 Christina Rønn Ingvardsen
Christina Rønn Ingvardsen Henrik Brinch-Pedersen
Henrik Brinch-Pedersen- Crop Genetics and Biotechnology, Department of Agroecology, Aarhus University, Slagelse, Denmark
Cannabis sativa L. is an ancient crop used for fiber and seed production and not least for its content of cannabinoids used for medicine and as an intoxicant drug. Due to the psychedelic effect of one of the compounds, tetrahydrocannabinol (THC), many countries had regulations or bands on Cannabis growing, also as fiber or seed crop. Recently, as many of these regulations are getting less tight, the interest for the many uses of this crop is increasing. Cannabis is dioecious and highly heterogenic, making traditional breeding costly and time consuming. Further, it might be difficult to introduce new traits without changing the cannabinoid profile. Genome editing using new breeding techniques might solve these problems. The successful use of genome editing requires sequence information on suitable target genes, a genome editing tool to be introduced into plant tissue and the ability to regenerate plants from transformed cells. This review summarizes the current status of Cannabis breeding, uncovers potentials and challenges of Cannabis in an era of new breeding techniques and finally suggests future focus areas that may help to improve our overall understanding of Cannabis and realize the potentials of the plant.
Introduction to medical Cannabis
Cannabis (Cannabis sativa L.) is an annual, predominantly dioecious (male and female flowers occur on separate plants), rarely monoecious (male and female flowers occur on the same plant) plant (Chandra et al., 2020). It is wind pollinated and flowers under short day conditions. Cannabis has a diploid genome (2n = 20) and its karyotype is composed of 9 pairs of autosomes and one pair of sexual chromosomes (X and Y) (Divashuk et al., 2014). The plants have been used by man for at least 6000 years (Li, 1974). The existence of an extant natural population is doubtful, and any ‘wild’ species might be naturalized from domesticated plants indicating that the current genetic variation is most probably due to the action of man (Small, 2017). Cannabis has traditionally been classified as a single species (Small and Cronquist, 1976). This is supported by molecular studies (Oh et al., 2016; Zhang et al., 2018). However, Cannabis is often divided into subspecies or groups based on chemotype, ecotype, crop-type (fiber or drug) or leaflet morphology (Grassi and McPartland, 2017; McPartland, 2017; Small, 2017). Grouping is often problematic, as the types readily inter-cross and a lot of hybrids exist and as classification based on crop-type is somewhat dependent on legislation. In this paper, we focus on Cannabis used for medical purposes, e.g., plants with a high level of cannabinoids, and will therefore use the term hemp for Cannabis grown for fiber and seeds and the term medical or drug type Cannabis for varieties grown for their cannabinoid content, independent of the tetrahydrocannabinol (THC)/cannabidiol (CBD) ratio.
Cannabis produces a range of secondary metabolites, the best known are the phytocannabinoids. Although as many as 120 different cannabinoids have been reported (see Radwan et al. (2017) for review), the most abundant cannabinoids are THC, CBD, cannabigerol (CBG) and cannabinol (CBN) (Flores-Sanchez and Verpoorte, 2008; Fischedick et al., 2010; ElSohly et al., 2016). Tetrahydrocannabinolic acid (THCA) and cannabidiolic acid (CBDA) are made from the common precursor, cannabigerolic acid (CBGA) whereas CBN is an oxidation product of THC. Acidic forms of cannabinoids are biosynthesized in the trichomes on the female flowers (Sirikantaramas et al., 2005; Livingston et al., 2020). The storage of the toxic cannabinoids in trichomes minimizes the risk of self-intoxication (Sirikantaramas et al., 2005). The acidic forms are spontaneously decarboxylated into THC and CBD by heat and light. THC is the main psychoactive compound in drug type Cannabis (Pertwee, 1988), but also beneficial effects are reported (Grotenhermen, 2003). Moreover, other cannabinoids, especially CBD, have attracted interest for their pharmacological properties. CBD has been reported to act as an antidepressant, to relieve pain and anxiety and to reduce inflammation. It is believed to have beneficial effects in a variety of diseases, such as Alzheimer’s and Parkinson’s diseases, multiple sclerosis, Huntington’s disease, epilepsy, and cancer (Pisanti et al., 2017). The resin from the glandular trichomes also contain terpenes (Flores-Sanchez and Verpoorte, 2008). Terpenes does not only add flavor to the product but also have medical properties, not least in synergic action (entourage effect) with cannabinoids (Ferber et al., 2020).
In planta, the secondary metabolites are believed to protect the plants against various pathogens and insects. Both cannabinoids and terpenes have been shown to possess antifungal activity (McPartland, 1984; Wanas et al., 2016) and to be toxic to insects (Taura et al., 2007; Mithöfer and Boland, 2012; Bedini et al., 2016). Although the active component is not known, Cannabis resin might have antibacterial activity (Radošević et al., 1962) and extract of Cannabis leaves is active against nematodes (Mukhtar et al., 2013).
Solving the many challenges in Cannabis sativa calls for the use of all available tools. In the current review, we discuss the challenges and possibilities for New Breeding Techniques, such as genome editing in medical Cannabis. To set the scene, we first present a brief status of Cannabis breeding and the level of genetic diversity. We then address the requirements for successful tissue culture and transformation with a focus on the status of micro propagation, regeneration and transient as well as stable transformation. Finally, we discuss the many opportunities for using genome editing in the improvement of medical Cannabis.
Breeding, genetic diversity and genetic markers
Breeding
Due to the high-value products, medical Cannabis is often produced in greenhouse or indoor facilities where the plants are propagated like a horticultural crop using stem cuttings (Vassilevska-Ivanova, 2019; Monthony et al., 2021c). The use of cuttings ensures that only female plants with a higher level of cannabinoids are used for production. The dioecy behavior of Cannabis, making it an obligately outbreeding species, the limited number of genetic markers and the anecdotal start of breeding of medical cultivars make breeding of drug type Cannabis challenging. Early breeding was selection, done by the illegal market with decades of interbreeding and hybridization without record of parentage (Barcaccia et al., 2020; Gilchrist et al., 2023). This means that the genetic identity of a medical Cannabis strains cannot be reliably inferred from its name, as studies have shown that some strains with different names were genetically similar, and some strains with identical names were genetically different (Sawler et al., 2015; Dufresnes et al., 2017; Punja et al., 2017; Reimann-Philipp et al., 2020; Adhikary et al., 2021). However, the fact that breeding has been illegal does not mean that it is inefficient, as the level of THC has increased (Mehmedic et al., 2010). For a comprehensive review on medical Cannabis breeding, readers are referred to Barcaccia et al. (2020).
The level and composition of cannabinoids and terpenoids as well as stability in production, flowering time and lower resource input are in focus in modern medical Cannabis breeding, with more focus on CBD and other non-psychoactive cannabinoids. Resistance against insects, pathogens and viruses is also in high demand. Healthy mother plants are essential. However, maintenance of mother plants in contained humid environments poses a challenge in relation to attacks by plant pathogens such as powdery mildew. This significant challenge can only be kept at tolerable levels by a strict growth control including air circulation, ventilation, and moisture control as the strict regulations for medical products do not permit any use of pesticides.
Doubled haploids
The production of doubled haploids (DH) in Cannabis would be highly advantageous, as it would be possible to produce female pure lines in one generation. Haploid plants have in other plant species been produced via androgenesis (anther or microspore culture), gynogenesis, parthenogenesis or wide hybridization-chromosome elimination (Ishii et al, 2016; Hooghvorst and Nogués, 2021). Later chromosome duplication in the haploid plants is performed, either spontaneously or by chemical treatment. Although DH production via microspore culture has been investigated in Cannabis, successful DH production has so far not been established (Adhikary et al., 2021). Cannabis seems to be recalcitrant to androgenesis induction, although very few embryos can be developed (Galán-Ávila et al., 2021a). The method used for successful doubled haploid production seems to be species dependent wherefor also the other methods should be investigated for their usefulness in Cannabis. Recently, CRISPR/Cas have been used to develop haploid-inducer lines in both monocot and dicot plants (Kelliher et al., 2019). As also suggested by others (Hesami et al., 2021a; Simiyu et al., 2022), this method might be very useful in Cannabis.
Polyploidization
Polyploidization is used as a tool in plant breeding to improve desirable plant characteristics such as larger organs and higher yield (Sattler et al., 2016). Even though the expected “giga” effect is not always achieved, higher production of secondary metabolites in autotetraploid medical plants is seen in several cases as reviewed by others (Dhawan and Lavania, 1996; Iannicelli et al., 2020; Niazian and Nalousi, 2020). As genomic stress occurs after polyploidization, genomic rearrangements, gene loss and/or changes in gene expression might occur (Iannicelli et al., 2020; Niazian and Nalousi, 2020). This might give a changed chemical profile with lower or even missing production of secondary metabolites, making it difficult to predict the outcome of polyploidization in new species or even other genotypes.
In Cannabis, the effect of polyploidization has been studied in hemp as well as drug-type Cannabis (Bagheri and Mansouri, 2015; Mansouri and Bagheri, 2017; Parsons et al., 2019; Hesami et al., 2021a). In both types, the tetraploid plants had broader leaves with bigger and less dense stomata, both clear signs of polyploidization. Cuttings of the drug-type Cannabis had reduced rooting ability, a phenomenon also observed in hop (Trojak-Goluch and Skomra, 2013; Parsons et al., 2019). When the level of cannabinoids was analyzed, only small changes were found. Levels of CBD was increased by 9%, whereas the level of THC was unchanged in drug-type Cannabis (Parsons et al., 2019). In the hemp-type plants, the level of THC was reduced in the female flowers with no change in the amount of CBD (Bagheri and Mansouri, 2015).
The terpene profile was not analyzed in the hemp-type Cannabis, but in the drug-type, the terpene profile changed after polyploidization, as mainly the contents of sesquiterpenes increased (Parsons et al., 2019). A change in terpene profile after polyploidization was also found in hop. Although the general level was lower, there was an increase in terpenes desirable for the brewing industry (Trojak-Goluch and Skomra, 2013).
These experiments did not show very promising results as far as an increase in cannabinoid is concerned. It should be reminded, however, that only one genotype per experiment gave tetraploid plants that could be analyzed. As there is often a difference between genotypes, the effect of polyploidization on the level of cannabinoids, terpenes and other important traits might have a different and more positive outcome in other trials, not least after crossing of polyploids with different genetic backgrounds.
Genetic structure and diversity
Several studies using STRUCTURE analysis position hemp and drug type Cannabis in separate clusters (Sawler et al., 2015; Lynch et al., 2016; Dufresnes et al., 2017). The genetic differences between the groups are distributed across the genome and are not restricted to loci involved in cannabinoid production (Dufresnes et al., 2017b; Sawler et al., 2015). Whether hemp or drug type Cannabis is having more heterozygosity seems to depend on the study, which probably reflects differences in the selected cultivars (Sawler et al., 2015; Lynch et al., 2016). The lower genetic diversity in drug type Cannabis compared to hemp found in some studies (Sawler et al., 2015) might be due to inbreeding and genetic bottlenecks in the illegal Cannabis production. However, studies including a broader set of genotypes are needed to create more knowledge about the heterozygosity of Cannabis from all regions (Kovalchuk et al., 2020). Such studies also provide the widest genetic background for medical Cannabis breeding.
Genetic markers such as single sequence repeats (SSRs) and Inter Simple Sequence Repeats (ISSR) have shown a high degree of genetic diversity in drug type Cannabis (Punja et al., 2017; Soler et al., 2017; de Oliveira Pereira Ribeiro et al., 2020). The analyses not only showed diversity between but also within cultivars (Punja et al., 2017; Soler et al., 2017). The high diversity within a cultivar means that hardly any reduction in the genetic variation was found after one round of selfing (Punja and Holmes, 2020). Some of this high variation found after selfing might be due to accumulation of somatic mutations within plants been propagated as cuttings for a long time (Adamek et al., 2022).
The high genetic diversity found in Cannabis is very useful for breeding new varieties. For medical Cannabis, however, homogenous material is needed, and varieties must be multiplied via cuttings. Introduction of single gene traits like disease resistance genes in Cannabis by traditional cross breeding, without affecting the genetic background and thereby the cannabinoid and terpene profile is difficult. Genome editing might solve this issue, see below.
Use of genetic markers
The use of genetic markers in drug-type Cannabis has mainly focused on analysis of Cannabis samples and plants to discriminate between hemp type and drug type material, to evaluate genetic variance and to identify female plants. Some examples will be highlighted here, for more comprehensive information, especially on early work, readers are referred to Onofri and Mandolino (2017); Punja et al. (2017) and Hesami et al., 2020. Further, a panel of 41 robust SSRs, with an average of four markers per chromosome, is provided by Barcaccia et al. (2020).
Markers for chemotype
As drug type Cannabis is illegal in many countries, there is a great need to be able to detect the presence of this type of Cannabis in seized samples. An important issue to consider here is the balance between speed, simplicity of the analysis, affordability, and accuracy. Many different assays have been developed, not only to discriminate between hemp and drug types, but also to establish from which geographic location the sample might originate.
The simplest form of markers is based on the genes for tetrahydrocannabinolic acid synthase (THCAS) and cannabidiolic acid synthase (CBDAS). As DNA extracted from the seized material is analyzed, there is no need to wait for plants to grow in case of seed material or to extract cannabinoids to analyze for the level of THC and CBD. The markers reported by Kojoma et al. (2006) and Rotherham and Harbison (2011) was based on THCAS, only. At the time, it was believed that the two enzymes belonged to the same locus so that plants having the THCA synthase would contain only THC or a mix of THC and CBD. Other markers are based on both THCAS and CBDAS sequences (Weiblen et al., 2015; Welling et al., 2016; Cascini et al., 2019; Toth et al., 2020), which is in line with newer information showing that the two enzymes are found in two closely linked loci (Laverty et al., 2019).
Other types of markers have also been shown to be useful; autosomal microsatellite markers and markers based on mitochondrial and chloroplast DNA. SSR’s have been used to differentiate between samples (Soler et al., 2016; Dufresnes et al., 2017; Houston et al., 2017; Soler et al., 2017; Houston et al., 2018; de Oliveira Pereira Ribeiro et al., 2020; Ioannidis et al., 2022b). It seems that a limited number of markers, from 6 to 13, is enough, not only to sort Cannabis from hop, but to individualize and differentiate between types (drug versus hemp) and to say something about geographic origin (Houston et al., 2017; de Oliveira Pereira Ribeiro et al., 2020). Often markers can be multiplexed (Dufresnes et al., 2017; Houston et al., 2017; de Oliveira Pereira Ribeiro et al., 2020), reducing labor and cost.
Thus, genetic markers are useful as forensic tool to give a confirmation whether a sample is Cannabis and to discriminate between hemp and drug type samples. This can otherwise be difficult, especially with seed samples where there is no obvious difference. Also, information about the geographical origin of samples can give valuable information about distribution routes of illegal products.
Sex markers
A mentioned, the karyotype is composed of 9 pairs of autosomes and one pair of sexual chromosomes (X and Y) (Divashuk et al., 2014). Sex determination in dioecious Cannabis is believed to function through a X-to-autosome balance system, where X/A = 1 are female (XX) and X/A = 0.5 are male (XY) rather than by a Y-active system (Ainsworth, 2000; Vyskot and Hobza, 2004). Monoecious hemp cultivars having both male and female flowers on the same plant, have two X chromosomes (Faux et al., 2014; Razumova et al., 2016). Different genetic markers have been developed to sort male and female plants (Mandolino et al., 1999; Mandolino et al., 2002; Törjék et al., 2002; Toth et al., 2020). It seems that the sex determination is somewhat leaky, as the environment, especially photoperiod, hormones and unknown genetic components other than the sex chromosomes also plays a role (Schaffner, 1921; Faux et al., 2014). In female drug-type Cannabis, spontaneous formation of hermaphroditic plants, with both female and male flowers, can be as high as 10% (Punja and Holmes, 2020). This ability of female plants to produce male flowers independent of the presence of the Y chromosome is used in the production of feminized seeds, as female plants can be treated with thiosulfate to produce male flowers (Lubell and Brand, 2018).
The prerequisites for genome editing
Due to the many uses of Cannabis, genome editing would be a very desirable tool. Not only would we get a deeper insight into the cannabinoid pathway, but also many other genes important for the many uses of Cannabis could be investigated. The successful use of genome editing requires a genome editing tool, sequence information of suitable target genes, introduction of the construct into plant tissue and the ability to regenerate shoots from explant tissue.
Gene editing techniques
New Breeding Techniques (NBT) have emerged as alternatives to classical plant breeding and conventional transgenesis. These new techniques facilitate development of novel varieties more precisely and faster than by classical breeding giving genome modifications indistinguishable from those introduced by conventional breeding and chemical or physical mutagenesis (Lusser et al., 2012). NBT include the sequence-specific nuclease (SSN) tools such as Zinc Finger Nucleases (ZNF´s) (Petolino, 2015), Transcription Activator-Like Effector Nucleases (TALENs) (Khan et al., 2017) and Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) (Chen et al., 2019; Montecillo et al., 2020), all allowing for targeted mutagenesis of candidates without unwanted side mutations.
A common feature for ZFNs, TALENs and CRISPR/Cas is the programmability to cleave in specific locations and generate DNA double strand breaks (DSBs) that stimulate standard cellular repair mechanisms including non-homologous end joining (NHJE) and homology-directed repair (HDR) (Voytas, 2013; Gupta et al., 2019). By NHEJ, the repair at the DSB site is often imprecise which leads to introduction of small deletions/insertions at the site of break, resulting in knockout of gene function via frame shift mutations. HDR requires a homologous DNA segment as template to correct or replace existing sequence. By HDR it is thus possible to make specific nucleotide changes.
ZNF´s and TALENs are hybrid proteins created by fusing ZF and TALE DNA-binding domain to the non-specific cleavage domain of FokI endonuclease (Petolino, 2015; Khan et al., 2017). The FokI endonuclease non-specific cleavage domain must dimerize to cleave the DNA target. In August 2013, CRISPR/Cas emerged as an alternative genome editing method in plants (Feng et al., 2013; Li et al., 2013; Nekrasov et al., 2013; Shan et al., 2013; Xie and Yang, 2013). The CRISPR method was originally discovered as an antiviral defense system widespread in prokaryotes (Wiedenheft et al., 2012). The system is based on a Cas9 nuclease which can be targeted to a specific genomic sequence by an easily engineered single guide RNA (sgRNA) of 20 base pair (bp). A protospacer adjacent motif (PAM) is needed adjacent to the 3’ end of the 20 bp target. Originally, the purpose of the PAM sequence was to distinguish self from non-self in prokaryotes.
The CRISPR/Cas technology has become the preferred method for making targeted mutations in plants without undesired side mutations, due to ease of use, precision, efficiency, and low cost (Shan et al., 2013; Kumar et al., 2020), and since first reported a long range of rapports on modifications in crops have been reported in multiple plant species (Gupta et al., 2019). Ease of multiplexing, i.e., the simultaneous targeting if several genes with a single molecular construct, is another major advantage of CRISPR/Cas9 technology compared to ZFN and TALEN (Shan et al., 2013; Lowder et al., 2016; Ma et al., 2016; Čermák et al., 2017; Kumar et al., 2020). Although many report the use of CRISPR techniques in the coding sequence of genes, they can be used in promoters and upstream open reading frames as well (Holme et al., 2017; Rodríguez-Leal et al., 2017; Si et al., 2020). Recently DNA editing in plant plastics has also become a possibility (Kang et al., 2021).
The technology is quickly developing (Zhu et al., 2020), and new CRISPR nucleases like Cas12a creating 5´overhangs or nucleases having different PAM recognition sites are constantly expanding the perspective (Jaganathan et al., 2018; Swarts and Jinek, 2018). Very recently, PAM-less CRISPR systems are emerging (Ren et al., 2021). Nucleases that only make a single-strand break (nickases) can be used in pairs, each requiring a sgRNA. By positioning the two nicks close to each other on opposite strands, a break is created. As two sgRNAs are required, this paired nicking dramatically increases the specificity and thus reduces off-targeting in unwanted places. HDR is still challenging in plants and several developments in the CRISPR techniques have been developed to overcome this problem. Adenine base editors (ABEs) changing adenine (A) to guanine (G) and cytosine base editors (CBEs) changing cytosine (C) to thymine (T) are now working in plants (Zong et al., 2018; Hua et al., 2020). A new technique for editing is the prime editing technique, where bases in the target site is edited based on the sequence of a prime editing guide RNA (Lin et al., 2020; Zhu et al., 2020; Zong et al., 2022). Further, multifunctional genome editing systems are now emerging, making it possible to do both base editing and knockout simultaneous using a single construct (Li et al., 2020). On top of local changes such as minor deletions/insertions and base editing, the introduction of bigger structural variations are now also possible (Lu et al., 2021). Some new techniques are firstly developed for monocots and optimization might be needed to use them in dicots such as Cannabis.
Online tools are available to guide the design of efficient sgRNAs. Five different sgRNA designing tools and their main characteristics have been reviewed by Hesami et al. (2021a). The algorithms in such programs are developed based on the assessment of many thousands of gRNAs targeting genes. However, most tools are not developed based on plant data and the predicted efficiency is not always in accordance with the found results (Pauwels et al., 2018; Naim et al., 2020). An updated tool based on plant data is needed to increase the efficiency of plant sgRNAs.
Status of available sequence data
There is now a reference genome for Cannabis, which can be found at NCBI (Supplementary Table S1). This reference genome, called cs10, is made from female material in the CBD-high isolate CBDDRx-18 (Grassa et al., 2021). On top of the reference genome, NCBI lists thirteen different Genbank accessions of genome assemblies, five of which are at chromosome level (Supplementary Table S1). These thirteen assemblies mostly represent female plants but are made from different types of Cannabis (TDC high, CBD high, mixed profile and hemp), making them a valuable tool for finding sequence data for potential candidate genes. As Cannabis is highly heterozygous, not least at the THCAS and CBCAS gene loci, the creation of a Cannabis pangenome is in high demand (Hurgobin et al., 2021). Six different transcriptome assemblies from hemp and medical Cannabis are also available from NCBI and the Cannabis Genome Browser (Supplementary Table S2). On top of this, very many sequences (Nucleotide, WGS, RNA-seq, Nanopore and Targeted amplicon sequences form the Phylos Bioscience/Open Cannabis Project) can be blasted from the NCBI homepage, if the gene of interest should not appear in the genome assemblies. Reference genomes and other assemblies for both the chloroplast and mitochondrial genomes are also available (Supplementary Table S3).
It must be stressed that for gene editing techniques, the precise genetic sequence of the cultivar at hand is needed in order to design guides. This means that both alleles of each candidate gene to be targeted must be sequenced. Further, the availability of genomic sequence information not only provide the candidate genes for targeting but also gives the opportunity to analyze for possible off-targets, not only in gene families with high sequence similarity but also unexpected off-targets in unrelated genome regions. However, a very accurate screening for off-targets might be difficult in the highly heterozygous Cannabis genome.
Tissue culture and plant regeneration
Micropropagation
The plant material for production of medical Cannabis is obtained from cuttings from mother plants. The maintenance of mother plants demands a lot of space and considerable effort to keep the material free from diseases. Further, there is a need to store valuable breeding material. Propagation and maintenance of material via in vitro culture would be a way to solve this issue and tissue culture has been a topic in several papers. Several papers have reported methods for in vitro propagation in Cannabis, methods that often are described as regeneration (Table 1) (Lata et al., 2016; Piunno et al., 2019; Smýkalová et al., 2019; Ioannidis et al., 2020; Wróbel et al., 2022). However, most papers do not describe regeneration from a single non-differentiated cell but instead report shoot formation from pre-existing meristems, once the apical dominance is broken. Summaries of published micropropagation studies were recently published (Hesami et al., 2021a; Monthony et al., 2021c). Readers with special interest in micro propagation and/or preservation are referred to these papers.
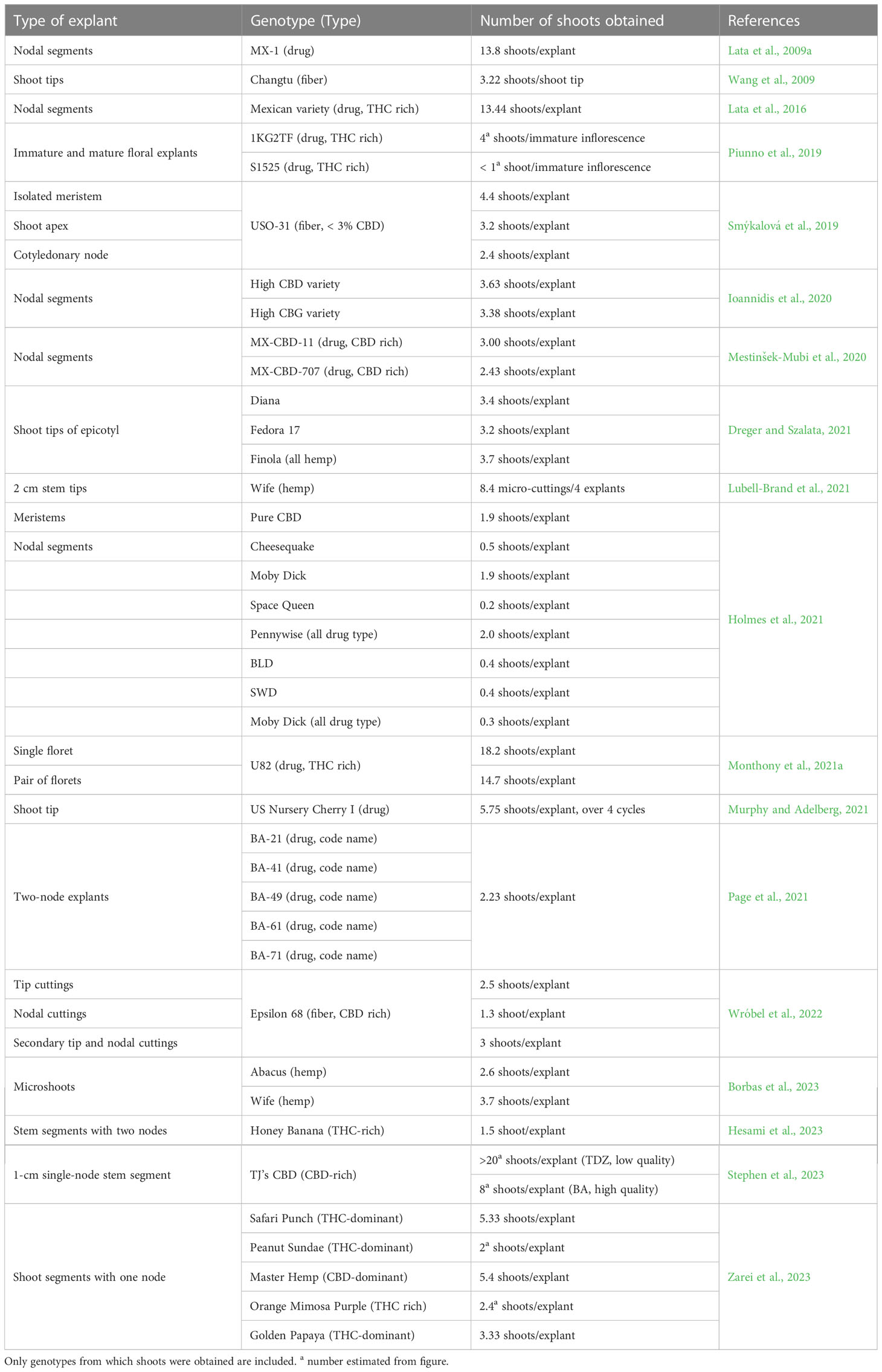
Table 1 List of Cannabis sativa studies reporting micropropagation from pre-existing meristems.
In general, finding the right growth medium seems to be very cultivar dependent (Grulichova et al., 2017; Codesido et al., 2020; Stephen et al., 2023). This makes it difficult to compare studies using different combinations of cultivars and media. It remains to be uncovered if this is due to a very limited number of plants being very willing to respond in vitro, or whether the ideal medium composition still needs to be discovered. This cultivar dependence implies that the use of hemp as a proxy for medicinal type Cannabis may not be successful (Page et al., 2021). Although there might not be a big difference in multiplication rate between fiber and drug types (Table 1). The results might also be dependent on whether the starting material is taken directly from the greenhouse or from tissue that has been in tissue culture for some time. The position (basal versus apical) of the stem explant also seems to affect multiplication rates (Hesami et al., 2023). There could be a lingering effect from plant growth regulators found in plants with roots that might influence the multiplication rate (Page et al., 2021). A high number of shoot proliferation has been reported in some cases (Lata et al., 2009a; Lata et al., 2016) but in most cases the number of shoots per explant is only between 2 and 3 (Table 1). The use of floral reversion might be a way to move forward, as this method increased the multiplication rate up to eightfold (Piunno et al., 2019; Monthony et al., 2021a). It should be noted that achieving the highest multiplication rate might give lower quality and lower rooting ability (Stephen et al., 2023). Rooting is very important for successful micropropagation and has been a topic in several papers (e.g.: Zarei et al., 2021; Ioannidis et al., 2022a; Kurtz et al., 2022; Stephen et al., 2023).
There is a general need for improvement in more cultivars, if micropropagation should be of general use. Scientific research can be successful, even though a very limited (or only one) number of plant cultivars can be used. However, in production, medical Cannabis growers need to be able to use the technique in all their material before they invest in tissue culture facilities. It is most probable that several companies already have developed successful protocols for micropropagation. Most of these are, however, kept as trade secrets, if not funded by public means (Adhikary et al., 2021; Zarei et al., 2023). Resent papers show that improvement of micropropagation is still an interest also in universities (Borbas et al., 2023; Hesami et al., 2023; Stephen et al., 2023) and new methods, such as the use of bioreactors (Rico et al., 2022), photosynthetic proficiency measurements (Pepe et al., 2022) and photoautotrophic micropropagation (Zarei et al., 2021) have been investigated.
Micropropagation using synthetic seeds has been investigated as an alternative solution for propagation and conservation of germplasm. The use of synthetic seeds in Cannabis was first reported by Lata et al. (2009b; Lata et al., 2012) using the cultivar MX. Recently, a paper on commercial scale synthetic seed production using the elite cultivar ‘Slurricane’ was published (Zarei et al., 2022). The developed method was very successful, as a regrowth rate of 100% was seen after storage for 150 days. Cryopreservation has also been investigated as a means for long term storage on in vitro material (Lata et al., 2019; Downey et al., 2021). Although these methods are not of immediate importance for genome editing, they represent valuable ways of storing high-value material such as modified cultivars.
Plant regeneration
In vitro propagation from pre-existing meristems is very useful for multiplying material for medical Cannabis production. However, if regeneration from single cells is optimized, a much higher multiplication rate might be obtained. Further, to produce CRISPR/Cas mutated plants, an efficient protocol for de novo regeneration of plants from single cells is needed to avoid chimeric plants. In Cannabis, such de novo regeneration seems very difficult to obtain from callus or tissue without preformed meristems. This recalcitrance to regeneration is the main obstacle to an efficient genome editing protocol in Cannabis (Monthony et al., 2021b).
Recalcitrance is a common problem in tissue culture (Altpeter et al., 2016). For years, many different combinations of explants and plant growth regulators have been used to try to solve the difficulty of regenerating plants from very many species, often without great success. The ability to regenerate seems to be not only species dependent but also cultivar dependent. The recalcitrance might be linked to a very high degree of apical dominance and/or difficulties in cellular reprogramming of already differentiated cells (Sugimoto et al., 2019), making the generation of shoots difficult. This calls for further research into whether there is a general explanation for recalcitrance to regenerate across plant species.
Testing many different combinations of explant type, explant age, type of gelling agent, type of carbohydrate source, type, and balance of PGRs and addition of other supplements such as Zn or polyamines is often tedious and time consuming, sometimes with a low success rate. This process might be optimized using the ability of machine learning to discover non-linear relationships and concealed interactions (García-Pérez et al., 2020). Machine learning has already been used in Cannabis to optimize in vitro seed germination (Hesami et al., 2021b; Pepe et al., 2021a; Aasim et al., 2022) and to study in vitro shoot growth and development (Pepe et al., 2021b) and callus morphology (Hesami and Jones, 2021). Further, as many research projects are performed by PhD students and post docs, with a strong demand for an outcome of scientific papers, the focus is often turned away from comprehensive, long-term research aiming at optimizing regeneration protocols. There might also be a lack of reports showing negative results. All this might have slowed down the progress of regeneration.
A direct comparison between experiments is sometimes difficult as results are given as responding explant % or number of shoots per shooting explant (Table 2). Although the lowest percentage of responding explants is reported in hemp (Slusarkiewicz-Jarzina et al., 2005), there is not a clear trend for a difference between hemp and drug type cultivars. The biggest difference can be explained by the type of explants used. Experiments using hypocotyl, stem, or stem nodes have a higher response rate in general (Table 2) (Wielgus et al., 2008; Galán-Ávila et al., 2020; Önol and Yildirim, 2021; Galán-Ávila et al., 2021b).
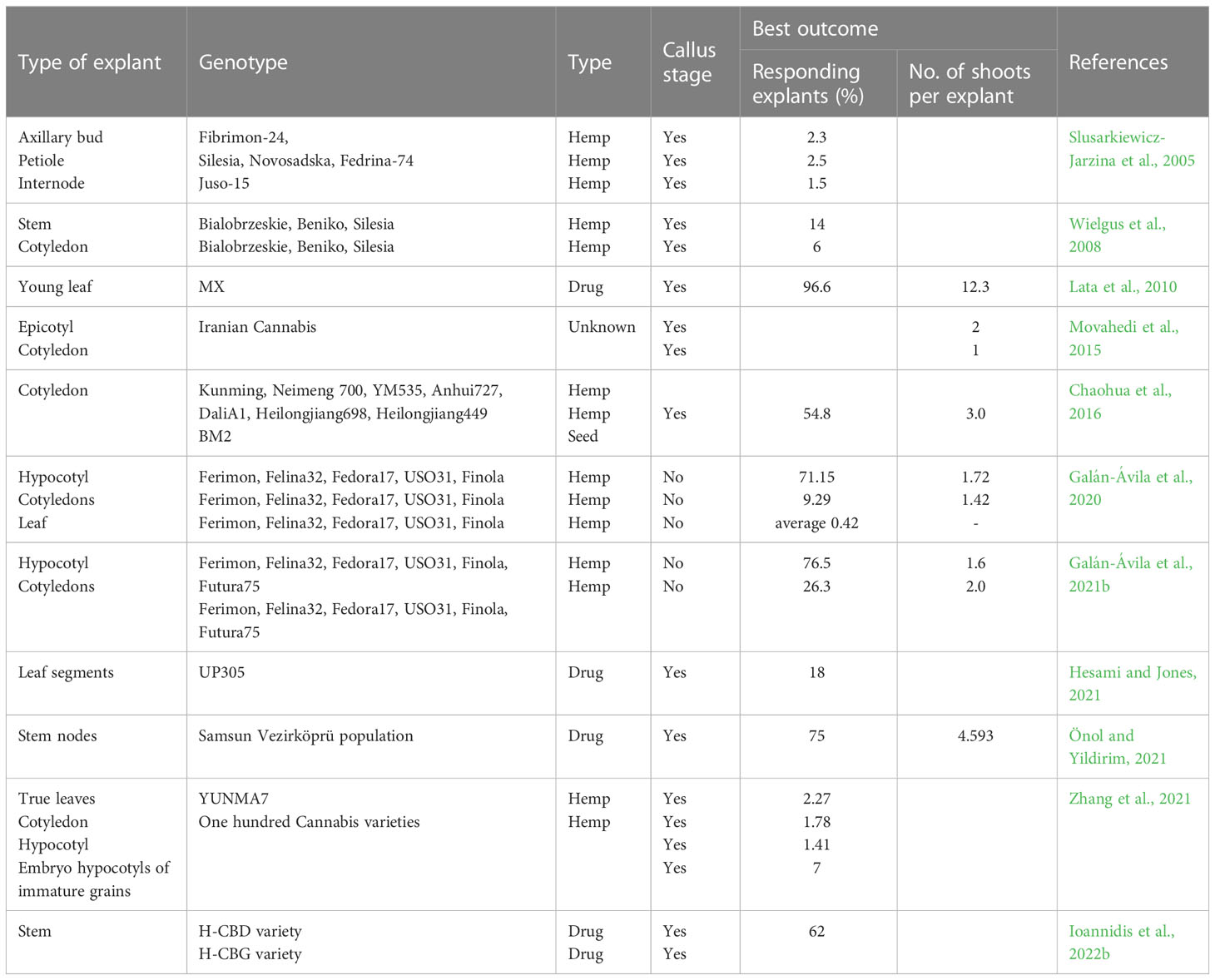
Table 2 List of Cannabis sativa studies reporting regeneration experiments without transformation.
One experiment, using leaf explants, stands out, as almost all explants gave shoots, with an average of 12.3 shoots per callus (Lata et al., 2010). This study has been replicated using ten other drug type Cannabis genotypes (Monthony et al., 2021b). Here, the experiment failed to induce shoots in all the genotypes tested, making it clear that regeneration might not only be tissue specific but also very dependent on genotype. The importance of genotype for regeneration in Cannabis is stressed by the fact that Zhang et al. found a regeneration rate varying from 0 to 7% when screening one hundred genotypes (Zhang et al., 2021). Further, the lack of reproducibility stresses the difficulty of transferring tissue culture methods to other laboratories.
Recently, Galán-Ávila et al. published two papers showing a very high percentage of direct regeneration without a callus phase from hypocotyls (Table 2) (Galán-Ávila et al., 2020; Galán-Ávila et al., 2021b). These results are very promising but remains to be seen whether this method can be transferred to medical Cannabis.
Transformation
Transformation in Cannabis has been a research topic for more than 20 years. Three main transformation techniques have been used, the transformation with Agrobacterium rhizogenes to get hairy roots, transient transformation, and stable transformation. The status of transformation in Cannabis has also been reviewed by others (Feeney and Punja, 2017; Simiyu et al., 2022). For an in-depth discussion of Agrobacterium strains, promoters and selection markers, readers are referred to Hesami et al. (2021a).
Hairy root cultures often have an enhanced ability to synthesize secondary metabolites (Srivastava and Srivastava, 2007). Hairy root cultures were established after transformation with A. rhizogenes in hemp as well as drug-type Cannabis (Wahby et al., 2013; Wahby et al., 2017). Several types of media containing a range of different combinations of hormones were tried, but although callus developed from the hairy root cultures, no shoots were obtained (Wahby et al., 2017). No cannabinoids were present in the hairy root cultures. Similarly, no cannabinoid production was found in cell suspension cultures (Flores-Sanchez et al., 2009). Contrary, hairy root cultures developed from callus without the use of A. rhizogenes produced a very low level of cannabinoids (Farag and Kayser, 2015). The level of cannabinoids might be dependent on the variety of plant and/or of the plant variety-bacterial stain combination. However, higher levels would probably be toxic to the cultures. The ability of callus cultures to form roots was also seen by Feeney and Punja (2003). Unfortunately, regeneration of plants from hairy roots, although possible in some species (Crane et al., 2006; Lütken et al., 2012; de la Torre et al., 2018), is often quite challenging and has not yet been reported in Cannabis.
Transformation using Agrobacterium tumefaciens is often used in plants, both for transient and stable transformation (Dunwell and Wetten, 2012; Krenek et al., 2015). Studies with wild-type A. tumefaciens (MacKinnon et al., 2001; Wahby et al., 2013) as well as recent papers (Tables 3, 4) (Galán-Ávila et al., 2021b; Zhang et al., 2021) show that genetic transformation of Cannabis is possible. However, there seems to be a cultivar difference in the susceptibility to A. tumefaciens. As the infection with Agrobacterium might be considered as a pathogen attack by the plant and the secondary metabolites is known to protect Cannabis against pathogens (McPartland, 1984; Wanas et al., 2016), the difference in susceptibility might be explained by a difference in the secondary metabolite profile (Sorokin et al., 2020). This might partly explain why all stable transformations in Cannabis is done in hemp cultivars (see below; Table 4).
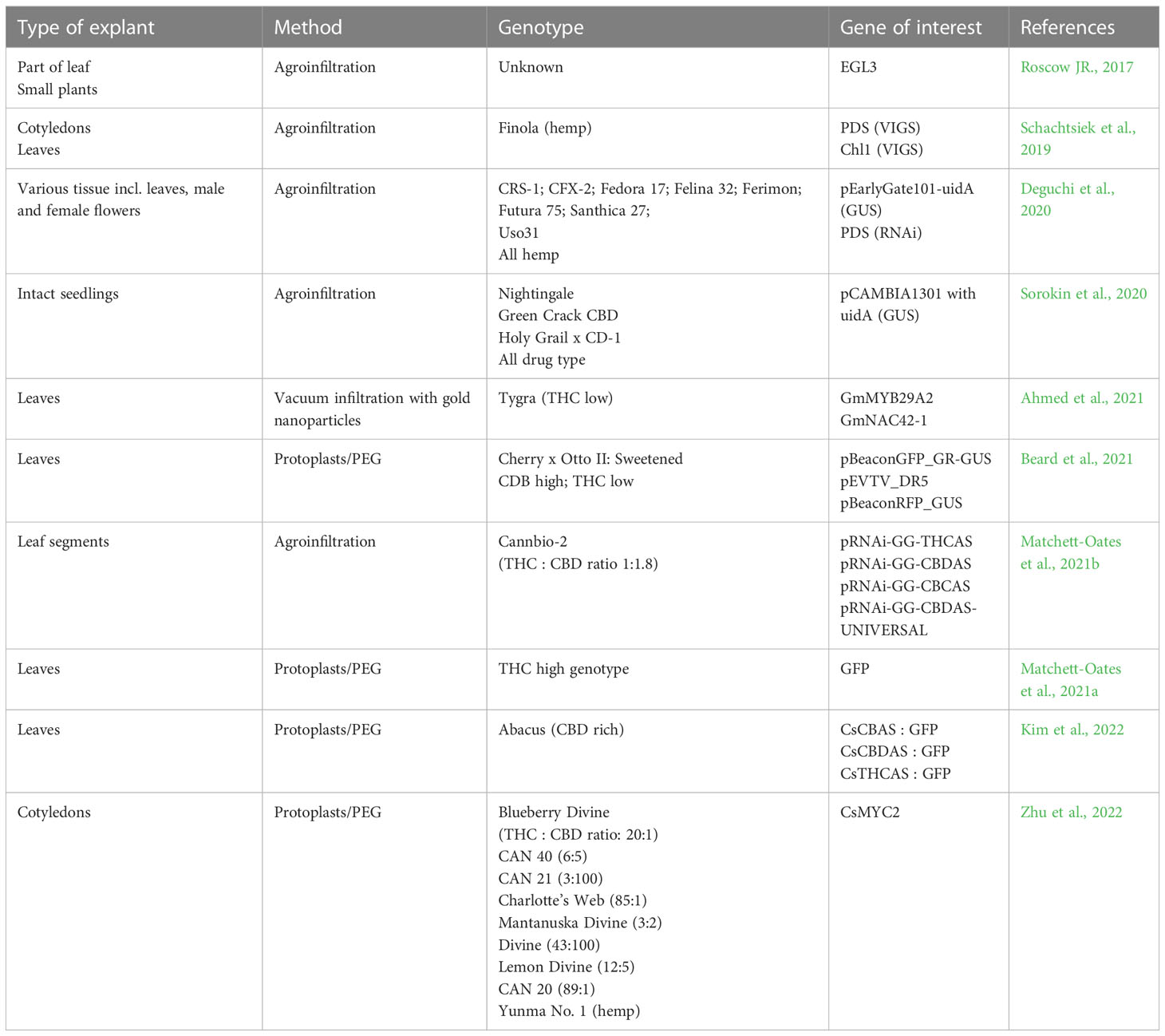
Table 3 List of Cannabis sativa studies reporting transient transformation.
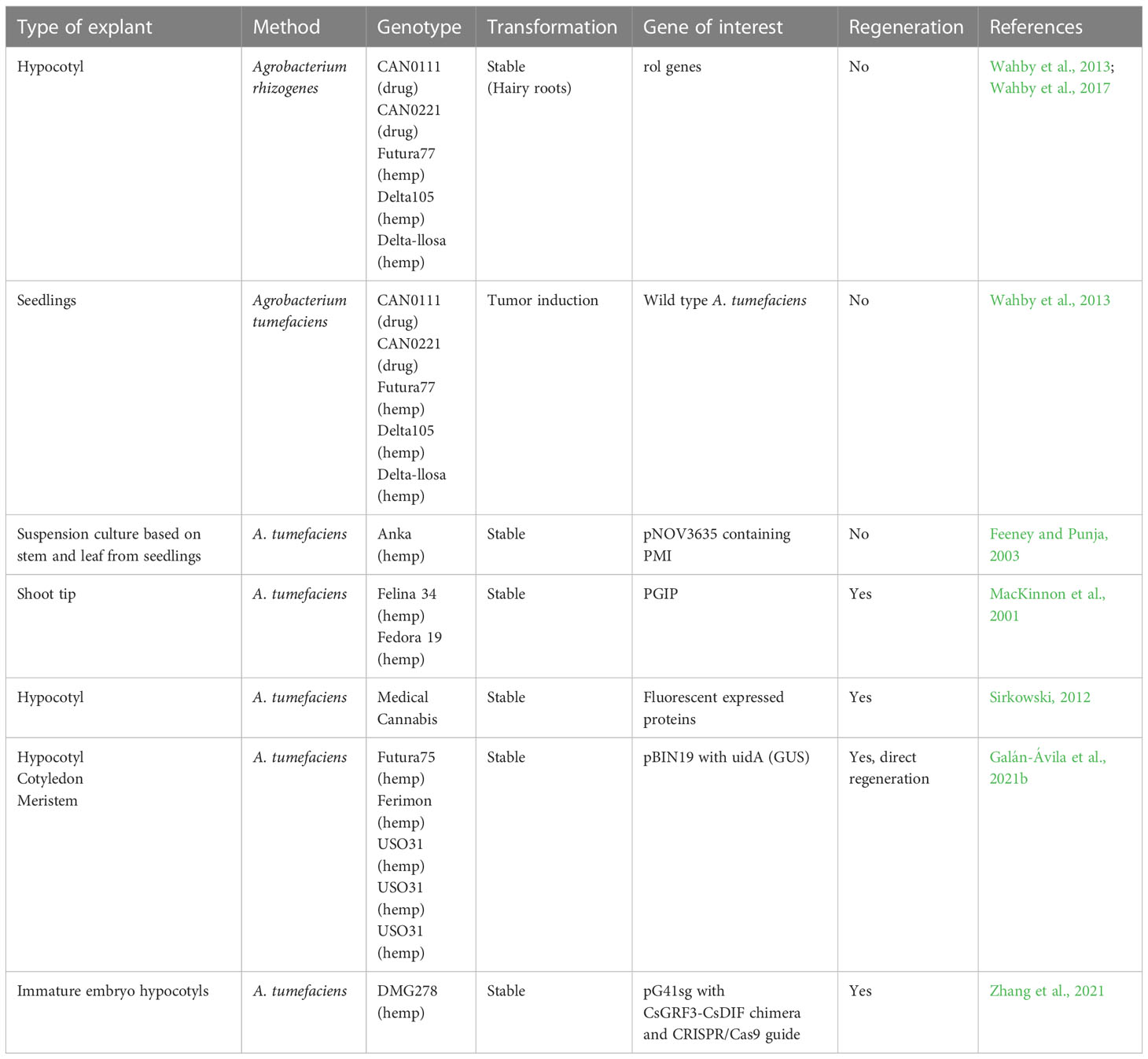
Table 4 List of Cannabis sativa studies reporting stable transformation.
Most probably, the recalcitrance in Cannabis explants is also due to the developmental state of the explants. In many plants, the first choice of explant material for transformation would be very young tissue, like that obtained from young seedlings. Such material is easily obtained from hemp, where seeds are available in big amounts. In medical Cannabis, however, very specific combinations of cannabinoids and terpenes might be lost if seeds must be produced. A method where the meristems of medical plants could be the starting material, is therefore highly adventitious. Recent reports show transformation on existing meristems, in vitro or in planta is possible (Galán-Ávila et al., 2021b; Pandey et al., 2022). Further, developmental regulators can be used to induce new meristems from somatic cells during the transformation process (Maher et al., 2020). One should be aware that the plants obtained from these meristematic techniques might have a higher degree of mosaic tissue than plantlets obtained by regeneration via callus.
Within the last six years several papers report transient transformation in Cannabis using not only reporter genes such as GUS or GFP, but also the down regulation of genes using virus induced gene silencing (VIGS) or RNAi (Table 3). A study from 2013 (Wahby et al., 2013) showed that both hemp and drug type Cannabis is susceptible to wild type A. tumefaciens infection and it is thus not surprising that the most frequently used method for transient transformation is agroinfiltration by vacuum or by using a needleless syringe. A patent by Roscow JR. (2017) describes transient methods to transform Cannabis by vacuum infiltration or by dipping the green parts into an Agrobacterium soup where after vacuum is applied. The treated plant parts were producing trichomes comprising secondary compounds on non-flowering parts of the plant. It is, however, unclear if this property was inherited to the next generation. Transient transformation using agroinfiltration is also reported by several others (Schachtsiek et al., 2019; Deguchi et al., 2020; Sorokin et al., 2020; Matchett-Oates et al., 2021b). Results show that the recalcitrance to be transformed might be due to the plants ability to protect itself against pathogen attack, as the addition of ascorbic acid, scavenging excess ROS, had a positive effect of the transformation efficiency (Deguchi et al., 2020). Other methods for transient transformation include vacuum infiltration of DNA coated gold nanoparticles (Ahmed et al., 2021) and transformation of protoplasts using PEG (Beard et al., 2021; Matchett-Oates et al., 2021a; Kim et al., 2022; Zhu et al., 2022).
Transient transformation, although not leading to stably transformed plants, is a very useful tool for overexpression and silencing studies of many genes. Valuable information about the interaction between Cannabis and Agrobacterium which might be used to improve stable transformation. It seems that transient transformation is possible in both hemp and drug-type material (Deguchi et al., 2020; Sorokin et al., 2020; Matchett-Oates et al., 2021b), which gives the hope that stable transformation will also be possible in medical Cannabis.
Stable transformation is likely needed for genome editing of Cannabis. The first successful stable transformation of hemp was reported by MacKinnon et al. (2001) in the Scottish Crop Research Institute Annual Report. The report gives very little information about the methods used and the transformation efficiency obtained. Hemp suspension culture cells were transformed by Feeney and Punja in 2003 (Feeney and Punja, 2003). Although showing a transformation rate up to 55%, no plant regeneration was obtained. After these two 20-year-old papers, stable transformation was not reported for almost 15 years, most probably reflecting the lack of success of regenerating transformants.
A patent filed by Sirkowski (2012) describes a method for Agrobacterium-mediated transformation of medical Cannabis. The material used is sections of hypocotyl and plants are regenerated from the tissue, but the efficiency is not mentioned. It is not clear in which genotypes the regeneration was successful and if plants were regenerated directly via a callus phase.
Very recently, two groups have been able to stably transform hemp (Galán-Ávila et al., 2020; Zhang et al., 2021) (Table 4). In the method developed by Galán-Ávila et al. (2021b) shoots are regenerated directly from hypocotyl, cotyledon, or meristems without a callus phase. This transformation system is very fast as tissue could be analyzed for GUS already one month after transformation. Both genotypic and explant differences were observed, with the least genotype dependency and the highest transformation rate found using hypocotyls. This method seems very promising, but the use of hypocotyls might be hampering the transferability to medical Cannabis that is usually propagated using cuttings. However, there was a low transformation rate using meristems as one transformant from one genotype was obtained from this material.
The first paper describing the use of genome editing in Cannabis was published in 2021 (Zhang et al., 2021). Using immature embryo hypocotyls from hemp as a starting material, shoots were regenerated from callus. The phytoene desaturase gene was knocked out using CRISPR/Cas9, giving an albino phenotype. Zhang et al. used a recently developed method, where co-transformation with a GRF-GIF chimeric protein showed a substantially increased efficiency and speed on regeneration, also in recalcitrant genotypes (Debernardi et al., 2020). This was also useful in Cannabis, as overexpression of the developmental regulator chimera CsGRF3-CsGIF1 almost doubled the regeneration rate (Zhang et al., 2021).
Genome editing in Cannabis - challenges and possibilities
During the last 5-10 years there has been a lot of progress in the prerequisites for genome editing in medical Cannabis. There is now a reference genome for Cannabis and a lot of additional sequence data available. This is a very important basis for the development of sgRNAs for different genome editing techniques. Not only can sequence for candidate genes be found, but also analyses for potential off-targets can be made. The editing techniques are in rapid development with new techniques and improvements emerging every year. The biggest leap forward is the recent reports of regeneration and Agrobacterium-mediated transformation of Cannabis. There is, however, still challenges when it comes to using genome editing in medical Cannabis. The two main challenges are genotype dependency and the selection of transformable explant material from cuttings.
As seen in recent experiments, very young hemp tissue can, although maybe not easily, be transformed and regenerated into plants (Galán-Ávila et al., 2021b; Zhang et al., 2021). There is without doubt genotype dependency, not only between chemotypes but also between hemp cultivars. The use of CsGRF3-CsGIF1 in Cannabis and the use of a WUSCHEL family gene in wheat, T. monococcum, triticale, barley, and maize are examples where developmental regulators can overcome at least some of the genotype dependency (Zhang et al., 2021; Wang et al., 2022). It is our belief that the future will bring more improvements and refinements as recalcitrance to regeneration and genotype dependency is found in several plant species.
Using hypocotyls from immature embryos for transformation in medical Cannabis is not the obvious choice. The plants are usually clonal propagated to maintain valuable cannabinoid and terpene profiles, which will be changed if plants are propagated by seeds. Recently, promising results showing regeneration from drug type Cannabis using leaf segments and stem nodes have emerged (Önol and Yildirim, 2021; Hesami and Jones, 2021). Methods avoiding young material from seeds also include transformation on existing meristems (Galán-Ávila et al., 2021b) and the induction of new meristems from somatic cells (Maher et al., 2020). All these methods need to be thoroughly investigated in many genotypes of medical Cannabis. The possible outcome of the combination of floral reversion and transformation should also be pursued.
Due to the multiple applications of Cannabis, many traits might be interesting to improve. Single gene traits are easier to work with than very complex traits. However, successful editing of several genes is reported in other species (Gao et al., 2017; Mercx et al., 2017; Morineau et al., 2017) and might also be feasible in medical Cannabis as soon as a versatile transformation platform is established.
Clean cannabinoid products can be obtained by producing them in other systems like yeast (Luo et al., 2019). However, the therapeutic response is often higher using plant products, probably due to a synergistic or entourage effect between various cannabinoids or between cannabinoids and terpenes (Ferber et al., 2020). Genome editing tools would be very useful to study and manipulate the biosynthetic pathways of cannabinoids and terpenes, not only for pure scientific purposes, but also to improve the products in the medical Cannabis industry. Gene editing facilitates gene knockout studies as well as studies altering gene expression levels and tailoring of specific genes. The review by Hesami et al. (2022) lists several ideas for the use of CRISPR-based methods for optimizing cannabinoid production using in vitro culture and heterologous systems.
The THCA and CBDA synthases are of special interest, as they are responsible for the synthesis of the two main cannabinoids, THC acid and CBD acid from the common precursor, CBGA. The sequences of these synthases are very similar, and studies have shown that they are positioned at two closely linked loci in a very complex region (Van Bakel et al., 2011; Laverty et al., 2019; Vergara et al., 2019). This complexity makes it challenging, although not impossible, to make changes using genome editing, as guides need to be designed to hit target genes without off-target effects.
As THC is psychoactive, there is a demand for plants completely free of THC. It is essential that drug formulations contain as low a THC content as possible as the THC contamination causes short- and long-term side effects (Volkow et al., 2014). The use of extra purified CBD or synthetic CBD instead of the plant extract results in a lack of other naturally occurring cannabinoids and bioactive compounds such as terpenes involved in the entourage effect. This may have a decisive influence on the therapeutic effect of the product (Russo, 2019). However, Cannabis varieties bred for high CBD content always contain small amounts of THC.
The hemp cultivar Finola has no THCA synthase (Laverty et al., 2019), but still a small amount of THCA is produced (Pavlovic et al., 2019). This presence of THC in CBD-varieties is due to a promiscuous CBDA synthase producing up to 5% THCA (Zirpel et al., 2018). In yeast model systems, site-directed mutagenesis has been performed in THCAS and CBDAS to investigate the importance of amino acids for activity and specificity. One amino acid change at A414V increased the catalytic activity 3.3-fold and caused a shift in specificity profile from CBDA to THCA production (Zirpel et al., 2018). Further studies are needed to uncover the potential of modulating the specificity of this enzyme in planta. CRISPR technology would be the obvious choice for inducing base editing in the CBDA synthase and modulate enzyme activity towards making the synthase completely specific for CBDA production.
Another need is to produce plants with a higher content of rare cannabinoids or to change the content of different terpenes independent of the cannabinoid profile. CBG is a compound having medical properties of its own, but it is often present in quite low levels. Knocking out both THCA and CBDA synthases by genome editing would result in a plant with a higher level of CBG.
Genome editing approaches have been used to obtain resistance against plant diseases caused by viruses, fungi, and bacteria (Borrelli et al., 2018), as inactivation of susceptibility genes often gives resistance. This is highly relevant in medical Cannabis production as the loss of production batches due to attack by fungal diseases is a serious problem. The plants cannot be treated with fungicides as traces of these compounds might be found in the final product. Controlling the humidity is highly energy demanding, but if not done, production batches might be lost.
Amino acid changes in the Mlo gene(s) are known to give resistance against powdery mildew in a range of plants (Kusch and Panstruga, 2017). In medical Cannabis, where the use of pesticides is not possible, the growth of resistant plants is highly desirable. When medical Cannabis is produced in greenhouses, there is a high humidity, which is the perfect environment for the development of the fungal disease powdery mildew. CRISPR-mediated changes in the Mlo gene would give resistance against powdery mildew without any site effects in the cannabinoid production. Pleiotropic effects might occur when the Mlo gene is mutated (Jørgensen, 1992). Whereas some Mlo mutations seem to be without or with only minor pleiotropic effects, TALEN-derived mlo wheat plants with the triple knockout mutations show strong chlorotic symptoms (Jørgensen, 1992; Acevedo-Garcia et al., 2017). This suggests that carefully selected base editing might be preferred over CRISPR/Cas-mediated knockout.
Grey mold due to the fungus Botrytis cinerea is a severe problem in indoor Cannabis production. Botrytis is a necrotrophic fungal pathogen, but it apparently starts its infection in a biotrophic manner (Veloso and van Kan, 2018). Rather than indiscriminately killing its host, Botrytis gently guides the host plant towards committing suicide through apoptosis, making the fungus able to colonize and digest the plant tissue. If the spore density is low, the infected plants might be without symptoms. When these asymptomatic plants approach flowering, the fungus might switch to necrotrophic lifestyle causing the host plant to succumb (Veloso and van Kan, 2018). This might be the reason why growers often see a very sudden, severe attack of grey mold. Although the molecular mechanisms are not fully understood, silencing of susceptibility genes has been shown to impede infection (Sun et al., 2017). This and similar research might lead the way for (some) resistance against Botrytis obtained by genome editing.
There is a strong apical dominance in Cannabis (Smýkalová et al., 2019). Varieties with less apical dominance might be easier to regenerate in tissue culture and would therefore be the obvious candidates for transformation. Removal of apical buds or the application of plant growth regulators such as phytohormones change plant architecture, giving lower and more branched plants (Kocjan Ačko et al., 2019; Burgel et al., 2020). These more uniform plants make harvest of inflorescences easier. Although apical bud removal in hemp gives higher seed yield (Kocjan Ačko et al., 2019), it is unclear whether higher cannabinoid yield will be found in plants with a changed architecture due to genetic changes. However, plants treated with phytohormones showed the same or reduced inflorescence dry weight, dependent on genotype, with no change in CBD content (Burgel et al., 2020). Although drug type Cannabis, due to selection, has become shorter and more densely branched, there is a great demand for varieties with a standardized plant type, not least with same height and branching, as such plants are suitable for automation. Thus, genes involved in apical dominance, branching and plant height would be very interesting candidates for genome editing.
As mentioned in the doubled haploid section, haploid inducer lines can be produced using CRISPR/Cas to knock out the centromere-specific histone H3 (CENH3) (Kelliher et al., 2019). This would be a very useful technique to use in Cannabis, where no successful doubled haploid technique is yet available. With a robust transformation platform in Cannabis, haploid inducer lines obtained with this technique is surely possible. Haploid-inducer lines with a hemp genetic background would be useful in medical Cannabis as well, as the haploids will not contain any parental DNA from the haploid-inducer parent. Once established, the haploid-induces lines can carry a CRISPR/Cas cassette that would give edited haploid offspring without the editing machinery (Wang et al., 2019).
There are several other traits of general interest for Cannabis production, also in hemp cultivars. Flowering time, fiber quality, lower phytic acid content of seeds, and soil remediation properties are obvious candidates for investigation via genome editing (Shiels et al., 2022).
Conclusion
The very high genetic diversity in Cannabis sativa is a great advantage for conventional breeding. However, conventional breeding through crossing and selection is very time consuming. It requires several rounds of backcrossing and is complicated by the dioecious nature of the plants. Further, the introduction of new traits might compromise the cannabinoid and terpenoid profile of medical Cannabis. Targeted improvements at predetermined positions in the genome by gene editing might mitigate some of these challenges.
Cannabis has traditionally been considered a recalcitrant species, in which techniques like genetic transformation and genome editing were very complicated. However, our review providing insight into recent progresses within tissue culture, genetic transformation, gene editing and necessary sequence information on Cannabis sativa indicates that this is not the case anymore. Currently, plant regeneration has been reported not only in hemp but also in a few medical Cannabis cultivars. Regeneration improvements using developmental regulators has facilitated the first report on CRISPR/Cas9 mediated genome editing in hemp. Our study suggests that future efforts could be directed towards development of a robust regeneration protocol for differentiated or meristematic tissue from medical Cannabis and thereby form the basis for future targeted improvements of medical Cannabis.
Genome editing is a great tool for scientific investigations and precision breeding for secondary metabolites such as cannabinoids and terpenes. Expression of genes related to these compounds can be fine-tuned or completely removed by editing promoters and/or genes. Precision breeding further gives the possibilities to add new traits such as disease resistance or changed plant architecture without meddling with a known cannabinoid profile. Conventional methods adding new genetic variation will continue to be a corner stone in breeding but once transformation is a routine tool in Cannabis, there are almost unlimited possibilities to improve a wide range of plant traits. Furthermore, since medical cannabis is often grown under contained facilities, any GM regulatory requirements of genome edited plants will be easier to meet.
Author contributions
HB-P designed the project. CI wrote the initial manuscript which was carefully revised by HB-P. All authors contributed to the article and approved the submitted version.
Funding
HB-P was supported by NovoCrops, Grant Reference No. NNF19OC0056580.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1154332/full#supplementary-material
References
Aasim, M., Katırcı, R., Akgur, O., Yildirim, B., Mustafa, Z., Nadeem, M. A., et al. (2022). Machine learning (ML) algorithms and artificial neural network for optimizing in vitro germination and growth indices of industrial hemp (Cannabis sativa l.). Ind. Crops Prod. 181, 114801. doi: 10.1016/j.indcrop.2022.114801
Acevedo-Garcia, J., Spencer, D., Thieron, H., Reinstädler, A., Hammond-Kosack, K., Phillips, A. L., et al. (2017). Mlo-based powdery mildew resistance in hexaploid bread wheat generated by a non-transgenic TILLING approach. Plant Biotechnol. J. 15, 367–378. doi: 10.1111/pbi.12631
Adamek, K., Jones, A. M. P., Torkamaneh, D. (2022). Accumulation of somatic mutations leads to genetic mosaicism in cannabis. Plant Genome 15, e20169. doi: 10.1002/tpg2.20169
Adhikary, D., Kulkarni, M., El-Mezawy, A., Mobini, S., Elhiti, M., Gjuric, R., et al. (2021). Medical cannabis and industrial hemp tissue culture: present status and future potential. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.627240
Ahmed, S., Gao, X., Jahan, M. A., Adams, M., Wu, N., Kovinich, N. (2021). Nanoparticle-based genetic transformation of cannabis sativa. J. Biotechnol. 326, 48–51. doi: 10.1016/j.jbiotec.2020.12.014
Ainsworth, C. (2000). Boys and girls come out to play: the molecular biology of dioecious plants. Ann. Bot. 86, 211–221. doi: 10.1006/anbo.2000.1201
Altpeter, F., Springer, N. M., Bartley, L. E., Blechl, A. E., Brutnell, T. P., Citovsky, V., et al. (2016). Advancing crop transformation in the era of genome editing. Plant Cell 28, 1510–1520. doi: 10.1105/tpc.16.00196
Bagheri, M., Mansouri, H. (2015). Effect of induced polyploidy on some biochemical parameters in cannabis sativa l. Appl. Biochem. Biotechnol. 175, 2366–2375. doi: 10.1007/s12010-014-1435-8
Barcaccia, G., Palumbo, F., Scariolo, F., Vannozzi, A., Borin, M. (2020). Potentials and challenges of genomics for breeding cannabis cultivars. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.573299
Beard, K. M., Boling, A. W., Bargmann, B. O. (2021). Protoplast isolation, transient transformation, and flow-cytometric analysis of reporter-gene activation in cannabis sativa l. Ind. Crops Prod. 164, 113360. doi: 10.1016/j.indcrop.2021.113360
Bedini, S., Flamini, G., Cosci, F., Ascrizzi, R., Benelli, G., Conti, B. (2016). Cannabis sativa and humulus lupulus essential oils as novel control tools against the invasive mosquito aedes albopictus and fresh water snail physella acuta. Ind. Crops Prod. 85, 318–323. doi: 10.1016/j.indcrop.2016.03.008
Borbas, L. N., Kurtz, L. E., Lubell-Brand, J. D. (2023). A comparison of two media formulations and two vented culture vessels for shoot multiplication and rooting of hemp shoot tip cultures. HortTechnology 33, 233–238. doi: 10.21273/HORTTECH05179-22
Borrelli, V. M., Brambilla, V., Rogowsky, P., Marocco, A., Lanubile, A. (2018). The enhancement of plant disease resistance using CRISPR/Cas9 technology. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01245
Burgel, L., Hartung, J., Schibano, D., Graeff-Hönninger, S. (2020). Impact of different phytohormones on morphology, yield and cannabinoid content of cannabis sativa l. Plants 9, 725. doi: 10.3390/plants9060725
Cascini, F., Farcomeni, A., Migliorini, D., Baldassarri, L., Boschi, I., Martello, S., et al. (2019). Highly predictive genetic markers distinguish drug-type from fiber-type cannabis sativa l. Plants 8, 496. doi: 10.3390/plants8110496
Čermák, T., Curtin, S. J., Gil-Humanes, J., Čegan, R., Kono, T. J., Konečná, E., et al. (2017). A multipurpose toolkit to enable advanced genome engineering in plants. Plant Cell 29, 1196–1217. doi: 10.1105/tpc.16.00922
Chandra, S., Lata, H., ElSohly, M. A. (2020). Propagation of cannabis for clinical research: an approach towards a modern herbal medicinal products development. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00958
Chaohua, C., Gonggu, Z., Lining, Z., Chunsheng, G., Qing, T., Jianhua, C., et al. (2016). A rapid shoot regeneration protocol from the cotyledons of hemp (Cannabis sativa l.). Ind. Crops Prod. 83, 61–65. doi: 10.1016/j.indcrop.2015.12.035
Chen, K., Wang, Y., Zhang, R., Zhang, H., Gao, C. (2019). CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 70, 667–697. doi: 10.1146/annurev-arplant-050718-100049
Codesido, V., Meyer, S., Casano, S. (2020). Influence of media composition and genotype for successful Cannabis sativa L. in vitro introduction. Acta Hortic 1285, 75–80. doi: 10.17660/ActaHortic.2020.1285.12
Crane, C., Wright, E., Dixon, R. A., Wang, Z.-Y. (2006). Transgenic medicago truncatula plants obtained from agrobacterium tumefaciens-transformed roots and agrobacterium rhizogenes-transformed hairy roots. Planta 223, 1344–1354. doi: 10.1007/s00425-006-0268-2
Debernardi, J. M., Tricoli, D. M., Ercoli, M. F., Hayta, S., Ronald, P., Palatnik, J. F., et al. (2020). A GRF–GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nat. Biotechnol. 38, 1274–1279. doi: 10.1038/s41587-020-0703-0
Deguchi, M., Bogush, D., Weeden, H., Spuhler, Z., Potlakayala, S., Kondo, T., et al. (2020). Establishment and optimization of a hemp (Cannabis sativa l.) agroinfiltration system for gene expression and silencing studies. Sci. Rep. 10, 1–11. doi: 10.1038/s41598-020-60323-9
de la Torre, M. C. P., Fernández, P., Greppi, J. A., Coviella, M. A., Fernández, M. N., Astigueta, F., et al. (2018). Transformation of mecardonia (Plantaginaceae) with wild-type agrobacterium rhizogenes efficiently improves compact growth, branching and flower related ornamental traits. Sci. Hortic. 234, 300–311. doi: 10.1016/j.scienta.2018.02.047
de Oliveira Pereira Ribeiro, L., Avila, E., Mariot, R. F., Fett, M. S., de Oliveira Camargo, F. A., Alho, C. S. (2020). Evaluation of two 13-loci STR multiplex system regarding identification and origin discrimination of Brazilian cannabis sativa samples. Int. J. Legal Med. 134, 1603–1612. doi: 10.1007/s00414-020-02338-5
Dhawan, O., Lavania, U. (1996). Enhancing the productivity of secondary metabolites via induced polyploidy: a review. Euphytica 87, 81–89. doi: 10.1007/BF00021879
Divashuk, M. G., Alexandrov, O. S., Razumova, O. V., Kirov, I. V., Karlov, G. I. (2014). Molecular cytogenetic characterization of the dioecious cannabis sativa with an XY chromosome sex determination system. PloS One 9, e85118. doi: 10.1371/journal.pone.0085118
Downey, C. D., Golenia, G., Boudko, E. A., Jones, A. M. P. (2021). Cryopreservation of 13 commercial cannabis sativa genotypes using in vitro nodal explants. Plants 10, 1794. doi: 10.3390/plants10091794
Dreger, M., Szalata, M. (2021). The effect of TIBA and NPA on shoot regeneration of cannabis sativa l. epicotyl explants. Agronomy 12, 104. doi: 10.3390/agronomy12010104
Dufresnes, C., Jan, C., Bienert, F., Goudet, J., Fumagalli, L. (2017). Broad-scale genetic diversity of cannabis for forensic applications. PloS One 12, e0170522. doi: 10.1371/journal.pone.0170522
Dunwell, J. M., Wetten, A. C. (2012). (Eds.), Transgenic Plants. Methods and Protocols, Methods in Molecular Biology, vol. 847, (Humana Totowa, NJ).
ElSohly, M. A., Mehmedic, Z., Foster, S., Gon, C., Chandra, S., Church, J. C. (2016). Changes in cannabis potency over the last 2 decades(1995–2014): analysis of current data in the united states. Biol. Psychiatry 79, 613–619. doi: 10.1016/j.biopsych.2016.01.004
Farag, S., Kayser, O. (2015). Cannabinoids production by hairy root cultures of cannabis sativa l. Am. J. Plant Sci. 6, 1874–1884. doi: 10.4236/ajps.2015.611188
Faux, A.-M., Berhin, A., Dauguet, N., Bertin, P. (2014). Sex chromosomes and quantitative sex expression in monoecious hemp (Cannabis sativa l.). Euphytica 196, 183–197. doi: 10.1007/s10681-013-1023-y
Feeney, M., Punja, Z. (2003). Tissue culture and agrobacterium-mediated transformation of hemp (Cannabis sativa l.). In Vitro Cell. Dev. Biol. Plant 39, 578–585. doi: 10.1079/IVP2003454
Feeney, M., Punja, Z. K. (2017). “The role of agrobacterium-mediated and other gene-transfer technologies in cannabis research and product development,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 343–363. doi: 10.1007/978-3-319-54564-6_16
Feng, Z., Zhang, B., Ding, W., Liu, X., Yang, D.-L., Wei, P., et al. (2013). Efficient genome editing in plants using a CRISPR/Cas system. Cell Res. 23, 1229–1232. doi: 10.1038/cr.2013.114
Ferber, S. G., Namdar, D., Hen-Shoval, D., Eger, G., Koltai, H., Shoval, G., et al. (2020). The “entourage effect”: terpenes coupled with cannabinoids for the treatment of mood disorders and anxiety disorders. Curr. Neuropharmacol. 18, 87–96. doi: 10.2174/1570159X17666190903103923
Fischedick, J. T., Hazekamp, A., Erkelens, T., Choi, Y. H., Verpoorte, R. (2010). Metabolic fingerprinting of cannabis sativa l., cannabinoids and terpenoids for chemotaxonomic and drug standardization purposes. Phytochemistry 71, 2058–2073. doi: 10.1016/j.phytochem.2010.10.001
Flores-Sanchez, I. J., Peč, J., Fei, J., Choi, Y. H., Dušek, J., Verpoorte, R. (2009). Elicitation studies in cell suspension cultures of cannabis sativa l. J. Biotechnol. 143, 157–168. doi: 10.1016/j.jbiotec.2009.05.006
Flores-Sanchez, I. J., Verpoorte, R. (2008). Secondary metabolism in cannabis. Phytochem. Rev. 7, 615–639. doi: 10.1007/s11101-008-9094-4
Galán-Ávila, A., García-Fortea, E., Prohens, J., Herraiz, F. J. (2020). Development of a direct in vitro plant regeneration protocol from cannabis sativa l. seedling explants: developmental morphology of shoot regeneration and ploidy level of regenerated plants. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00645
Galán-Ávila, A., García-Fortea, E., Prohens, J., Herraiz, F. J. (2021a). Microgametophyte development in cannabis sativa l. and first androgenesis induction through microspore embryogenesis. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.669424
Galán-Ávila, A., Gramazio, P., Ron, M., Prohens, J., Herraiz, F. J. (2021b). A novel and rapid method for agrobacterium-mediated production of stably transformed cannabis sativa l. plants. Ind. Crops Prod. 170, 113691. doi: 10.1016/j.indcrop.2021.113691
Gao, W., Long, L., Tian, X., Xu, F., Liu, J., Singh, P. K., et al. (2017). Genome editing in cotton with the CRISPR/Cas9 system. Front. Plant Sci. 8 1364. doi: 10.3389/fpls.2017.01364
García-Pérez, P., Lozano-Milo, E., Landin, M., Gallego, P. P. (2020). Machine learning unmasked nutritional imbalances on the medicinal plant bryophyllum sp. cultured in vitro. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.576177
Gilchrist, E. J., Wang, S., Quilichini, T. D. (2023). “The impact of biotechnology and genomics on an ancient crop: Cannabis sativa,” in Genomics and the Global Bioeconomy (Cambridge, Massachusetts: Academic Press), 177–204. doi: 10.1016/B978-0-323-91601-1.00009-2
Grassa, C. J., Weiblen, G. D., Wenger, J. P., Dabney, C., Poplawski, S. G., Timothy Motley, S., et al. (2021). A new Cannabis genome assembly associates elevated cannabidiol (CBD) with hemp introgressed into marijuana. New Phytologist 230, 1665–1679. doi: 10.1111/nph.17243
Grassi, G., McPartland, J. M. (2017). “Chemical and morphological phenotypes in breeding of cannabis sativa l,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 137–160. doi: 10.1007/978-3-319-54564-6_6
Grotenhermen, F. (2003). Clinical pharmacokinetics of cannabinoids. J. Cannabis Ther. 3, 3–51. doi: 10.2165/00003088-200342040-00003
Grulichova, M., Mendel, P., Lalge, A. B., Slamova, N., Trojan, V., Vyhnanek, T., et al. (2017). Effect of different phytohormones on growth and development of micropropagated cannabis sativa l. Mendel Net 2017, 618–623.
Gupta, D., Bhattacharjee, O., Mandal, D., Sen, M. K., Dey, D., Dasgupta, A., et al. (2019). CRISPR-Cas9 system: a new-fangled dawn in gene editing. Life Sci. 232, 116636. doi: 10.1016/j.lfs.2019.116636
Hesami, M., Adamek, K., Pepe, M., Jones, A. M. P. (2023). Effect of explant source on phenotypic changes of In vitro grown cannabis plantlets over multiple subcultures. Biology 12, 443. doi: 10.3390/biology12030443
Hesami, M., Baiton, A., Alizadeh, M., Pepe, M., Torkamaneh, D., Jones, A. M. P. (2021a). Advances and perspectives in tissue culture and genetic engineering of cannabis. Int. J. Mol. Sci. 22, 5671. doi: 10.3390/ijms22115671
Hesami, M., Jones, A. M. P. (2021). Modeling and optimizing callus growth and development in cannabis sativa using random forest and support vector machine in combination with a genetic algorithm. Appl. Microbiol. 105, 5201–5212. doi: 10.1007/s00253-021-11375-y
Hesami, M., Pepe, M., Alizadeh, M., Rakei, A., Baiton, A., Jones, A. M. P. (2020). Recent advances in cannabis biotechnology. Ind. Crops Prod. 158, 113026. doi: 10.1016/j.indcrop.2020.113026
Hesami, M., Pepe, M., Baiton, A., Jones, A. M. P. (2022). Current status and future prospects in cannabinoid production through in vitro culture and synthetic biology. Biotechnol. Adv. 108074. doi: 10.1016/j.biotechadv.2022.108074
Hesami, M., Pepe, M., Monthony, A. S., Baiton, A., Jones, A. M. P. (2021b). Modeling and optimizing in vitro seed germination of industrial hemp (Cannabis sativa l.). Ind. Crops Prod. 170, 113753. doi: 10.1016/j.indcrop.2021.113753
Holme, I. B., Wendt, T., Gil-Humanes, J., Deleuran, L. C., Starker, C. G., Voytas, D. F., et al. (2017). Evaluation of the mature grain phytase candidate HvPAPhy_a gene in barley (Hordeum vulgare l.) using CRISPR/Cas9 and TALENs. Plant Mol. Biol. 95, 111–121. doi: 10.1007/s11103-017-0640-6
Holmes, J. E., Lung, S., Collyer, D., Punja, Z. K. (2021). Variables affecting shoot growth and plantlet recovery in tissue cultures of drug-type cannabis sativa l. Front. Plant Sci. 12, 732344. doi: 10.3389/fpls.2021.732344
Hooghvorst, I., Nogués, S. (2021). Chromosome doubling methods in doubled haploid and haploid inducer-mediated genome-editing systems in major crops. Plant Cell Rep. 40, 255–270. doi: 10.1007/s00299-020-02605-0
Houston, R., Birck, M., Hughes-Stamm, S., Gangitano, D. (2017). Developmental and internal validation of a novel 13 loci STR multiplex method for cannabis sativa DNA profiling. Leg. Med. 26, 33–40. doi: 10.1016/j.legalmed.2017.03.001
Houston, R., Birck, M., LaRue, B., Hughes-Stamm, S., Gangitano, D. (2018). Nuclear, chloroplast, and mitochondrial data of a US cannabis DNA database. Int. J. Legal Med. 132, 713–725. doi: 10.1007/s00414-018-1798-4
Hua, K., Tao, X., Liang, W., Zhang, Z., Gou, R., Zhu, J. K. (2020). Simplified adenine base editors improve adenine base editing efficiency in rice. Plant Biotechnol. J. 18, 770–778. doi: 10.1111/pbi.13244
Hurgobin, B., Tamiru-Oli, M., Welling, M. T., Doblin, M. S., Bacic, A., Whelan, J., et al. (2021). Recent advances in cannabis sativa genomics research. New Phytol. 230, 73–89. doi: 10.1111/nph.17140
Iannicelli, J., Guariniello, J., Tossi, V. E., Regalado, J. J., Di Ciaccio, L., Van Baren, C. M., et al. (2020). The “polyploid effect“ in the breeding of aromatic and medicinal species. Sci. Hortic. 260, 108854. doi: 10.1016/j.scienta.2019.108854
Ioannidis, K., Dadiotis, E., Mitsis, V., Melliou, E., Magiatis, P. (2020). Biotechnological approaches on two high CBD and CBG cannabis sativa l. (Cannabaceae) varieties: In vitro regeneration and phytochemical consistency evaluation of micropropagated plants using quantitative 1H-NMR. Molecules 25, 5928. doi: 10.3390/molecules25245928
Ioannidis, K., Tomprou, I., Mitsis, V. (2022a). An alternative In vitro propagation protocol of cannabis sativa L.(Cannabaceae) presenting efficient rooting, for commercial production. Plants 11, 1333. doi: 10.3390/plants11101333
Ioannidis, K., Tomprou, I., Mitsis, V., Koropouli, P. (2022b). Genetic evaluation of in vitro micropropagated and regenerated plants of cannabis sativa l. using SSR molecular markers. Plants 11, 2569. doi: 10.3390/plants11192569
Ishii, T., Karimi-Ashtiyani, R., Houben, A. (2016). Haploidization via chromosome elimination: means and mechanisms. Annu. Rev. Plant Biol. 67, 421–438. doi: 10.1146/annurev-arplant-043014-114714
Jaganathan, D., Ramasamy, K., Sellamuthu, G., Jayabalan, S., Venkataraman, G. (2018). CRISPR for crop improvement: an update review. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00985
Jørgensen, I. H. (1992). Discovery, characterization and exploitation of mlo powdery mildew resistance in barley. Euphytica 63, 141–152. doi: 10.1007/BF00023919
Kang, B.-C., Bae, S.-J., Lee, S., Lee, J. S., Kim, A., Lee, H., et al. (2021). Chloroplast and mitochondrial DNA editing in plants. Nat. Plants 7, 899–905. doi: 10.1038/s41477-021-00943-9
Kelliher, T., Starr, D., Su, X., Tang, G., Chen, Z., Carter, J., et al. (2019). One-step genome editing of elite crop germplasm during haploid induction. Nat. Biotechnol. 37, 287–292. doi: 10.1038/s41587-019-0038-x
Khan, Z., Khan, S. H., Mubarik, M. S., Sadia, B., Ahmad, A. (2017). Use of TALEs and TALEN technology for genetic improvement of plants. Plant Mol. Biol. Rep. 35, 1–19. doi: 10.1007/s11105-016-0997-8
Kim, A. L., Yun, Y. J., Choi, H. W., Hong, C.-H., Shim, H. J., Lee, J. H., et al. (2022). Establishment of efficient cannabis (Cannabis sativa l.) protoplast isolation and transient expression condition. Plant Biotechnol. Rep. 16, 613–619. doi: 10.1007/s11816-022-00786-6
Kocjan Ačko, D., Flajšman, M., Trdan, S. (2019). Apical bud removal increased seed yield in hemp (Cannabis sativa l.). Acta Agric. Scand. B Soil Plant Sci. 69, 317–323. doi: 10.1080/09064710.2019.1568540
Kojoma, M., Seki, H., Yoshida, S., Muranaka, T. (2006). DNA Polymorphisms in the tetrahydrocannabinolic acid (THCA) synthase gene in “drug-type” and “fiber-type” cannabis sativa l. Forensic Sci. Int. 159, 132–140. doi: 10.1016/j.forsciint.2005.07.005
Kovalchuk, I., Pellino, M., Rigault, P., Van Velzen, R., Ebersbach, J., Ashnest, J., et al. (2020). The genomics of cannabis and its close relatives. Annu. Rev. Plant Biol. 71, 713–739. doi: 10.1146/annurev-arplant-081519-040203
Krenek, P., Samajova, O., Luptovciak, I., Doskocilova, A., Komis, G., Samaj, J. (2015). Transient plant transformation mediated by agrobacterium tumefaciens: principles, methods and applications. Biotechnol. 33, 1024–1042. doi: 10.1016/j.biotechadv.2015.03.012
Kumar, K., Gambhir, G., Dass, A., Tripathi, A. K., Singh, A., Jha, A. K., et al. (2020). Genetically modified crops: current status and future prospects. Planta 251, 1–27. doi: 10.1007/s00425-020-03372-8
Kurtz, L. E., Borbas, L. N., Brand, M. H., Lubell-Brand, J. D. (2022). Ex vitro rooting of cannabis sativa microcuttings and their performance compared to retip and stem cuttings. HortScience 57, 1576–1579. doi: 10.21273/HORTSCI16890-22
Kusch, S., Panstruga, R. (2017). Mlo-based resistance: an apparently universal “weapon” to defeat powdery mildew disease. Mol. Plant-Microbe Interac. 30, 179–189. doi: 10.1094/MPMI-12-16-0255-CR
Lata, H., Chandra, S., Khan, I., ElSohly, M. A. (2009a). Thidiazuron-induced high-frequency direct shoot organogenesis of. In Vitro Cell. Dev. Biol. Plant 45, 12–19. doi: 10.1007/s11627-008-9167-5
Lata, H., Chandra, S., Khan, I. A., ElSohly, M. A. (2009b). Propagation through alginate encapsulation of axillary buds of cannabis sativa l. - an important medicinal plant. Physiol. Mol. Biol. Plants 15, 79–86. doi: 10.1007/s12298-009-0008-8
Lata, H., Chandra, S., Khan, I. A., ElSohly, M. A. (2010). High frequency plant regeneration from leaf derived callus of high Δ9-tetrahydrocannabinol yielding cannabis sativa l. Planta Med. 76, 1629–1633. doi: 10.1055/s-0030-1249773
Lata, H., Chandra, S., Mehmedic, Z., Khan, I. A., ElSohly, M. A. (2012). In vitro germplasm conservation of high Δ 9-tetrahydrocannabinol yielding elite clones of cannabis sativa l. under slow growth conditions. Acta Physiol. Plant 34, 743–750. doi: 10.1007/s11738-011-0874-x
Lata, H., Chandra, S., Techen, N., Khan, I. A., ElSohly, M. A. (2016). In vitro mass propagation of cannabis sativa l.: a protocol refinement using novel aromatic cytokinin meta-topolin and the assessment of eco-physiological, biochemical and genetic fidelity of micropropagated plants. J. Appl. Res. Med. Aromat. Plants 3, 18–26. doi: 10.1016/j.jarmap.2015.12.001
Lata, H., Uchendu, E., Chandra, S., Majumdar, C. G., Khan, I. A., ElSohly, M. A. (2019). Cryopreservation of axillary buds of cannabis sativa l. by V-cryoplate droplet-vitrification: the critical role of sucrose preculture. Cryoletters 40, 291–298.
Laverty, K. U., Stout, J. M., Sullivan, M. J., Shah, H., Gill, N., Holbrook, L., et al. (2019). A physical and genetic map of cannabis sativa identifies extensive rearrangements at the THC/CBD acid synthase loci. Genome Res. 29, 146–156. doi: 10.1101/gr.242594.118
Li, H.-L. (1974). An archaeological and historical account of cannabis in China. Econ. Bot. 28, 437–448. doi: 10.1007/BF02862859
Li, J.-F., Norville, J. E., Aach, J., McCormack, M., Zhang, D., Bush, J., et al. (2013). Multiplex and homologous recombination–mediated genome editing in arabidopsis and nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 31, 688–691. doi: 10.1038/nbt.2654
Li, C., Zong, Y., Jin, S., Zhu, H., Lin, D., Li, S., et al. (2020). SWISS: multiplexed orthogonal genome editing in plants with a Cas9 nickase and engineered CRISPR RNA scaffolds. Genome Biol. 21, 1–15. doi: 10.1186/s13059-020-02051-x
Lin, Q., Zong, Y., Xue, C., Wang, S., Jin, S., Zhu, Z., et al. (2020). Prime genome editing in rice and wheat. Nat. Biotechnol. 38, 582–585. doi: 10.1038/s41587-020-0455-x
Livingston, S. J., Quilichini, T. D., Booth, J. K., Wong, D. C., Rensing, K. H., Laflamme-Yonkman, J., et al. (2020). Cannabis glandular trichomes alter morphology and metabolite content during flower maturation. Plant J. 101, 37–56. doi: 10.1111/tpj.14516
Lowder, L., Malzahn, A., Qi, Y. (2016). Rapid evolution of manifold CRISPR systems for plant genome editing. Front. Plant Sci. 7, 1683. doi: 10.3389/fpls.2016.01683
Lu, Y., Wang, J., Chen, B., Mo, S., Lian, L., Luo, Y., et al. (2021). A donor-DNA-free CRISPR/Cas-based approach to gene knock-up in rice. Nat. Plants 7, 1445–1452. doi: 10.1038/s41477-021-01019-4
Lubell, J. D., Brand, M. H. (2018). Foliar sprays of silver thiosulfate produce male flowers on female hemp plants. HortTechnology 28, 743–747. doi: 10.21273/HORTTECH04188-18
Lubell-Brand, J. D., Kurtz, L. E., Brand, M. H. (2021). An in vitro–ex vitro micropropagation system for hemp. HortTechnology 31, 199–207. doi: 10.21273/HORTTECH04779-20
Luo, X., Reiter, M. A., d’Espaux, L., Wong, J., Denby, C. M., Lechner, A., et al. (2019). Complete biosynthesis of cannabinoids and their unnatural analogues in yeast. Nature 567, 123–126. doi: 10.1038/s41586-019-0978-9
Lusser, M., Parisi, C., Plan, D., Rodríguez-Cerezo, E. (2012). Deployment of new biotechnologies in plant breeding. Nat. Biotechnol. 30, 231–239. doi: 10.1038/nbt.2142
Lütken, H., Clarke, J. L., Müller, R. (2012). Genetic engineering and sustainable production of ornamentals: current status and future directions. Plant Cell Rep. 31, 1141–1157. doi: 10.1007/s00299-012-1265-5
Lynch, R. C., Vergara, D., Tittes, S., White, K., Schwartz, C., Gibbs, M. J., et al. (2016). Genomic and chemical diversity in cannabis. CRC Crit. Rev. Plant Sci. 35, 349–363. doi: 10.1080/07352689.2016.1265363
Ma, X., Zhu, Q., Chen, Y., Liu, Y.-G. (2016). CRISPR/Cas9 platforms for genome editing in plants: developments and applications. Mol. Plant 9, 961–974. doi: 10.1016/j.molp.2016.04.009
MacKinnon, L., McDougall, G., Aziz, N., Millam, S. (2001). Progress towards transformation of fibre hemp. Annu. Rep. Scottish Crop Res. Institute 2000/2001, 84–86.
Maher, M. F., Nasti, R. A., Vollbrecht, M., Starker, C. G., Clark, M. D., Voytas, D. F. (2020). Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 38, 84–89. doi: 10.1038/s41587-019-0337-2
Mandolino, G., Carboni, A., Bagatta, M., Moliterni, V., Ranalli, P. (2002). Occurrence and frequency of putatively y chromosome linked DNA markers in cannabis sativa l. Euphytica 126, 211–218. doi: 10.1023/A:1016382128401
Mandolino, G., Carboni, A., Forapani, S., Faeti, V., Ranalli, P. (1999). Identification of DNA markers linked to the male sex in dioecious hemp (Cannabis sativa l.). Theor. Appl. Genet. 98, 86–92. doi: 10.1007/s001220051043
Mansouri, H., Bagheri, M. (2017). “Induction of polyploidy and its effect on cannabis sativa l,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 365–383. doi: 10.1007/978-3-319-54564-6_17
Matchett-Oates, L., Mohamaden, E., Spangenberg, G., Cogan, N. (2021a). Development of a robust transient expression screening system in protoplasts of cannabis. In Vitro Cell. Dev. Biol. Plant 57, 1040–1050. doi: 10.1007/s11627-021-10178-0
Matchett-Oates, L., Spangenberg, G. C., Cogan, N. O. (2021b). Manipulation of cannabinoid biosynthesis via transient RNAi expression. Front Plant Sci. 2766. doi: 10.3389/fpls.2021.773474
McPartland, J. (1984). Pathogenicity of phomopsis ganjae on cannabis sativa and the fungistatic effect of cannabinoids produced by the host. Mycopathologia 87, 149–153. doi: 10.1007/BF00436900
McPartland, J. M. (2017). “Cannabis sativa and cannabis indica versus “Sativa“ and “Indica,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 101–121. doi: 10.1007/978-3-319-54564-6_4
Mehmedic, Z., Chandra, S., Slade, D., Denham, H., Foster, S., Patel, A. S., et al. (2010). Potency trends of Δ9-THC and other cannabinoids in confiscated cannabis preparations from 1993 to 2008. J. Forensic Sci. 55, 1209–1217. doi: 10.1111/j.1556-4029.2010.01441.x
Mercx, S., Smargiasso, N., Chaumont, F., De Pauw, E., Boutry, M., Navarre, C. (2017). Inactivation of the β (1, 2)-xylosyltransferase and the α (1, 3)-fucosyltransferase genes in nicotiana tabacum BY-2 cells by a multiplex CRISPR/Cas9 strategy results in glycoproteins without plant-specific glycans. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00403
Mestinšek-Mubi, Š., Svetik, S., Flajšman, M., Murovec, J. (2020). In vitro tissue culture and genetic analysis of two high-CBD medical cannabis (Cannabis sativa l.) breeding lines. Genetika 52, 925–941. doi: 10.2298/GENSR2003925M
Mithöfer, A., Boland, W. (2012). Plant defense against herbivores: chemical aspects. Annu. Rev. Plant Biol. 63, 431–450. doi: 10.1146/annurev-arplant-042110-103854
Montecillo, J. A. V., Chu, L. L., Bae, H. (2020). CRISPR-Cas9 system for plant genome editing: current approaches and emerging developments. Agron. 10, 1033. doi: 10.3390/agronomy10071033
Monthony, A., Bagheri, S., Zheng, Y., Jones, A. (2021a). Flower power: floral reversion as a viable alternative to nodal micropropagation in cannabis sativa. In Vitro Cell. Dev. Biol. Plant 57, 1018–1030. doi: 10.1007/s11627-021-10181-5
Monthony, A. S., Kyne, S. T., Grainger, C. M., Jones, A. M. P. (2021b). Recalcitrance of cannabis sativa to de novo regeneration; a multi-genotype replication study. PloS One 16, e0235525. doi: 10.1371/journal.pone.0235525
Monthony, A. S., Page, S. R., Hesami, M., Jones, A. M. P. (2021c). The past, present and future of cannabis sativa tissue culture. Plants 10, 185. doi: 10.3390/plants10010185
Morineau, C., Bellec, Y., Tellier, F., Gissot, L., Kelemen, Z., Nogué, F., et al. (2017). Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid camelina sativa. Plant Biotechnol. J. 15, 729–739. doi: 10.1111/pbi.12671
Movahedi, M., Ghasemi-Omran, V., Torabi, S. (2015). The effect of different concentrations of TDZ and BA on in vitro regeneration of Iranian cannabis (Cannabis sativa) using cotyledon and epicotyl explants. J. Plant Mol. Breed. 3, 20–27. doi: 10.22058/jpmb.2015.15371
Mukhtar, T., Kayani, M. Z., Hussain, M. A. (2013). Nematicidal activities of cannabis sativa l. and zanthoxylum alatum roxb. against meloidogyne incognita. Ind. Crops Prod. 42, 447–453. doi: 10.1016/j.indcrop.2012.06.027
Murphy, R., Adelberg, J. (2021). Physical factors increased quantity and quality of micropropagated shoots of cannabis sativa l. in a repeated harvest system with ex vitro rooting. In Vitro Cell. Dev. Biol. Plant 57, 923–931. doi: 10.1007/s11627-021-10166-4
Naim, F., Shand, K., Hayashi, S., O’Brien, M., McGree, J., Johnson, A. A., et al. (2020). Are the current gRNA ranking prediction algorithms useful for genome editing in plants? PloS One 15, e0227994. doi: 10.1371/journal.pone.0227994
Nekrasov, V., Staskawicz, B., Weigel, D., Jones, J. D., Kamoun, S. (2013). Targeted mutagenesis in the model plant nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 31, 691–693. doi: 10.1038/nbt.2655
Niazian, M., Nalousi, A. M. (2020). Artificial polyploidy induction for improvement of ornamental and medicinal plants. Plant Cell Tissue Organ Cult. 142, 447–469. doi: 10.1007/s11240-020-01888-1
Oh, H., Seo, B., Lee, S., Ahn, D.-H., Jo, E., Park, J.-K., et al. (2016). Two complete chloroplast genome sequences of cannabis sativa varieties. Mitochondrial DNA Part A 27, 2835–2837. doi: 10.3109/19401736.2015.1053117
Onofri, C., Mandolino, G. (2017). “Genomics and molecular markers in cannabis sativa l,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 319–342. doi: 10.1007/978-3-319-54564-6_15
Önol, B., Yildirim, M. U. (2021). Breaking seed dormancy and regeneration in cannabis sativa l. Int. J. Agric. Environ. Food Sci. 5, 709–719. doi: 10.31015/jaefs.2021.4.32
Page, S. R., Monthony, A. S., Jones, A. M. P. (2021). DKW basal salts improve micropropagation and callogenesis compared with MS basal salts in multiple commercial cultivars of cannabis sativa. Botany 99, 269–279. doi: 10.1139/cjb-2020-0179
Pandey, K., Karthik, K., Singh, S. K., Vinod, Sreevathsa, R., Srivastav, M. (2022). Amenability of an agrobacterium tumefaciens-mediated shoot apical meristem-targeted in planta transformation strategy in mango (Mangifera indica l.). GM Crops Food 13, 342–354. doi: 10.1080/21645698.2022.2141014
Parsons, J. L., Martin, S. L., James, T., Golenia, G., Boudko, E. A., Hepworth, S. R. (2019). Polyploidization for the genetic improvement of cannabis sativa. Front. Plant Sci. 476. doi: 10.3389/fpls.2019.00476
Pauwels, L., De Clercq, R., Goossens, J., Iñigo, S., Williams, C., Ron, M., et al. (2018). A dual sgRNA approach for functional genomics in arabidopsis thaliana. G3-Genes Genom. Genet. 8, 2603–2615. doi: 10.1534/g3.118.200046
Pavlovic, R., Panseri, S., Giupponi, L., Leoni, V., Citti, C., Cattaneo, C., et al. (2019). Phytochemical and ecological analysis of two varieties of hemp (Cannabis sativa l.) grown in a mountain environment of Italian Alps. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01265
Pepe, M., Hesami, M., Jones, A. M. P. (2021a). Machine learning-mediated development and optimization of disinfection protocol and scarification method for improved in vitro germination of cannabis seeds. Plants 10, 2397. doi: 10.3390/plants10112397
Pepe, M., Hesami, M., Small, F., Jones, A. M. P. (2021b). Comparative analysis of machine learning and evolutionary optimization algorithms for precision micropropagation of cannabis sativa: prediction and validation of in vitro shoot growth and development based on the optimization of light and carbohydrate sources. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.757869
Pepe, M., Leonardos, E. D., Marie, T. R., Kyne, S. T., Hesami, M., Jones, A. M. P., et al. (2022). A noninvasive gas exchange method to test and model photosynthetic proficiency and growth rates of In vitro plant cultures: preliminary implication for cannabis sativa l. Biology 11, 729. doi: 10.3390/biology11050729
Pertwee, R. G. (1988). The central neuropharmacology of psychotropic cannabinoids. Pharmacol. Ther. 36, 189–261. doi: 10.1016/0163-7258(88)90106-4
Petolino, J. F. (2015). Genome editing in plants via designed zinc finger nucleases. In Vitro Cell. Dev. Biol. Plant 51, 1–8. doi: 10.1007/s11627-015-9663-3
Pisanti, S., Malfitano, A. M., Ciaglia, E., Lamberti, A., Ranieri, R., Cuomo, G., et al. (2017). Cannabidiol: state of the art and new challenges for therapeutic applications. Pharmacol. Ther. 175, 133–150. doi: 10.1016/j.pharmthera.2017.02.041
Piunno, K. F., Golenia, G., Boudko, E. A., Downey, C., Jones, A. M. P. (2019). Regeneration of shoots from immature and mature inflorescences of cannabis sativa. Can. J. Plant Sci. 99, 556–559. doi: 10.1139/cjps-2018-0308
Punja, Z. K., Holmes, J. E. (2020). Hermaphroditism in marijuana (Cannabis sativa l.) inflorescences–impact on floral morphology, seed formation, progeny sex ratios, and genetic variation. Front. Plant Sci. 718. doi: 10.3389/fpls.2020.00718
Punja, Z. K., Rodriguez, G., Chen, S. (2017). “Assessing genetic diversity in cannabis sativa using molecular approaches,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 395–418. doi: 10.1007/978-3-319-54564-6_19
Radošević, A., Kupinić, M., Grlić, L. (1962). Antibiotic activity of various types of cannabis resin. Nature 195, 1007–1009. doi: 10.1038/1951007a0
Radwan, M. M., Wanas, A. S., Chandra, S., ElSohly, M. A. (2017). “Natural cannabinoids of cannabis and methods of analysis,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 161–182. doi: 10.1007/978-3-319-54564-6_7
Razumova, O. V., Alexandrov, O. S., Divashuk, M. G., Sukhorada, T. I., Karlov, G. I. (2016). Molecular cytogenetic analysis of monoecious hemp (Cannabis sativa l.) cultivars reveals its karyotype variations and sex chromosomes constitution. Protoplasma 253, 895–901. doi: 10.1007/s00709-015-0851-0
Reimann-Philipp, U., Speck, M., Orser, C., Johnson, S., Hilyard, A., Turner, H., et al. (2020). Cannabis chemovar nomenclature misrepresents chemical and genetic diversity; survey of variations in chemical profiles and genetic markers in Nevada medical cannabis samples. Cannabis Cannabinoid Res. 5, 215–230. doi: 10.1089/can.2018.0063
Ren, Q., Sretenovic, S., Liu, S., Tang, X., Huang, L., He, Y., et al. (2021). PAM-less plant genome editing using a CRISPR–SpRY toolbox. Nat. Plants 7, 25–33. doi: 10.1038/s41477-020-00827-4
Rico, S., Garrido, J., Sánchez, C., Ferreiro-Vera, C., Codesido, V., Vidal, N. (2022). A temporary immersion system to improve cannabis sativa micropropagation. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.895971
Rodríguez-Leal, D., Lemmon, Z. H., Man, J., Bartlett, M. E., Lippman, Z. B. (2017). Engineering quantitative trait variation for crop improvement by genome editing. Cell 171, 470–480.e478. doi: 10.1016/j.cell.2017.08.030
Roscow JR., R. F. (2017). Enhanced cannabis plants and methods of making and using the same. Patent with International Publication Number WO 2017/1810180 A1 (Geneva, Switzerland: World Intellectual Property Organization).
Rotherham, D., Harbison, S. (2011). Differentiation of drug and non-drug cannabis using a single nucleotide polymorphism (SNP) assay. Forensic Sci. Int. 207, 193–197. doi: 10.1016/j.forsciint.2010.10.006
Russo, E. B. (2019). The case for the entourage effect and conventional breeding of clinical cannabis: no “strain,“ no gain. Front. Plant Sci. 1969. doi: 10.3389/fpls.2018.01969
Sattler, M. C., Carvalho, C. R., Clarindo, W. R. (2016). The polyploidy and its key role in plant breeding. Planta 243, 281–296. doi: 10.1007/s00425-015-2450-x
Sawler, J., Stout, J. M., Gardner, K. M., Hudson, D., Vidmar, J., Butler, L., et al. (2015). The genetic structure of marijuana and hemp. PloS One 10 (8), e0133292. doi: 10.1371/journal.pone.0133292
Schachtsiek, J., Hussain, T., Azzouhri, K., Kayser, O., Stehle, F. (2019). Virus-induced gene silencing (VIGS) in. Plant Methods 15, 1–9. doi: 10.1186/s13007-019-0542-5
Schaffner, J. H. (1921). Influence of environment on sexual expression in hemp. Bot. Gaz. 71, 197–219. doi: 10.1086/332818
Shan, Q., Wang, Y., Li, J., Zhang, Y., Chen, K., Liang, Z., et al. (2013). Targeted genome modification of crop plants using a CRISPR-cas system. Nat. Biotechnol. 31, 686–688. doi: 10.1038/nbt.2650
Shiels, D., Prestwich, B. D., Koo, O., Kanchiswamy, C. N., O'Halloran, R., Badmi, R. (2022). Hemp genome editing–challenges and opportunities. Front. Genome Ed. 4. doi: 10.3389/fgeed.2022.823486
Si, X., Zhang, H., Wang, Y., Chen, K., Gao, C. (2020). Manipulating gene translation in plants by CRISPR–Cas9-mediated genome editing of upstream open reading frames. Nat. Protoc. 15, 338–363. doi: 10.1038/s41596-019-0238-3
Simiyu, D. C., Jang, J. H., Lee, O. R. (2022). Understanding cannabis sativa l.: current status of propagation, use, legalization, and haploid-inducer-mediated genetic engineering. Plants 11, 1236. doi: 10.3390/plants11091236
Sirikantaramas, S., Taura, F., Tanaka, Y., Ishikawa, Y., Morimoto, S., Shoyama, Y. (2005). Tetrahydrocannabinolic acid synthase, the enzyme controlling marijuana psychoactivity, is secreted into the storage cavity of the glandular trichomes. Plant Cell Physiol. 46, 1578–1582. doi: 10.1093/pcp/pci166
Sirkowski, E. E. (2012). Marked Cannabis for indicating medical marijuana. United States Patent Application, Pub. No.: US 2012/0311744 A1. (United States Patent and Trademark Office).
Slusarkiewicz-Jarzina, A., Ponitka, A., Kaczmarek, Z. (2005). Influence of cultivar, explant source and plant growth regulator on callus induction and plant regeneration of cannabis sativa l. Acta Biol. Crac. Ser. Bot. 47, 145–151.
Small, E. (2017). “Classification of cannabis sativa l. @ in relation to agricultural, biotechnological, medical and recreational utilization,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 1–62. doi: 10.1007/978-3-319-54564-6_1
Small, E., Cronquist, A. (1976). A practical and natural taxonomy for cannabis. Taxon 25, 405–435. doi: 10.2307/1220524
Smýkalová, I., Vrbová, M., Cvečková, M., Plačková, L., Žukauskaitė, A., Zatloukal, M., et al. (2019). The effects of novel synthetic cytokinin derivatives and endogenous cytokinins on the in vitro growth responses of hemp (Cannabis sativa l.) explants. Plant Cell Tissue Organ Cult. 139, 381–394. doi: 10.1007/s11240-019-01693-5
Soler, S., Borràs, D., Vilanova, S., Sifres, A., Andújar, I., Figàs, M. R., et al. (2016). Use of embryos extracted from individual cannabis sativa seeds for genetic studies and forensic applications. J. Forensic Sci. 61, 494–500. doi: 10.1111/1556-4029.12995
Soler, S., Gramazio, P., Figàs, M. R., Vilanova, S., Rosa, E., Llosa, E. R., et al. (2017). Genetic structure of cannabis sativa var. indica cultivars based on genomic SSR (gSSR) markers: implications for breeding and germplasm management. Ind. Crops Prod. 104, 171–178. doi: 10.1016/j.indcrop.2017.04.043
Sorokin, A., Yadav, N. S., Gaudet, D., Kovalchuk, I. (2020). Transient expression of the β-glucuronidase gene in cannabis sativa varieties. Plant Signal. Beh. 15 (8), 1780037. doi: 10.1080/15592324.2020.1780037
Srivastava, S., Srivastava, A. K. (2007). Hairy root culture for mass-production of high-value secondary metabolites. Crit. Rev. Biotechnol. 27, 29–43. doi: 10.1080/07388550601173918
Stephen, C., Zayas, V. A., Galic, A., Bridgen, M. P. (2023). Micropropagation of hemp (Cannabis sativa l.). HortScience 58, 307–316. doi: 10.21273/HORTSCI16969-22
Sugimoto, K., Temman, H., Kadokura, S., Matsunaga, S. (2019). To regenerate or not to regenerate: factors that drive plant regeneration. Curr. Opin. Plant Biol. 47, 138–150. doi: 10.1016/j.pbi.2018.12.002
Sun, K., van Tuinen, A., van Kan, J. A., Wolters, A.-M. A., Jacobsen, E., Visser, R. G., et al. (2017). Silencing of DND1 in potato and tomato impedes conidial germination, attachment and hyphal growth of botrytis cinerea. BMC Plant Biol. 17, 1–12. doi: 10.1186/s12870-017-1184-2
Swarts, D. C., Jinek, M. (2018). Cas9 versus Cas12a/Cpf1: structure–function comparisons and implications for genome editing. Wiley Interdiscip. Rev. RNA 9, e1481. doi: 10.1002/wrna.1481
Taura, F., Sirikantaramas, S., Shoyama, Y., Shoyama, Y., Morimoto, S. (2007). Phytocannabinoids in cannabis sativa: recent studies on biosynthetic enzymes. Chem. Biodivers. 4, 1649–1663. doi: 10.1002/cbdv.200790145
Törjék, O., Bucherna, N., Kiss, E., Homoki, H., Finta-Korpelová, Z., Bócsa, I., et al. (2002). Novel male-specific molecular markers (MADC5, MADC6) in hemp. Euphytica 127, 209–218. doi: 10.1023/A:1020204729122
Toth, J. A., Stack, G. M., Cala, A. R., Carlson, C. H., Wilk, R. L., Crawford, J. L., et al. (2020). Development and validation of genetic markers for sex and cannabinoid chemotype in cannabis sativa l. Glob. Change Biol. Bioenergy 12, 213–222. doi: 10.1111/gcbb.12667
Trojak-Goluch, A., Skomra, U. (2013). Artificially induced polyploidization in humulus lupulus l. and its effect on morphological and chemical traits. Breed. Sci. 63, 393–399. doi: 10.1270/jsbbs.63.393
Van Bakel, H., Stout, J. M., Cote, A. G., Tallon, C. M., Sharpe, A. G., Hughes, T. R., et al. (2011). The draft genome and transcriptome of cannabis sativa. Genome Biol. 12, 1–18. doi: 10.1186/gb-2011-12-10-r102
Vassilevska-Ivanova, R. (2019). Biology and ecology of genus cannabis: genetic origin and biodiversity. In vitro production of cannabinoids. Genet. Plant Physiol. 9, 75–98.
Veloso, J., van Kan, J. A. (2018). Many shades of grey in botrytis–host plant interactions. Trends Plant Sci. 23, 613–622. doi: 10.1016/j.tplants.2018.03.016
Vergara, D., Huscher, E. L., Keepers, K. G., Givens, R. M., Cizek, C. G., Torres, A., et al. (2019). Gene copy number is associated with phytochemistry in cannabis sativa. AoB Plants 11, plz074. doi: 10.1093/aobpla/plz074
Volkow, N. D., Baler, R. D., Compton, W. M., Weiss, S. R. (2014). Adverse health effects of marijuana use. N. Engl. J. Med. 370, 2219–2227. doi: 10.1056/NEJMra1402309
Voytas, D. F. (2013). Plant genome engineering with sequence-specific nucleases. Annu. Rev. Plant Biol. 64, 327–350. doi: 10.1146/annurev-arplant-042811-105552
Vyskot, B., Hobza, R. (2004). Gender in plants: sex chromosomes are emerging from the fog. Trends Genet. 20, 432–438. doi: 10.1016/j.tig.2004.06.006
Wahby, I., Caba, J. M., Ligero, F. (2013). Agrobacterium infection of hemp (Cannabis sativa l.): establishment of hairy root cultures. J. Plant Interact. 8, 312–320. doi: 10.1080/17429145.2012.746399
Wahby, I., Caba, J. M., Ligero, F. (2017). “Hairy root culture as a biotechnological tool in c. sativa,” in Cannabis sativa l. – botany and biotechnology. Eds. Chandra, S., Lata, H., ElSohly, M. (Cham: Springer), 299–317. doi: 10.1007/978-3-319-54564-6_14
Wanas, A. S., Radwan, M. M., Mehmedic, Z., Jacob, M., Khan, I. A., Elsohly, M. A. (2016). Antifungal activity of the volatiles of high potency cannabis sativa l. against cryptococcus neoformans. Rec. Nat. Prod. 10, 214.
Wang, R., He, L.-S., Xia, B., Tong, J.-F., Li, N., Peng, F. (2009). A micropropagation system for cloning of hemp (Cannabis sativa l.) by shoot tip culture. Pak. J. Bot. 41, 603–608.
Wang, K., Shi, L., Liang, X., Zhao, P., Wang, W., Liu, J., et al. (2022). The gene TaWOX5 overcomes genotype dependency in wheat genetic transformation. Nat. Plants 8, 110–117. doi: 10.1038/s41477-021-01085-8
Wang, B., Zhu, L., Zhao, B., Zhao, Y., Xie, Y., Zheng, Z., et al. (2019). Development of a haploid-inducer mediated genome editing system for accelerating maize breeding. Mol. Plant 12, 597–602. doi: 10.1016/j.molp.2019.03.006
Weiblen, G. D., Wenger, J. P., Craft, K. J., ElSohly, M. A., Mehmedic, Z., Treiber, E. L., et al. (2015). Gene duplication and divergence affecting drug content in cannabis sativa. New Phytol. 208, 1241–1250. doi: 10.1111/nph.13562
Welling, M. T., Shapter, T., Rose, T. J., Liu, L., Stanger, R., King, G. J. (2016). A belated green revolution for cannabis: virtual genetic resources to fast-track cultivar development. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.01113
Wiedenheft, B., Sternberg, S. H., Doudna, J. A. (2012). RNA-Guided genetic silencing systems in bacteria and archaea. Nature 482, 331–338. doi: 10.1038/nature10886
Wielgus, K., Luwanska, A., Lassocinski, W., Kaczmarek, Z. (2008). Estimation of cannabis sativa l. tissue culture conditions essential for callus induction and plant regeneration. J. Nat. Fibers 5, 199–207. doi: 10.1080/15440470801976045
Wróbel, T., Dreger, M., Wielgus, K., Słomski, R. (2022). Modified nodal cuttings and shoot tips protocol for rapid regeneration of cannabis sativa l. J. Nat. Fibers 19, 536–545. doi: 10.1080/15440478.2020.1748160
Xie, K., Yang, Y. (2013). RNA-Guided genome editing in plants using a CRISPR–cas system. Mol. Plant 6, 1975–1983. doi: 10.1093/mp/sst119
Zarei, A., Behdarvandi, B., Tavakouli Dinani, E., Maccarone, J. (2021). Cannabis sativa l. photoautotrophic micropropagation: a powerful tool for industrial scale in vitro propagation. In Vitro Cell. Dev. Biol. 57, 932–941. doi: 10.1007/s11627-021-10167-3
Zarei, A., Davis, B., Feyissa, B. A., Dinani, E. T., Simons, B. (2023). Improvement of mineral nutrition and rooting efficiency of cannabis sativa l. for in vitro large-scale propagation. In Vitro Cell. Dev. Biol. 59, 95–105. doi: 10.1007/s11627-022-10320-6
Zarei, A., Feyissa, B. A., Davis, B., Tavakouli Dinani, E. (2022). Cannabis synthetic seeds: an alternative approach for commercial scale of clonal propagation and germplasm conservation. Plants 11, 3186. doi: 10.3390/plants11233186
Zhang, Q., Chen, X., Guo, H., Trindade, L. M., Salentijn, E. M., Guo, R., et al. (2018). Latitudinal adaptation and genetic insights into the origins of cannabis sativa l. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01876
Zhang, X., Xu, G., Cheng, C., Lei, L., Sun, J., Xu, Y., et al. (2021). Establishment of an agrobacterium-mediated genetic transformation and CRISPR/Cas9-mediated targeted mutagenesis in hemp (Cannabis sativa l.). Plant Biotechnol. J. 19, 1979–1987. doi: 10.1111/pbi.13611
Zhu, H., Li, C., Gao, C. (2020). Applications of CRISPR–cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 21, 661–677. doi: 10.1038/s41580-020-00288-9
Zhu, P., Zhao, Y., You, X., Zhang, Y. J., Vasseur, L., Haughn, G., et al. (2022). A versatile protoplast system and its application in Cannabis sativa l. Botany. doi: 10.1139/cjb-2021-0178
Zirpel, B., Kayser, O., Stehle, F. (2018). Elucidation of structure-function relationship of THCA and CBDA synthase from cannabis sativa l. J. Biotechnol. 284, 17–26. doi: 10.1016/j.jbiotec.2018.07.031
Zong, Y., Liu, Y., Xue, C., Li, B., Li, X., Wang, Y., et al. (2022). An engineered prime editor with enhanced editing efficiency in plants. Nat. Biotechnol. 40, 1394–1402. doi: 10.1038/s41587-022-01254-w
Keywords: Cannabis sativa, medical Cannabis, drug type Cannabis, genome editing, plant regeneration, transformation
Citation: Ingvardsen CR and Brinch-Pedersen H (2023) Challenges and potentials of new breeding techniques in Cannabis sativa. Front. Plant Sci. 14:1154332. doi: 10.3389/fpls.2023.1154332
Received: 30 January 2023; Accepted: 24 May 2023;
Published: 08 June 2023.
Edited by:
Marcello Iriti, University of Milan, ItalyReviewed by:
Mohsen Hesami, University of Guelph, CanadaPankaj Kumar Bhowmik, National Research Council Canada (NRC), Canada
Copyright © 2023 Ingvardsen and Brinch-Pedersen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christina Rønn Ingvardsen, Y2hyaXN0aW5hLmluZ3ZhcmRzZW5AYWdyby5hdS5kaw==
‡ORCID: Christina Rønn Ingvardsen, orcid.org/0000-0003-0445-5793
Henrik Brinch-Pedersen, orcid.org/0000-0002-9773-8903