 Sławomir Nowak
Sławomir Nowak Marcin Górniak2
Marcin Górniak2 Natalia Olędrzyńska
Natalia Olędrzyńska Przemysław Baranow
Przemysław Baranow- 1Department of Plant Taxonomy and Nature Conservation, Faculty of Biology, The University of Gdansk, Gdansk, Poland
- 2Department of Evolutionary Genetics and Biosystematics, Faculty of Biology, The University of Gdansk, Gdansk, Poland
Introduction: The monotypic genus Ambrella H. Perrier, endemic to Madagascar and traditionally placed within the subtribe Angraecinae, has long poses a challenge in terms of its phylogenetic classification.
Methods: To clarify the evolutionary position of Ambrella, analyzed nuclear (ITS) and plastid (matK) markers applying maximum likelihood methods for phylogenetic inference and estimating divergence times using a Bayesian relaxed molecular clock model implemented in BEAST.
Results: Our results show that Ambrella forms a strongly supported clade with the Malagasy genera Cryptopus Lindl., Oeonia Lindl., and Neobathiea Schltr. Despite the phylogenetic proximity, Ambrella remains morphologically distinct, particularly in terms of its floral architecture. According to molecular dating, the lineage leading to Ambrella diverged during the Late Miocene to Early Pliocene (ca. 8.8–3.3 Mya), a period of substantial ecological change in Madagascar.
Discussion: The results of this study provide new insight into the evolutionary history of Ambrella and clarify its phylogenetic position within Vandeae. Despite forming a well-supported clade with other Malagasy genera such as Cryptopus, Oeonia, and Neobathiea, Ambrella maintains clear morphological distinctiveness, particularly in its floral architecture. This suggests that its most striking morphological features may have evolved independently.
1 Introduction
The systematics of orchids, as one of the largest families of flowering plants, has always been challenging. However, it is only in the past two decades, with the development of phylogenetic and now phylogenomic studies, that many changes have occurred in the way orchids are classified (Górniak et al., 2010; Chase et al., 2015; Pérez‐Escobar et al., 2021; Pérez-Escobar et al., 2024). Nevertheless, it seems that the phylogeny of this group is more complex than it might originally appear, and there are still many questions to be resolved. In this group of plants, as in no other, phenomena, such as hybridization, transfer of genetic material via other organisms, or incomplete lineage sorting, play an important role and complicate the pattern of phylogenetic relationships obtained through simple analyses of genetic markers.
One of the most diverse groups in Orchidaceae are species of the tribe Vandeae comprising mainly epiphytes from tropical regions. Recently, they have been classified under Epidendroideae, the largest subfamily among orchids (Chase et al., 2015; Freudenstein and Chase, 2015; Pérez-Escobar et al., 2024). The tribe includes four subtribes, Aeridinae, Adrorhizinae, Polystachyinae, and Angraecinae, with a relatively well-studied phylogeny (Russell et al., 2010; Mytnik-Ejsmont, 2011; Mytnik-Ejsmont et al., 2015; Simo-Droissart et al., 2018; Kim et al., 2020). However, there are still many gaps to be filled, including the need to place some taxa on the phylogenetic tree. One of these is the genus Ambrella H. Perrier, which has been included in the subtribe Angraecinae since it was first described (De La Bathie, 1934; Dressler, 1993; Szlachetko, 1995; Chase et al., 2015). The main objective of this work is to verify and check the placement of this unusual and somewhat mysterious genus in the angraecoid orchids based on molecular data.
Ambrella H. Perrier was described in 1934 in the Bulletin de la Société Botanique de France as a monotypic genus with the only representative being A. longituba H. Perrier, a species unique among angraecoid orchids in terms of flower morphology. Interestingly, in the protologue, the author compared his discovery with Sobennikofia Schltr. and Oeoniella Schltr., which are genera that differ significantly from Ambrella in many respects (Figure 1). Perrier De la Bathie (1934) emphasized its distinctness in the shape of the labellum and the arrangement of the reproductive organs, its much smaller size, broad, flat leaves, and greenish flowers when compared with the two genera mentioned. In the first description, Ambrella was characterized as unique and distinctly different from Sobennikofia and Oeoniella by having a long tubular labellum, hairy inside the tube, with a base that completely hides the column, and the median lobe of the rostellum thickened and exceeding the laterals, pollinia with tegulas, and separate, free viscidia (Perrier De la Bathie 1934).

Figure 1. Plant with inflorescence (A) and frontal view of the flower (B) of Ambrella longituba H. Perrier, fragment of the inflorescence showing the flowers of Oeoniella polystachys (Thouars) Schltr. (C), close-up of the flower of Sobennikoffia humbertiana H. Perrier (D) [photographs (A, B) taken by Stefano Pagnoni; (C, D) taken by Ron Parson].
Ambrella longituba was discovered in Madagascar, in Central Highlands, on the borders of the northern sector of the Eastern Highlands. The detailed location in the protologue states the river Makys, at approximately 800 m, on the northern side of the base of Mount Ambre, as an epiphyte on the branches of Calliandra alternans Benth., hanging over waterfalls or rapids. The species is known from only a few collections and is recognized as threatened (Bachman et al., 2024). As far as we know, the species is known in herbarium resources, only by the original material and collections made by Dupuy s.n. (34) from 1992 to 1995 in the same location, the vicinity of Joffreville, Mt. Ambre. All specimens are deposited in the Muséum National d’Histoire Naturelle in Paris, France. The species was also observed later (in 2022 by J. Hermans), and the population is estimated at fewer than 50 individuals and decreasing (Rabarimanarivo and Ramandimbisoa, 2024). Although the species is reported from Montagne d’Ambre National Park, it is facing the same threats as in neighboring areas, including forest degradation caused by illegal logging, illegal agricultural practices, fires, and grazing.
Considering the small population size in a limited distribution area and the threats, it is understandable that Ambrella longituba has been assessed as critically endangered (CR) according to the IUCN (Rabarimanarivo and Ramandimbisoa, 2024). Therefore, such steps as including the species in Madagascar’s list of protected species and its transfer to Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) are fully justified. In addition, active protection through propagation and introduction into the environment, combined with education and the involvement of the local community in efforts to address the problem of the disappearance of species, especially endemic and extremely rare ones, which are of enormous value to Madagascar, is worth considering.
2 Materials and methods
For phylogenetic reconstruction, we used 183 taxa that represent the subtribe Angraecinae, with Polystachya species as an outgroup. The accession numbers of sequences downloaded from GenBank (www.ncbi.nlm.nih.gov), as well as those newly generated for Ambrella, are provided in Supplementary Table S1 (Supplementary Materials).
2.1 DNA isolation, amplification, and sequencing
Genomic DNA was extracted from approximately 20 mg of dried leaf tissue using the Genomic Mini Plant Kit (A&A Biotechnology, Poland) following the manufacturer’s instructions. Polymerase chain reaction (PCR) amplifications were performed in a total volume of 25 µl containing 1× buffer, 2 mM MgCl2, 0.2 mM dNTPs, 0.2 µM of each primer, 1.0 U of Color Perpetual Taq DNA polymerase (EURx Ltd. Poland), and DNA template (~30 ng). Amplified products were purified using the Wizard SV Gel and PCR Clean-UP System (Promega, USA).
The same primer pairs were used for both amplification and sequencing. Amplification of the ITS region (ITS1–5.8S–ITS2) was performed using primers 17SE and 26SE (Sun et al., 1994), while for the matK region, primers −19F (Wilson et al., 2000) and 1326R (Cuénoud et al., 2002) were used. PCR thermal cycling conditions began with an initial denaturation at 94°C for 4 min, followed by 30 cycles of denaturation at 94°C for 45 s, annealing at 52°C for 45 s, and an extension step at 72°C for either 1 min (ITS) or 2 min (matK) depending on the target region. Sequencing of the purified PCR products (~80 ng and 2.5 μM primer) was performed externally by Macrogen (Amsterdam, the Netherlands). Both DNA strands were sequenced to ensure accuracy in base calling. The sequences were edited using FinchTV v. 1.4.0 (Geospiza, Inc.), and the two complementary strands were assembled with AutoAssembler (ABI).
2.2 Phylogenetic analyses
DNA sequences were aligned using MAFFT v.7 (Katoh and Standley, 2013). Separate alignments were prepared for the ITS region (850 bp) and matK gene (1,361 bp). For each dataset, the best-fit evolutionary model was estimated using IQ-TREE v.2. Due to the absence of significant topological conflicts, nuclear and plastid DNA data were combined into a single concatenated matrix (2,211 bp) for further analyses. Additional details about the datasets analyzed are presented in Table 1.
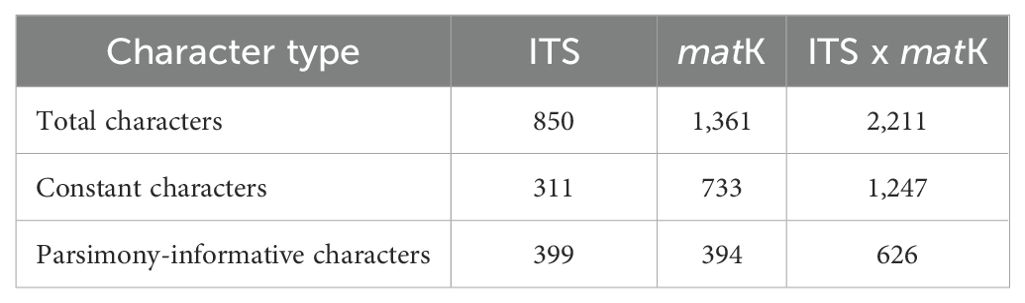
Table 1. Summary of character statistics for the analyzed datasets.
Phylogenetic reconstruction was performed using the maximum likelihood (ML) method in IQ-TREE v.2 (Minh et al., 2020). Clade support was assessed through a nonparametric bootstrap analysis with 1,000 replicates in the same software. Additionally, divergence time estimation within the studied group was conducted using BEAST v.1.8.4, executed on the CIPRES Science Gateway server (Miller et al., 2010). The analysis was run in two independent replicates, each consisting of 40 million generations. An uncorrelated relaxed molecular clock (lognormal) and the Yule model of speciation were applied. Two calibration points, following Givnish et al. (2015), were used: 27.34 million years ago (Mya) for the Polystachyinae node and 21.21 Mya for the ingroup. The results from both independent runs were assessed in Tracer v.1.6 to ensure convergence and subsequently combined in LogCombiner v.1.8.4 where 25% of the initial generations from each run were discarded as burn-in. The final maximum clade credibility (MCC) tree was generated using TreeAnnotator v.1.8.4.
3 Results
The results of our analyses are presented on a maximum clade credibility tree derived from a molecular clock analysis illustrating the divergence times of the studied lineages (Figure 2). Support values for individual clades are indicated above the branches as posterior probability (PP)/bootstrap support (BS).
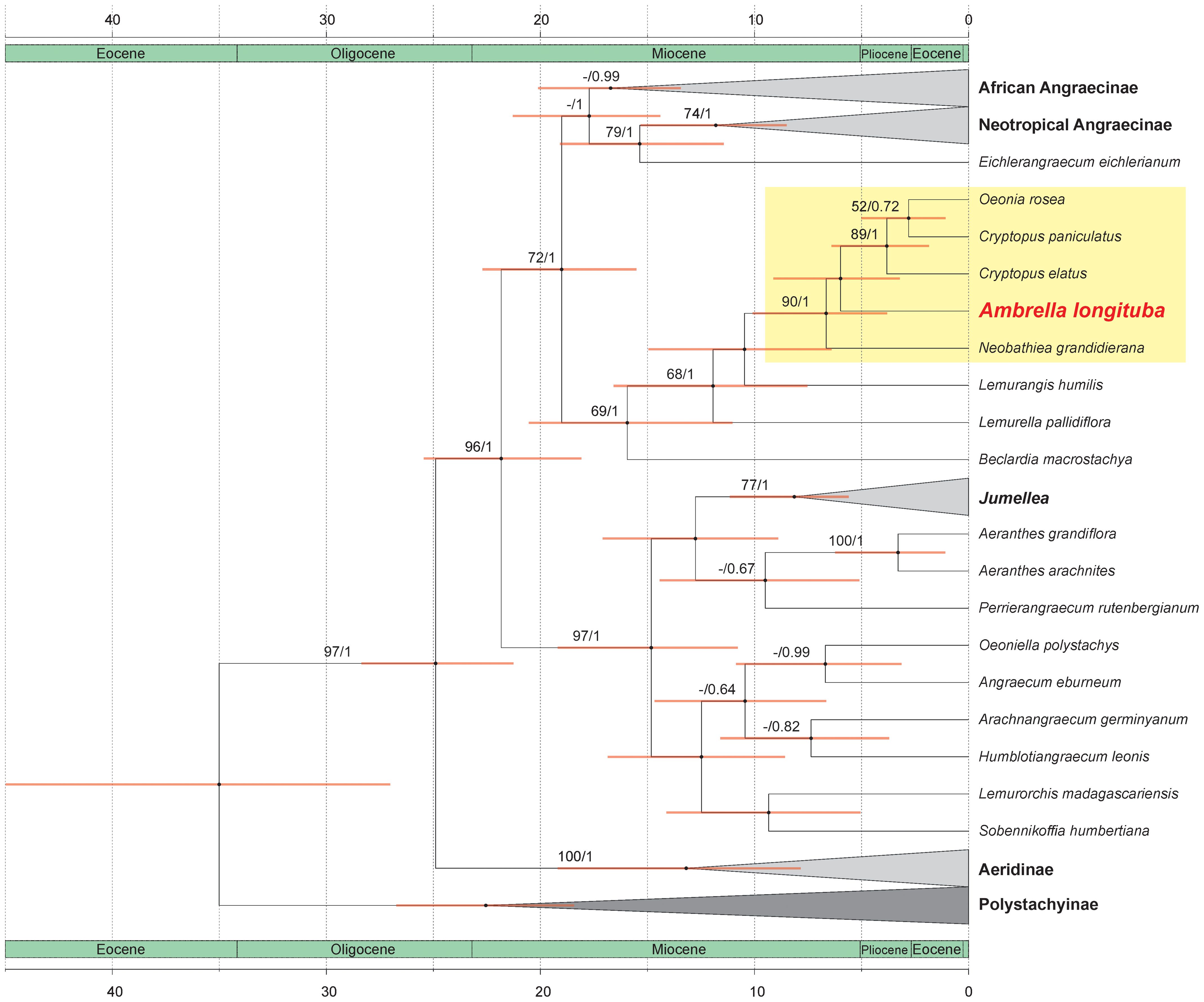
Figure 2. The maximum clade credibility tree presenting relationships within subtribe Angraecinae obtained from combined (nr ITS and plastid matK) dataset using Bayesian inference. The divergence times (in Mya) are shown on the scale bars located at the top and bottom of the figure. Node support values are shown as PP/BS.
The overall topology of the inferred phylogenetic tree is congruent with previous studies (Simo-Droissart et al., 2018; Farminhão et al., 2021) further supporting the robustness of the recovered relationships.
Our analyses strongly support the placement of the genus Ambrella, represented by A. longituba, within a well-supported clade (PP/BS = 90/1), grouping it with Oeonia rosea Ridl., Cryptopus paniculatus H. Perrier, C. elatus (Thouars) Lindl., and Neobathiea grandidieriana (Rchb.f.) Garay. This suggests a close evolutionary relationship among these taxa. The estimated divergence time for the most recent common ancestor (MRCA) of this group falls within the late Miocene to early Pliocene, approximately 8.83 to 3.31 Mya.
4 Discussion
4.1 Phylogenetic placement
Our investigations confirm the previous phylogenetic framework for Vandeae and especially for angraecoid orchids (Simo-Droissart et al., 2018). Ambrella is embedded in the clade along with the so-called vitsyangraecoides (Farminhão et al., 2021), which includes a few taxa found mainly in Madagascar and also in eastern Africa. The phylogenetic position of Ambrella, along with Cryptopus Lindl. and Oeonia Lindl., is strongly supported. Interestingly, both of these genera are very different from Ambrella (Figure 3). In both, we can observe elongated, ascending stems with loosely spaced leaves, at least several-flowered, elongated inflorescences, and wide-open flowers with a freely accessible gynostemium and spur entrance. The generative parts in representatives of both genera relate in structure to Angraecum Bory s.str., i.e., the rostellum is dome shaped, with a weakly marked middle lobe. In contrast, in Ambrella, Cryptopus, and Oeonia, the pollinia are attached to two tegulae, at the base of which separate viscidia are present. The genus Neobathiea Schltr. is then sister to them, followed by the genus Lemurella Schltr., with the East African Lemurangis humilis (Summerh.) Szlach., Mytnik & Grochocka, a representative of the Angraecum complex (=A. humile Summerh.), and finally Beclardia A. Rich., which is sister to all of them. Of the genera mentioned, elongated stems with significantly distant leaves can be observed in Neobathiea (e.g., N. spatulata H. Perrier), but already in N. perrieri (Schltr.) Schltr. or N. hirtula H. Perrier and Lemurella, the leaves have a similar form and distribution on the stem as in Ambrella. One could speculate that the primary feature in this evolutionary line, maintained in Ambrella, is the shortened stem and its relatively dense foliage. In Ambrella ancestors, there is a tendency to elongate the stem by making it ascending. This trait is undoubtedly an adaptation to habitat conditions. In this clade, a clear differentiation in the morphology of the floral parts is observed, while the structure of the generative parts is maintained. In all these genera, with the exception of Beclardia, the short, dome-shaped rostellum, with a weakly marked central patch, is present, and the pollinia are attached to two tegulae, with an independent viscidium on each. In B. macrostachya (Thouars) A. Rich., the rostellum is strongly elongated, and the equally elongated tegula is attached to a single viscidium. Both pollinia are attached to the apical, disc-shaped extended part of the tegula.

Figure 3. Single flower (A) and inflorescence (B) of Cryptopus paniculatus H. Perrier, close-up of the flower of Cryptopus elatus (Thouars) Lindl. (C), single-flowered inflorescences of Neobathiea grandidieriana (Rchb.f) Garay (D), single flower (E) and inflorescence (F) of Oeonia rosea Ridl. (photographs taken by Ron Parson).
Such a resolution of phylogeny is consistent with previous studies (Simo-Droissart et al., 2018; Farminhão et al., 2021). Interestingly, the vitsyangraecoides group is not most closely related to the other Malagasy angraecoids and is sister to the Afroneotropical clade. Thus, Ambrella is not closely related to the Oeoniella and Sobennikoffia compared in the protologue. In both genera, the stem is elongated, as are the leaves, and the inflorescences are multiflowered. The lip in Oeoniella is tubular, with a relatively narrow spur entrance and access to the gynostemium. The structure of the gynostemium in the two genera is different. In Oeoniella, the lateral lobes of the rostellum are elongate, the median lobe is strongly reduced, and the pollinia are attached to two tegulae that connect to a single viscidium. In Sobennikoffia, the lateral lobes of the rostellum are strongly reduced, with both pollinia attached to a common tegula, to which two independent viscidia are attached on the other side.
Despite being related to other taxa than assumed, Ambrella is morphologically a completely separate taxon. Vegetative traits in all Angraecinae are confusing, and taxonomic distinctiveness cannot usually be recognized, but flower architecture resolves this problem. The inflorescence of Ambrella longituba, with one to three flowers, does not resemble the multifloral inflorescence of Cryptopus and Oeonia or uniflowered inflorescence in Neobathiea. Finally, the tubular labellum, with hairs inside and very short spur observed in Ambrella, is completely different from the spurless flowers of Cryptopus and Oeonia, which have a flat labellum, at most slightly tubular at the base, through fusion with the gynostemium. Moreover, the Neobathiea has a very long, downward-hanging spur, which is typical of many angraecoid orchids.
4.2 Divergence time
Ambrella longituba falls within a clade of taxa that separated in the Early Miocene (ca. 8.8–3.3 Mya), and diversification within it continued up to the Pliocene. The Miocene period is a time of significant changes in the vegetation of Africa (Couvreur et al., 2021). Due to the cooling of the climate in the middle Miocene, many forest formations experienced decline and division especially on the continent, but it could also be the reason for the lack of diversification or extinction in Madagascar (Buerki et al., 2013). The estimated divergence time for the most recent common ancestor of Ambrella and sister genera falls within the Late Miocene to Early Pliocene, approximately 8.83 to 3.31 Mya. This is a time of ongoing changes in the vegetation of Madagascar and the development and expansion of humid forests (Buerki et al., 2013), which provide the main habitat for Ambrella and closely related taxa, promoting their cladogenesis.
4.3 Pollination possibilities
Representatives of the Angraecinae are fairly well known for their mechanisms of pollination by long-proboscis insects (Lepidoptera), which is related to the presence in many species of a long spur containing nectar (Micheneau et al., 2014; Netz and Renner, 2017). However, other groups have also been observed as effective pollinators in the subtribe such as Diptera and Hymenoptera (Singer and Cocucci, 1999) for Campylocentrum aromaticum Barb.Rodr. or even Orthoptera (Micheneau et al., 2010) for Hadrangis cadetii (Bosser) Szlach., Mytnik & Grochocka (=Angraecum cadetii Bosser).
Nothing is known, so far, about pollination in Ambrella longituba. According to the note of Perrier left under the protologue (1924), it may be associated with a dependence on a narrow group of potential pollinators. The insect that inserts its head into a small window at the base of the lip to pump nectar from the spur can only exit through a window covered by two pollen-connecting stipes. The narrowness of this window must allow pollination of this flower only by a few very exceptional insects. The role of the hairs inside the tubular labellum remains unclear. Such a feature is often an adaptation to zoogamy, and in many plants, such a closed pathway to the reproductive organs is evidence of trap flowers (Case and Bradford, 2009; Matallana-Puerto et al., 2024). The hairs may also be part of the attraction and food for pollinators (Davies et al., 2002).
Finally, self-pollination cannot be eliminated, which has been confirmed for other closely related taxa, such as Cryptopus elatus, Oeonia rosea, and Beclardia macrostachya (Thouars) A.Rich (Agnew, 1986; Jacquemyn et al., 2005). Only for Neobathiea grandidieriana was pollination by Lepidoptera observed (Nilsson et al., 1987), which is obviously related to the presence of a spur, which is also present in Ambrella, but certainly not in this form. Therefore, pollination in Ambrella remains an interesting question to investigate.
5 Conclusion
The results presented here confirm the previous position of the genus Ambrella within the subtribe Angraecinae. However, Ambrella is more closely related to genera such as Cryptopus, Oeonia, and Neobathiea than Oeoniella and Sobennikoffia, with which it has so far been compared. Therefore, the tubular lip that surrounds the gynostemium arose twice independently in Ambrella and Oeoniella. Both genera belong to two separate clades that diverged in the early Miocene. Ambrella falls within a group of several species-poor genera found mainly in Madagascar, sister to the Afro-Neotropical Angraeciinae, while Oeoniella is a part of a clade of Malagasy taxa that have reached a high diversity on the island.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
SN: Investigation, Writing – original draft, Visualization, Validation, Formal analysis, Conceptualization, Methodology, Writing – review & editing, Data curation. MG: Methodology, Writing – original draft, Formal analysis, Writing – review & editing, Investigation, Validation. NO: Writing – review & editing, Investigation, Conceptualization, Validation, Writing – original draft, Methodology, Formal analysis, Visualization, Data curation. PB: Data curation, Writing – original draft, Methodology, Investigation, Validation, Formal analysis, Writing – review & editing. DS: Investigation, Conceptualization, Writing – review & editing, Writing – original draft, Methodology, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The research was funded by the Biology Faculty of the University of Gdańsk.
Acknowledgments
We thank Mr. Marcin Kaczmarski, from whose private collection the living plants were provided. We are grateful to Ron Parson and Stefano Pagnoni for giving us permission to use their photographs in this paper. We thank the reviewers for their valuable comments, which contributed to the improvement of the article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1602122/full#supplementary-material
Supplementary Table 1 | Genbank accession numbers of the sequences used in this study. Newly generated sequences are marked with an asterisk (*).
References
Agnew, J. D. (1986). Self-compatibility/incompatibility in some orchids of the subfamily vandoideae. Plant Breed. 97, 183–186. doi: 10.1111/j.1439-0523.1986.tb01049.x
Bachman, S. P., Brown, M. J. M., Leão, T. C. C., Nic Lughadha, E., and Walker, B. E. (2024). Extinction risk predictions for the world’s flowering plants to support their conservation. New Phytol. 242, 797–808. doi: 10.1111/nph.19592
Buerki, S., Devey, D. S., Callmander, M. W., Phillipson, P. B., and Forest, F. (2013). Spatio-temporal history of the endemic genera of Madagascar. Bot. J. Linn. Soc. 171, 304–329. doi: 10.1111/boj.12008
Case, M. and Bradford, Z. (2009). Enhancing the trap of lady’s slippers: a new technique for discovering pollinators yields new data from Cypripedium parviflorum (Orchidaceae). Bot. J. Linn. Soc. 160, 1–10. doi: 10.1111/j.1095-8339.2009.00962.x
Chase, M. W., Cameron, K. M., Freudenstein, J. V., Pridgeon, A. M., Salazar, G., van den Berg, C., et al. (2015). An updated classification of Orchidaceae. Bot. J. Linn. Soc. 177, 151–174. doi: 10.1111/boj.12234
Couvreur, T. L. P., Dauby, G., Blach-Overgaard, A., Deblauwe, V., Dessein, S., Droissart, V., et al. (2021). Tectonics, climate and the diversification of the tropical African terrestrial flora and fauna. Biol. Rev. 96, 16–51. doi: 10.1111/brv.12644
Cuénoud, P., Savolainen, V., Chatrou, L. W., Powell, M., Grayer, R. J., and Chase, M. W. (2002). Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences. Am. J. Bot. 89, 132–144. doi: 10.3732/ajb.89.1.132
Davies, K. L., Roberts, D. L., and Turner, M. P. (2002). Pseudopollen and food-hair diversity in polystachya hook. (Orchidaceae). Ann. Bot. 90, 477–484. doi: 10.1093/aob/mcf220
Dressler, R. L. (1993). Phylogeny and Classification of the Orchid Family (Cambridge: Cambridge University Press).
Farminhão, J. N. M., Verlynde, S., Kaymak, E., Droissart, V., Simo-Droissart, M., Collobert, G., et al. (2021). Rapid radiation of angraecoids (Orchidaceae, Angraecinae) in tropical Africa characterised by multiple karyotypic shifts under major environmental instability. Mol. Phylogenet. Evol. 159, 107105. doi: 10.1016/j.ympev.2021.107105
Freudenstein, J. V. and Chase, M. W. (2015). Phylogenetic relationships in Epidendroideae (Orchidaceae), one of the great flowering plant radiations: progressive specialization and diversification. Ann. Bot. 115 (4), 665–681.
Givnish, T. J., Spalink, D., Ames, M., Lyon, S. P., Hunter, S. J., Zuluaga, A., et al. (2015). Orchid phylogenomics and multiple drivers of their extraordinary diversification. Proc. Biol. Sci. 282, 20151553. doi: 10.1098/rspb.2015.1553
Górniak, M., Paun, O., and Chase, M. W. (2010). Phylogenetic relationships within Orchidaceae based on a low-copy nuclear coding gene, Xdh: Congruence with organellar and nuclear ribosomal DNA results. Mol. Phylogenet. Evol. 56, 784–795. doi: 10.1016/j.ympev.2010.03.003
Jacquemyn, H., Micheneau, C., Roberts, D. L., and Pailler, T. (2005). Elevational gradients of species diversity, breeding system and floral traits of orchid species on Réunion Island. J. Biogeogr. 32, 1751–1761. doi: 10.1111/j.1365-2699.2005.01307.x
Katoh, K. and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kim, Y.-K., Jo, S., Cheon, S.-H., Kwak, M., Kim, Y.-D., and Kim, K.-J. (2020). Plastome evolution and phylogeny of subtribe Aeridinae (Vandeae, Orchidaceae). Mol. Phylogenet. Evol. 144, 106721. doi: 10.1016/j.ympev.2019.106721
Matallana-Puerto, C. A., Brito, V. L. G., Kuster, V. C., Oliveira, P. E., and Cardoso, J. C. F. (2024). Sex, flies and flower trap: Trapping trichomes and their function in pollination. Funct. Ecol. 38, 2261–2270. doi: 10.1111/1365-2435.14633
Micheneau, C., Fournel, J., and Pailler, T. (2014). “Pollination biology and evolutionary history of angraecoid orchids: from Darwin to the present day,”. Eds. Edens-Meier, R. and Bernhardt, P. (University of Chicago Press, Chicago, IL, USA: Darwin’s Orchids Then and Now), 201–227. Available at: http://www.press.uchicago.edu/ucp/books/book/chicago/D/bo18659332.html.
Micheneau, C., Fournel, J., Warren, B. H., Hugel, S., Gauvin-Bialecki, A., Pailler, T., et al. (2010). Orthoptera, a new order of pollinator. Ann. Bot. 105, 355–364. doi: 10.1093/aob/mcp299
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). Creating the CIPRES Science Gateway for inference of large phylogenetic trees. 2010 Gateway Comput. Environ. Works. (GCE) 1–8. doi: 10.1109/GCE.2010.5676129
Minh, B. Q., Schmidt, H. A., Chernomor, O., Schrempf, D., Woodhams, M. D., Von Haeseler, A., et al. (2020). IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 37, 1530–1534. doi: 10.1093/molbev/msaa015
Mytnik-Ejsmont, J. (2011). A monograph of the subtribe Polystachyinae schltr. (Orchidaceae) (Mytnik-Ejsmont - Gdańsk: Fundacja Rozwoju Uniwersytetu Gdańskiego).
Mytnik-Ejsmont, J., Szlachetko, D. L., Baranow, P., Jolliffe, K., and Górniak, M. (2015). Phylogenetic placement and taxonomy of the genus hederorkis (Orchidaceae). PloS One 10, e0122306. doi: 10.1371/journal.pone.0122306
Netz, C. and Renner, S. S. (2017). Long-spurred Angraecum orchids and long-tongued sphingid moths on Madagascar: A time frame for Darwin’s predicted Xanthopan/Angraecum coevolution. Biol. J. Linn. Soc. 122, 469–478. doi: 10.1093/biolinnean/blx086
Nilsson, L. A., Johnsson, L., Ralison, L., and Randrianjohany, E. (1987). Angraecoid orchids and hawkmoths in central Madagascar: specialized pollination systems and generalist foragers. Biotropica 19, 310–318. doi: 10.2307/2388628
Perrier De la Bathie, H. (1934). Ambrella, genre nouveau d’Orchidées (Angraecoïdées) de Madagascar. Bull. la Société Bot. France 81, 655–657. doi: 10.1080/00378941.1934.10834012
Pérez-Escobar, O. A., Bogarín, D., Przelomska, N. A. S., Ackerman, J. D., Balbuena, J. A., Bellot, S., et al. (2024). The origin and speciation of orchids. New Phytol. 242, 700–716. doi: 10.1111/nph.19580
Pérez-Escobar, O. A., Dodsworth, S., Bogarín, D., Bellot, S., Balbuena, J. A., Schley, R. J., et al. (2021). Hundreds of nuclear and plastid loci yield novel insights into orchid relationships. Am. J. Bot. 108, 1166–1180. doi: 10.1002/ajb2.1702
Rabarimanarivo, M. and Ramandimbisoa, B. (2024). Ambrella longituba. The IUCN Red List of Threatened Species 2024.
Russell, A., Samuel, R., Rupp, B., Barfuss, M. H. J., Šafran, M., Besendorfer, V., et al. (2010). Phylogenetics and cytology of a pantropical orchid genus Polystachya (Polystachyinae, Vandeae, Orchidaceae): Evidence from plastid DNA sequence data. TAXON 59, 389–404. doi: 10.1002/tax.592005
Simo-Droissart, M., Plunkett, G. M., Droissart, V., Edwards, M. B., Farminhão, J. N. M., Ječmenica, V., et al. (2018). New phylogenetic insights toward developing a natural generic classification of African angraecoid orchids (Vandeae, Orchidaceae). Mol. Phylog. Evol. 126, 241–249. doi: 10.1016/j.ympev.2018.04.021
Singer, R. B. and Cocucci, A. A. (1999). Pollination mechanism in southern Brazilian orchids which are exclusively or mainly pollinated by halictid bees. Pl Syst. Evol. 217, 101–117. doi: 10.1007/BF00984924
Szlachetko, D. L. (1995). Systema Orchidalium. Fragmenta Floristica et Geobotanica Polonica. Supplementum 3, 1–152.
Sun, Y., Skinner, D. Z., Liang, G. H., and Hulbert, S. H. (1994). Phylogenetic analysis of Sorghum and related taxa using internal transcribed spacers of nuclear ribosomal DNA. Theor. Appl. Genet. 89, 26–32. doi: 10.1007/BF00226978
Wilson, K., Morrison, D., Molvray, M., Kores, P., and Chase, M. (2000). Polyphyly of mycoheterotrophic orchids and functional influences on floral and molecular characters. Available online at: https://www.semanticscholar.org/paper/Polyphyly-of-mycoheterotrophic-orchids-and-on-and-Wilson-Morrison/563adef9cb4b5732ecc46d6116a9f10887d26ccf (Accessed March 27, 2025).
Keywords: Angraecinae, divergence time estimation, molecular taxonomy, Orchidaceae, phylogenetics
Citation: Nowak S, Górniak M, Olędrzyńska N, Baranow P and Szlachetko DL (2025) Phylogenetic placement of the mysterious Malagasy genus Ambrella H. Perrier (Vandeae, Orchidaceae). Front. Plant Sci. 16:1602122. doi: 10.3389/fpls.2025.1602122
Received: 28 March 2025; Accepted: 29 May 2025;
Published: 19 June 2025.
Edited by:
Jeremie Benjamin Fant, Chicago Botanic Garden, United StatesReviewed by:
Fernando Rocha Vento, Northwestern University, United StatesEric Smidt, Federal University of Paraná, Brazil
Copyright © 2025 Nowak, Górniak, Olędrzyńska, Baranow and Szlachetko. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Natalia Olędrzyńska, bmF0YWxpYS5vbGVkcnp5bnNrYUB1Zy5lZHUucGw=