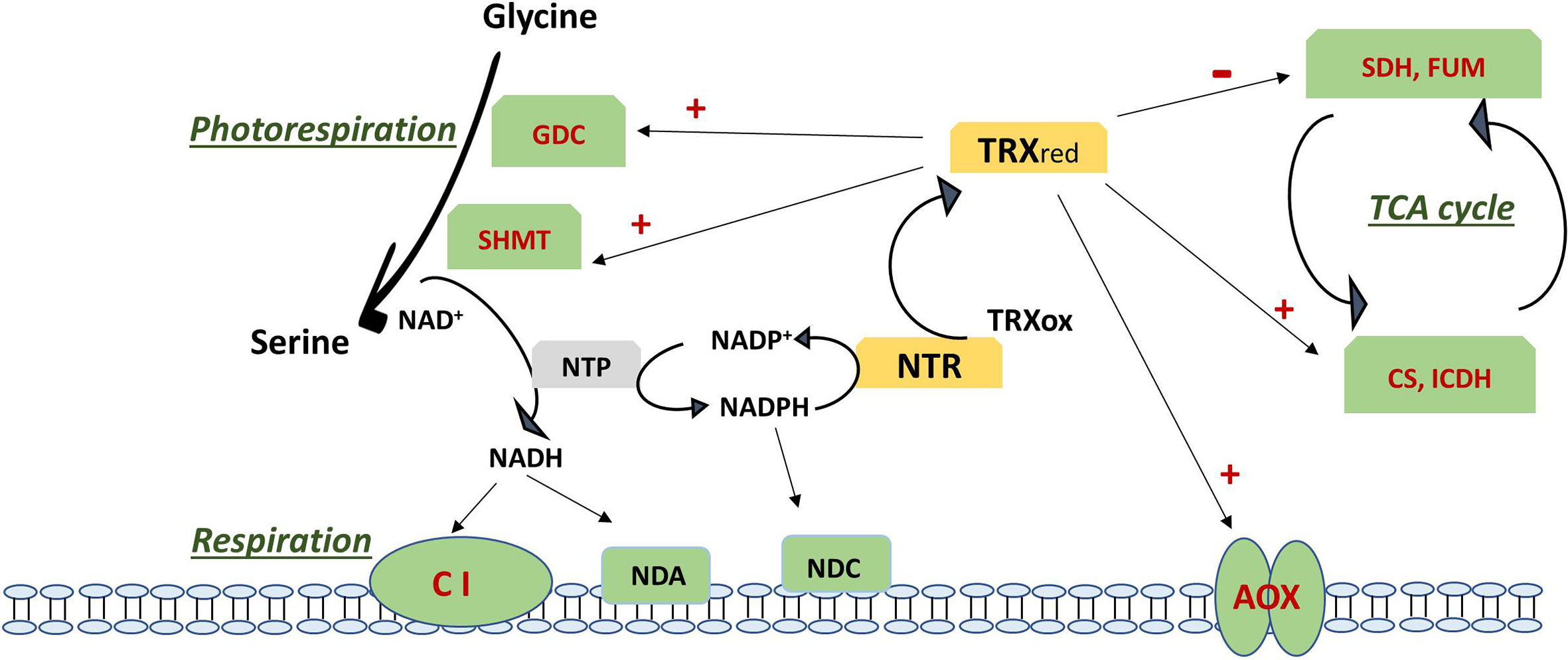
Figure 2 Regulation of photorespiration, respiration and TCA cycle via the action of thioredoxin on specific targets (in red). Negative regulation is shown as – and positive regulation as +. TCA cycle is regulated by inhibition of SDH and FUM and the activation of CS and ICDH. Respiration is regulated via activation of AOX and by the NADH generated by an active photorespiration with the Gly to Ser conversion regulated by the TRX-mediated activation of GDC an SHMT, which generates an increase in NADH. NADH excess can be oxidised via complex I and internal dehydrogenase NDA or can be transhydrogenated by NTP with formation of NADPH. NADPH is used by NTR to reduce back TRX from its oxidised form or can be oxidized via the internal dehydrogenase NDC. Updated scheme based on the previously reported one (Bykova and Igamberdiev, 2016). AOX, alternative oxidase; CI, complex I; CS, citrate synthase; FUM, fumarase; GDC, glycine decarboxylase complex; ICDH, isocitrate dehydrogenase; NDA, internal NADH dehydrogenase; NDC, internal NADPH dehydrogenase; NTP, NAD(P)transhydrogenase; NTR, NADPH thioredoxin reductase; Ox, oxidized; Red, reduced; SDH, succinate dehydrogenase; SHMT, serine hydroxymethyl transferase; TCA, tricarboxylic acid; TRX, thioredoxin.
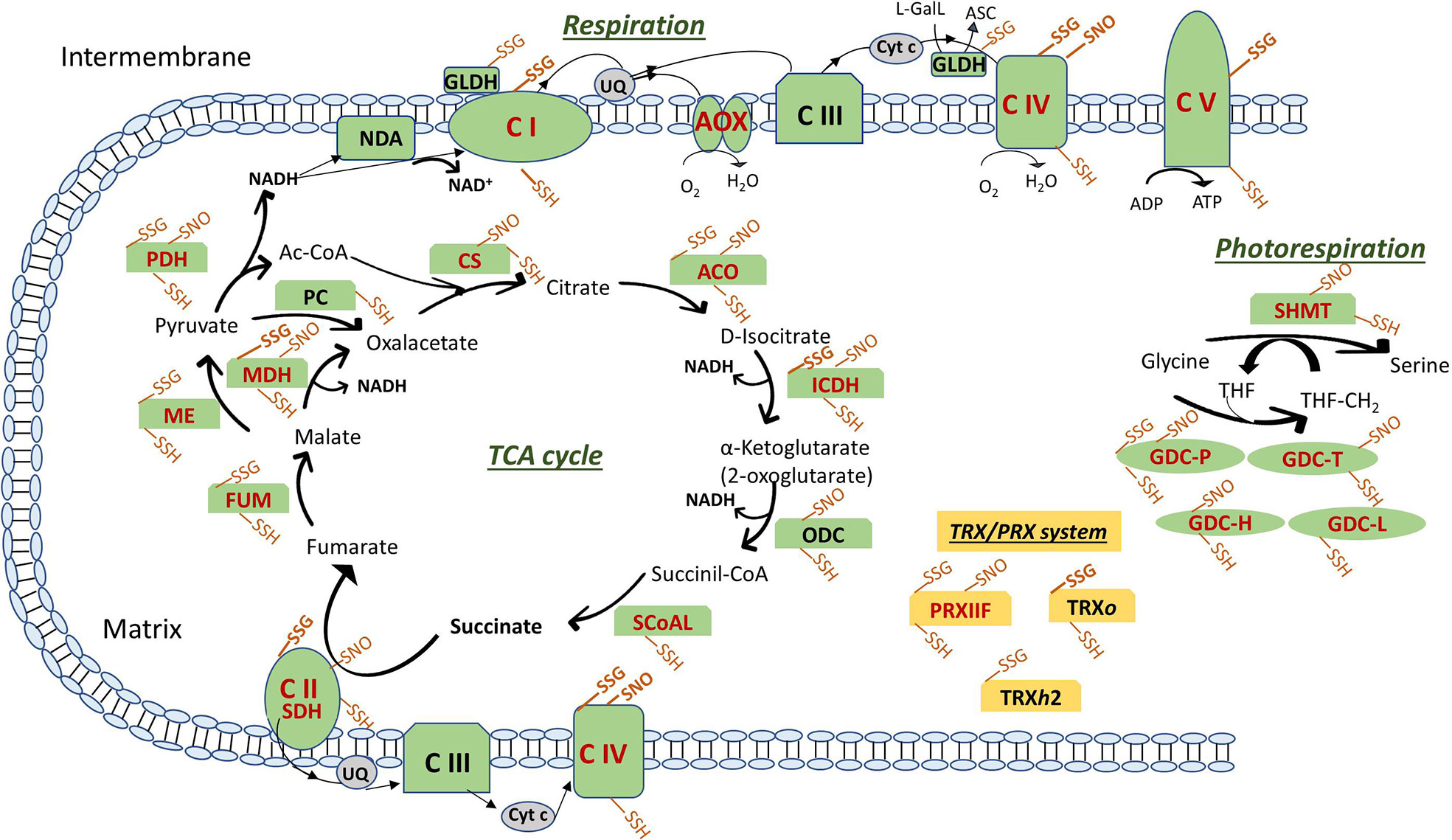
Figure 3 Posttranslational modifications of cysteine residues of ETC components, TCA cycle, photorespiration enzymes and TRX/PRX system in plant mitochondria. Modifications are shown as SSG (S-glutathionylation), SNO (S-nitrosylation) and SSH (Persulfidation). In bold the modifications described only in mitochondria from animal systems. Enzymes are in red when reported as thioredoxin targets (see text for details and references). ASC, ascorbate; ETC, electron transport chain: NDA, internal NADH dehydrogenase; Complex (C) I: NADH Dehydrogenase; CII: SDH, succinate dehydrogenase; CIII: Cytochrome (Cyt) b-c reductase; CIV: Cyt c oxidase; CV: ATP synthase; AOX: alternative oxidase; GLDH, L-galactono-1,4-lactone dehydrogenase; L-GalL, L-galactono-1,4-lactone; UQ, ubiquinone. TCA, tricarboxylic acid cycle: ACO, aconitase; CS, citrate synthase; FUM, fumarase; ICDH, isocitrate dehydrogenase; MDH, malate dehydrogenase; ME, malic enzyme; ODC, 2-oxoglutarate decarboxylase; PC, pyruvate carboxylase; PDH, pyruvate dehydrogenase; SCoAL, Succinyl-CoA synthetase. Photorespiration: GDC, glycine decarboxylase (P, T, H and L proteins); SHMT, serine hydroxymethyltransferase; THF, tetrahydrofolate; THF-CH2, N-5, N-10-methylenetetrahydrofolate. TRX/PRX system: PRX, peroxiredoxin; TRX, thioredoxin.
Targets of S-Oxidation
TRXs are small ubiquitous proteins with a conserved CPGC motif in the active center enabling the reduction of disulfides bonds of specific target proteins through a dithiol-disulfide exchange mechanism. In Arabidopsis, at least 20 TRX genes have been reported, classified into eight subgroups (f, h, m, o, s, x, y, and z-type) (Alkhalfioui et al., 2008; Meyer et al., 2012). The mitochondrial TRX isoforms belong to the TRXh and TRXo groups. The presence of TRX and NADPH-TRX reductase (NTR) activities was initially detected in plant mitochondria (Marcus et al., 1991; Konrad et al., 1996), but the native proteins were not identified and characterized. Laloi et al. (2001) identified AtTRX-o1 and AtTRX-o2 genes encoding two TRXo isoforms in Arabidopsis, being TRXo1 unequivocally present in mitochondria whereas the TRXo2 cellular location is still not clear (Zannini et al., 2018). Later on, Gelhaye et al. (2004) found that poplar mitochondria also contain a TRX isoform belonging to the TRXh type (PtTRXh2), earlier identified in the cytosol. More recently, a new TRXo1 gene was identified in Pisum sativum L. and located in both mitochondria and nucleus under physiological non –stress conditions (Martí et al., 2009). TRXs regeneration relies on the NTRA and/or NTRB being both genes expressed in mitochondria and cytosol, although NTRA has been also found in the nucleus (Reichheld et al., 2005).
In Arabidopsis, a recent structure-function analysis of both TRXo isoforms, has demonstrated that both recombinant proteins expressed in E. coli bind a Fe-S cluster. In TRXo2, the Fe-S cluster ligation depends on the cysteine residues found in the active conserved motif, but the physiological relevance of this observation remains unclear. Furthermore, it was observed a possible connection between TRXs o with the mitochondrial Fe-S cluster as both TRXs o shown the same capacity to reduce oxidized nitrogen-fixation-subunit-U 4 (NFU4) or NFU5. These NFU proteins were previously isolated as putative TRX partners and known to participate in the maturation of certain mitochondrial Fe-S proteins (Braymer and Lill, 2017; Ciofi-Baffoni et al., 2018).
An important advance in the research on the physiological role of TRX in plants has been carried out in the last two decades, but the specific role in mitochondria remains to be fully elucidated and only Arabidopsis and pea TRXo1 have been deeper studied. Several redox-proteomics studies using TRX affinity chromatography approaches have identified potentially redox regulated proteins in plant mitochondria. Thus, the TRXo1 function has been related with mitochondrial processes including, the respiratory AOX pathway, the detoxification of ROS via mitochondrial PRXIIF, the TCA cycle enzymes, the energy (ATP) synthesis and protein translation (Balmer et al., 2004; Barranco-Medina et al., 2008; Martí et al., 2009; Yoshida et al., 2013). In parallel, transcriptional regulation and redox regulation via TRXs is believed to play a crucial role in adapting photorespiration to changing environmental conditions (Reinholdt et al., 2019a).
Over-reducing conditions of mitochondrial ETC activate AOX by promoting the conversion of its dimeric oxidase form into the active reduced dimers as a consequence of the reduction of a functional disulfide bond (Day et al., 1994). Although AOX protein has not been identified as a putative mitochondrial TRX target using affinity chromatography, it has been shown to be activated by TRX-dependent reversible thiol-disulfide switch, being the mitochondrial TRX the candidate to this redox regulation (Lázaro et al., 2013; Selinski et al., 2018). AOX reduction and activation by mitochondrial PtTRXh2 and its effector pyruvate was initially described (Gelhaye et al., 2004; Umbach et al., 2005). Further, the addition of recombinant NADPH/NTR/PsTRXo1 to isolated pea and soybean mitochondria preparations induced both the reduction of AOX homodimers through the thiol redox switch of the protein and the activation of the AOX capacity (Martí et al., 2009). A similar induced AOX thiol redox switch has been linked to recombinant TRXo1 system in Arabidopsis and recently in thermogenic skunk cabbage (Yoshida et al., 2013; Umekawa and Ito, 2019). However, aside from these studies in isolated mitochondria, there is still no conclusive evidence showing that AOX is indeed regulated by the TRX system in vivo (Sevilla et al., 2015a; Geigenberger et al., 2017) (Figure 2).
PRXIIF is considered as partner of TRXo and was captured by PsTRXo1 affinity chromatography using mitochondrial preparations (Martí et al., 2009). PRXs are dimeric thiol-based peroxidases involved in H2O2 and alkyl hydroperoxide reduction to water and the corresponding alcohol, respectively. PRXs play an important role as redox sensors, antioxidant defense and in signaling. The mitochondrial location of Arabidopsis and pea PRXIIF was reported by Finkemeier et al. (2005) and Barranco-Medina et al. (2007), respectively. A biochemical characterization analysis, the use of recombinant proteins and microcalorimetric (ITC) titrations techniques reported the binding of PsTRXo1 and PsPrxIIF, sustaining a role of TRXo1 as a physiological electron donor to PrxIIF (Barranco-Medina et al., 2007; Barranco-Medina et al., 2008). A common characteristic for PRXs from plants and mammalians is the presence of at least one conserved cysteine at the active site called peroxidatic cysteine Cp (Barranco-Medina et al., 2007; Dietz, 2011; Toledo et al., 2011). Under the normal catalytic mechanism, reaction of Cp with H2O2 is very fast generating the sulfenic acid (Cys-SOH), which then form an inter-or intramolecular disulfide bond with a resolving cysteine (Cr) (Toledo et al., 2011). The disulfide or sulfenic from of PRXs is subsequently reduced by TRX/GRX (Barranco-Medina et al., 2009; Meyer et al., 2012). The cysteine sulfenic form may be overoxidized to the sulfinic form (Cys-SO2H), which can be reduced by SRX in the presence of ATP. Finally, a thiosulfinate intermediate between both PRX and SRX is formed (Iglesias-Baena et al., 2010). The regeneration step is catalyzed by a thiol reductant like NTR/TRX and/or GSH (Iglesias-Baena et al., 2011; Liebthal et al., 2018). In pea and Arabidopsis plants, the small thiol reductase protein, SRX has also been described to be localized in mitochondria and in chloroplasts (Iglesias-Baena et al., 2010; Iglesias-Baena et al., 2011). In mitochondria, SRX presents a broader specificity than in chloroplasts, catalyzing the retro-reduction of hyperoxidized (sulfinic) atypical mitochondrial PRXIIF and atypical human PRXV (Rey et al., 2007; Iglesias-Baena et al., 2010; Iglesias-Baena et al., 2011).
Some TCA enzymes were described to be highly susceptible to oxidative stress as citrate synthase (CS) and isocitrate dehydrogenase (ICDH) (Geigenberger et al., 2017). CS activity in Arabidopsis was demonstrated to be inhibited by oxidation (Stevens et al., 1997). Later on, the mitochondrial CS isoform was also shown to be regulated by TRXo (Schmidtmann et al., 2014). In contrast to other TCA cycle enzymes, CS is exclusively localized in mitochondria in green tissues, so the TCA cycle cannot be bypassed via cytosolic isoforms in those tissues. By site-directed mutagenesis of its six cysteine residues the authors reported that oxidation inhibited enzyme activity by the formation of diverse disulfide bridges, as the partially oxidized enzyme forms large redox-dependent aggregates. TRX can cleave those intra- and intermolecular disulfide bridges, reversing the enzyme to the active state and strongly enhancing the activity (Figure 2). Daloso et al. (2015) described an activation by TRXo1 in vitro although an increase in the leaf enzyme activities in trxo1 and ntra ntrb mutants suggested that this activation may not occur in vivo. Mitochondrial NAD+-dependent isocitrate dehydrogenase (mICDH) was also captured using TRX affinity chromatography. This enzyme forms only intramolecular disulfide bridges under oxidizing conditions (Yoshida and Hisabori, 2014). Recombinant Arabidopsis mICDH has a heterodimeric structure composed of two subunits: ICDH-c and ICDH-r and unlike yeast ICDH, it did not show adenylated-dependent enzyme activity. Upon oxidation of the subunit ICDH-r, ICDH activity was largely diminished via intermolecular disulfide–mediated oligomer formation of ICDH-r. A recent study has confirmed that AtTRXo was effective in the reduction of oxidized ICDH-r likely leading to a recovery of ICDH activity (Yoshida and Hisabori, 2014) (Figure 2). In the first studies using the double T-DNA mutant of Arabidopsis with downregulated NTRA and B expression (ntra ntrb) and the mitochondrial located thioredoxin o1 (trxo1) mutant, Daloso et al. (2015) demonstrated that the mitochondrial TRX system regulates in vivo the activity of different enzymes of the TCA cycle. In particular, the mitochondrial TRXo1 deactivates fumarase (FUM) and succinate dehydrogenase (SDH) both previously reported as potential TRX target proteins (Yoshida et al., 2013). These enzymes bear a considerable proportion of the flux control of TCA pathway, with SDH linking the TCA cycle and ETC. Curiously, TRXo activates the cytosolic citrate lyase. On the basis on these findings, the authors point out a role of mitochondrial TRXo as crucial mediator of mitochondria-cytosol cross-talk controlling carbon fluxes in the cell (Daloso et al., 2015). The enzyme ATP-citrate lyase is involved in fatty acid biosynthesis, a noncyclic mode of TCA function, using citrate exported from the mitochondria. It has been suggested that this TRXo1/citrate lyase regulation might be favored under reducing conditions characterized by a high NAD(P)H/NAD(P)+ ratio, which allows a deceleration from the TCA cycle (Møller, 2015) (Figure 2). Other TCA cycle enzyme that was trapped on TRX affinity columns is malate dehydrogenase (MDH). In plants, two mitochondrial NAD-dependent MDHs have been described with a role in central metabolism and redox homeostasis (Selinski et al., 2014; Huang et al., 2017). Initially, it was reported that mitMDH could be regulated via mitochondrial TRXo. However, Daloso et al. (2015) described an unaltered effect by TRXo1 finding similar activity in WT and trxo1 and ntra ntrb mutant plants (Figure 2). Additionally, mitMDH activity has been shown to be regulated by adenin nucleotides (Yoshida and Hisabori, 2016).
Photorespiration or “respiration in light” links photosynthetic carbon assimilation with nitrogen and sulfur assimilation and carbon metabolism, contributing to balancing the redox state between mitochondria, chloroplasts, peroxisomes and cytoplasm. In mitochondria, a key step in photorespiration is the conversion of glycine from peroxisomes into serine which will be back to peroxisomes for its deamination (Engel et al., 2007). This step is carried out by the action of a multienzyme system composed of the complex glycine decarboxylase (GDC, with 4 subunits: P, T, L and H) and serine hydroxymethyl transferase (SHMT; Bauwe and Kolukisaoglu, 2003). The NADH generated by GDC is oxidized by the respiratory ETC leading to higher NADH/NAD+ ratio in the organelle and increased cytosolic ATP/ADP under photorespiratory conditions (Eisenhut et al., 2019) (Figure 2). GDC proteins have been identified as potential targets of mitochondrial TRXo1 in different plants as well as in Synechocystis PCC 6803. In pea, mitochondrial P and T components were identified as PsTRXo1 targets together with SHMT (Martí et al., 2009). More recent research has shown that, the lack of TRXo1 caused a slow-down of GDC activity with an impaired glycine to serine turnover. Results evidenced that TRXo1 contributes to redox-regulation of GDC due to TRXo-mediated redox regulation of lipoamide dehydrogenase (GDC-L or LPD), since its activity decreases in plant and cyanobacteria when reduced via NTRA/TRXo1 system. Redox regulation of LPD has also been suggested in Chlamydomonas reinhardtii (Pérez-Pérez et al., 2017). It was concluded that TRXo1 contributes to adjust photorespiration in response to environmental fluctuations via the regulation of GDC and possibly other mitochondrial multienzyme systems in which LPD is involved (Reinholdt et al., 2019a). Not only TRXo1, but also AtTRXh2, contributes to the redox regulation of mitochondrial photorespiratory metabolism (Da Fonseca-Pereira et al., 2019a) (Figure 2). AtTRXh2 regulates the redox state of GDC-L protein, which is altered in trxh2 mutants in vivo. The recombinant TRXh2 can also deactivate GDC-L in vitro. Decreased abundance of SHMT and GDC H and L subunits as well as reduced NADH/NAD+ ratio, were also found.
All the described evidences allow us to conclude that a consistent overall picture is emerging indicating that central carbon metabolism is controlled by protein thiol switches that affect enzymatic activity. To determine the physiological and molecular function of mitochondrial TRX on the in vivo regulation of the identified targets and metabolism, plant mutants have been recently probe to be useful, therefore plants mutants needs to be analyzed further.
Targets of Glutathionylation
Glutathionylation in cells represents the major form of S-thiolation mainly occurring under oxidative stress conditions although it may also be important under normal conditions and the exact mechanism in vivo is not clearly elucidated (Gao et al., 2009). Several methods have been developed to glutathionylate proteins in vitro and also to induce glutathionylation by oxidative stress in proteomic studies. Among them, GSSG-biotin is considered a component of oxidation because of the shift in the glutathione redox towards the oxidized state (Brennan et al., 2006). Other glutathionylating agents as GSH-biotin or biotin-amide that do not induce oxidative stress are usually used together with oxidants as diamide, ter-buthyl hydroperoxide (TBHP) or H2O2. Also, anti-GSH antibody is used to identify thiolation in protein extracts or in specific purified proteins after immunoprecipitation and western blot assay, although a major concern is related to its specificity and sensitivity (Gao et al., 2009).
Some TCA cycle enzymes have been identified as thiolated such as FUM, malic enzyme and pyruvate decarboxylase in Arabidopsis cell cultures treated with GSSG-biotin and TBHP and identification by MS (Dixon et al., 2005). Glutathionylation may be also important to modulate ACO activity under oxidative and nitrosative stress, due to the fact that apart from the cysteine thiol oxidation to sulfonic acid, using anti-GSH, the purified enzyme was shown to be glutathionylated, and both modifications decreased its activity (Han et al., 2005). Other TCA enzymes identified as glutathionylated but in different animal systems are succinate dehydrogenase (SDH) and MDH (Kil and Park, 2005; Niture et al., 2005; Chen et al., 2007) (Figure 3).
Among respiratory enzymes, in plants only GLDH is sensitive to oxidative stress induced by H2O2 (Leferink et al., 2009) due to the modification of Cys-340 presumably via reversible S-glutathionylation. In this way, glutathione (GSH) may protect the enzyme from overoxidation allowing production of ASC. However, in animal systems, the glutathionylation of mitochondrial respiratory proteins such as complex I, II, ATP synthase, and COX have been described as a modulator of mitochondrial redox status (Fratelli et al., 2003; Chen et al., 2007; Hurd et al., 2008; García et al., 2010) (Figure 3).
Related to glutathionylation of photorespiratory enzymes in mitochondria, the inhibition of GDC-P subunit activity by glutathionylation has been described by Palmieri et al. (2010) using partially purified P subunit from Arabidopsis leaf mitochondria. The fact that NO is also able to modulate GDC activity suggests the possibility of certain competitive effect of S-glutathionylation and S-nitrosation which may depend of the susceptibility of each protein to undergo these modifications (Figure 3).
Some possible targets of TRXs has been identified as glutathionylated in mammals, so the overlap between regulation by TRX and glutathionylation may be important to better understand the complex network of redox-regulated processes in plant cells (Gao et al., 2009). Human thioredoxin was identified as glutathionylated and the modification likely decreased its activity (Casagrande et al., 2002). In plants, Arabidopsis chloroplastic TRXf was identified as glutathionylated (Michelet et al., 2005) and interestingly, thiolation of poplar mitochondrial TRXh2 in an additional cysteine not belonging to the active site was shown to increase the redox potential of the enzyme (Gelhaye et al., 2004) (Figure 3). Also, PRXs undergo glutathionylation as in pea chloroplastic 2-Cys PRX in which the glutathionylation of the decameric form induced a change to its dimeric glutathionylated form (Calderón et al., 2017b) or in the poplar 1-Cys PRXIIB with a conformational dissociation of homodimers into monomers (Noguera-Mazon et al., 2006). This suggested the existence of a redox-dependent oligomerization switch in the PRX family by thiolation although it does not occur for all PRXs: this is the case of mitochondrial PRXIIF in which glutathionylation did not provoke a change in its oligomeric state, being GSSG able to glutathionylate both peroxidatic (Cys59) and resolving (Cys84) cysteine, provoking a decrease in the peroxidatic activity of the protein (Calderón et al., 2017b) (Figure 3). Interestingly, pea 2-Cys PRX and PRXIIF were differentially sensitive to small changes in GSH/GSSG ratio pointing to a fine regulation by the intensity of the oxidative stress. This could be a key step during cellular metabolism but mainly during stress conditions modulating the peroxidase activity and thus affecting H2O2 signaling. Both pea chloroplastic 2-Cys PRX and mitochondrial PRXIIF are regenerated from their overoxidized forms by pea SRX, which is then reduced by TRX (Iglesias-Baena et al., 2010; Iglesias-Baena et al., 2011). Interestingly, SRX deglutathionylated pea 2-Cys PRX but not PRXIIF, so the glutathionylation/deglutathionylation processes may have an important role during plant development and response to stress in which redox changes influence the posttranslational regulation of key proteins as the ones involved in the TRX/PRX/SRX system.
Targets of S-Nitrosation
A large number of mass spectrometry (MS)-based analytical methods has been developed to face the challenge of analyzing S-nitrosylated proteins. However, the most often-used method is the biotin-switch assay (BST) (Jaffrey and Snyder, 2001). This methodology includes firstly the blocking of all the free thiols groups of cysteine residues, then selective reduction of modified cysteine and switch with a stable functional group usually with a biotin molecule that also serves as a handle for enrichment and detection. Despite this method is time consuming, tedious, and unsuitable for monitoring dynamic changes of this PTM in vivo (Feng et al., 2019), it has allowed to gain insight into the processes affected by S-nitrosation and into the study of the mechanism and effect that this PTM has on protein functions. Recently, newer strategies have been reported to overcome these issues. A S-nitrosoproteomic analysis using iodo-TMT (iodo-tandem mass tag) labelling, affinity enrichment, and high-resolution LC-MS/MS has been used to identify S-nitrosated proteins in tea plants (Qiu et al., 2019). Also, in animals, a new chemical proteomics strategy for quantitative analysis of reversibly modified cysteine using bioorthogonal cleavable-linker and switch technique (Cys-BOOST) has been reported. This method shows a higher sensitivity and considerably higher specificity and precision than other methods. It allows the proteome wide identification of SNO even from low abundance proteins and under basal conditions (Mnatsakanyan et al., 2019).
Similar to glutathionylation, several TCA enzymes including ACO, MDH, ICDH, α-ketoglutarate dehydrogenase, FUM, CS, and pyruvate dehydrogenase have been found to be S-nitrosylated in several plant species using different methodological approaches (Fares et al., 2011; Puyaubert et al., 2014; Hu et al., 2015; Lindermayr et al., 2015; Qiu et al., 2019) (Figure 3). However, the mechanisms by which this PTM happens, how S-nitrosation affects the enzymes activity or how S-nitrosation is affected by stress conditions are not well known. Previously it has been reported that ACO is reversibly inhibited by NO by a mechanism which promotes the loss of the iron-sulfur cluster, which can subsequently be reassembled under the proper conditions (Drapier, 1997). Recently it has been also shown that this inhibition under hypoxia results in accumulation of citrate, the latter in turn induces AOX and causes a shift of metabolism towards amino acid biosynthesis (Gupta et al., 2012). Regarding MDH, it has also been found to be S-nitrosated in peroxisomes which activity has been shown to be inhibited by NO donors (Ortega-Galisteo et al., 2012). Additionally, ICDH has been reported to be slightly over-nitrosylated in Arabidopsis cell suspension submitted to salt stress (Fares et al., 2011). In plants, the effect of S-nitrosation on ICDH has not been described unlike in animals, where this PTM leads to an inactivation of the enzyme and to a pro-oxidant condition in the cell (Yang et al., 2002; Lee et al., 2003).
SDH in plants is susceptible to S-nitrosation in vivo (Fares et al., 2011; Hu et al., 2015; Qiu et al., 2019) (Figure 3) under normal but not under saline stress conditions (Camejo et al., 2013). To our knowledge, in plants the effect of the S-nitrosylation in SDH activity is not known, although an inhibitory effect has been described in animals (Rizza et al., 2016).
Among the enzymes involved in the photorespiratory process, GDH has been reported to be S-nitrosated in vivo in Arabidopsis (Palmieri et al., 2010; Puyaubert et al., 2014; Hu et al., 2015) (Figure 3). As previously mentioned, in vitro studies have shown the effect of S-nitrosation on GDC-P subunit activity (Palmieri et al., 2010). Additionally, our laboratory has also found that in pea, both subunits P and T of GDC showed the same S-nitrosation pattern in control plants and salt-treated plants, with no changes in protein levels during plant development and salt stress, although loosing S-nitrosation in older plants (Camejo et al., 2013). S-nitrosation of SHMT has been reported by Camejo et al. (2013); Hu et al. (2015) and Tanou et al. (2009) in Arabidopsis, pea, and citrus plants, respectively (Figure 3). Despite of previous report where photorespiration has been shown to increase under salinity (Hoshida et al., 2000), our lab has reported that SHMT did not change its protein levels during salt stress although it was found S-nitrosated in control pea mitochondria but not in those from stressed plants. We hypothesized that its denitrosation after long period of salt stress may allow photorespiration to be functional under these conditions.
Among the RNS targets in plant mitochondria are some ETC components, with NO inhibiting cytochrome c pathway whereas AOX is only partially inhibited (Martí et al., 2012). The different inhibitory effect may be involved in the regulation of ROS generation and energy metabolism in the organelle collaborating in the stress response. In fact, the reported effect of NO on different enzymes components of the antioxidant system as the lack of inhibition on Mn-SOD or on the majority of the ASC-GSH cycle except APX in pea plants could be part of a NO redox signalling through the H2O2 and NO cross-talk (Martí et al., 2012). However, similar to AOX, COX has not been found to be S-nitrosated in plants according to the references checked for this review, although it has been reported to be S-nitrosated in animals (Figure 3) (Zhang et al., 2005).
Under both, normal and stress conditions, PRXs from animals and plants have been shown to be targets of S-nitrosylation (Figure 3). In Arabidopsis and citrus plants under normal and salinity conditions, respectively, PRX is one of the proteins reported to be S-nitrosated (Tanou et al., 2009; Lindermayr et al., 2015). Deeper studies have also shown that S-nitrosation of plants PRXs leads to a change in their protein activity. In this sense, in chloroplasts, both the hydroperoxide-reducing peroxidase activity during the plant hypersensitive disease resistance response and the ONOO− detoxification activity of PRXIIE are inhibited by S-nitrosation (Romero-Puertas et al., 2007; Romero-Puertas et al., 2008). Recently, mitochondrial PRXIIF has been found by our lab to be S-nitrosated in vivo under long saline stress conditions, a situation in which NO levels were also increased (Camejo et al., 2013). The effect of NO and S-nitrosation on the protein function has been further investigated in our lab (Camejo et al., 2015). Under in vitro conditions, we found that both catalytic cysteines of PsPRXIIF (C59 and/or C84) are susceptible of S-nitrosation depending of its oligomerization state. We also found that the S-nitrosation leads to a conformational change that inhibits the PsPRXII peroxidase activity in favour of the transnitrosylase activity. These data together with the previous finding of PRXIIF being nitrosated only under long saline conditions and the reversibility of S-nitrosation and, consequently, of its peroxidase activity, suggest that the S-nitrosation of PRXIIF might act as a mechanism that is activated under oxidative and nitrosative stress, a situation that is presented in long saline conditions and that is reversed by reducing conditions, in which the TRX system may function in the mitochondria.
The level of S-nitrosated proteins depends on NO levels in the cells, which are regulated by several mechanisms. They depend on GSNO which is regulated by GSNOR activity (Liu et al., 2001; Holzmeister et al., 2011) and on Tyr nitration of proteins which consumes NO. Another important mechanism affecting S-nitrosation state of proteins may be through TRX activity, which similarly to GSNOR, can degrade S-nitrosothiols to increase their turnover ratio (Benhar et al., 2008). Several S-nitrosated enzymes that are summarized in this review have been also shown to be targets of mitochondrial TRX (see review in Møller et al., 2020). From plant S-nitrosation studies under abiotic stresses (Fares et al., 2011; Camejo et al., 2013; Puyaubert et al., 2014), it could be suggested that under those conditions, the effect of the stress does not produce large changes on the S-nitrosation status of the cells, probably due to other mechanisms that could be competing with the S-nitrosation process. Therefore, in the future, it might be more important to answer which cysteine residues in a specific protein are differentially S-nitrosated under normal and stress conditions and in which proportion, and also, which are the biochemical mechanisms involved.
Targets of Persulfidation
The chemical reactivity of H2S provokes the modification of cysteine residues to form persulfides, a PTM that may cause functional changes in protein structures and activities.This modification has been reported to increase under oxidative stress and it has been postulated as a defense mechanism against protein oxidative damage, avoiding the formation of dangerous -SO2H or irreversible -SO3H. Trying to identify the target proteins of persulfidation, a few works have been carried out in plant extracts. In a first approach, using a modified BST with S-methyl-methanothiosulfonate (MMTS) to block free thiols and the thiol-specific biotinylating agent biotin-HPDP, 106 proteins were identified as persulfidated in Arabidopsis mainly involved in photosynthesis, protein synthesis, and cell organization (Aroca et al., 2015). More recently, using methylsulfonylbenzothiazole (MSBT) to block both thiols and persulfide groups and cyanoacetate-based reagent CN-biotin as labelling agent, more than 3,000 proteins were identified as possible targets in cytosol-enriched leaf extracts of Arabidopsis plants grown under physiological conditions (Aroca et al., 2017). The bioinformatic analysis revealed that persulfidated cysteines are involved regulating important processes such as plant growth, plant response to abiotic and biotic stresses, carbon metabolism, and RNA translation. Several mitochondrial proteins were described as possible targets of persulfidation, including among others, proteins involved in ATP synthesis, respiration, chaperone function, protein synthesis, and all the TCA and photorespiratory enzymes (Figure 3). Interestingly, TRXo1 and almost all the described PsTRXo1 targets (Martí et al., 2009) were also found as persulfidated (ATP synthase subunit alpha, thiosulfate/3-mercaptopyruvate sulfurtransferase, elongation factor Tu and PRXIIF), implying that the role of this mitochondrial TRXo1 is related to persulfidation as it has been described for other TRXs (Filipovic and Jovanović, 2017) (Figure 3). Some of them were also demonstrated as S-nitrosylated and/or glutathionylated as described above, that reinforces the role of these PTMs in the processes in which the proteins are involved in mitochondria.
Another mechanism of action of H2S independent of its persulfidation effect is its coordination with the metal center of metalloproteins, attaching covalently to heme porphyrins. In this way, H2S acts as a potent inhibitor of plant and animal COX. Additionally, both COX and NADH dehydrogenase have been found among the persulfidated proteins in Arabidopsis (Aroca et al., 2017) (Figure 3).
A beneficial effect of exogenous application of H2S on plants subjected to different abiotic stresses has been described (see review by Corpas and Palma, 2020), including heavy metals, arsenic, low and high temperature, salinity, or drought. In fact, a reduction of oxidative stress on lipids and proteins as well as an increase in antioxidant components have been observed in the treated plants, contributing to a better response under the stress situation. However, to our knowledge, the effect on specific mitochondrial targets of persulfidation is scarcely reported.
Cross Talk Among H2S and Reactive Oxygen and Nitrogen Species
The crosstalk between H2S and RNS/ROS is important for the biological functions of all these molecules (Scuffi et al., 2014; Corpas et al., 2019b). H2S can act as a reductant reacting with biological oxidants, such as NO·, H2O2, O2·−, peroxynitrite, hypochlorite, and S-nitrosothiols although the direct reactions have not been quantified in plants (Gotor et al., 2019). Several examples point to the existence of the mentioned crosstalk. It has been described that H2S enhanced antioxidant capacity and salt tolerance of cucumber hypocotyls and radicles (Yu et al., 2013). The overlap between persulfidation and sulfenylation has been described by proteomic studies [reviewed by Zhou (2020)], with 437 proteins being modified by both. However, the effect of each modification can be opposite. As examples, activities of glyceraldehyde- 3-phosphate dehydrogenase (GAPC) and cytosolic APX are inhibited by sulfenylation and increased by persulfidation (Kitajima et al., 2008; Zaffagnini et al., 2013; Aroca et al., 2015), pointing to a fine regulation of the proteins by different PTMs. Related to RNS, an increase in NO occurs after H2S treatments of nitrate-treated tomato (Guo et al., 2018). Also, among 927 S-nitrosated proteins in Arabidopsis, 639 may be persulfidated (Hu et al., 2015; Aroca et al., 2018) with different functions on specific targets: both PTMs increased cAPX activity while S-nitrosation inhibited GAPC and persulfidation increased it (Begara-Morales et al., 2014; Lindermayr et al., 2015; Yang et al., 2015). As another example, exogenous treatments of H2S and NO alleviated some abiotic stresses and maintained the quality of post-harvested fruits (Ziogas et al., 2015; Corpas and Palma, 2018). Although the cascade of action is not fully elucidated, some experiments have reported that H2S could act upstream of NO signaling during stomatal closure while downstream in response to abiotic stress (Scuffi et al., 2014).
All these examples evidence that the interaction among H2S, RNS, and ROS may occur to regulate physiological processes not only under control conditions during plant development but also under stressed environments, allowing signal transduction pathways through the posttranslational regulation of key proteins involved in the response. The identification of the target proteins of each modification driven by H2S, RNS, and ROS and their effects on enzyme structure and activity are essential to understand the complexity of interactions and will help to reveal the modulating effect of redox components.
The TRX-PRX System Under Stress Conditions
Stress conditions provoke changes in cellular redox homeostasis mainly by dangerous increase in ROS/RNS generation (Noctor et al., 2015; Calderón et al., 2018a). APX, a component of the ASC-GSH cycle together with monodehydroascorbate reductase (MDHAR), dehydroascorbate (DHA) reductase (DHAR) and glutathione reductase (GR), is the responsible of H2O2 scavenging in mitochondria (Jiménez et al., 1997; Locato et al., 2018). Also, GPX, GRX, and TRX/PRX system play an important role in cellular ROS homeostasis. In this context, either mitochondria as chloroplasts are essential in maintaining the cellular redox balance under stress conditions (Cejudo et al., 2014; Sevilla et al., 2015b; Ojeda et al., 2017).
Changes in redox state and its significance in cellular signaling process can be evaluated in plant tissues using different approaches. Mutants are useful tools for investigating the involvement of specific proteins in vivo on redox metabolism in plant cells. Also, a parallel approach is to analyze stress conditions affecting the intracellular redox balance. In this sense, loss of function trxo1-1 and trxo1-2 mutants have been used not only to corroborate a role for the TRX system in regulating different metabolic processes in mitochondria but also to define the importance of mitochondrial TRXs under stress conditions (Daloso et al., 2015; Calderón et al., 2018a). However, in these studies, no extreme phenotype has been described, either in standard or stress conditions, possibly due to the redundancy or overlapping functions with mitochondrial or cytosolic proteins as other TRXs and GRXs (Daloso et al., 2015; Geigenberger et al., 2017; Ortiz-Espín et al., 2017; Calderón et al., 2018a; Calderón et al., 2018b; Sánchez-Guerrero et al., 2019). The information related to the physiological mitochondrial TRXo1/TRXh2 function in plant stress acclimation is scarce. In silico studies on AtTRXo1 gene expression have revealed that it does not vary as a response to different environment conditions including salinity. Contrary, in pea plants, an adaptative response to a short-term high salinity levels (150 mM NaCl), including increased PsTRXo1 gene expression was demonstrated, while longer salinity treatment downregulated it, with a parallel increase in PsTRXo1 protein and activity. Interestingly, these TRXo1 related changes were correlated with an increase in the AOX capacity in purified mitochondria preparations as well as a higher demand to regenerate the oxidized form of PRXIIF involved in ROS detoxification. Also, a maintained MnSOD gene expression, protein and activity levels were observed. Overall, the results demonstrated the participation of PsTRXo1 as a component in the mitochondrial antioxidant response allowing plant salinity adaptation (Martí et al., 2011). In order to gain further insight into the physiological and metabolic function of the mitochondrial TRXo1 in the plant acclimation to saline stress, we have recently used two independent mitochondrial Attrxo1 mutants (Attrxo1-1, Attrxo1-2) (Calderón et al., 2018b). Their responses to salinity fit well with those reported by Martí et al. (2011) in pea plants, all pointing that TRXo1 is required for the proper functioning of the antioxidant metabolism including compensatory mechanisms, as higher levels of H2O2 and NO, an upregulation of all SOD isoforms, catalase and GR, alterations in the glutathione redox state, due to maintained GSH content in Attrxo1 plants and changes in stomatal density, stomatal closure with lower water loss, which may be also a key factor for the adaptative response to salinity (Calderón et al., 2018b) (Figure 4). At mitochondrial level, a more recent study has revealed the impact of the lack of TRXo1 in the acclimation to salinity, with an important in vivo reorganization of the respiratory and antioxidant enzymes as well as metabolic responses (Sánchez-Guerrero et al., 2019). Compelling evidences from several plants indicated that AOX transcript and protein increase during salinity (Smith et al., 2009; Martí et al., 2011; Lázaro et al., 2013; Del-Saz et al., 2016). Using Attrxo1-1 and Attrxo1-2 mutants under a long-term saline stress, we described that AOX protein displayed a reduction of its in vivo activity in all genotypes and that exits a higher electron partitioning to the AOX pathway under salinity which denotes a relatively higher response of the AOX that can act preventing the generation of O2.-at the UQ level (Purvis, 1997; Cvetkovska and Vanlerberghe, 2012). In parallel, the high constitutive mitochondrial GR and DHAR activities in Atttrxo1 mutants, could act helping to decrease H2O2 content and to increase GSH recycling. Importantly, we observed a change in AOX isoforms pattern but AOX protein was not decreased and was also maintained in its reduced state under control and saline conditions in both Attrxo1 mutants. These observations suggested that in vivo, the mitochondrial TRXo1 system could perform a maintenance of reductive function rather than an AOX regulation, as it has been proposed. This possibility was also suggested recently by Nietzel et al. (2017) (Figure 4). Furthermore, a pronounced decrease on glucose and fructose levels occurred in both Attrxo1 mutants under control and salinity, coinciding with an increased in vivo respiration through the cytochrome c (COX) pathway (Sánchez-Guerrero et al., 2019). These results indicated a reorganization in central carbon metabolism and reflect a higher use of these sugars in the glycolytic pathway, causing an increased respiration, probably driven by an increased carbon flow through the TCA cycle as previously suggested (Daloso et al., 2015). The increase in ATP-coupled respiration is an indicator of a higher demand on the leaf energy of the mutants under control conditions (Figure 3).
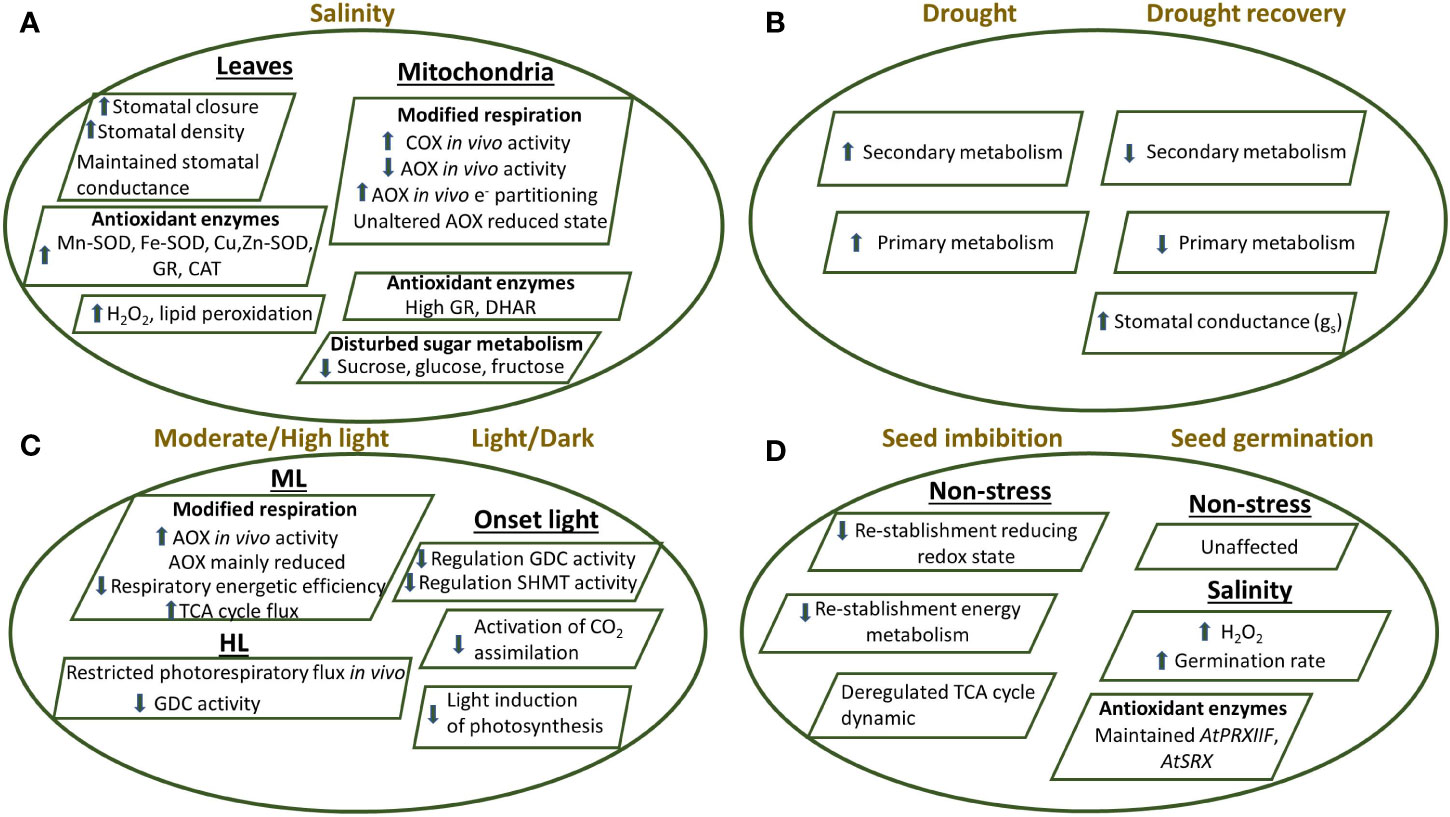
Figure 4 Proposed model showing the response of Arabidopsis thaliana plants knockout of the mitochondrial TRXo1 under different stress conditions as (A) salinity in leaves and mitochondria, (B) drought and drought recovery, (C) moderate light/high light and (D) during seed imbibition and germination, as detailed in the main text (Daloso et al., 2015; Ortiz-Espín et al., 2017; Calderón et al., 2018b; Florez-Sarasa et al., 2019; Sánchez-Guerrero et al., 2019; Da Fonseca-Pereira et al., 2019b; Da Fonseca-Pereira et al., 2019c). Effect on stomatal behaviour, antioxidant and oxidative metabolism, respiration and photorespiration, photosynthesis, TCA cycle as well as primary and secondary metabolism are shown in the KO plants compared to non-transformed plants. AOX, alternative oxidase; CAT, catalase; COX, cytochrome oxidase; Cu/Zn-SOD, copper-zinc superoxide dismutase; DHAR, dehydroascorbate reductase; GDC, glycine decarboxylase; Fe-SOD, iron superoxide dismutase; GR, glutathione reductase; Mn-SOD, manganese superoxide dismutase; PRXIIF, peroxiredoxin IIF; SHMT, serine hydroxymethyl transferase; SRX, sulfiredoxin; TCA, tricarboxylic acid.
A similar metabolic adjustment could be also taking place during seed germination, a process in which a function for AtTRXo1 has also been described (Ortiz-Espín et al., 2017). Arabidopsis AtTrxo1 transcript levels were particularly high in dry seeds and cotyledons, where it reached a maximum coinciding with 50% germination. However, expression was lower in seeds germinating under salinity. We reported that seeds of both Attrxo1-1 and Attrxo1-2 mutant lines failed to show any important difference in the germination rate compared to WT, but the physiological and metabolic analysis of these mutants showed diverse and complex responses, showing higher H2O2 levels in dry seeds. Moreover, Attrxo1 mutant seeds germinated faster and accumulated higher H2O2 content under salinity (Figure 4). This peak of H2O2 at the beginning of germination might be a factor in the observed early germination rate and fits well with that proposed for the accumulated mitochondrial H2O2 at early stages of germination (Zhang et al., 2014). Thus, a role for AtTRXo1 in redox homeostasis during seed germination, acting as a possible sensor of saline stress and/or an inducer of H2O2 accumulation was proposed. This specific role of TRXo1 under stress may in turn be related to its targets following seed germination, an aspect that deserves further investigations (Ortiz-Espín et al., 2017).
Recently, the earliest events in energy and redox metabolism of Arabidopsis seeds at imbibition, as a physiological step of metabolic reactivation, have been investigated (Nietzel et al., 2020). RoGFP-based in vivo sensing experiments, have suggested that the reestablishment to a more reducing thiol redox status of the mitochondrial matrix and the cytosol within minutes in intact seeds, is intimately linked to the reestablishment of energy metabolism. Redox proteomic analysis has shown that active site cysteine peptides of GR 2, NTR a/b, and TRXo1 present the strongest change in redox state. Seeds germination of the three mutants gr2, ntra/b, and trx-o1 was associated with increased respiratory rates and deregulated TCA cycle dynamics, suggesting decreased resource efficiency of energy metabolism. NAD-malic enzyme (NAD-ME) and aconitase (ACO) also showed a quantitative redox-dependent response, but this was not the case for enzymes as 2-oxoglutarate dehydrogenase (OGDH), NAD-isocitrate dehydrogenase (ICDH), and ICDH which only showed a marginal response at imbibition. These differences indicated that the functional impact of cysteine redox switch operation is enzyme-specific. An important contribution of redox regulation to efficient metabolism during early seed germination was proposed.
The importance of the mitochondrial NTR/TRX system under drought episodes was recently investigated using both Attrxo1 mutant and Atntra ntrb double mutant plants (Da Fonseca-Pereira et al., 2019b). Under these conditions, all the genotypes lacking functional NTR/TRX system, showed enhanced drought tolerance. Interestingly, TRXo1 transcript was more highly expressed under drought, an effect even stronger mainly during repetitive drought/recovery events. Results on the changes in secondary metabolites with a large number of metabolites increasing in at least one of the Attrxo1 mutants following two cycles of drought, reinforces the idea that secondary metabolism is redox regulated by TRX system (Figure 4). Finally, the analysis of the increased stomatal conductance following drought recovery suggested a TRXo1 redox regulation of stoma function (Figure 4) which fits well with the previously reported involvement of TRXo1 mediated redox regulation of stomatal dynamic and function under salinity (Calderón et al., 2018b).
Regarding the effects of different light conditions, it has been observed that an altered in vivo AOX activity and carbon metabolism occur in both Arabidopsis trxo1 mutants under medium light (ML) and high light (HL) conditions (Florez-Sarasa et al., 2019). Contrary to the effects under salinity (Sánchez-Guerrero et al., 2019), the results showed that the in vivo AOX activity was higher in the Attrxo1 mutants at ML while the AOX redox state was apparently unaltered as we have commented above. Moreover, the authors claim that the negative regulation of the TCA cycle by the TRX system is coordinated with the increased input of electrons into the AOX pathway. Under HL conditions, AOX and photosynthesis displayed similar patterns in the mutants. Furthermore, changes on photorespiration were observed under HL conditions, being restricted at the level of glycine decarboxylation, most likely as a consequence of the redox imbalance. These results denote the relevance of TRXo1 on the interaction between mitochondrial redox and carbon metabolism under light stress conditions (Figure 4). In a parallel study, the alteration of photorespiratory activity in the absence of TRXo1 was described. In this sense, it has been shown that a functional TRXo1 allowing the rapid induction of mitochondrial steps of the photorespiration process via GDC system in conjunction with SHMT is necessary to facilitate light-induction of photosynthesis (Reinholdt et al., 2019b).
Some other TRXo1 functional studies were focused on determining the effect of high PsTRXo1 expression on processes linked to oxidative treatment and on the functional PsTRXo1 role in the nucleus. Under a situation of high H2O2 level treatment, the analysis of different hallmarks of programmed cell death (PCD) demonstrated that the over-expression of PsTrxo1 in tobacco (Nicotiana tabacum) BY-2 cells, was able to increase the cell viability in that oxidative situation, contrasting with a severe decrease in viability and marked oxidative stress, with a rapid cell death, observed in non-overexpressing lines. The decreased content in endogenous H2O2, an increased catalase activity, with the practically maintained PRXII expression, would be involved in the delayed cell death found in over-expressing cells, in which changes in oxidative parameters and GSH/ASC redox state were less extended after the H2O2 treatment, than in non-overexpressing lines. These data pointed to PsTRXo1 as an important factor responsible for the delay in the PCD provoked by the oxidative treatment (Ortiz-Espín et al., 2015) (Figure 5). In pea leaves TRXo1 is constitutively found in mitochondria and nucleus (Martí et al., 2009). Nuclear localization has been reported for several cytosolic TRXh isoforms but only during oxidative stress (Pulido et al., 2009). While subsequent reports describing a functional TRX system in the plant nucleus have appeared, little evidence on the nuclear TRXs targets in vivo has been reported and only a few candidate protein targets have been experimentally validated (Delorme-Hinoux et al., 2016). Among them, proliferating cellular nuclear antigen (PCNA) has been demonstrated to interact with PsTRXo1 in the nucleus (Calderón et al., 2017a). In Medicago and barley seed embryos, PCNA was also reported as a candidate targets of TRXh, but the interaction was not conclusively demonstrated (Alkhalfioui et al., 2007; Hägglund et al., 2008). In TBY-2 cells it was reported that the over-expressed PsTRXo1 was localized in mitochondria and nucleus and the over-expression correlated with changes in the growth of the culture, increase in the rate of cell proliferation and a decrease in the total cellular GSH content, but maintained nuclear GSH accumulation (Calderón et al., 2017a). Changes in GSH content were accompanied by a higher mitotic index, unlike non-expressing TBY-2 cells. All these findings suggest that TRXo1 is involved in the cell cycle progression of TBY-2 cultures, possibly through its link with both GSH and PCNA (Figure 5).
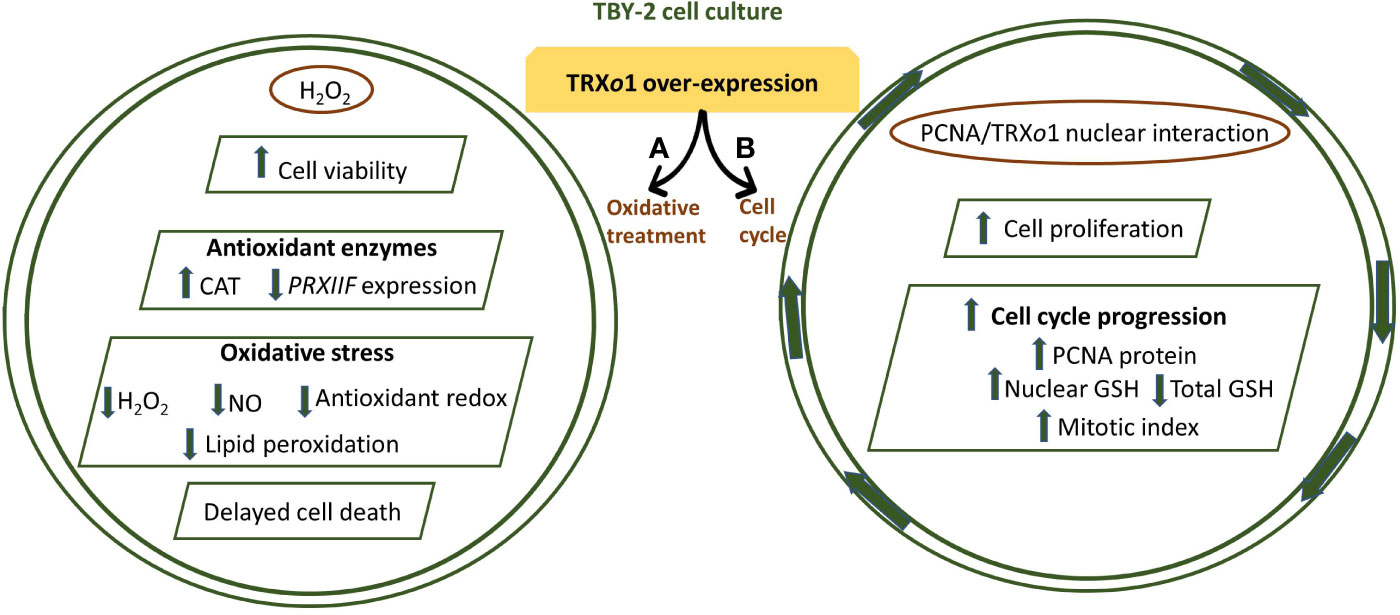
Figure 5 (A) PsTRXo1 function in cell response to high H2O2 treatment. TBY-2 cell lines over-expressing PsTRXo1 showed higher and maintained viability than control lines. The high catalase (CAT) activity, decrease in ASC and GSH redox state are parallel and might contribute to decreased H2O2, NO, and lipid peroxidation, protecting together with TRXo1, against increased oxidative stress, thus allowing delayed programmed cell death (PCD) (Ortiz-Espín et al., 2015). (B) Proliferating cell nuclear antigen (PCNA) interacts with PsTRXo1 in the nucleus and showed an efficient in vitro disulfide reductase activity reducing oxidized PCNA. Flow cytometry showed that TRXo1 increased the rate of cell proliferation and cell cycle progression. Higher amount of PCNA protein in over-expressing TRXo1 cells occurred in the growth phases when high mitotic index and percentage of cells in S-G2/M phases were more evident and different to that in control lines. These changes were parallel to maintained GSH level in the nucleus but decreased in total cellular GSH in over-expressing lines, when the percentage of cells in G2/M phase was higher. Overall, these changes suggest that TRXo1 is involved in cell cycle progression providing a reductive environment and interacting with PCNA (Calderón et al., 2017a).
The information on the involvement of the mitochondrial PRXIIF in stress is scarce, being the chloroplastic and cytosolic isoforms more studied. Among ROS sensors during stress, reactive thiols may play a key role in the signal transduction, and as example, the action of SRX on the oxidized PRX has been reported (Lázaro et al., 2013). A link between redox changes and gene regulation has been described during the stress response (Astier et al., 2011) being AtPRXIIF one of the responsive genes in oxidative-induced stress situations such as treatments of Arabidopsis cell cultures with H2O2, menadione or antimycin A (Sweetlove et al., 2002) or in pea and poplar leaves under cold stress (Barranco-Medina et al., 2007; Gama et al., 2007). Also under salinity and Cd stress in pea leaves but not in roots, both transcripts and protein levels were upregulated (Barranco-Medina et al., 2007), while in more resistant species as Arabidopsis, the leaf mRNA level did not change significantly under salt, ozone, H2O2, or light stress (Horling et al., 2002; Horling et al., 2003). In another study, PRXIIF presented a biphasic response in pea plants under salinity, being the gene upregulated after 5 days of 150 mM NaCl treatment while downregulated after 14 days (Martí et al., 2011). However, the protein content remained constant although a regulation by S-nitrosylation was described affecting structure and function as commented above (Camejo et al., 2013; Camejo et al., 2015). Also, under these conditions salinity provoked a change in the oligomerization pattern of PRXIIF, increasing the oligomeric forms of the protein, similar to that occurring when recombinant protein was treated with H2O2 (Camejo et al., 2015). Also, the analysis of mutants lacking AtPRXIIF has allowed the knowledge of the physiological role of the protein not only under control (where a compensatory mechanism by mitochondrial antioxidant system seems to occur) but also under stress situations (Lázaro et al., 2013). The mutants’ behaviour has revealed a role for PRXIIF in root growth under Cd stress in Arabidopsis seedlings (Finkemeier et al., 2005), water stress in Vitis vinifera and in Arabidopsis mature leaves (Gama et al., 2007; Vidigal et al., 2013). All these examples imply the important involvement of PRXIIF in the antioxidant defence and redox signalling in plant cells.
Conclusion and Perspectives
In the last years, the advance in the analysis and characterization of TRXo1 mutants has allowed to obtain wide information on the function of this protein, unveiling the key processes by which TRX system regulates mitochondrial respiration, TCA cycle, photorespiration, plant acclimation to stress through a proper functioning of the antioxidant metabolism, as well as its link with the germination process and cell cycle progression. In general, in the scarce studies describing processes as germination and plants acclimation under different conditions, the lack of TRXo1 generates a decrease in the energetic efficiency, but the connection between TRXo1 and in vivo respiratory AOX, needs to be explored in more detail. Also, the compelling evidences that both, TRXh2 and o1 systems play a role in the redox regulation of the mitochondrial photorespiratory metabolism needs to be further investigated. Another interesting point is the possible existence of compensatory and redundant mechanisms as the existence of other TRX isoforms. The use of multiple mutants in different redox systems present in the cellular compartments will aid to unravel their functionality in which, the PTMs may play a key role. Understanding which is the signature of the signal that triggers the specific PTM associated to a specific stress and also how plants distinguish these signals, will help to get insight on how plants responds to the different environmental cues. Hopefully, the development of new mass-spectrometry proteomic techniques will allow in the near future to expand the knowledge on the dynamic and interaction among the different PTMs. This is important not only for fundamental research but also for crop improvement in the actual climate-changing environment.
Author Contributions
MM, AJ, and FS conceived the idea and wrote the manuscript.
Funding
The review was funded by a Saavedra Fajardo 20402/SF/17 (Fundación Séneca, Murcia, Spain) and a Ramón y Cajal (Ministry of Science and Innovation, Spain) Fellowship both awarded to MM and Spanish grants MINECO/FEDER (BFU2017-86585-P) and Seneca Excellence Project (19876/GERM/15).
Conflict of Interest
The authors declare that this review article was written in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors apologize to the scientists that are not cited because of space limitation.
References
Albert, N. A., Sivanesan, H., Vanlerberghe, G. C. (2017). The occurrence and control of nitric oxide generation by the plant mitochondrial electron transport chain. Plant Cell Environ. 40, 1074–1085. doi: 10.1111/pce.12884
Alkhalfioui, F., Renard, M., Frendo, P., Keichinger, C., Meyer, Y., Gelhaye, E., et al. (2008). A Novel Type of Thioredoxin Dedicated to Symbiosis in Legumes. Plant Physiol. 148, 424–435. doi: 10.1104/pp.108.123778
Alkhalfioui, F., Renard, M., Vensel, W. H., Wong, J., Tanaka, C. K., Hurkman, W. J., et al. (2007). Thioredoxin-linked proteins are reduced during germination of Medicago truncatula seeds. Plant Physiol. 144, 1559–1579. doi: 10.1104/pp.107.098103
Álvarez, C., García, I., Romero, L. C., Gotor, C. (2012). Mitochondrial sulfide detoxification requires a functional isoform O-acetylserine (thiol) lyase C in Arabidopsis thaliana. Mol. Plant 5, 1217–1226. doi: 10.1093/mp/sss043
Aroca, A., Serna, A., Gotor, C., Romero, L. C. (2015). S-sulfhydration: a cysteine posttranslational modification in plant systems. Plant Physiol. 168, 334–342. doi: 10.1104/pp.15.00009
Aroca, A., Benito, J. M., Gotor, C., Romero, L. C. (2017). Persulfidation proteome reveals the regulation of protein function by hydrogen sulfide in diverse biological processes in Arabidopsis. J. Exp. Bot. 68, 4915–4927. doi: 10.1093/jxb/erx294
Aroca, A., Gotor, C., Romero, L. C. (2018). Hydrogen Sulfide Signaling in Plants: Emerging Roles of Protein Persulfidation. Front. Plant Sci. 9, 1369. doi: 10.3389/fpls.2018.01369
Astier, J., Rasul, S., Koen, E., Manzoor, H., Besson-Bard, A., Lamotte, O., et al. (2011). S-nitrosylation: an emerging post-translational protein modification in plants. Plant Sci. 181, 527–533. doi: 10.1016/j.plantsci.2011.02.011
Astier, J., Gross, I., Durner, J. (2018). Nitric oxide production in plants: an update. J. Exp. Bot. 69, 3401–3411. doi: 10.1093/jxb/erx420
Balmer, Y., Vensel, W. H., Tanaka, C. K., Hurkman, W. J., Gelhaye, E., Rouhier, N., et al. (2004). Thioredoxin links redox to the regulation of fundamental processes of plant mitochondria. Proc. Natl. Acad. Sci. U.S.A. 101, 2642–2647. doi: 10.1073/pnas.0308583101
Barranco-Medina, S., Krell, T., Finkemeier, I., Sevilla, F., Lázaro, J. J., Dietz, K. J. (2007). Biochemical and molecular characterization of the mitochondrial peroxiredoxin PsPrxII F from Pisum sativum. Plant Physiol. Biochem. 45, 729–739. doi: 10.1016/j.plaphy.2007.07.017
Barranco-Medina, S., Bernier-Villamor, L., Krell, T., Sevilla, F., Lázaro, J. J., Dietz, K. J. (2008). Hexameric oligomerization of mitochondrial peroxiredoxin PrxII F and formation of an ultrahigh affinity complex with its electron donor thioredoxin Trx-o. J. Exp. Bot. 59, 3259–3269. doi: 10.1093/jxb/ern177
Barranco-Medina, S., Lázaro, J. J., Dietz, K. J. (2009). The oligomeric conformation of peroxiredoxins links redox state to function. FEBS Lett. 583, 1809–1816. doi: 10.1016/j.febslet.2009.05.029
Bauwe, H., Kolukisaoglu, U. (2003). Genetic manipulation of glycine decarboxylation. J. Exp. Bot. 54, 1523–1535. doi: 10.1093/jxb/erg171
Bedhomme, M., Adamo, M., Marchand, C. H., Couturier, J., Rouhier, N., Lemaire, S. D., et al. (2012). Glutathionylation of cytosolic glyceraldehyde-3-phosphate dehydrogenase from the model plant Arabidopsis thaliana is reversed by both glutaredoxins and thioredoxins in vitro. Biochem. J. 445, 337–347. doi: 10.1042/BJ20120505
Beer, S. M., Taylor, E. R., Brown, S. E., Dahm, C. C., Costa, N. J., Runswick, M. J., et al. (2004). Glutaredoxin 2 catalyzes the reversible oxidation and glutathionylation of mitochondrial membrane thiol proteins: implications for mitochondrial redox regulation and antioxidant defense. J. Biol. Chem. 279, 47939–47951. doi: 10.1074/jbc.M408011200
Begara-Morales, J. C., Sánchez-Calvo, B., Chaki, M., Valderrama, R., Mata-Pérez, C., López-Jaramillo, J., et al. (2014). Dual regulation of cytosolic ascorbate peroxidase (APX) by tyrosine nitration and S-nitrosylation. J. Exp. Bot. 65, 527–538. doi: 10.1093/jxb/ert396
Benhar, M., Forrester, M. T., Hess, D. T., Stamler, J. S. (2008). Regulated protein denitrosylation by cytosolic and mitochondrial thioredoxins. Science 320, 1050–1054. doi: 10.1126/science.1158265
Braymer, J. J., Lill, R. (2017). Iron–sulfur cluster biogenesis and trafficking in mitochondria. J. Biol. Chem. 292, 12754–12763. doi: 10.1074/jbc.R117.787101
Brennan, J. P., Miller, J. I., Fuller, W., Wait, R., Begum, S., Dunn, M. J., et al. (2006). The utility of N,N-biotinyl glutathione disulfide in the study of protein S-glutathiolation. Mol. Cell Proteom. 5, 215–225. doi: 10.1074/mcp.M500212-MCP200
Bykova, N. V., Igamberdiev, A. U. (2016). “Redox state in plant mitochondria and its role in stress tolerance”, in Redox State as a Central Regulator of Plant-Cell Stress Responses. Ed. Gupta, D. K., Palma, J. M., Corpas, F. J. (Switzerland: Springer International Publishing), 93–115.
Calderón, A., Ortiz-Espín, A., Iglesias-Fernández, R., Carbonero, P., Pallardó, F. V., Sevilla, F., et al. (2017a). Thioredoxin (Trxo1) interacts with proliferating cell nuclear antigen (PCNA) and its overexpression affects the growth of tobacco cell culture. Redox Biol. 11, 688–700. doi: 10.1016/j.redox.2017.01.018
Calderón, A., Lázaro-Payo, A., Iglesias-Baena, I., Camejo, D., Lázaro, J. J., Sevilla, F., et al. (2017b). Glutathionylation of pea chloroplast 2-Cys Prx and mitochondrial PrxIIF affects their structure and peroxidase activity and sulfiredoxin deglutathionylates only the 2-Cys Prx. Front. Plant Sci. 8, 118. doi: 10.3389/fpls.2017.00118
Calderón, A., Sevilla, F., Jiménez, A. (2018a). “Redox protein thioredoxins: function under salinity, drought and extreme temperature condition,” in Antioxidants and Antioxidant Enzymes in Higher Plants. Ed. Gupta, D. K., Palma, J. M., Corpas, F. J. (Switzerland: Springer International Publishing), 132–162.
Calderón, A., Sánchez-Guerrero, A., Ortiz, A., Martínez-Alcalá, I., Camejo, D., Jiménez, A., et al. (2018b). Lack of mitochondrial thioredoxin o1 is compensated by antioxidant components under salinity in Arabidopsis thaliana plants. Physiol. Plant 164, 251–267. doi: 10.1111/ppl.12708
Camejo, D., Romero-Puertas, M. C., Rodríguez-Serrano, M., Sandalio, L. M., Lázaro, J. J., Jiménez, A., et al. (2013). Salinity-induced changes in S-nitrosylation of pea mitochondrial proteins. J. Proteom. 79, 87–99. doi: 10.1016/j.jprot.2012.12.003
Camejo, D., Ortiz-Espín, A., Lázaro, J. J., Romero-Puertas, M. C., Lázaro-Payo, A., Sevilla, F., et al. (2015). Functional and structural changes in plant mitochondrial PrxIIF caused by NO. J. Proteom. 119, 112–125. doi: 10.1016/j.jprot.2015.01.022
Casagrande, S., Bonetto, V., Fratelli, M., Gianazza, E., Eberini, I., Massignan, T., et al. (2002). Glutathionylation of human thioredoxin: a possible crosstalk between the glutathione and thioredoxin systems. Proc. Natl. Acad. Sci. U. S. A. 99, 9745–9749. doi: 10.1073/pnas.152168599
Cejudo, F. J., Meyer, A. J., Reichheld, J.-P., Rouhier, N., Traverso, J. A., Huber, S. C. (2014). Thiol-based redox homeostasis and signaling. Front. Plant Sci. 5, 266. doi: 10.3389/fpls.2014.00266
Chen, Y.-R., Chen, C. H.-L., Pfeiffer, D. R., Zweier, J. L. (2007). Mitochondrial complex II in the post-ischemic heart. Oxidative injury and the role of proteins-glutathionylation. J. Biol. Chem. 282, 32640–32654. doi: 10.1074/jbc.M702294200
Chew, O., Whelan, J., Millar, A. H. (2003). Molecular definition of the ascorbate-glutathione cycle in Arabidopsis mitochondria reveals dual targeting of antioxidant defenses in plants. J. Biol. Chem. 278, 46869–46877. doi: 10.1074/jbc.M307525200
Ciofi-Baffoni, S., Nasta, V., Banci, L. (2018). Protein networks in the maturation of human iron–sulfur proteins. Metallomics 10, 4972. doi: 10.1039/C7MT00269F
Corpas, F. J., Palma, J. M. (2018). Nitric oxide on/off in fruit ripening. Plant Biol. 20, 805–807. doi: 10.1111/plb.12852
Corpas, F. J., Palma, J. M. (2020). H2S signaling in plants and applications in agriculture. J. Adv. Res. 24, 131–137. doi: 10.1016/j.jare.2020.03.011
Corpas, F. J., Alché, J. D., Barroso, J. B. (2013). Current overview of S-nitrosoglutathione (GSNO) in higher plants. Front. Plant Sci. 4, 126. doi: 10.3389/fpls.2013.00126
Corpas, F. J., Río, L. A. D., Palma, J. M. (2019a). Impact of Nitric Oxide (NO) on the ROS Metabolism of Peroxisomes. Plants (Basel) 8, e37. doi: 10.3390/plants8020037
Corpas, F. J., González-Gordo, S., Cañas, A., Palma, J. M. (2019b). Nitric oxide and hydrogen sulfide in plants: which comes first? J. Exp. Bot. 70, 4391–4404. doi: 10.1093/jxb/erz031
Cvetkovska, M., Vanlerberghe, G. C. (2012). Alternative oxidase impacts the plant response to biotic stress by influencing the mitochondrial generation of reactive oxygen species. Plant Cell Environ. 36, 721–732. doi: 10.1111/pce.12009
Da Fonseca-Pereira, P., Souza, P. V. L., Hou, L. Y., Schwab, S., Geigenberger, P., Nunes-Nesi, A., et al. (2019a). Thioredoxin h2 contributes to the redox regulation of mitocondrial photorespiratory metabolism. Plant Cell Environ. 43, 188–208. doi: 10.1111/pce.13640
Da Fonseca-Pereira, P., Daloso, D. M., Gago, J., Magnum, F., Silva, O., Condori-Apfata, J. A., et al. (2019b). The Mitochondrial Thioredoxin System Contributes to the Metabolic Responses Under Drought Episodes in Arabidopsis. Plant Cell Physiol. 60, 213–229. doi: 10.1093/pcp/pcy194
Da Fonseca-Pereira, P., Daloso, D. M., Gago, J., Nunes-Nesi, A., Araújo, W. L. (2019c). On the role of the plant mitochondrial thioredoxin system during abiotic stress. Plant Signal. Behav. 14, 1592536. doi: 10.1080/15592324.2019.1592536
Daloso, D. M., Müller, K., Obata, T., Florian, A., Tohge, T., Bottcher, A., et al. (2015). Thioredoxin, a master regulator of the tricarboxylic acid cycle in plant mitochondria. Proc. Natl. Acad. Sci. U.S.A. 112, 1392–1400. doi: 10.1073/pnas.1424840112
Day, D. A., Millar, A. H., Wiskich, J. T., Whelan, J. (1994). Regulation of Alternative Oxidase Activity by Pyruvate in Soybean Mitochondria. Plant Physiol. 106, 1421–1427. doi: 10.1104/pp.106.4.1421
Delorme-Hinoux, V., Bangash, S. A., Meyer, A. J., Reichheld, J. P. (2016). Nuclear thiol redox systems in plants. Plant Sci. 243, 84–95. doi: 10.1016/j.plantsci.2015.12.002
Del-Saz, N. F., Florez-Sarasa, I., Clemente-Moreno, M. J., Mhadhbi, H., Flexas, J., Fernie, A. R., et al. (2016). Salinity tolerance is related to cyanide-resistant alternative respiration in Medicago truncatula under sudden severe stress. Plant Cell Environ. 39, 2361–2369. doi: 10.1111/pce.12776
Del-Saz, N. F., Ribas-Carbo, M., McDonald, A. E., Lambers, H., Fernie, A. R., Florez-Sarasa, I. (2017). An In Vivo Perspective of the Role(s) of the Alternative Oxidase Pathway. Trends Plant Sci. 23, 206–219. doi: 10.1016/j.tplants.2017.11.006
Dietz, K.-J. (2011). Peroxiredoxins in plants and cyanobacteria. Antioxid. Redox Signal. 15, 1129–1159. doi: 10.1089/ars.2010.3657
Dixon, D. P., Skipsey, M., Grundy, N. M., Edwards, R. (2005). Stress-induced protein Sglutathionylation in Arabidopsis. Plant Physiol. 138, 2233–2244. doi: 10.1104/pp.104.058917
Drapier, J. C. (1997). Interplay between NO and [Fe-S] clusters: relevance to biological systems. Methods 11, 319–329. doi: 10.1006/meth.1996.0426
Dumont, S., Rivoal, J. (2019). Consequences of Oxidative Stress on Plant Glycolytic and Respiratory Metabolism. Front. Plant Sci. 10, 166. doi: 10.3389/fpls.2019.00166
Eisenhut, M., Roelland, M. S., Weber, A. P. M. (2019). Mechanistic understanding of photorespiration paves the way to a new green revolution. New Phytol. 223, 1762–1769. doi: 10.1111/nph.15872
Engel, N., van den Daele, K., Kolukisaoglu, U., Morgenthal, K., Weckwerth, W., Pärnik, T., et al. (2007). Deletion of glycine decar-boxylase in Arabidopsis is lethal under non photorespiratory conditions. Plant Physiol. 144, 1328–1335. doi: 10.1104/pp.107.099317
Fares, A., Rossignol, M., Peltier, J. B. (2011). Proteomics investigation of endogenous S-nitrosylation in Arabidopsis. Biochem. Biophys. Res. Commun. 416, 331–336. doi: 10.1016/j.bbrc.2011.11.036
Fenech, M., Amaya, I., Valpuesta, V., Botella, M. A. (2019). Vitamin C Content in Fruits: Biosynthesis and Regulation. Front. Plant Sci. 9:2006:2006. doi: 10.3389/fpls.2018.02006
Feng, J., Chen, L., Zuo, L. (2019). Protein S-Nitrosylation in plants: Current progresses and challenges. J. Integr. Plant Biol. 61, 1206–1223. doi: 10.1111/jipb.12780
Fernández-García, N., Martí, M. C., Jiménez, A., Sevilla, F., Olmos, E. (2009). Sub-cellular distribution of glutathione in an Arabidopsis mutant (vtc1) deficient in ascorbate. J. Plant Physiol. 166, 2004–2012. doi: 10.1016/j.jplph.2009.06.006
Filipovic, M. R., Jovanović, V. M. (2017). More than just an intermediate: hydrogen sulfide signalling in plants. J. Exp. Bot. 68, 4733–4736. doi: 10.1093/jxb/erx352
Finkemeier, I., Goodman, M., Lankemeyer, P., Kandlbinder, A., Sweetlove, L. J., Dietz, K. J. (2005). The mitochondrial type II peroxiredoxin F is essential for redox homeostasis and root growth of Arabidopsis thaliana under stress. J. Biol. Chem. 280, 12168–12180. doi: 10.1074/jbc.M413189200
Florez-Sarasa, I., Obata, T., Del-Saz, N. S. F. N., Reichheld, J. P., Meyer, E. H., Rodriguez-Concepcion, M., et al. (2019). The lack of mitochondrial thioredoxin TRXo1 affects in vivo alternative oxidase activity and carbon metabolism under different light conditions. Plant Cell Physiol. 60, 2369–2381. doi: 10.1093/pcp/pcz123
Foyer, C. H., Noctor, G. (2011). Ascorbate and Glutathione: The Heart of the Redox Hub. Plant Physiol. 155, 2–18. doi: 10.1104/pp.110.167569
Foyer, C. H., Noctor, G. (2013). Redox signaling in plants. Antioxid. Redox Signal. 18, 2087–2090. doi: 10.1089/ars.2013.5278
Foyer, C. H., Baker, A., Wright, M., Sparkes, I. A., Mhamdi, A., Schippers, J. H. M., et al. (2020). On the move: redox-dependent protein relocation in plants. J. Exp. Bot. 71, 620–631. doi: 10.1093/jxb/erz330
Fratelli, M., Demol, H., Puype, M., Casagrande, S., Villa, P., Eberini, I., et al. (2003). Identification of proteins undergoing glutathionylation in oxidatively stressed hepatocytes and hepatoma cells. Proteomics 3, 1154–1161. doi: 10.1002/pmic.200300436
Gama, O., Keech, F., Eymery, F., Finkemeier, I., Gelhaye, E., Gardeström, P., et al. (2007). The mitochondrial type II peroxiredoxin from poplar. Physiol. Plant 129, 196–206. doi: 10.1111/j.1399-3054.2006.00785.x
Gao, X. H., Bedhomme, M., Veyel, D., Zaffagnini, M., Lemaire, S. D. (2009). Methods for Analysis of Protein Glutathionylation and their Application to Photosynthetic Organisms. Mol. Plant 2, 218–235. doi: 10.1093/mp/ssn072
García, J., Han, D., Sancheti, H., Yap, L. P., Kaplowitz, N., Cadenas, E. (2010). Regulation of mitochondrial glutathione redox status and protein glutathionylation by respiratory substrates. J. Biol. Chem. 285, 39646–39654. doi: 10.1074/jbc.M110.164160
García, I., Gotor, C., Romero, L. C. (2015). “Cysteine homeostasis,” in Amino Acids in Higher Plants. Ed. D’Mello, J. P. F. (Wallingford, UK: CABI Publishing), 219–233.
Geigenberger, P., Thormählen, I., Daloso, D. M., Fernie, A. R. (2017). The unprecedented versatility of the plant thioredoxin system. Trends Plant Sci. 22, 249–262. doi: 10.1016/j.tplants.2016.12.008
Gelhaye, E., Rouhier, N., Gérard, J., Jolivet, Y., Gualberto, J., Navrot, N., et al. (2004). A specific form of thioredoxin h occurs in plant mitochondria and regulates the alternative oxidase. Proc. Natl. Acad. Sci. U.S.A. 101, 14545–14550. doi: 10.1073/pnas.0405282101
Gleason, C., Huang, S., Thatcher, L. F., Foley, R. C., Anderson, C. R., Carroll, A. J., et al. (2011). Mitochondrial complex II has a key role in mitochondrial-derived reactive oxygen species influence on plant stress gene regulation and defense. Proc. Natl. Acad. Sci. U.S.A. 108, 10768–10773. doi: 10.1073/pnas.1016060108
Gotor, C., García, I., Aroca, Á., Laureano-Marín, A. M., Arenas-Alfonseca, L., Jurado-Flores, A., et al. (2019). Signaling by hydrogen sulfide and cyanide through post-translational modification. J. Exp. Bot. 70, 4251–4265. doi: 10.1093/jxb/erz225
Guo, F. Q., Crawford, N. M. (2005). Arabidopsis nitric oxide synthase 1 is targeted to mitochondria and protects against oxidative damage and dark-induced senescence. Plant Cell 17, 3436–3450. doi: 10.1105/tpc.105.037770
Guo, Z., Liang, Y., Yan, J., Yang, E., Li, K., Xu, H. (2018). Physiological response and transcription profiling analysis reveals the role of H2S in alleviating excess nitrate stress tolerance in tomato roots. Plant Physiol. Biochem. 124, 59–69. doi: 10.1016/j.plaphy.2018.01.006
Gupta, K. J., Igamberdiev, A. U. (2016). Reactive nitrogen species in mitochondria and their implications in plant energy status and hypoxic stress tolerance. Front. Plant Sci. 7, 369. doi: 10.3389/fpls.2016.00369
Gupta, K. J., Shah, J. K., Brotman, Y., Jahnke, K., Willmitzer, L., Kaiser, W. M., et al. (2012). Inhibition of aconitase by nitric oxide leads to induction of the alternative oxidase and to a shift of metabolism towards biosynthesis of amino acids. J. Exp. Bot. 63, 1773–1784. doi: 10.1093/jxb/ers053
Hägglund, P., Bunkenborg, J., Maeda, K., Svensson, B. (2008). Identification of thioredoxin disulfide targets using a quantitative proteomics approach based on isotope-coded affinity tags. J. Prot. Res. 7, 5270–5276. doi: 10.1021/pr800633y
Han, D., Canali, R., García, J., Aguilera, R., Gallaher, T. K., Cadenas, E. (2005). Sites and mechanisms of aconitase inactivation by peroxynitrite: modulation by citrate and glutathione. Biochemistry 44, 11986–11996. doi: 10.1021/bi0509393
Hernández, J. A., Corpas, F. J., Gómez, M., del Río, L. A., Sevilla, F. (1993). Salt-induced oxidative stress mediated by activated oxygen species in pea leaf mitochondria. Physiol. Planta 89, 103–110. doi: 10.1111/j.1399-3054.1993.tb01792.x
Hess, D. T., Matsumoto, A., Kim, S. O., Marshall, H. E., Stamler, J. S. (2005). Protein S-nitrosylation: Purview and parameters. Nat. Rev. Mol. Cell Biol. 6, 150–166. doi: 10.1038/nrm1569
Holzmeister, C., Fröhlich, A., Sarioglu, H., Bauer, N., Durner, J., Lindermayr, C. (2011). Proteomic analysis of defense response of wildtype Arabidopsis thaliana and plants with impaired NO-homeostasis. Proteomics 11, 1664–1683. doi: 10.1002/pmic.201000652
Horling, F., König, J., Dietz, K. J. (2002). Type II peroxiredoxin C, a member of the peroxiredoxin family of Arabidopsis thaliana: its expression and activity in comparison with other peroxiredoxins. Plant Physiol. Biochem. 40, 491–499. doi: 10.1016/S0981-9428(02)01396-7
Horling, F., Lamkemeyer, P., Konig, J., Finkemeier, I., Kandlbinder, A., Baier, M., et al. (2003). Divergent light, ascorbate, and oxidative stress-dependent regulation of expression of the peroxiredoxin gene family in Arabidopsis. Plant Physiol. 131, 317–325. doi: 10.1104/pp.010017
Hoshida, H., Tanaka, K., Hibino, T., Hayashi, Y., Tanaka, A., Takabe, T., et al. (2000). Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast glutamine synthetase. Plant Mol. Biol. 43, 103–111. doi: 10.1023/A:1006408712416
Hu, J., Huang, X., Chen, L., Sun, X., Lu, C., Zhang, L., et al. (2015). Site-specific nitrosoproteomic identification of endogenously S-nitrosylated proteins in Arabidopsis. Plant Physiol. 167, 1731–1746. doi: 10.1104/pp.15.00026
Huang, J., Niazi, A. K., Young, D., Rosado, L. A., Vertommen, D., Bodra, N. (2017). Self-protection of cytosolic malate dehydrogenase against oxidative stress in Arabidopsis. J. Exp. Bot. 69, 3491–3505. doi: 10.1093/jxb/erx396
Huang, J., Willems, P., Wei, B., Tian, C., Ferreira, R. B., Bodra, N., et al. (2019). Mining for protein S-sulfenylation in Arabidopsis uncovers redox-sensitive sites. PNAS 116, 21256–21261. doi: 10.1073/pnas.1906768116
Hurd, T. R., Raquejo, R., Filipovska, A., Brown, S., Prime, T. A., Robinson, A. J., et al. (2008). Complex I within oxidatively-stressed bovine heart mitochondria is glutathionylated on Cys-531 and Cys-704 of the 75-kDa subunit: potential role of Cys residues in decreasing oxidative damage. J. Biol. Chem. 283, 24801–24815. doi: 10.1074/jbc.M803432200
Iglesias-Baena, I., Barranco-Medina, S., Lázaro-Payo, A., López-Jaramillo, F. J., Sevilla, F., Lázaro, J. J. (2010). Characterization of plant sulfiredoxin and role of sulphinic form of 2-Cys peroxiredoxin. J. Exp. Bot. 6, 1509–1521. doi: 10.1093/jxb/erq016
Iglesias-Baena, I., Barranco-Medina, S., Sevilla, F., Lázaro, J. J. (2011). The dual targeted plant sulfiredoxin retroreduces the sulfinic form of atypical mitochondrial peroxiredoxin. Plant Physiol. 155, 944–955. doi: 10.1104/pp.110.166504
Ischiropoulos, H., Zhu, L., Chen, J., Tsai, M., Martin, J. C., Smith, C. D. (1992). Peroxynitrite-mediated tyrosine nitration catalyzed by superoxide dismutase. Arch. Biochem. Biophys. 298, 431–437. doi: 10.1016/0003-9861(92)90431-u
Jaffrey, S. R., Snyder, S. H. (2001). The biotin switch method for the detection of S-nitrosylated proteins. Sci. Signal. 2001, pl1. doi: 10.1126/stke.2001.86.pl1
Jayawardhane, J., Cochrane, D. W., Vyas, P. V., Bykova, N. V., Vanlerberghe, G. C., Igamberdiev, A. U. (2020). Roles for Plant Mitochondrial Alternative Oxidase Under Normoxia, Hypoxia, and Reoxygenation Conditions. Front. Plant Sci. 11, 566. doi: 10.3389/fpls.2020.00566
Jiménez, A., Hernández, J. A., del Rio, L. A., Sevilla, F. (1997). Evidence for the Presence of the Ascorbate-Glutathione Cycle in Mitochondria and Peroxisomes of Pea Leaves. Plant Physiol. 114, 275–284. doi: 10.1104/pp.114.1.275
Jiménez, A., Hernández, J. A., Barcelo, A. R., Sandalio, L. M., del Rio, L. A., Sevilla, F. (1998). Mitochondrial and peroxisomal ascorbate peroxidase of pea leaves. Physiol. Planta 104, 687–692. doi: 10.1034/j.1399-3054.1998.1040424.x
Jin, Z., Wang, Z., Ma, Q., Sun, L., Zhang, L., Liu, Z., et al. (2017). Hydrogen sulfide mediates ion fluxes inducing stomatal closure in response to drought stress in Arabidopsis thaliana. Plant Soil 419, 141–152. doi: 10.1007/s11104-017-3335-5
Keilin, D., Hartree, E. F. (1937). Reaction of nitric oxide with haemoglobin and methaemoglobin. Nature 139, 548. doi: 10.1038/139548a0
Kil, I. S., Park, J.-W. (2005). Regulation of Mitochondrial NADP+-dependent Isocitrate Dehydrogenase Activity by Glutathionylation. J. Biol. Chem. 280, 10846–10854. doi: 10.1074/jbc.M411306200
Kitajima, S., Kurioka, M., Yoshimoto, T., Shindo, M., Kanaori, K., Tajima, K., et al. (2008). A cysteine residue near the propionate side chain of heme is the radical site in ascorbate peroxidase. FEBS J. 275, 470–480. doi: 10.1111/j.1742-4658.2007.06214.x
Kolbert, Zs., Barroso, J. B., Brouquisse, R., Corpas, F. J., Gupta, K. J., Lindermayr, C., et al. (2019). A forty year journey: The generation and roles of NO in plants. Nitric. Oxide 93, 53–70. doi: 10.1016/j.niox.2019.09.006
Konrad, A., Banze, M., Follman, H. (1996). Mitochondria of plant leaves contain two thioredoxins. Completion of the thioredoxin profile of higher plants. J. Plant Physiol. 149, 317–321. doi: 10.1016/S0176-1617(96)80128-3
Laloi, C., Rayapuram, N., Chartier, Y., Grienenberger, J. M., Bonnard, G., Meyer, Y. (2001). Identification and characterization of a mitochondrial thioredoxin system in plants. Proc. Natl. Acad. Sci. U.S.A. 98, 14144–14149. doi: 10.1073/pnas.241340898
Lamotte, O., Bertoldo, J. B., Besson-Bard, A., Rosnoblet, C., Aimé, S., Hichami, S., et al. (2015). Protein S-nitrosylation: specificity and identification strategies in plants. Front. Chem. 2:114:114. doi: 10.3389/fchem.2014.00114
Lázaro, J. J., Jiménez, A., Camejo, D., Iglesias-Baena, I., Martí, M., Lázaro-Payo, A., et al. (2013). Dissecting the integrative antioxidant and redox systems in plant mitochondria. Effect of stress and S-nitrosylation. Front. Plant Sci. 4, 460. doi: 10.3389/fpls.2013.00460
Lee, J. H., Yang, E. S., Park, J. W. (2003). Inactivation of NADP+-dependent Isocitrate Dehydrogenase by Peroxynitrite: implications for cytotoxicity and alcohol-induced liver injury. J. Biol. Chem. 27, 51360–51371. doi: 10.1074/jbc.M302332200
Leferink, N. G. H., van Duijn, E., Barendregt, A., Heck, A. J. R., van Berkel, W. J. H. (2009). Galactonolactone Dehydrogenase Requires a Redox-Sensitive Thiol for Optimal Production of Vitamin C. Plant Physiol. 150, 596–605. doi: 10.1104/pp.109.136929
Leitner, M., Vandelle, E., Gaupels, F., Bellin, D., Delledonne, M. (2009). NO signals in the haze. Nitric oxide signalling in plant defence. Curr. Opin. Plant Biol. 12, 451–458. doi: 10.1016/j.pbi.2009.05.012
Liebthal, M., Maynard, D., Dietz, K.-J. (2018). Peroxiredoxins and Redox Signaling in Plants. Antioxid. Redox Signal. 28, 609–624. doi: 10.1089/ars.2017.7164
Lindermayr, C., Saalbach, G., Durner, J. (2015). Proteomic identification of S-Nitrosylated proteins in Arabidopsis. Plant Physiol. 137, 921–930. doi: 10.1104/pp.104.058719
Lindermayr, C. (2018). Crosstalk between reactive oxygen species and nitric oxide in plants: Key role of S-nitrosoglutathione reductase. Free Radic. Biol. Med. 122, 110–115. doi: 10.1016/j.freeradbiomed.2017.11.027
Liu, L., Hausladen, A., Zeng, M., Que, L., Heitman, J., Stamler, J. S. (2001). A metabolic enzyme for S-nitrosothiol conserved from bacteria to humans. Nature 410, 490–494. doi: 10.1038/35068596
Locato, V., Cimini, S., De Gara, L. (2018). ROS and redox balance as multifaceted players of cross-tolerance: epigenetic and retrograde control of gene expression. J. Exp. Bot. 69, 3373–3391. doi: 10.1093/jxb/ery168
Mailloux, R. J., Jin, X., Willmore, W. G. (2014). Redox regulation of mitochondrial function with emphasis on cysteine oxidation reactions. Redox Biol. 2, 123–139. doi: 10.1016/j.redox.2013.12.011
Marcus, F., Chamberlain, S. H., Che, C., Masiarz, F. R., Shin, S., Yee, B. C., et al. (1991). Plant thioredoxin h: an animal-like thioredoxin occurring in multiple cell compartments. Arch. Biochem. Biophys. 287, 195–198. doi: 10.1016/0003-9861(91)90406-9
Martí, M. C., Olmos, E., Calvete, J. J., Díaz, I., Barranco-Medina, S., Whelan, J., et al. (2009). Mitochondrial and nuclear localization of a novel pea thioredoxin: identification of its mitochondrial target proteins. Plant Physiol. 150, 646–657. doi: 10.1104/pp.109.138073
Martí, M. C., Florez-Sarasa, I., Camejo, D., Ribas-Carbó, M., Lázaro, J. J., Sevilla, F., et al. (2011). Response of the mitochondrial antioxidant redox system and respiration to salinity in pea plants. J. Exp. Bot. 62, 3863–3874. doi: 10.1093/jxb/err076
Martí, M. C., Florez-Sarasa, I., Camejo, D., Pallol, B., Ortiz, A., Ribas-Carbó, M., et al. (2012). Response of mitocondrial antioxidant system and respiratory pathways to reactive nitrogen species in pea leaves. Physiol. Plant. 147, 194–206. doi: 10.1111/j.1399-3054.2012.01654.x
Martínez-Ruiz, A., Araújo, I. M., Izquierdo-Álvarez, A., Hernansanz-Agustín, P., Lamas, S., Serrador, J. (2013). Specificity in S-nitrosylation: A short-range mechanism for NO signaling? Antioxid. Redox Signal. 19, 1220–1235. doi: 10.1089/ars.2012.5066
Mata-Pérez, C., Sánchez-Calvo, B., Padilla, M. N., Begara-Morales, J. C., Valderrama, R., Corpas, F. J., et al. (2017). Nitro-fatty acids in plant signaling: New key mediators of nitric oxide metabolism. Redox Biol. 11, 554–561. doi: 10.1016/j.redox.2017.01.002
Maxwell, D. P., Wang, Y., McIntosh, L. (1999). The alternative oxidase lowers mitochondrial reactive oxygen production in plant cells. Proc. Natl. Acad. Sci. U.S.A. 96, 8271–8276. doi: 10.1016/0003-9861(91)90406-9
Meyer, Y., Belin, C., Delorme-Hinoux, V., Reichheld, J. P., Riondet, C. (2012). Thioredoxin and glutaredoxin systems in plants: molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox Signal. 17, 1124–1160. doi: 10.1089/ars.2011.4327
Mhamdi, A., Van Breusegem, F. (2018). Reactive oxygen species in plant development. Development 145, dev164376. doi: 10.1242/dev.164376
Michelet, L., Zaffagnini, M., Marchand, C., Collin, V., Decottignies, P., Tsan, P., et al. (2005). Glutathionylation of chloroplast thioredoxin f is a redox signaling mechanism in plants. Proc. Natl. Acad. Sci. U. S. A. 102, 16478–16483. doi: 10.1073/pnas.0507498102
Millar, A. H., Whelan, J., Soole, K. L., Day, D. A. (2011). Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol. 62, 79–104. doi: 10.1146/annurev-arplant-042110-103857
Mnatsakanyan, R., Markoutsa, S., Walbrunn, K., Roos, A., Verhelst, S. H. L., Zahedi, R. P. (2019). Proteome-wide detection of S-nitrosylation targets and motifs using bioorthogonal cleavable-linker-based enrichment and switch technique. Nat. Commun. 10, 2195. doi: 10.1038/s41467-019-10182-4
Møller, I. M., Igamberdiev, A. U., Bykova, N. V., Finkemeier, I., Rasmusson, L. G., Schwarzländer, M. (2020). Matrix Redox Physiology Governs the Regulation of Plant Mitochondrial Metabolism through Posttranslational Protein Modifications. Plant Cell 32, 573–594. doi: 10.1105/tpc.19.00535
Møller, I. M. (2015). Mitochondrial metabolism is regulated by thioredoxin. Proc. Natl. Acad. Sci. U.S.A. 112, 3180–3181. doi: 10.1073/pnas.1502425112
Nietzel, T., Mostertz, J., Hochgräfe, F., Schwarzländer, M. (2017). Redox regulation of mitochondrial proteins and proteomes by cysteine thiol switches. Mitochondrion 33, 72–83. doi: 10.1016/j.mito.2016.07.010
Nietzel, T., Mostertz, J., Ruberti, C., Née, G., Fuchs, P., Wagner, S., et al. (2020). Redox-mediated kickstart of mitochondrial energy metabolism drives resource-efficient seed germination. Proc. Natl. Acad. Sci. U.S.A. 117, 741–751. doi: 10.1073/pnas.1910501117
Niture, S. K., Velu, C. S., Bailey, N. I., Srivenugopal, K. S. (2005). S-thiolation mimicry: quantitative and kinetic analysis of redox status of protein cysteines by glutathione-affinity chromatography. Arch. Biochem. Biophys. 444, 174–184. doi: 10.1016/j.abb.2005.10.013
Noctor, G., De Paepe, R., Foyer, C. H. (2007). Mitochondrial redox biology and homeostasis in plants. Trends Plant Sci. 12, 125–134. doi: 10.1016/j.tplants.2007.01.005
Noctor, G., Lelarge-Trouverie, C., Mhamdi, A. (2015). The metabolomics of oxidative stress. Phytochemistry 112, 33–53. doi: 10.1016/j.phytochem.2014.09.002
Noguera-Mazon, V., Lemoine, J., Walker, O., Rouhier, N., Salvador, A., Jacquot, J. P., et al. (2006). Glutathionylation induces the dissociation of 1-Cys D-peroxiredoxin non-covalent homodimer. J. Biol. Chem. 281, 31736–31742. doi: 10.1074/jbc.M602188200
Ojeda, V., Pérez-Ruiz, J. M., González, M., Nájera, V. A., Sahrawy, M., Serrato, A. J., et al. (2017). NADPH thioredoxin reductase C and thioredoxins act concertedly in seedling development. Plant Physiol. 174, 1436–1448. doi: 10.1104/pp.17.00481
Ortega-Galisteo, A. P., Rodríguez-Serrano, M., Pazmiño, D. M., Gupta, D. K., Sandalio, L. M., Romero-Puertas, M. C. (2012). S-Nitrosylated proteins in pea (Pisum sativum L.) leaf peroxisomes: changes under abiotic stress. J. Exp. Bot. 63, 2089–2103. doi: 10.1093/jxb/err414
Ortiz-Espín, A., Locato, V., Camejo, D., Schiermeyer, A., De Gara, L., Sevilla, F., et al. (2015). Over-expression of Trxo1 increases the viability of tobacco BY-2 cells under H2O2 treatment. Ann. Bot. 116, 571–582. doi: 10.1093/aob/mcv076
Ortiz-Espín, A., Iglesias-Fernández, R., Calderón, A., Carbonero, P., Sevilla, F., Jiménez, A. (2017). Mitochondrial AtTrxo1 is transcriptionally regulated by AtbZIP9 and AtAZF2 and affects seed germination under saline conditions. J. Exp. Bot. 68, 1025–1038. doi: 10.1093/jxb/erx012
Palmieri, M. C., Lindermayr, C., Bauwe, H., Steinhauser, C., Durner, J. (2010). Regulation of plant glycine decarboxylase by s-nitrosylation and glutathionylation. Plant Physiol. 152, 1514–1528. doi: 10.1104/pp.109.152579
Paul, B. D., Snyder, S. H. (2012). H2S signaling through protein sulfhydration and beyond. Nat. Rev. Mol. Cell Biol. 13, 499–507. doi: 10.1038/nrm3391
Pérez-Pérez, M. E., Mauriès, A., Maes, A., Tourasse, N. J., Hamon, M., Lemaire, S. D., et al. (2017). The deep thioredoxome in Chlamydomonas reinhardtii: New insights into redox regulation. Mol. Plant 10, 1107–1125. doi: 10.1016/j.molp.2017.07.009
Pulido, P., Cazalis, R., Cejudo, F. J. (2009). An antioxidant redox system in the nucleus of wheat seed cells suffering oxidative stress. Plant J. 57, 132–145. doi: 10.1111/j.1365-313X.2008.03675.x
Purvis, A. C. (1997). Role of the alternative oxidase in limiting superoxide production in plant mitochondria. Physiol. Plant 100, 165–170. doi: 10.1111/j.1399-3054.1997.tb03468.x
Puyaubert, J., Fares, A., Reze, N., Peltier, J. B., Baudouin, E. (2014). Identification of endogenously S-nitrosylated proteins in Arabidopsis plantlets: effect of cold stress on cysteine nitrosylation level. Plant Sci. 215-216, 150–156. doi: 10.1016/j.plantsci.2013.10.014
Qiu, C., Sun, J., Wang, Y., Sun, L., Xie, H., Ding, Y., et al. (2019). First nitrosoproteomic profiling deciphers the cysteine S-nitrosylation involved in multiple metabolic pathways of tea leaves. Sci. Rep. 9, 17525. doi: 10.1038/s41598-019-54077-2
Rasmusson, A. G., Wallström, S. V. (2010). Involvement of mitochondria in the control of plant cell NAD(P)H reduction levels. Biochem. Soc Trans. 38, 661–666. doi: 10.1042/bst0380661
Reichheld, J. P., Meyer, E., Khafif, M., Bonnard, G., Meyer, Y. (2005). AtNTRB is the major mitochondrial thioredoxin reductase in Arabidopsis thaliana. FEBS Lett. 579, 337–342. doi: 10.1016/j.febslet.2004.11.094
Reinholdt, O., Schwab, S., Zhang, Y., Reichheld, J.-P., Fernie, A. R., Hagemann, M., et al. (2019a). Redox-regulation of photorespiration through mitochondrial thioredoxin o1. Plant Physiol. 181, 442–457. doi: 10.1104/pp.19.00559
Reinholdt, O., Bauwe, H., Hagemann, M., Timm, S. (2019b). Redox-regulation of mitochondrial metabolism through thioredoxin o1 facilitates light induction of photosynthesis. Plant Signal. Behav. 14, 1674607. doi: 10.1080/15592324.2019.1674607
Rey, P., Becuwe, N., Barrault, M. B., Rumeau, D., Havaux, M., Biteau, B., et al. (2007). The Arabidopsis thaliana sulfiredoxin is a plastidic cysteine-sulfinic acid reductase involved in the photooxidative stress response. Plant J. 49, 505–514. doi: 10.1111/j.1365-313X.2006.02969.x
Riemenschneider, A., Wegele, R., Schmidt, A., Papenbrock, J. (2005). Isolation and characterization of a D-cysteine desulfhydrase protein from Arabidopsis thaliana. FEBS J. 272, 1291–1304. doi: 10.1111/j.1742-4658.2005.04567.x
Rizza, S., Montagna, C., Cardaci, S., Maiani, E., Di Giacomo, G., Sanchez-Quiles, V., et al. (2016). S-nitrosylation of the Mitochondrial Chaperone TRAP1 Sensitizes Hepatocellular Carcinoma Cells to Inhibitors of Succinate Dehydrogenase. Cancer Res. 76, 4170–4182. doi: 10.1158/0008-5472.CAN-15-2637
Rodríguez-Ruiz, M., Mateos, R. M., Codesido, V., Corpas, F. J., Palma, J. M. (2017). Characterization of the galactono-1,4-lactone dehydrogenase from pepper fruits and its modulation in the ascorbate biosynthesis. Role Nitric. Oxide Redox Biol. 12, 171–181. doi: 10.1016/j.redox.2017.02.009
Romero-Puertas, M. C., Sandalio, L. M. (2016). Nitric Oxide Level Is Self-Regulating and Also Regulates Its ROS Partners. Front. Plant Sci. 7, 316. doi: 10.3389/fpls.2016.00316
Romero-Puertas, M. C., Laxa, M., Matte, A., Zaninotto, F., Finkemeier, I., Jones, A. M., et al. (2007). S-Nitrosylation of peroxiredoxin II E promotes peroxynitrite-mediated tyrosine nitration. Plant Cell 19, 4120–4130. doi: 10.1105/tpc.107.055061
Romero-Puertas, M. C., Campostrini, N., Matte, A., Righetti, P. G., Perazzolli, M., Zolla, L., et al. (2008). Proteomic analysis of S-nitrosylated proteins in Arabidopsis thaliana undergoing hypersensitive response. Proteomics 8, 1459–1469. doi: 10.1002/pmic.200700536
Sánchez-Guerrero, A., Fernández del-Saz, N., Florez-Sarasa, I., Ribas-Carbó, M., Ferni, A. R., Jiménez, A., et al. (2019). Coordinated responses of mitochondrial antioxidative enzymes, respiratory pathways and metabolism in Arabidopsis thaliana thioredoxin trxo1 mutants under salinity. Env. Exp. Bot. 162, 212–222. doi: 10.1016/j.envexpbot.2019.02.026
Sandalio, L. M., Gotor, C., Romero, L. C., Romero-Puertas, M. C. (2019). Multilevel regulation of peroxisomal proteome by posttranslational modifications. Int. J. Mol. Sci. 20, 4881. doi: 10.3390/ijms20194881
Schimmeyer, J., Bock, R., Meyer, E. H. (2016). L-Galactono-1,4-lactone dehydrogenase is an assembly factor of the membrane arm of mitochondrial complex I in Arabidopsis. Plant Mol. Biol. 90, 117–126. doi: 10.1007/s11103-015-0400-4
Schmidtmann, E., König, A.-C., Orwat, A., Leister, D., Hartl, M., Finkemeier, I. (2014). Redox regulation of Arabidopsis mitochondrial citrate synthase. Mol. Plant 7, 156–169. doi: 10.1093/mp/sst144
Scuffi, D., Álvarez, C., Laspina, N., Gotor, C., Lamattina, L., García-Mata, C. (2014). Hydrogen sulfide generated by L-cysteine desulfhydrase acts upstream of nitric oxide to modulate abscisic acid-dependent stomatal closure. Plant Physiol. 166, 2065–2076. doi: 10.1104/pp.114.245373
Selinski, J., König, N., Wellmeyer, B., Hanke, G. T., Linke, V., Neuhaus, H. E., et al. (2014). The plastid-localized NAD-dependent malate dehydrogenase is crucial for energy homeostasis in developing Arabidopsis thaliana seeds. Mol. Plant 7, 170–186. doi: 10.1093/mp/sst151
Selinski, J., Scheibe, R., Day, D. A., Whelan, J. (2018). Alternative Oxidase Is Positive for Plant Performance. Trends Plant Sci. 23, 588–597. doi: 10.1016/j.tplants.2018.03.012
Sevilla, F., Camejo, D., Ortiz-Espín, A., Calderón, A., Lázaro, J. J., Jiménez, A. (2015a). The thioredoxin/peroxiredoxin/sulfiredoxin system: current overview on its redox function in plants and regulation by reactive oxygen and nitrogen species. J. Exp. Bot. 66, 2945–2955. doi: 10.1093/jxb/erv146
Sevilla, F., Jiménez, A., Lázaro, J. J. (2015b). “What do the plant mitochondrial antioxidant and redox systems have to say in salinity, drought and extreme temperature abiotic stress situations?,” in Reactive oxygen species and oxidative damage in plants under stress. Eds. Gupta, D. K., Palma, J. M., Corpas, F. J. (Switzerland: Springer Cham), 23–55.
Smith, C. A., Melino, V. J., Sweetman, C., Soole, K. L. (2009). Manipulation of alternative oxidase can influence salt tolerance in Arabidopsis thaliana. Physiol. Plant 137, 459–472. doi: 10.1111/j.1399-3054.2009.01305.x
Stevens, F. J., Li, A. D., Lateef, S. S., Anderson, L. E. (1997). Identification of potential inter-domain disulfides in three higher plant mitochondrial citrate synthases: paradoxical differences in redox-sensitivity as compared with the animal enzyme. Photosynth. Res. 54, 185–197. doi: 10.1023/A:1005991423503
Sweetlove, L. J., Heazlewood, J. L., Herald, V., Holtzapffel, R., Day, D. A., Leaver, C. J., et al. (2002). The impact of oxidative stress on Arabidopsis mitochondria. Plant J. 32, 891–904. doi: 10.1046/j.1365-313X.2002.01474.x
Tanou, G., Job, C., Rajjou, L., Arc, E., Belghazi, M., Diamantidis, G., et al. (2009). Proteomics reveals the overlapping roles of hydrogen peroxide and nitric oxide in the acclimation of citrus plants to salinity. Plant J. 60, 795–804. doi: 10.1111/j.1365-313X.2009.04000.x
Toledo, J. C., Audi, R., Ogusucu, R., Monteiro, G., Netto, L. E. S., Augusto, O. (2011). Horseradish peroxidase compound I as a tool to investigate reactive protein-cysteine residues: from quantification to kinetics. Free Radic. Biol. Med. 50, 1032–1038. doi: 10.1016/j.freeradbiomed.2011.02.020
Turrens, J. F. (2003). Mitochondrial formation of reactive oxygen species. J. Physiol. 552, 335–344. doi: 10.1111/j.1469-7793.2003.00335.x
Umbach, A. L., Fiorani, F., Siedow, J. N. (2005). Characterization of transformed Arabidopsis with altered alternative oxidase levels and analysis of effects on reactive oxygen species in tissue. Plant Physiol. 139, 1806–1820. doi: 10.1104/pp.105.070763
Umekawa, Y., Ito, K. (2019). Thioredoxin o- mediated reduction of mitocondrial alternative oxidase. J. Biol. Chem. 165, 57–65. doi: 10.1093/jb/mvy082
Van Ruyskensvelde, V., Van Breusegem, F., Van Der Kelen, K. (2018). Post-transcriptional regulation of the oxidative stress response in plants. Free Rad. Biol. Med. 122, 181–192. doi: 10.1016/j.freeradbiomed.2018.02.032
Vidigal, P., Carvalho, R., Amancio, S., Carvalho, L. (2013). Peroxiredoxins are involved in two independent signalling pathways in the abiotic stress protection in Vitis vinifera. Biol. Planta 57, 675–683. doi: 10.1007/s10535-013-0346-9
Vishwakarma, A., Tetali, S. D., Selinski, J., Scheibe, R., Padmasree, K. (2015). Importance of the alternative oxidase (AOX) pathway in regulating cellular redox and ROS homeostasis to optimize photosynthesis during restriction of the cytochrome oxidase pathway in Arabidopsis thaliana. Annu. Bot. 116, 555–569. doi: 10.1093/aob/mcv122
Wang, Y., Berkowitz, O., Selinski, J., Xu, Y., Hartmann, A., Whelan, J. (2018). Stress responsive mitochondrial proteins in Arabidopsis thaliana. Free Rad. Biol. Med. 122, 28–39. doi: 10.1016/j.freeradbiomed.2018.03.031
Waszczak, C., Carmody, M., Kangasjärvi, J. (2018). Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 69, 209–236. doi: 10.1146/annurev-arplant-042817-040322
Wedmann, R., Onderka, C., Wei, S., Szijártó, I. A., Miljkovic, J. L., Mitrovic, A., et al. (2016). Improved tag-switch method reveals that thioredoxin acts as depersulfidase and controls the intracellular levels of protein persulfidation. Chem. Sci. 7, 3414–3426. doi: 10.1039/c5sc04818d
Yang, E. S., Richter, C. H., Chun, J. S., Huh, T. L., Kang, S. S., Park, J. W. (2002). Inactivation of NADP-dependent isocitrate dehydrogenase by nitric oxide. Free Radic. Biol. Med. 33, 927–937. doi: 10.1016/s0891-5849(02)00981-4
Yang, H., Mu, J., Chen, L., Feng, J., Hu, J., Li, L., et al. (2015). S-nitrosylation positively regulates ascorbate peroxidase activity during plant stress responses. Plant Physiol. 167, 1604–1615. doi: 10.1104/pp.114.255216
Yoshida, K., Hisabori, T. (2014). Mitochondrial isocitrate dehydrogenase is inactivated upon oxidation and reactivated by thioredoxin-dependent reduction in Arabidopsis. Front. Environ. Sci. 2, 38. doi: 10.3389/fenvs.2014.00038
Yoshida, K., Hisabori, T. (2016). Adenine nucleotide-dependent and redox-independent control of mitochondrial malate dehydrogenase activity in Arabidopsis thaliana. Biochim. Biophys. Acta 1857, 810–818. doi: 10.1016/j.bbabio.2016.03.001
Yoshida, K., Noguchi, K., Motohashi, K., Hisabori, T. (2013). Systematic exploration of thioredoxin target proteins in plant mitochondria. Plant Cell Physiol. 54, 875–892. doi: 10.1093/pcp/pct037
Yu, L.-X., Zhang, C.-J., Shang, H.-Q., Wang, X.-F., Wei, M., Yang, F.-J., et al. (2013). Exogenous hydrogen sulfide enhanced antioxidant capacity, amylase activities and salt tolerance of cucumber hypocotyls and radicles. J. Integr. Agric. 12, 445–456. doi: 10.1016/S2095-3119(13)60245-2
Zaffagnini, M., Fermani, S., Costa, A., Lemaire, S. D., Trost, P. (2013). Plant cytoplasmic GAPDH: redox post-translational modifications and moonlighting properties. Front. Plant Sci. 4, 450. doi: 10.3389/fpls.2013.00450
Zannini, F., Roret, T., Przybyla-Toscano, J., Dhalleine, T., Rouhier, N., Couturier, J. (2018). Mitochondrial Arabidopsis thaliana TRXo Isoforms Bind an Iron–Sulfur Cluster and Reduce NFU Proteins In Vitro. Antioxidants 7, 142. doi: 10.3390/antiox7100142
Zechmann, B. (2014). Compartment-specific importance of glutathione during abiotic and biotic stress. Front. Plant Sci. 5, 566. doi: 10.3389/fpls.2014.00566
Zhang, J., Jin, B., Li, L., Block, E. R., Patel, J. M. (2005). Nitric oxide-induced persistent inhibition and nitrosylation of active site cysteine residues of mitochondrial cytochrome-c oxidase in lung endothelial cells. Am. J. Physiol. Cell Physiol. 288, C840–C849. doi: 10.1152/ajpcell.00325.2004
Zhang, H., Ye, Y. K., Wang, S. H., Luo, J. P., Tang, J., Ma, D. F. (2009). Hydrogen sulfide counteracts chlorophyll loss in sweetpotato seedling leaves and alleviates oxidative damage against osmotic stress. Plant Growth Regul. 58, 243–250. doi: 10.1007/s10725-009-9372-1
Zhang, H., Wu, Z., Wang, C., Li, Y., Xu, J.-R. (2014). Germination and infectivity of microconidia in the rice blast fungus Magnaporthe oryzae. Nat. Commun. 5, 4518. doi: 10.1038/ncomms5518
Zhou, H., Zhang, J., Shen, J., Zhou, M., Yuan, X., Xie, Y. (2020). Redox-based protein persulfidation in guard cell ABA signaling. Plant Signal. Behav. 15, 1741987. doi: 10.1080/15592324.2020.1741987
Keywords: abiotic stress, mitochondria, persulfidation, redox regulation, S-glutathionylation, S-nitrosation, S-oxidation, thioredoxin
Citation: Martí MC, Jiménez A and Sevilla F (2020) Thioredoxin Network in Plant Mitochondria: Cysteine S-Posttranslational Modifications and Stress Conditions. Front. Plant Sci. 11:571288. doi: 10.3389/fpls.2020.571288
Received: 10 June 2020; Accepted: 08 September 2020;
Published: 23 September 2020.
Edited by:
José Manuel Palma, Consejo Superior de Investigaciones Científicas (CSIC), SpainReviewed by:
Cecilia Gotor, Institute of Plant Biochemistry and Photosynthesis (IBVF), SpainIrene García, Institute of Plant Biochemistry and Photosynthesis (IBVF), Spain
Copyright © 2020 Martí, Jiménez and Sevilla. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francisca Sevilla, ZnNldmlsbGFAY2ViYXMuY3NpYy5lcw==