 Julio Cezar Souza Vasconcelos1
Julio Cezar Souza Vasconcelos1 Caio Simplicio Arantes1
Caio Simplicio Arantes1 Eliane Aparecida Gomes2*
Eliane Aparecida Gomes2* Christiane Abreu de Oliveira-Paiva2
Christiane Abreu de Oliveira-Paiva2 Sylvia Morais de Sousa2
Sylvia Morais de Sousa2 Eduardo Antonio Speranza3
Eduardo Antonio Speranza3 João Francisco Gonçalves Antunes3
João Francisco Gonçalves Antunes3 Ubiraci Gomes de Paula Lana2
Ubiraci Gomes de Paula Lana2 Geraldo Magela de Almeida Cançado3*
Geraldo Magela de Almeida Cançado3*- 1Fundação de Apoio à Pesquisa e ao Desenvolvimento (FAPED), Campinas, São Paulo, Brazil
- 2Embrapa Maize and Sorghum, Sete Lagoas, Minas Gerais, Brazil
- 3Embrapa Digital Agriculture, Campinas, São Paulo, Brazil
Climate change exacerbates drought stress, posing challenges to global soybean grain yield. This study assesses the effectiveness of microbial inoculants derived from two Bacillus velezensis (strains 5D5, 6E9) and one Bacillus subtilis (strain 1A11), which were previously selected in vitro to promote growth and enhance drought resilience in soybeans (Glycine max [L.] Merr.), and evaluated through agronomic metrics and remote sensing. A greenhouse experiment was conducted to evaluate the performance of these inoculants under both irrigated and drought conditions. The inoculants were applied at the dose-range of 1, 2, 3 and 4 mL Kg-1 of seed to identify the optimal dose. The greenhouse results indicated that for many tested doses, the three bacterial strains significantly increased shoot fresh weight, shoot dry weight, and root dry weight compared to control treatments. Multi-location field-trials in Brazil (Birigui, Itapira and Piracicaba) were conducted during the growing seasons of 2022–2023 and 2023–2024, using 3 mL Kg-1 of seed as a reference dose. These field-trials revealed yield improvements of 11.3 to 18\% for inoculated treatments, with B. subtilis 1A11 achieving the highest grain yield of 620 Kg ha-1 over the control. However, all three microbial inoculants significantly enhanced soybean development and grain yield relative to non-inoculated controls. Vegetation indices, particularly the Enhanced Vegetation Index 2 (EVI2), derived from PlanetScope satellite and Unmanned Aerial Vehicle (UAV) imagery, demonstrated a high overlap between field data and model predictions, confirming the value of remote sensing as a predictive tool. Climatic variability significantly impacted the yield in field-trials, with 2022–2023 (4.28 t ha-1 outperforming 2023–2024 (3.34 t ha-1) due to higher temperatures (>40 °C) and lower rainfall in the last season. Meanwhile, locations with balanced precipitation, like Itapira, showed superior grain yield. Statistical modeling confirmed inoculant efficacy and EVI2's utility in production measurement. This study emphasizes that microbial inoculants can serve as sustainable strategies to mitigate the impacts of drought. By integrating Bacillus-based bioinoculants into soybean cultivation and utilizing both agronomic and remote sensing metrics for validation, we can enhance resilience and ultimately support food security amid climate variability.
1 Introduction
Many of the world’s agricultural regions are experiencing increasingly intense changes in climate patterns. These changes have affected the intensity and regularity of rainfall, caused significant fluctuations in historical temperature averages, and intensified the occurrence of pests, diseases, and various harmful abiotic effects during the crop cycle (Tchonkouang et al., 2024).
To mitigate the negative impacts of these environmental challenges, various strategies have been implemented to enhance the adaptive capacity of agricultural species. One approach involves incorporating genetic traits linked to stress tolerance into breeding programs, enabling plants to better withstand environmental limitations. Another strategy focuses on modifying the production environment to optimize conditions for crop growth. However, these interventions can sometimes compromise ecosystem sustainability. Fortunately, sustainable and economically viable alternatives are emerging that can be integrated into existing production systems without causing environmental harm. These alternatives not only support crop development but also maintain or even enhance productivity, contributing to global food security. The use of microorganisms in association with cultivated plants has become a promising alternative to conventional agricultural inputs (Singh et al., 2011). Commercially available products include biofertilizers as substitutes for inorganic fertilizers, biopesticides as alternatives to synthetic pesticides, and various biostimulants that promote plant growth (O’Callaghan et al., 2022). These microbial-based solutions offer a sustainable approach to improving crop performance while reducing reliance on chemical inputs.
Soybean (G. max) is a crop that particularly benefits from microbiological inoculants, which enhance field performance and grain yield (Schmidt et al., 2015). A key example is biological nitrogen fixation (BNF), a process mediated by diazotrophic bacteria of the genus Bradyrhizobium spp. These bacteria form symbiotic relationships with leguminous plants, including soybeans, enabling the conversion of atmospheric nitrogen into a biologically usable form. This process reduces the need for synthetic nitrogen fertilizers, offering both economic and environmental benefits. Brazil has been a pioneer in adopting this technology, with estimates suggesting annual savings of approximately US$15 billion due to the replacement of nitrogen fertilizers with biologically fixed nitrogen (Döbereiner, 1997; Hungria and Mendes, 2015; Hungria and Nogueira, 2019).
Several studies have demonstrated that plant growth-promoting bacteria (PGPB) provides numerous direct and indirect benefits to crop plants (Sarma and Saikia, 2014; Rho et al., 2018). These microorganisms are known to establish close associations with host plants, contributing significantly to enhanced stress tolerance, including drought (Pandey et al., 2016; Zhang et al., 2019; Dubey et al., 2021; Liu et al., 2025). Among the various PGPB, species belonging to the genus Bacillus are of particular interest due to their ability to form endospores, which enables them to survive under extreme abiotic stresses such as temperature and pH, radiation, desiccation, ultraviolet exposure, and pesticide contamination (Bahadir et al., 2018). For instance, B. velezensis has been shown to facilitate the establishment of a specific symbiosis between soybean and Bradyrhizobium strains, as well as to promote root development under water-limited conditions (Kondo et al., 2025). In rice, B. velezensis contributes to improved drought tolerance by reducing the accumulation of reactive oxygen species (Park et al., 2024). Recent studies also have demonstrated that B. subtilis plays a significant role in enhancing plant drought tolerance. It primarily alleviates drought stress by secreting various metabolites and modulating plant hormone levels (Azeem et al., 2023; Kuramshina and Khairullin, 2023; Ren et al., 2025). Furthermore, B. subtilis promotes root development and improves the soil microbial environment, thereby increasing water uptake efficiency (Khan et al., 2024) and crop productivity (Ren et al., 2025).
According to Bettiol (2022), bioproducts used in agriculture are classified into three main categories: biofertilizers, biostimulants, and biopesticides. In this study, we evaluated the efficacy of three Bacillus-based microbiological inoculants applied as biostimulants to soybean seeds at the time of sowing. The aim was to assess their potential to mitigate developmental and yield constraints under real field conditions. In addition to traditional agronomic evaluation methods, we employed remote sensing technologies, including drone and satellite imagery, to calculate vegetation indices (VIs). These indices were used to monitor crop development and correlate with agronomic performance metrics.
The integration of remote sensing with agricultural practices has become a widely used tool in the field, including the evaluation of irrigation practices on the field (Ozdogan et al., 2010; Bégué et al., 2018). The use of images obtained from UAVs and satellites has enabled the development of increasingly accurate predictive production models. Vasconcelos et al. (2025) and Vasconcelos et al. (2023), while working with sugarcane crops and temporal images obtained via satellites and drones, developed models based on VIs capable of predict sugarcane production months ahead of the harvest date. This capability offers numerous advantages for sugarcane growers, mills, and the overall production chain associated with this crop. Similarly, Santos et al. (2024), utilizing aerial images captured by drones in sugarcane experimental fields, demonstrated the effectiveness of a phosphate-solubilizing microbiological inoculant compared to traditional cultivation methods that do not use the inoculant. Particularly for soybean crops, recent studies have shown the advantages of using remote sensing to monitor the production cycle and estimate grain yield (Prince Czarnecki et al., 2025). Hu et al. (2023) used images in the visible spectrum collected by UAV and field data to develop a high-accuracy model based on machine learning to estimate vegetation cover fraction, chlorophyll content and breeding maturity. Joshi et al. (2024) developed a comparative study of several artificial intelligence (AI) models for predicting soybean grain yield, based on high spatial resolution multispectral images obtained from PlanetScope nanosatellites. The results showed that combinations of images obtained at different growth stages help to increase the accuracy of models evaluated. Amaral et al. (2024) used a machine learning model based on random forests and multispectral images from the Sentinel-2 satellite to define more appropriate vegetation indices to identify the spatiotemporal variability of vegetation vigor and canopy structure of soybean plants.
This study investigated the effectiveness of three bacterial inoculantss — B. velezensis (strains 5D5 and 6E9) and B. subtilis (strain 1A11) — in increasing the grain yield of soybean (G. max) when applied to seeds before sowing. The agronomic performance was assessed using grain yield metrics and VIs, with the results compared to non-inoculated controls to quantify the effects of the treatments.
2 Materials and methods
2.1 Isolation, selection, characterization and identification of bacterial strains
To carry out this study, strains of the bacteria B. velezensis (5D5 and 6E9) and B. subtilis (1A11) were isolated and selected in vitro based on their potential to promote growth in plants, as well as their ability to survive under osmotic stress as described by de Oliveira-Paiva et al. (2024). The microorganisms were isolated from soil samples, starting with the screening of a total of 414 strains samples. In the subsequent stages of evaluation and screening, which included the capacity to grow in medium with a high concentration of sorbitol, assessing exopolysaccharide (EPS) production capacity, biofilm production, and a qualitative evaluation of siderophore and growth phytoregulators, the three strains identified as 5D5 (germplasm code BRM051757), 6E9 (germplasm code BRM051761), and 1A11 (germplasm code BRM051734) were selected for further analysis. The methods and protocols used during this stage of the study are outlined below.
2.1.1 Identification of Bacillus strains
Bacterial genomic DNA was extracted with the Wizard Genomic DNA Purification Kit (Promega, USA) and 16S rRNA gene amplified with the primers 8F and 1492R (Turner et al., 1999). PCR reactions were performed with 30 ng of bacterial genomic DNA, 2.5 µL 10X PCR buffer (20 mM Tris–HCl pH 8.4, 50 mM KCl), 0.4 µM of each primer, 100 µM dNTP, 2.5 mM MgCl2, and 1 U Taq DNA Polymerase (Invitrogen, USA) in a total volume of 25 µL. PCR was performed with the following conditions: 95°C for 2 min, 30 cycles of 30 s at 94°C, 30 s at 55°C and 2 min at 72°C. Finally, reactions were incubated for 10 min at 72°C. The amplification products were purified with the ExoSAP-IT Kit (USB, USA), and sequenced with the primers 8F, 1492R, 515F (Turner et al., 1999), and 902R (Hodkinson and Lutzoni, 2009) using Big Dye Terminator v3.1 kit (Applied Biosystems, USA). The samples were analyzed in the automated sequencer ABI PRISM 3500 XL Genetic Analyzer (Applied Biosystems, USA), and DNA sequences were compared with the GenBank DNA sequence database using the BlastN program (Altschul et al., 1997).
2.1.2 Bacterial strains selection
To conduct this study, the bacterial strains 5D5 (BRM051757) and 6E9 (BRM051761) of B. velezensis, as well as strain 1A11 (BRM051734) of B. subtilis, were isolated and characterized by Embrapa Maize and Sorghum´s research group. The strains were obtained from soil samples collected in the Caatinga biome, a semi-arid region located in Ceará, Brazil, and were deposited in the Collection of Multifunctional and Phytopathogenic Microorganisms at Embrapa Maize and Sorghum. Experimental procedures to assess in vitro plant growth-promoting properties and drought stress tolerance were conducted at the Embrapa Maize and Sorghum research facilities.
2.1.3 Bacterial growth in medium with high concentration of sorbitol
The strains were inoculated onto tryptone soy agar (TSA; g L−1: tryptone 15, papain-digested soybean 5, NaCl 5, agar 15) enriched with 10% (w/v) sorbitol at concentrations of 520 and 780 g L−1, corresponding to water activities (aw) of 0.897 and 0.807, respectively. Plates were incubated at 40°C for 72 hours.
2.1.4 Exopolysaccharide, biofilm and siderophore production
EPS production was evaluated following Paulo et al. (2012). Sterile filter paper discs (5 mm Ø) were placed in Petri dishes containing the culture medium described by Guimarães et al. (1999) and inoculated with 5 µL of each bacterial isolate grown in TSB. Plates were incubated at 30°C for 24 h, and EPS production was indicated by a mucoid colony surrounding the discs. Production was confirmed by mixing the mucoid substance with 2 mL of absolute ethanol: a precipitate indicated positive EPS production, while turbidity indicated a negative result. Biofilm formation was assessed using the spectrophotometric method described by Stepanović et al. (2007). Qualitative siderophore production analysis was performed as previously described by Ribeiro et al. (2018) and Arora and Verma (2017). Absorbance was measured at 630 nm using a Perkin Elmer spectrophotometer (USA), and siderophore production was estimated following the method described by Ribeiro et al. (2018).
2.1.5 IAA-like molecules production
Each strain was grown in a TSB medium supplemented with 1.0 mg mL−1 tryptophan as an IAA precursor and incubated at 30°C for five days at 100 rpm in the dark (Ribeiro et al., 2018). After centrifugation at 4250 × g for 10 min, 0.1 mL of the supernatant was mixed with 0.1 mL of the Salkowski reagent (Loper and Schroth, 1986) and incubated for 20 min in the dark. The concentration of IAA-like molecules in the supernatant was determined in triplicate by the colorimetric measurement at 540 nm and compared to a standard curve prepared from commercial IAA at concentrations ranging from 0 to 100 µg mL−1 (Patten and Glick, 1996).
2.2 Greenhouse experiments and plant material
The greenhouse experiments were carried out at Embrapa Maize and Sorghum, located in Sete Lagoas, MG, Brazil, in 2023. The study evaluated the dose-range effect of inoculants individually formulated with the strains 1A11, 5D5, and 6E9 on the growth of soybean seedlings. This assessment was carried out under conditions of adequate moisture availability, as well as during episodes of drought stress.
The inoculants were prepared with bacterial colony-forming unit concentration for each strain as follows: 1A11 = 8.67x109 bacterials mL−1; 5D5 = 1.62x1010 bacterial mL-1 to CFU; and 6E9 = 9.41x109 bacterial mL-1 to CFU. Soybean seeds were treated with doses of 1, 2, 3, and 4 mL kg−1 for all tested strains, with application occurring just before sowing. The soybean seeds were homogenized with the liquid inoculant by mixing them in a container until all seeds were thoroughly moistened and covered by the inoculant. As a standard practice for soybean cultivation to enhance symbiotic nitrogen fixation, it was applied to all treatments the liquid inoculant SimbioseNod™, formulated from strains SEMIA 5079 and SEMIA 5080 of Bradyrhizobium japonicum, at a concentration of 7.2 x 109 bacterial mL-1 to CFU of inoculant, at a dose of 3 mL kg−1 of seed. The B. japonicum inoculant was applied separately, immediately after the tested Bacillus inoculants were administered. The control group received only the B. japonicum and this inoculant was mixed with the seed mass in a manner similar to the process used for the Bacillus inoculants.
For each experimental plot, 20 L pots filled with clayey red latosol, typical of the Brazilian Cerrado biome, were utilized. Soil fertilization and liming were consistent across all treatments, adhering to recommendations based on soil physicochemical analysis. Two soybean plants of the cultivar ‘BRS Valiosa RR’, were maintained in each pot. The experimental design was completely randomized, comprising two sets of 26 treatments with three replicates per treatment. The data were analyzed by the R statistical package.
Of the 78 experimental units, 39 were sustained under adequate irrigation, with soil field capacity continuously maintained at -18 kPa. The other 39 experimental units underwent drought stress conditions starting 27 days after germination, simulated by keeping the soil at a water tension of -138 kPa for 14 days. Daily monitoring of soil water availability for both irrigated and non-irrigated treatments was conducted using a tensiometer installed at a depth of 20 cm in each pot. At the conclusion of the drought stress period, the irrigated and non-irrigated plants were collected at the early flowering stage (R1), and the weights of fresh and dry matter for both the aerial parts and roots were evaluated.
2.3 Field-trials and plant material
To assess the impact of microbiological inoculants on soybean crops, field-trials were established in three different locations in Brazil: Birigui (geographical coordinate -21.318220, -50.355165), Itapira (-22.401927, -46.778187) and Piracicaba (-22.769027, -47.582643) and evaluated over two consecutive growing seasons (2022–2023 and 2023–2024). The soybean varieties was chosen based in its agronomic recommendation for each region and in its precocity of the cycle. In Birigui was cultivated the variety ‘BMX Nexus i2X’ of the maturity group 6.4, in Itapira was cultivated the variety ‘Monsoy 6601 i2X’ of the maturity group 6.6, and in Piracicaba was cultivated the variety ‘NS 5933 iPro’ of the group of maturity 6.1. To establish a stand of approximately 260,000 plants per hectare, about 13 seeds were sown per linear meter of furrow, maintaining a spacing of 50 cm between rows and the planting depth was set at 5 cm. During the planting of seeds was applied granulated fertilizer with a composition of 3-21-21 (NPK, YaraBasa™) at rates based on the soil’s chemical analysis for each location: 150 kg ha−1 in Birigui; 130 kg ha−1 in Itapira; and 180 kg ha−1 in Piracicaba and the fertilizer was positioned next to the seed, 5 cm below it. During the phenological phases between V3 and V6, a foliar micronutrient fertilizer (Micromax Foliar Concentrate™, Aqua do Brasil) was applied at a rate of 500 g ha−1.
The field-trials of Birigui and Itapira consisted of 16 experimental plots for each location and twenty rows of soybean measuring 0.5 m between furrows and 100 m in length, while the field-trial in Piracicaba also consisted of 16 experimental plots but with 10 rows of soybean measuring 0.5 m between furrows and 25 m in length. For each soybean variety, there were 4 blocks per field-trial, totaling 48 experimental plots per season and 96 experimental plots considering two harvest seasons. The random block design with four blocks per field-trial was adopted in this study to enable the measurement of random environmental variation. All experimental fields followed management practices aligned with standard commercial soybean cultivation in Brazil, ensuring consistent agronomic standards across all varieties. Soybean plots were harvested and measured in tons of soy per hectare (t ha−1) at the end of their growth cycle, once the seeds reached peak of maturation (13% of humidity based on the dry weight). Yield was assessed based on the average grain weight obtained from experimental replicates for each treatment across all locations (Birigui, Itapira and Piracicaba) over two consecutive growth cycles: 2022–2023 and 2023–2024.
2.4 Field inoculation
To assess the impact of three microbiological inoculants on soybean crops, each bacterial strain—5D5, 6E9, and 1A11—formulated as liquid inoculant was applied individually at a rate of 3 mL of liquid inoculant per kilogram of soybean seeds at the same concentration of the greenhouse study. A control treatment without inoculant application was included for comparison.
The inoculants containing strains 5D5, 6E9, and 1A11 were applied to the seeds in a similar manner to the procedure described in the greenhouse experiment. After application, the seeds were allowed to dry for five minutes before sowing, making sure to avoid direct sunlight exposure. Additionally, after the Bacillus inoculants application, B. japonicum inoculant was applied to the seeds of all treatments, including the control, using the same commercial product at the same concentration and dosage as in the greenhouse experiment.
2.5 Weather data
The climate data for the field-trials in Birigui, Itapira and Piracicaba were compiled from the NASA Power (NASA POWER Project, 2024) database as indicated by Monteiro et al. (2018). The dataset, which spans the entire soybean growing season for each location and harvest season, was sourced directly from the NASA POWER Project (2024).
2.6 Vegetation indices
The EVI2 VI (Jiang et al., 2008) was calculated for all field-trials during the phenological phase of vegetative growing. This VI can be used as an alternative to the traditional NDVI (Normalized Difference Vegetation Index) without changing the spectral bands used in its calculation (red and near infrared), which often reach saturation points in the most advanced stages of crop development, making it difficult to observe differences between treatments (Santos et al., 2024). For the fields of Birigui and Itapira were used satellite images from the PlanetScope PSB.SD instrument (Frazier and Hemingway, 2021), provided by RedeMAIS/MJSP source, with spatial resolution of 3 m/pixel and daily temporal resolution, regarding the following bands: blue (B): 465–515 nm; green (G): 547–583 nm; red (R): 650–680 nm; red edge (RE): 697–713 nm; and near-infrared (NIR): 845–885 nm. For the field of Piracicaba, due to the smaller size of the experimental area, were used images obtained from missions performed by a drone, model DJI Phantom 4 Pro equipped with RTK and two cameras: one RGB, for images in the visible spectrum; and another for multispectral array, covering the following bands: blue (B): 450 ± 16 nm; green (G): 560 ± 16 nm; red (R): 650 ± 16 nm; red edge (RE): 730 ± 16 nm; and near-infrared (NIR): 840 ± 26 nm. In this case, for each mission, orthomosaics were generated by the Agisoft Metashape software, considering the aforementioned bands with a ground sample distance (GSD) of approximately 10 cm. The satellite images or the orthomosaic assembled from drone images were then used to extract and calculate the VIs of the field-trials. For both cases, only the images or othomosaic that best represented the phenological stadium of vegetative growth (approximately 70 to 90 days) were effectively chosen to extract the EVI2 values. The average EVI2 value of the internal pixels of each experimental plot, excluding the pixels considered as borders, was then calculated and considered in this study.
2.7 Evaluation of variance inflation factor, data normality and homoscedasticity
In order to enhance the prediction model’s stability and address potential multicollinearity concerns, all predictor variables were incorporated into the initial analysis. The assessment of the Variance Inflation Factor (VIF) (Belsley et al., 1980) was performed to detect and eliminate variables with high collinearity, leading to a more streamlined and reliable selection of variables. The Shapiro-Wilk test was used for normality (Shapiro and Wilk, 1965) and the Breusch-Pagan test for homoscedasticity (Breusch and Pagan, 1979).
2.8 Variables definition
To investigate the determinants of soybean productivity, we define a set of explanatory variables that comprise both categorical and continuous covariates. The response variable is the grain yield, denoted by yi, measured in tons per hectare (t ha−1). Each observation i = 1,…,96 corresponds to a unique combination of experimental conditions. The variables are structured as follows:
● yi: Grain yield (t ha−1);
● xi1: Block factor (Block 1 to Block 4). Since it contains four levels, three binary (dummy) variables are created: bi1,bi2,bi3;
● xi2: Treatment factor (Control, 1A11, 5D5, and 6E9), represented using three dummy variables: ti1,ti2,ti3;
● xi3: Location factor (Birigui, Itapira, and Piracicaba), modeled with two dummy variables: li1,li2;
● xi4: Year of cultivation (2022–2023 and 2023–2024), encoded by a single dummy variable: wi1;
● xi5: Total accumulated precipitation during the crop cycle (in millimeters), a continuous variable;
● xi6: Mean value of the Enhanced Vegetation Index 2 (EVI2) over the crop cycle, a continuous variable.
Categorical variables (xi1 to xi4) were transformed into binary indicators through dummy encoding to ensure compatibility with linear modeling frameworks. Continuous variables (xi5 and xi6) were retained in their original form to preserve their numeric information. This structure enables the application of linear regression models and facilitates the interpretation of individual effects on soybean yield.
2.9 Statistical model
A linear regression model was used to evaluate the effect of explanatory variables on soybean grain-yield, measured in tons per hectare (t ha−1). The model includes both categorical and continuous covariates previously described and is given by:
where yi represents the response variable in observation i. The term β0 corresponds to the model intercept, which indicates the value of the response variable when all the explanatory variables are zero. The coefficients βj are the regression parameters associated with the explanatory variables xij, for j = 1,2,…,p, which quantify the influence of each of these variables on the response variable. The variables xij are the explanatory variables that can directly influence the response variable. Finally, the term ϵi represents the random error associated with observation i, which captures the variations unexplained by the variables included in the model. It is assumed that the error ϵi is a random variable with a normal distribution, mean zero, and constant variance, i.e., ϵi ∼ N(0,σ2). Thus, the equation expresses how the response variable is affected by the explanatory variables, incorporating an error component that reflects the unexplained variability.
The model was fitted using the lm() function from the R software environment (R Core Team, 2024), which estimates the regression coefficients using the Ordinary Least Squares (OLS) method. This method aims to minimize the sum of squared residuals:
where is the fitted value for observation i. In matrix form, the estimator is given by:
where X is the design matrix, y is the response vector, and is the vector of estimated coefficients. The formulation and estimation are based on standard theory for linear models as described in Chambers (1992).
3 Results and discussion
3.1 Strain in vitro characterization
The selection of Bacillus strains from the Active Germplasm Bank of Multifunctional and Phytopathogenic Microorganisms at Embrapa Maize and Sorghum focused on laboratory tests to assess their ability to grow in vitro under high osmotic potential conditions (Table 1). This included evaluating colony growth in a medium with high sorbitol concentration and low water activity. Additionally, the selected strains were examined for mechanisms that may help mitigate plant drought stress, such as the ability to exude biofilm and exopolysaccharides. Some characteristics associated with growth promotion of roots and canopy, including the synthesis of indole-3-acetic acid (IAA) and siderophores, were also evaluated. Based on the results obtained from strains 5D5 (B. velezensis), 6E9 (B. velezensis), and 1A11 (B. subtilis), the next step was to evaluate the behavior of these three strains in a biological system involving both plant and soil in a controlled growth environment. For this purpose, soybean was selected due to its agronomic significance and because it is a crop with sensitivity to moderate and extreme variations in temperature and soil humidity—conditions that are commonly observed in many agricultural systems in Brazil.
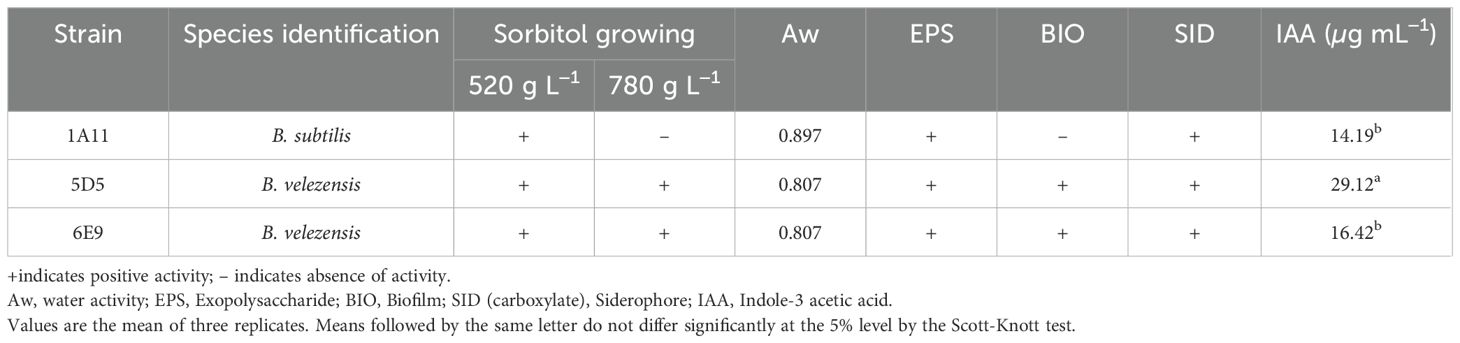
Table 1. Plant-growing promotion attributes of three selected Bacillus strains.
The identification of microorganisms that can thrive in low humidity and simultaneously produce biologically active substances holds significant promise for various applications, including enhancing plant tolerance in marginal environments. The bacterial strains were initially isolated from arid soils and the strains capable of growing in culture medium containing sorbitol at concentrations of 520 g L−1 and/or 780 g L−1 proceeded to the next stages of selection and evaluation. Further, a series of screenings was carried out to identify multifunctional mechanisms for plant growth-promoting (PGP) related to drought stress tolerance, particularly those related to exopolysaccharide (EPS) and biolfim production and capacity of growing in a low water activity. Water activity (aw) refers to the amount of water available for biological processes, and a value of 0.807, where 5D5 and 6E9 strains could grow, indicates a reduced moisture availability in the soil. For example, most microbes cease cell division below 0.9, and few prokaryotic strains have been shown to continue cell division at aw smaller than 0.755 (Amina and Lotfi, 2024). However, the strains 5D5, 1A11, and 6E9 screened in water-restricted culture media were efficient in producing EPS in vitro.
EPS are high-molecular-weight carbohydrates attached to the external surface of bacteria, related to the formation of biofilms and the attachment of bacterial cells to surfaces, including plant roots and soil particles. Since EPS are hydrated compounds, including 97% water in a polymeric matrix, they can enhance growth and ensure the survival of plants under drought stress (Vu et al., 2009; Nocker et al., 2012). In addition, they can protect the plant against desiccation due to the formation of hydrophilic biofilms on the root surface (Rossi et al., 2012). Although EPS is the main constituent of the biofilm, the 1A11 strain did not show biofilm production in vitro. Since the presence of EPS and biofilm are closely related, the difference between the number of bacteria producing these two compounds can be explained by the need for other substances to be present for biofilm formation, such as extracellular proteins or molecules related to hydrophobicity, which makes it more or less soluble (Vlamakis et al., 2013; Zhang et al., 2015). The production of EPS by bacteria is important because it increases soil permeability and aggregation, maintaining high water potential near the roots (Alami et al., 2000), which facilitates the diffusion of nutrients and the mass flow of soluble substances, increasing plant absorption and therefore, promoting the mitigation of water deficit. Inoculation experiments of maize seeds with EPS-producing bacterial strains, in combination with their respective purified EPS, resulted in increased soil moisture content, plant biomass, root and shoot length, and leaf area (Zhang et al., 2015). The microenvironments provided by EPS retain water and dehydrate more slowly than the surrounding environment, thus protecting the rhizosphere from desiccation.
The Bacillus isolates in this study also produced Indole-3 acetic acid (IAA), ranging from 14.19 to 29.12 µg mL−1. IAA is a member of the group of phytohormones normally considered the most important native auxins. It has been frequently reported as an important mechanism for plant growth promotion, improving root size and distribution, and ensuring an increase in nutrient uptake from the soil. The production of IAA by bacteria differs between species and can vary between strains, but it is also influenced by culture conditions, availability of vitamins, salts, oxygen, pH value, temperature, growth rate, and available nitrogen source (Duca et al., 2014), while the tryptophan, an amino acid used as an IAA precursor, can be provided by the plant roots.
3.2 Comparison of treatment means in the greenhouse environment
To assess the ability of B. velezensis strains 5D5 and 6E9, and B. subtilis strain 1A11 to enhance growth in soybean plants in comparison to a control inoculated only with B. japonicum, an experiment using pots were carried out in a greenhouse with controlled environmental conditions. The experiments consisted of dose-range of the three inoculants, evaluating the effects of 1, 2, 3, and 4 mL of inoculant per kg of seeds. The experiment was divided into two groups: one with adequate moisture for optimal plant development and the other subjected to water restriction, simulating a drought episode.
Table 2 displays the response of soybean plants in terms of Shoot Fresh Weight (SFW), Shoot Dry Weight (SDW), Root Dry Weight (RDW), Number of Nodules per Root (NNR), and Mass of Functional Nodules per Root (MFNR) under irrigated and non-irrigated conditions. In both conditions, it was observed that the average treatment effects for certain inoculant doses were higher than the average of the control groups (T13 and T26 for irrigated and non-irrigated, respectively).
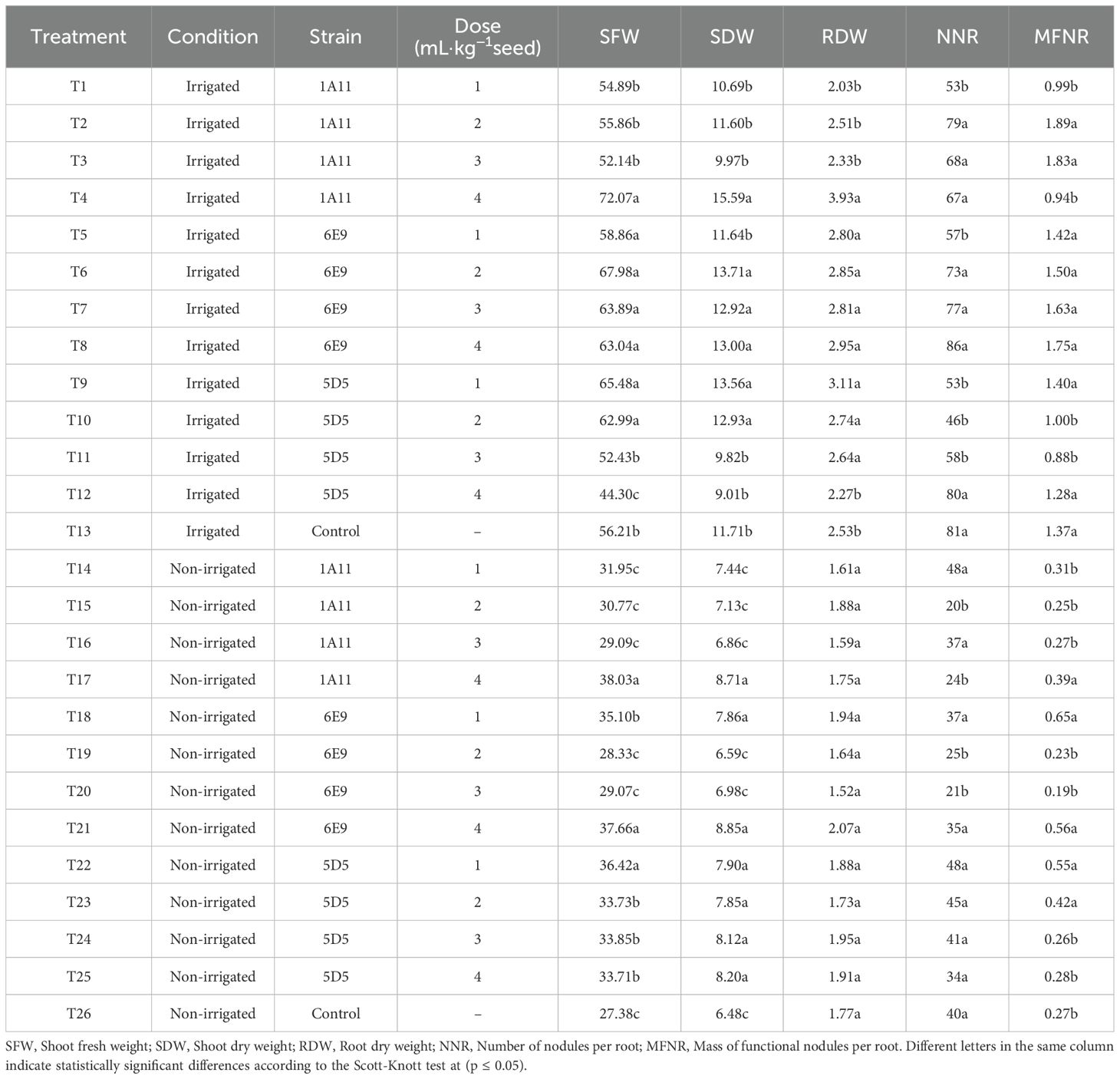
Table 2. Mean values of shoot and root biomass, number of nodules, and mass of functional nodules under irrigated and non-irrigated conditions.
Under irrigated conditions (T1 to T13), treatment T4 (1A11, 4 mL·kg−1) exhibited the highest mean shoot fresh weight (SFW), not differing significantly from treatments T5 (6E9, 1 mL·kg−1), T6 (6E9, 2 mL·kg−1), T7 (6E9, 3 mL·kg−1), T8 (6E9, 4 mL·kg−1), T9 (5D5, 1 mL·kg−1), and T10 (5D5, 2 mL·kg−1). All these treatments outperformed the non-inoculated control (T13). A similar pattern was observed for shoot dry weight (SDW) and root dry weight (RDW), where the same treatments, except T5 for SDW, showed significantly higher values compared to the control. In contrast, no significant improvements were observed in the number of nodules (NNR) and the mass of functional nodules (MFNR) in inoculated treatments compared to the control (T13).
Under non-irrigated conditions (T14 to T26), for the variable shoot fresh weight (SFW), the treatments T17 (1A11, 4 mL·kg−1), T21 (6E9, 4 mL·kg−1), and T22 (5D5, 1 mL·kg−1) exhibited the highest shoot fresh weight (SFW) means, significantly surpassing the non-inoculated control (T26). Similarly, treatments T18 (6E9, 1 mL·kg−1), T23 (5D5, 2 mL·kg−1), T24 (5D5, 3 mL·kg−1), and T25 (5D5, 4 mL·kg−1) showed intermediate performance, but still significantly higher than the control. For shoot dry weight (SDW), treatments T17 (1A11, 4 mL·kg−1), T18 (6E9, 1 mL·kg−1), T21 (6E9, 4 mL·kg−1), and all doses of 5D5 (T22 through T25) recorded superior means compared to the control. In contrast, no significant differences were observed for root dry weight (RDW), suggesting that root development was uniformly constrained under water-limited conditions. Regarding the number of nodules (NNR), none of the treatments outperformed the control. However, for the mass of functional nodules (MFNR), treatment T18 had the highest mean, statistically comparable to T17, T22, and T23, all of which were significantly superior to the control.
The results revealed a strain- and dose-dependent effect of inoculation on plant growth and nodulation parameters, with notable distinctions between irrigated and non-irrigated conditions. Under irrigation, strains 1A11 and 6E9 significantly enhanced shoot fresh weight (SFW) and the mass of functional nodules per root (MFNR), particularly at moderate to high doses. Strain 1A11 reached the highest SFW (72.07 g) at 4 mL·kg−1, while 6E9 showed consistent performance across doses. In contrast, strain 5D5 exhibited reduced performance at higher doses, suggesting a possible inhibitory effect, which may be attributed to microbial competition or metabolic burden on the host plant (Etesami, 2025). It is also important to highlight that each microbial strain possesses distinct mechanisms for promoting drought tolerance, such as the production of phytohormones, enzymes, and biofilm formation. Additionally, the applied dose influences the number of viable cells and spores available to colonize the seed, thereby affecting both the survival and colonization efficiency of the inoculant. Notably, very high or very low inoculum doses may not necessarily benefit the plant, as the concentration of bioactive compounds, such as phytohormones, may become either excessive or insufficient, potentially leading to suboptimal plant responses (Etesami, 2025).
Under non-irrigated conditions, although the overall performance of inoculated plants was reduced compared to irrigated treatments, certain inoculant-strain combinations still demonstrated potential to confer drought resilience. For instance, 6E9 at 1 mL·kg−1 improved SFW (35.10 g) and MFNR (0.65 g) relative to the non-inoculated control (27.38 g and 0.27 g, respectively), and 5D5 at the same dose yielded the highest SFW (36.42 g) under drought. These results suggest that low-dose inoculation with well-adapted strains can mitigate drought stress effects, likely through improved symbiotic efficiency and modulation of root architecture, as previously documented in studies on plant-microbe interactions under water-limited conditions (Timmusk et al., 2014; Ngumbi and Kloepper, 2016; Vurukonda et al., 2016). In contrast, 1A11 enhanced SFW (38.03 g) and SDW (8.71 g), relative to the non-inoculated control under drought condition (27.38 g and 6.48 g, respectively).
Among the evaluated variables, SFW, SDW, and MFNR were the most positively responsive to inoculation, highlighting their utility as indicators of inoculant performance under contrasting moisture regimes. These findings reinforce the importance of selecting compatible strain-dose combinations and support the strategic use of plant growth-promoting bacteria to enhance legume productivity in both irrigated and drought-prone environments.
3.3 The impact of field weather on soybean yield
Abrupt and extreme climate variations have become one of the main challenges for agricultural practices, particularly in Brazil, a tropical country with a strong agricultural tradition. Regular planting and cultivation seasons are changing due to shifts in rainfall patterns, as well as variations in average maximum and minimum temperatures (Dos Santos et al., 2023). These changes increase the level of risk and make agricultural activities more unpredictable.
To mitigate these effects and identify their occurrence, farmers are adopting technological alternatives as part of their strategy to reduce losses and ensure economic returns. In this context, microbiological inoculants that improve plant resilience during mild to moderate water deficits—such as the bacterial strains evaluated in this study—are fundamental for maintaining crop grain yield. Some products are already available on the market, including those aimed at increasing phosphorus solubilization (de Oliveira-Paiva et al., 2024) and microbial-based solutions to reduce the effects of drought (Kavamura et al., 2013).
Additionally, incorporating technologies like remote sensing with time-series satellite or drone images enables producers to track in advance the environmental impacts on crop development. This allows them to take proactive palliative or corrective measures.
During the field-trials with soybeans, significant variations were observed in rainfall patterns and temperature peaks (Figure 1). In the 2022–2023 season, rainfall was more regularly distributed and of a greater volume compared to the 2023–2024 season. The maximum temperatures recorded during 2022–2023 generally stayed below 35°C, whereas during 2023–2024, temperatures frequently exceeded 40°C, negatively affecting the development of soybean crops.
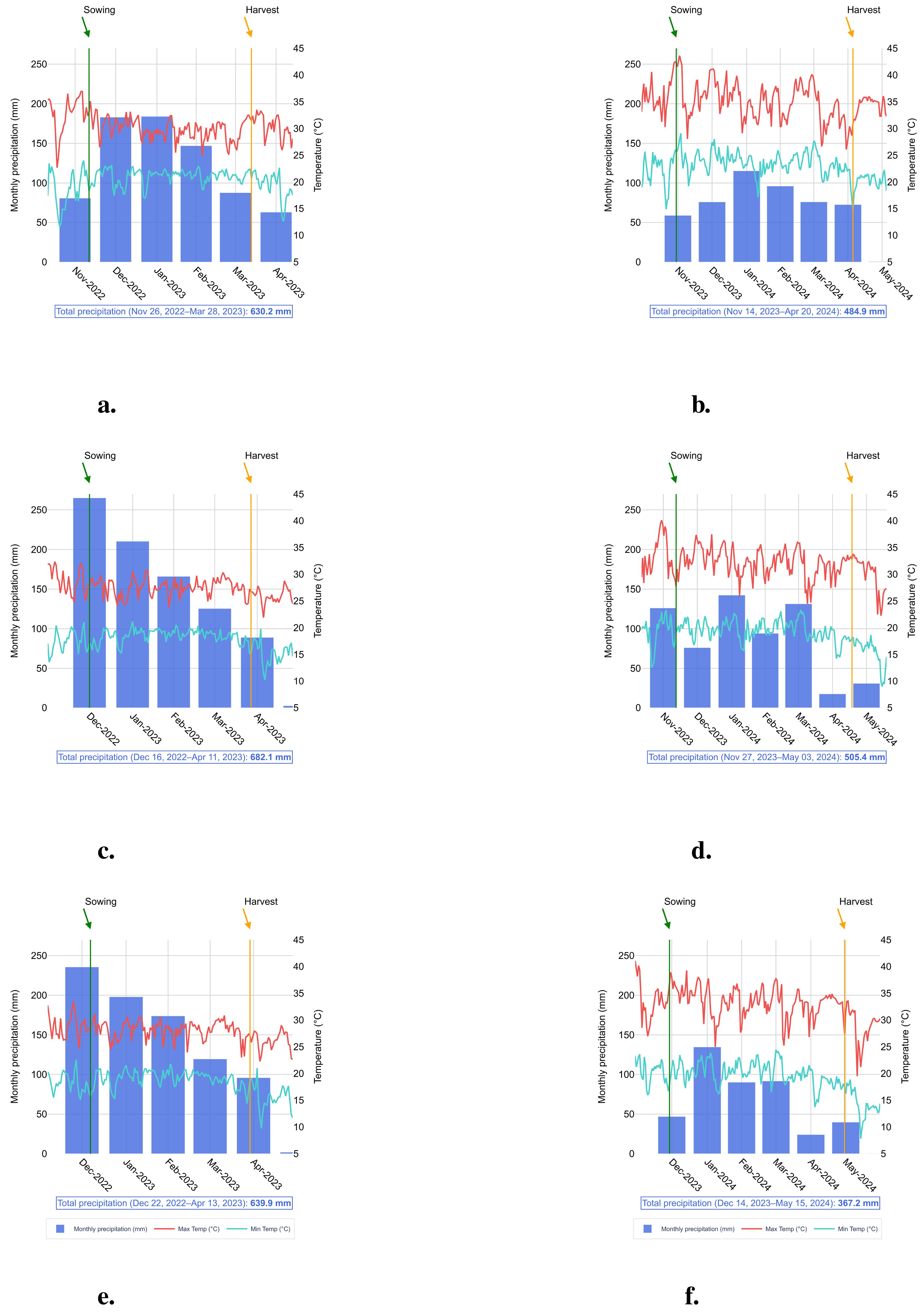
Figure 1. Monthly accumulated precipitation (mm), maximum temperature (°C), and minimum temperature (°C) during the experiment period. Locations, seasons and precipitation: (a) Birigui, season 2022/2023 and total accumulated precipitation of 630.2 mm, (b) Birigui, season 2023/2024 and total accumulated precipitation of 484.9 mm, (c) Itapira, season 2022/2023 and total accumulated precipitation of 682.1 mm, (d) Itapira, season 2023/2024 and total accumulated precipitation of 505.4 mm, (e) Piracicaba, season 2022/2023 and total accumulated precipitation of 639.9 mm, and (f) Piracicaba, season 2023/2024 and total accumulated precipitation of 367.2 mm.
As a result, the 2022–2023 harvest yielded an average of nearly one ton more than the 2023–2024 harvest average across the three locations involved in the study (Figure 2c). This clearly indicates the detrimental effects of climate extremes on soybean production. The analysis also revealed that Itapira had slightly higher average grain yield than Birigui and Piracicaba, likely due to better rainfall averages, especially durin7g the 2023–2024 season (Figure 2b). Beyond climate variations across locations (Figure 1), the differences in environmental conditions—particularly soil physicochemical properties—and the distinct genetic backgrounds of the three soybean varieties may also influence the results. This is worth mentioning that the three soybean varieties utilized in this study are categorized into maturity groups ranging from 6.1 (‘NS 5933 iPro’) and 6.4 (‘BMX Nexus i2X’) to 6.8 (‘Monsoy 6601 i2X’). Therefore, these varieties are suitable for the southeastern region of Brazil, where the field trials were carried out, and it also provided a wider planting window for the locations. This flexibility allowed to plant later in the season, exposing the crops to a period of lower rainfall and reduced soil humidity, and thus evaluate the effect of water shortage on the Bacillus/soybean crop association under conditions that are appropriate to water deficit.
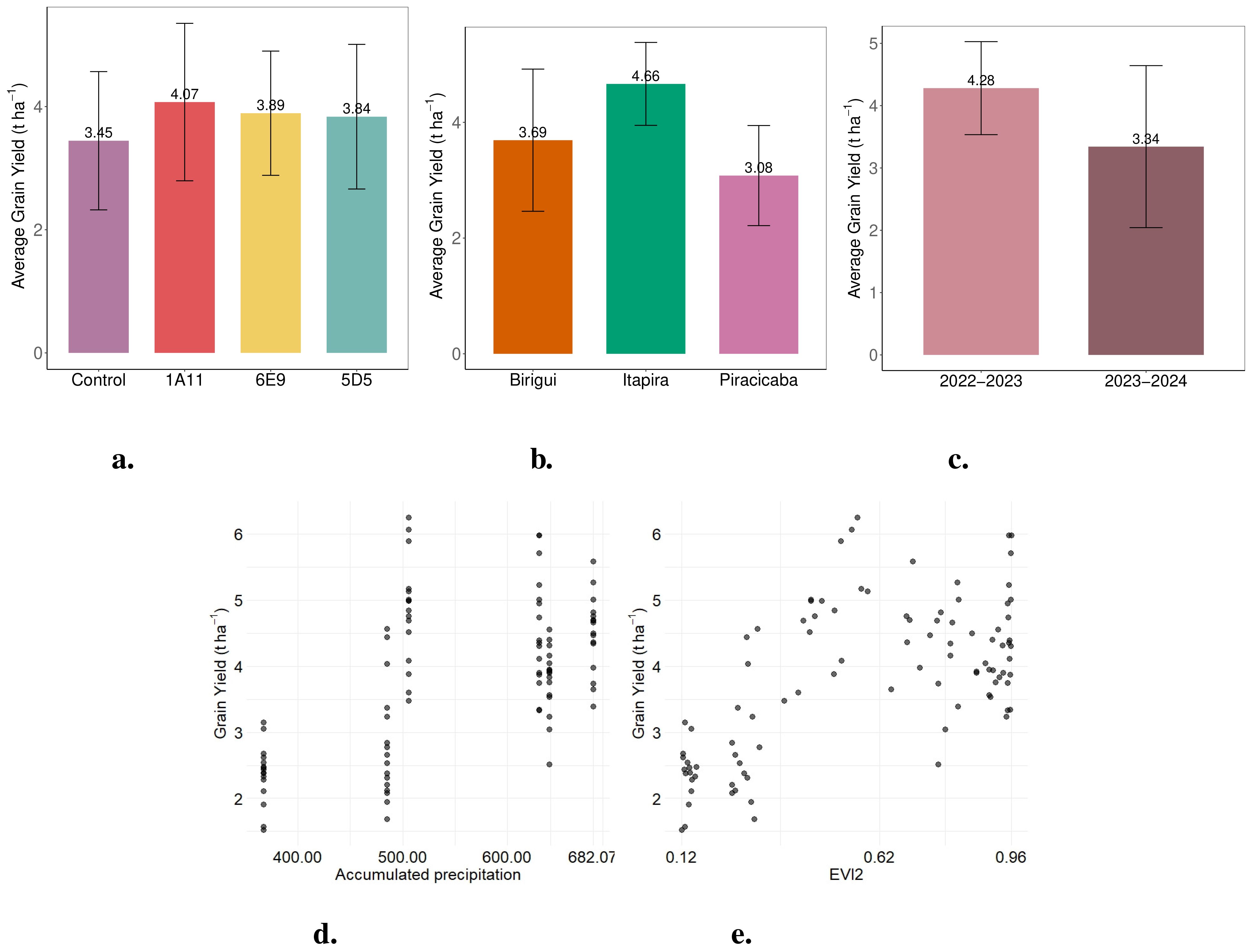
Figure 2. Bar plots of average grain yield (t ha−1) from the field trials: (a) by treatment, which received a dose of 3 mL·kg−1 of seeds, (b) by location, and (c) by crop year (2022–2023 and 2023–2024). Scatter plots from the field experiment: (d) cumulative precipitation and (e) mean EVI2 during the crop growth period versus grain yield (t ha−1).
3.4 Descriptive analysis of field experiments
Figure 2a shows that microbial strain 1A11 achieved the highest soybean yield across the two growing seasons and three tested locations, producing an average of 620 kg ha−1 more than the control (an 18% increase). Strains 5D5 and 6E9 also led to yield improvements over the control, with 5D5 yielding 390 kg ha−1 (11.3% increase) and 6E9 yielding 440 kg ha−1 (12.8% increase). These results suggest that the application of these treatments resulted in yield gains relative to the control.
Among the locations, Itapira achieved the highest average yield (4.66 t ha−1), followed by Birigui (3.69 t ha−1) and Piracicaba (3.08 t ha−1), as shown in Figure 2b. Furthermore, significant differences in yield were observed between the two growing seasons, with 2022–2023 (4.28 t ha−1) outperforming 2023–2024 (3.34 t ha−1), as shown in Figure 2c. According to Figure 2, increased cumulative precipitation (Figure 2d) and higher EVI2 (Figure 2e) values are associated with indications of greater soybean grain yield, suggesting that these variables may have a positive influence on crop yield.
Table 3 summarizes the mean and standard deviation of grain yield (t ha−1) and the EVI2 index for different locations, treatments, and years. The data cover two seasons (2022–2023 and 2023–2024) and three locations: Birigui, Itapira, and Piracicaba. For each location and year, values for four treatments—1A11, 5D5, 6E9, and control—are provided. In 2022–2023 harvest, grain yield means ranged from 3.58 t ha−1 in the control treatment at Birigui to 5.14 t ha−1 in treatment 1A11 at the same location. EVI2 values were relatively stable across all treatments and locations in 2022–2023 season, with mean values close to 0.95. In 2023–2024 harvest, a noticeable decrease in grain yield was observed at most locations and treatments, particularly at Piracicaba, where the control treatment showed the lowest grain yield (1.78 t ha−1). EVI2 values remained consistent across treatments, with some variations, especially in Itapira and Birigui. These results reflect both grain yield levels and vegetation dynamics (as indicated by EVI2) across the two seasons.
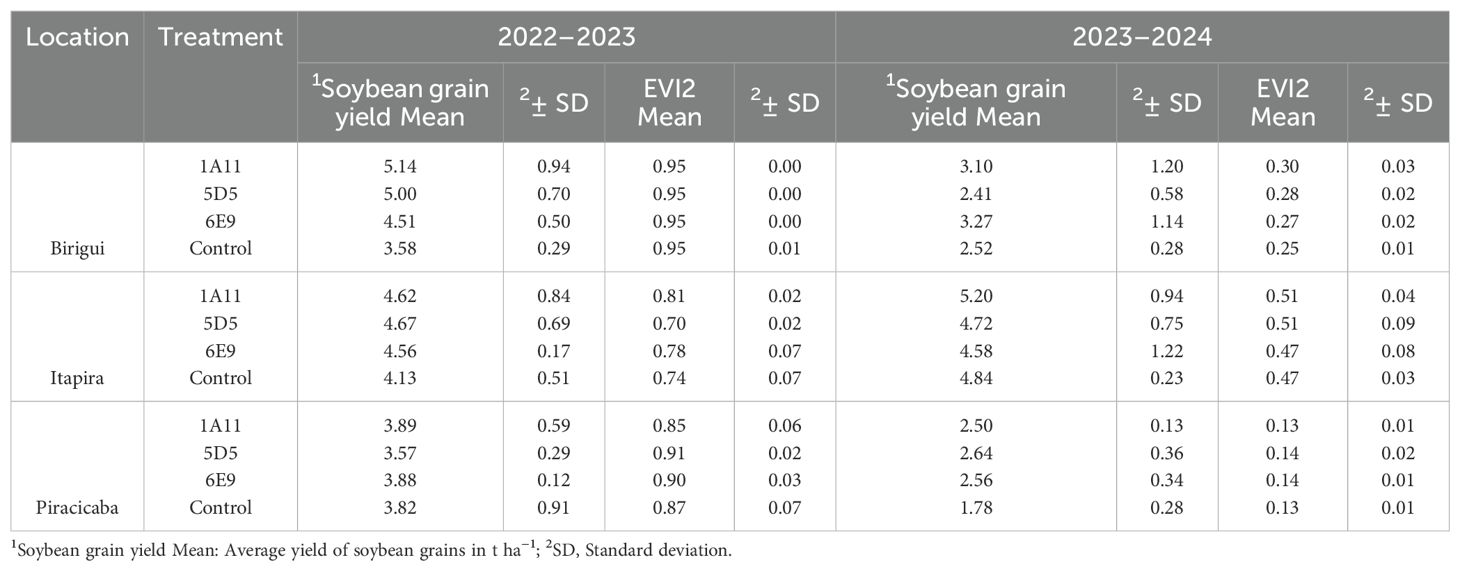
Table 3. Mean and standard deviation of soybean grain yield (t ha−1) and Enhanced Vegetation Index 2 (EVI2) by location, treatment, and year under field conditions.
Additionally, Table 3 highlights that treatments with strains 1A11, 5D5, and 6E9 performed better in terms of EVI2 compared to the control. However, in Birigui during the 2022–2023 season, all treatments showed the same average EVI2 values, which were close to the maximum of the EVI2 scale (0.95), suggesting potential saturation under these specific conditions.
The high EVI2 values in Birigui during 2022–2023 may be attributed to the greater canopy development of soybean plants in all treatment groups, compared to other locations or years. Environmental variations in open field conditions likely contributed to these discrepancies.
3.5 Statistical modeling of field experiments
Based on the multicollinearity analysis, the explanatory variable Year (xi4) was excluded due to its high VIF in relation to the other covariates. The final model, without this variable, was fitted to represent the relationship between the dependent variable yi and the selected explanatory variables. The model equation is given by:
This revised model, excluding the variable with high multicollinearity, results in a more robust and precise selection of explanatory variables.
The results of the statistical model (Table 4) indicated that treatments 1A11, 5D5, and 6E9 showed significant differences compared to the control (represented by the intercept), with statistical significance determined by p-values less than 0.10, however, these treatments did not differ from each other (Table 5). These findings highlight the efficacy of strains 1A11, 5D5, and 6E9 in enhancing soybean development under non-irrigated, open-field conditions. The observed grain yield gains were statistically significant compared to the not inoculated control, underscoring the potential of microbial inoculants to improve crop performance even in challenging agricultural environments.
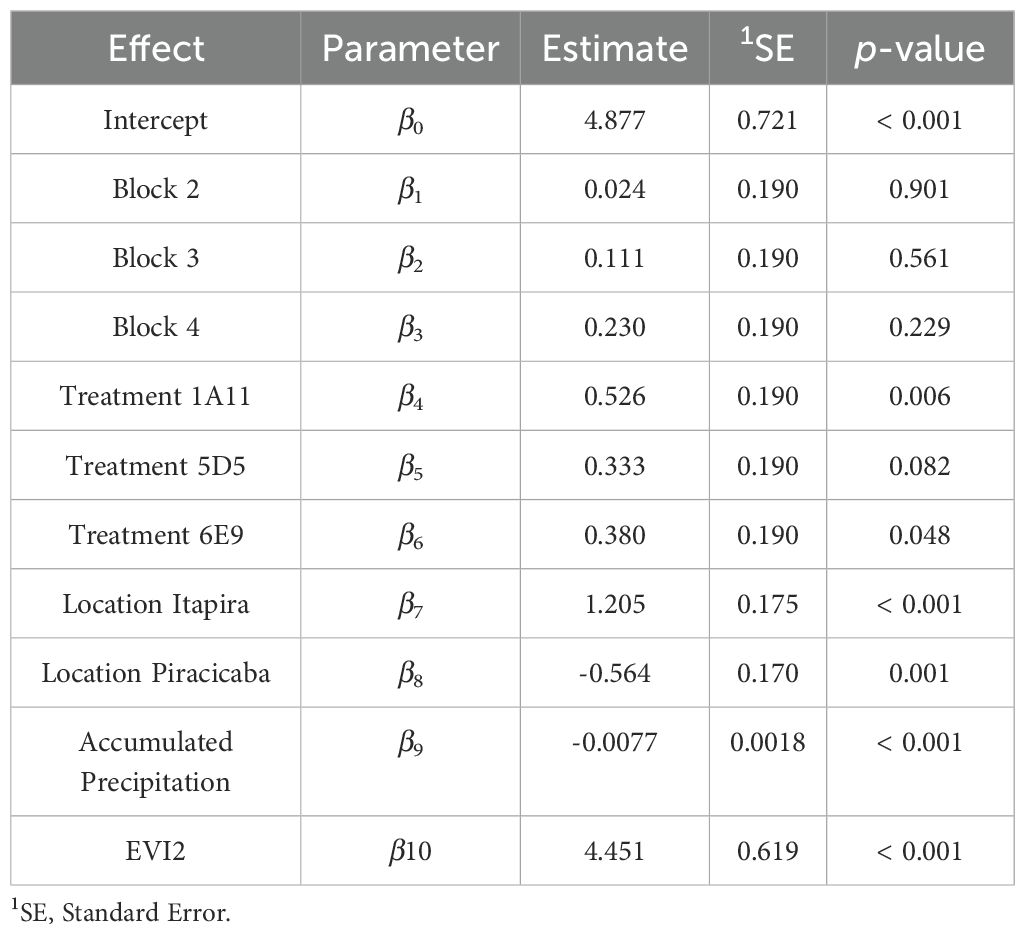
Table 4. Estimated coefficients for the fitted model using field experiment data.
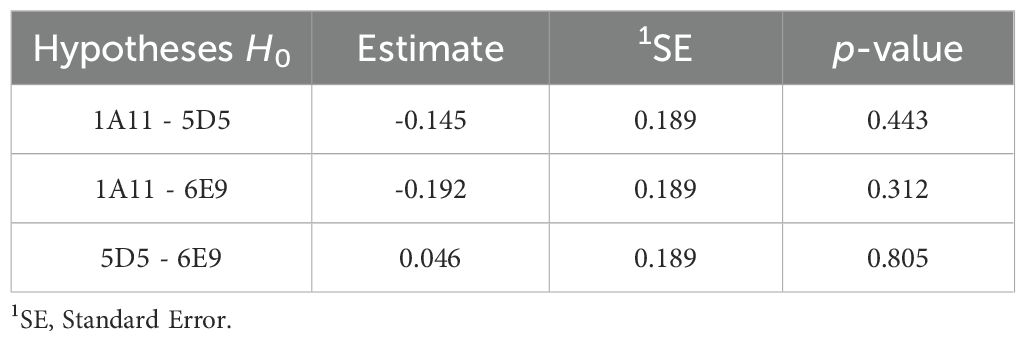
Table 5. Results of multiple comparisons of treatment considering regression model.
Regarding location, Itapira showed significantly higher average grain yield compared to the other sites, as illustrated in Figure 2b. The bar plot indicates that the mean yield in Itapira was greater than in other locations, particularly when compared to Birigui. This difference may reflect environmental and management variability between locations and the availability of rainwater throughout the crop cycle, with the rainfall regime being more favorable in some of the locations where the tests were carried out than in others (Figure 1).
As for cumulative precipitation, the model results indicate that this variable was statistically significant. However, excessive precipitation appears to be associated with a slight reduction in soybean grain yield. This trend is evident in Figure 2d, which shows a small decrease in grain yield at the highest levels of accumulated rainfall during the early stages of the experiment.
Finally, the EVI2 index showed a significant positive effect on grain yield: according to the fitted model, for each one-unit increase in EVI2, soybean yield increased by an average of 4.45 t/ha. It is worth noting that, as the crop approaches its final growth stages, a natural decline in vegetation indices is expected due to plant maturation and senescence.
The half-normal plot (Moral et al., 2017) with simulated envelope (Figure 3a) showed that all standardized residuals fell within the envelope. This suggests that the linear model provides an adequate fit to the data, with residuals behaving as expected under the assumptions of normality and homoscedasticity.
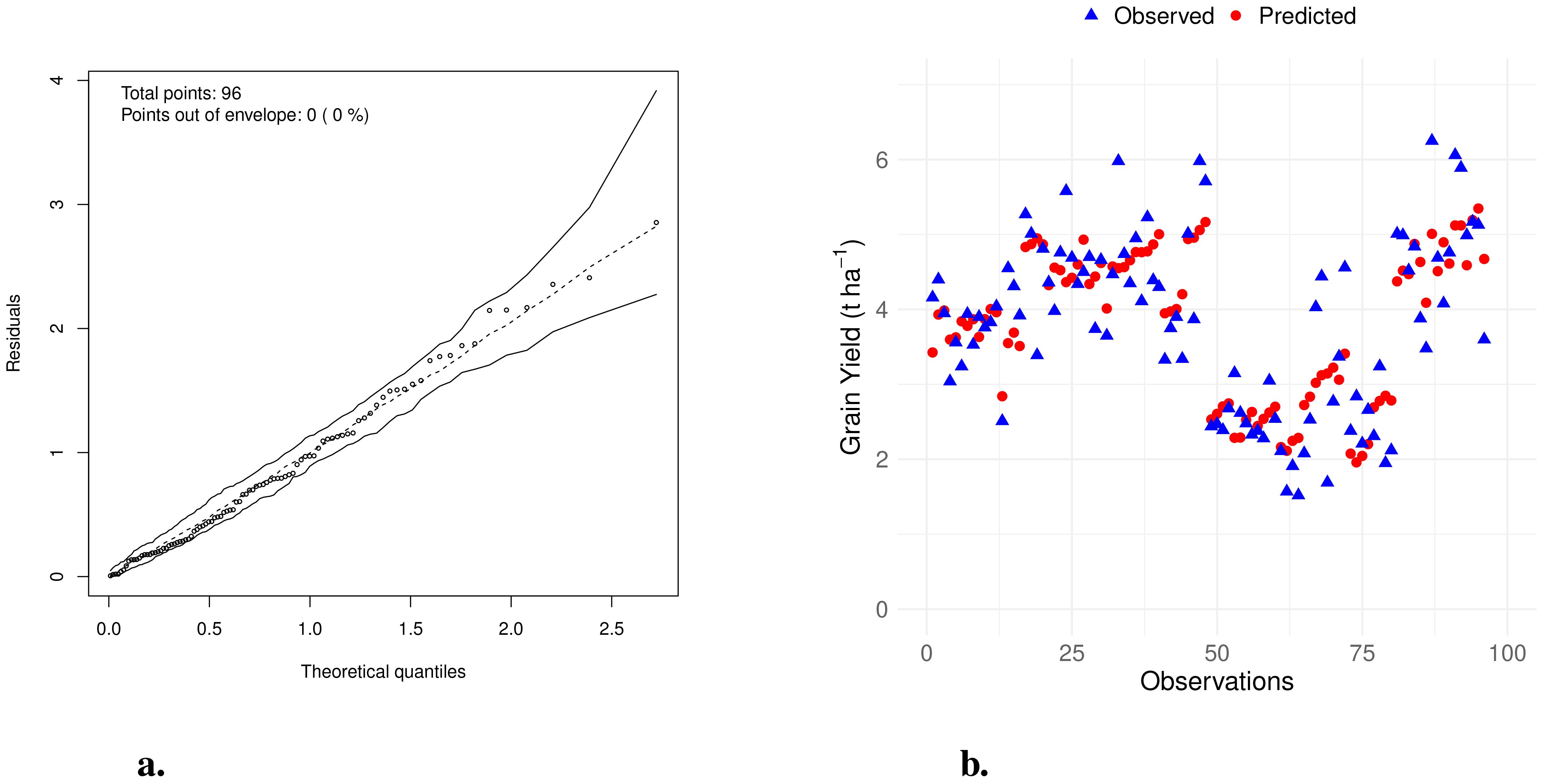
Figure 3. (a) Half-normal plot with simulated envelope for the regression model. (b) Observed vs. predicted values from the multiple linear regression model for soybean grain yield.
The Shapiro-Wilk test yielded W = 0.9898 with a p-value of 0.6756, and the Breusch-Pagan test produced BP = 12.508 with df = 10 and a p-value of 0.2525. Both tests were non-significant, indicating that the residuals are normally distributed and exhibit constant variance, thus supporting the adequacy of the linear model.
Additionally, the model achieved a coefficient of determination (R2) of 0.71, indicating that approximately 71% of the variability in soybean grain yield was explained by the included explanatory variables.
Figure 3b illustrates the comparison between observed and predicted grain yield values obtained from the multiple linear regression model. The close alignment between observed and predicted points demonstrates that the model effectively captures the variability in soybean grain yield. In the figure, blue triangles represent the observed values, while red circles indicate the predicted values. The overlapping of these points reflects the natural variability in the data and supports the model’s accuracy in predicting grain yield.
Research utilizing remote sensing tools to study microbiological inoculants is surprisingly limited in the literature, revealing a significant gap but also presenting opportunities to improve the assessment of these inoculants’ effectiveness on agronomic crop performance. For example, de Souza et al. (2022) conducted a study on common beans, using UAVs to evaluate and monitor the biofertilization process. They applied VIs to map spectral changes in common beans after the application of phosphate-solubilizing bacteria in a remote and non-destructive manner. Similarly, Santos et al. (2024) investigated phosphate-solubilizing bacterial inoculants in sugarcane crops developed a predictive statistical model that accurately forecasted sugarcane grain yield in fields treated with these microbiological inoculants, producing predictions months ahead of harvest that closely aligned with actual field data.
4 Conclusion
This study demonstrates that B. velezensis (5D5, 6E9) and B. subtilis (1A11) microbiological inoculants significantly enhance soybean resilience and grain yield under drought stress, validating their role as scalable, sustainable tools for climate-smart agriculture. The Bacillus strains 1A11, 5D5 and 6E9, isolated from arid soils, showed multifunctional mechanisms for plant growth-promoting (PGP) related to drought stress tolerance, particularly those related to exopolysaccharide (EPS) and biolfim production and capacity of growing in a low water activity. Greenhouse trials revealed that doses of microbial biostimulants above 2 mL Kg−1 of seed application were able to increase biomass accumulation, with inoculated plants under drought stress surpassing non-inoculated controls. Field-trials across three Brazilian agroclimatic zones under non-irrigated conditions further confirmed the grain yield superiority of Bacillus of 1A11, 5D5 and 6E9 compared to the control treatment without seed inoculation. These results position microbial biostimulants as critical alternatives to synthetic inputs, aligning with Brazil’s strategic priorities to reduce agrochemical dependency while safeguarding food security. The integration of satellite and UAV-derived EVI2 allowed for precise, non-destructive monitoring of treatment effectiveness, ensuring a consistent overlap between field data and model predictions. This dual assessment framework—combining agronomic metrics with remote sensing—provides a novel, cost-effective strategy for real-time crop performance evaluation.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
JV: Writing – original draft, Writing – review & editing, Investigation, Formal Analysis, Methodology, Data curation. CA: Formal Analysis, Data curation, Writing – original draft, Investigation. EG: Writing – original draft, Formal Analysis, Data curation, Conceptualization, Methodology, Investigation, Writing – review & editing, Project administration, Funding acquisition. CO: Investigation, Formal Analysis, Conceptualization, Writing – review & editing, Writing – original draft, Methodology. SS: Writing – review & editing, Methodology, Writing – original draft, Investigation, Conceptualization, Formal Analysis. ES: Writing – original draft, Writing – review & editing, Formal Analysis, Methodology, Data curation, Investigation. JA: Methodology, Formal Analysis, Writing – review & editing, Investigation, Writing – original draft. UL: Methodology, Writing – original draft, Formal Analysis, Data curation, Writing – review & editing, Investigation. GC: Formal Analysis, Writing – original draft, Methodology, Project administration, Conceptualization, Validation, Resources, Investigation, Supervision, Writing – review & editing, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded through collaborative grants from the Brazilian Agricultural Research Corporation (Embrapa) and the São Paulo State Sugarcane Growers’ Cooperative (Coplacana) [Grant SEG 30.19.90.005.00.00], as well as, the Simbiose Company and the Brazilian Funding Authority for Studies and Projects (Finep) [Grant SEG 20.24.00.110.00.00]. The funders and companies had no role in study design, data access, data collection/analysis, or publication decisions. Julio Cezar Souza Vasconcelos and Caio Simplicio Arantes are fellows of the research project that funded the study and receive a grant from an Embrapa foundation that administers the project's resources (FAPED). The other authors are affiliated with Embrapa, a public research company.
Acknowledgments
The authors thank A.S.P. Junior, G. Medeiros, R. Klaic, D. Christofoletti, F. J. Severino, and L.A.F. Barbosa for their assistance during the research execution. They also thank RedeMAIS/MJSP for providing the satellite images from PlanetScope, and Coplacana for their invaluable support throughout the field experiments.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alami, Y., Achouak, W., Marol, C., and Heulin, T. (2000). Rhizosphere soil aggregation and plant growth promotion of sunflowers by an exopolysaccharide-producing rhizobium sp. strain isolated from sunflower roots. Appl. Environ. Microbiol. 66, 3393–3398. doi: 10.1128/AEM.66.8.3393-3398.2000
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped blast and psi-blast: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Amaral, L. R., Oldoni, H., Baptista, G. M., Ferreira, G. H., Freitas, R. G., Martins, C. L., et al. (2024). Remote sensing imagery to predict soybean yield: a case study of vegetation indices contribution. Precis. Agric. 25, 2375–2393. doi: 10.1007/s11119-024-10174-5
Amina, M. and Lotfi, G. (2024). An overview of extremophile: Microbial diversity, adaptive strategies, and potential applications. Microbiol. Biotechnol. Lett. 52, 233–254. doi: 10.48022/mbl.2406.06007
Arora, N. K. and Verma, M. (2017). Modified microplate method for rapid and efficient estimation of siderophore produced by bacteria. 3 Biotech. 7, 381. doi: 10.1007/s13205-017-1008-y
Azeem, M., Javed, S., and Zahoor, A. (2023). Bacillus species as potential plant growth promoting rhizobacteria for drought stress resilience. Russian J. Plant Physiol. 70, 59. doi: 10.1134/S1021443723600538
Bahadir, P. S., Liaqat, F., and Eltem, R. (2018). Plant growth promoting properties of phosphate solubilizing bacillus species isolated from the aegean region of Turkey. Turkish J. Bot. 42, 183–196. doi: 10.3906/bot-1706-51
Bégué, A., Arvor, D., Bellon, B., Betbeder, J., De Abelleyra, D., PD Ferraz, R., et al. (2018). Remote sensing and cropping practices: A review. Remote Sens. 10, 99. doi: 10.3390/rs10010099
Belsley, D. A., Kuh, E., and Welsch, R. E. (1980). Regression Diagnostics: Identifying Influential Data and Sources of Collinearity (New York, USA: John Wiley & Sons).
Bettiol, W. (2022). Pesquisa, desenvolvimento e inovação com bioinsumos. Eds. Meyer, M. C., Bueno, A. F., and Mazaro, S. M., (Brazilian, DF, Brazil: Embrapa) 21–28.
Breusch, T. S. and Pagan, A. R. (1979). A simple test for heteroscedasticity and random coefficient variation. Econometrica 47, 1287–1294. doi: 10.2307/1911963
Chambers, J. M. (1992). “Linear models. Chap. 4,” in Statistical Models in S. Eds. Chambers, J. M. and Hastie, T. J. (Wadsworth & Brooks/Cole, Pacific Grove, CA), 95–144.
de Oliveira-Paiva, C. A., Bini, D., de Sousa, S. M., Ribeiro, V. P., Dos Santos, F. C., de Paula Lana, U. G., et al. (2024). Inoculation with bacillus megaterium cnpms b119 and bacillus subtilis cnpms b2084 improve p-acquisition and maize yield in Brazil. Front. Microbiol. 15, 1426166. doi: 10.3389/fmicb.2024.1426166
de Souza, A. E. S., Barbosa Júnior, M. R., de Almeida Moreira, B. R., da Silva, R. P., and Lemos, L. B. (2022). Uav multispectral data: A reliable approach for managing phosphate-solubilizing bacteria in common bean. Agronomy 12, 2284. doi: 10.3390/agronomy12102284
Döbereiner, J. (1997). Biological nitrogen fixation in the tropics: social and economic contributions. Soil Biol. Biochem. 29, 771–774. doi: 10.1016/S0038-0717(96)00226-X
Dos Santos, E., Fortini, R., Cardoso, L., and Zanuncio, J. (2023). Climate change in Brazilian agriculture: vulnerability and adaptation assessment. Int. J. Environ. Sci. Technol. 20, 10713–10730. doi: 10.1007/s13762-022-04730-7
Dubey, A., Saiyam, D., Kumar, A., Hashem, A., Abd Allah, E. F., and Khan, M. L. (2021). Bacterial root endophytes: Characterization of their competence and plant growth promotion in soybean (glycine max (l.) merr.) under drought stress. Int. J. Environ. Res. Public Health 18, 931. doi: 10.3390/ijerph18030931
Duca, D., Lorv, J., Patten, C. L., Rose, D., and Glick, B. R. (2014). Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek 106, 85–125. doi: 10.1007/s10482-013-0095-y
Etesami, H. (2025). The dual nature of plant growth-promoting bacteria: Benefits, risks, and pathways to sustainable deployment. Curr. Res. Microb. Sci. 9, 100421. doi: 10.1016/j.crmicr.2025.100421
Frazier, A. E. and Hemingway, B. L. (2021). A technical review of planet smallsat data: Practical considerations for processing and using planetscope imagery. Remote Sens. 13, 3930. doi: 10.3390/rs13193930
Guimarães, D. P., Costa, F., Rodrigues, M., and Maugeri, F. (1999). Optimization of dextran syntesis and acidic hydrolisis by surface response analysis. Braz. J. Chem. Eng. 16, 129–139. doi: 10.1590/S0104-66321999000200004
Hodkinson, B. P. and Lutzoni, F. (2009). A microbiotic survey of lichen-associated bacteria reveals a new lineage from the rhizobiales. Symbiosis 49, 163–180. doi: 10.1007/s13199-009-0049-3
Hu, J., Yue, J., Xu, X., Han, S., Sun, T., Liu, Y., et al. (2023). Uav-based remote sensing for soybean fvc, lcc, and maturity monitoring. Agriculture 13, 692. doi: 10.3390/agriculture13030692
Hungria, M. and Mendes, I. C. (2015). Nitrogen fixation with soybean: the perfect symbiosis? Biol. Nitrogen Fixation 2, 1009–1024. doi: 10.1002/9781119053095.ch99
Hungria, M. and Nogueira, M. A. (2019). Tecnologias de inoculação da cultura da soja: Mitos, verdades e desafios (Rondonópolis: Fundação MT), 50–62.
Jiang, Z., Huete, A. R., Didan, K., and Miura, T. (2008). Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ. 112, 3833–3845. doi: 10.1016/j.rse.2008.06.006
Joshi, D. R., Clay, S. A., Sharma, P., Rekabdarkolaee, H. M., Kharel, T., Rizzo, D. M., et al. (2024). Artificial intelligence and satellite-based remote sensing can be used to predict soybean (glycine max) yield. Agron. J. 116, 917–930. doi: 10.1002/agj2.21473
Kavamura, V. N., Santos, S. N., da Silva, J. L., Parma, M. M., Ávila, L. A., Visconti, A., et al. (2013). Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol. Res. 168, 183–191. doi: 10.1016/j.micres.2012.12.002
Khan, W., Zhu, Y., Khan, A., Zhao, L., Yang, Y.-M., Wang, N., et al. (2024). Above-and below-ground feedback loop of maize is jointly enhanced by plant growth-promoting rhizobacteria and arbuscular mycorrhizal fungi in drier soil. Sci. Total Environ. 917, 170417. doi: 10.1016/j.scitotenv.2024.170417
Kondo, T., Sibponkrung, S., Tittabutr, P., Boonkerd, N., Ishikawa, S., Teaumroong, N., et al. (2025). Bacillus velezensis s141 improves the root growth of soybean under drought conditions. Biosci. Biotechnol. Biochem. 89, 304–312. doi: 10.1093/bbb/zbae168
Kuramshina, Z. and Khairullin, R. (2023). Endophytic strains of bacillus subtilis promote drought resistance of plants. Russian J. Plant Physiol. 70, 45. doi: 10.1134/S1021443722603172
Liu, K., Deng, F., Zeng, F., Chen, Z.-H., Qin, Y., and Chen, G. (2025). Plant growth-promoting rhizobacteria improve drought tolerance of crops: a review. Plant Growth Regul. 105, 567–581. doi: 10.1007/s10725-025-01300-y
Loper, J. and Schroth, M. (1986). Influence of bacterial sources of indole-3-acetic acid on root elongation of sugar beet. Phytopathology 76, 386–389. doi: 10.1094/Phyto-76-386
Monteiro, L. A., Sentelhas, P. C., and Pedra, G. U. (2018). Assessment of nasa/power satellite-based weather system for Brazilian conditions and its impact on sugarcane yield simulation. Int. J. Climatol. 38, 1571–1581. doi: 10.1002/joc.5282
Moral, R. A., Hinde, J., and Demétrio, C. G. B. (2017). Half-normal plots and overdispersed models in r: the hnp package. J. Stat. Softw. 81, 1–23. doi: 10.18637/jss.v081.i10
NASA POWER Project (2024). POWER Data Access Viewer [Dataset]. (NASA Langley Research Center, POWER Project). Available online at: https://power.larc.nasa.gov/data-access-viewer/ (Accessed October 30, 2024).
Ngumbi, E. and Kloepper, J. (2016). Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 105, 109–125. doi: 10.1016/j.apsoil.2016.04.009
Nocker, A., Fernández, P. S., Montijn, R., and Schuren, F. (2012). Effect of air drying on bacterial viability: a multiparameter viability assessment. J. Microbiol. Methods 90, 86–95. doi: 10.1016/j.mimet.2012.04.015
O’Callaghan, M., Ballard, R. A., and Wright, D. (2022). Soil microbial inoculants for sustainable agriculture: Limitations and opportunities. Soil Use Manage. 38, 1340–1369. doi: 10.1111/sum.12811
Ozdogan, M., Yang, Y., Allez, G., and Cervantes, C. (2010). Remote sensing of irrigated agriculture: Opportunities and challenges. Remote Sens. 2, 2274–2304. doi: 10.3390/rs2092274
Pandey, P. K., Singh, S., Singh, A. K., Samanta, R., Yadav, R. N. S., and Singh, M. C. (2016). Inside the plant: Bacterial endophytes and abiotic stress alleviation. J. Appl. Natural Sci. 8, 1899–1904. doi: 10.31018/jans.v8i4.1059
Park, D., Jang, J., Seo, D. H., Kim, Y., and Jang, G. (2024). Bacillus velezensis gh1–13 enhances drought tolerance in rice by reducing the accumulation of reactive oxygen species. Front. Plant Sci. 15, 1432494. doi: 10.3389/fpls.2024.1432494
Patten, C. L. and Glick, B. R. (1996). Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 42, 207–220. doi: 10.1139/m96-032
Paulo, E. M., Vasconcelos, M. P., Oliveira, I. S., Affe, H. M. d. J., Nascimento, R., Melo, I., et al. (2012). An alternative method for screening lactic acid bacteria for the production of exopolysaccharides with rapid confirmation. Food Sci. Technol. 32, 710–714. doi: 10.1590/S0101-20612012005000094
Prince Czarnecki, J. M., Samiappan, S., Bheemanahalli, R., Huang, Y., and Shammi, S. A. (2025). A brief history of remote sensing of soybean. Agron. J. 117, e70004. doi: 10.1002/agj2.70004
R Core Team (2024). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing).
Ren, P., Zhou, B., Bi, Y., Chen, X., Yao, S., and Yang, X. (2025). Bacillus subtilis can promote cotton phenotype, yield, nutrient uptake and water use efficiency under drought stress by optimizing rhizosphere microbial community in arid area. Ind. Crops Prod. 227, 120784. doi: 10.1016/j.indcrop.2025.120784
Rho, H., Hsieh, M., Kandel, S. L., Cantillo, J., Doty, S. L., and Kim, S.-H. (2018). Do endophytes promote growth of host plants under stress? a meta-analysis on plant stress mitigation by endophytes. Microb. Ecol. 75, 407–418. doi: 10.1007/s00248-017-1054-3
Ribeiro, V. P., Marriel, I. E., Sousa, S., Lana, P. U., Mattos, B. B., Oliveira, C., et al. (2018). Endophytic bacillus strains enhance pearl millet growth and nutrient uptake under low-p. Braz. J. Microbiol. 49, 40–46. doi: 10.1016/j.bjm.2018.06.005
Rossi, F., Micheletti, E., Bruno, L., Adhikary, S. P., Albertano, P., and De Philippis, R. (2012). Characteristics and role of the exocellular polysaccharides produced by five cyanobacteria isolated from phototrophic biofilms growing on stone monuments. Biofouling 28, 215–224. doi: 10.1080/08927014.2012.663751
Santos, D., Soares, A., de Medeiros, G., Christofoletti, D., Arantes, C. S., Vasconcelos, J. C. S., et al. (2024). Evaluation of sugarcane yield response to a phosphate-solubilizing microbial inoculant: Using an aerial imagery-based model. Sugar Tech 26, 143–159. doi: 10.1007/s12355-023-01326-4
Sarma, R. K. and Saikia, R. (2014). Alleviation of drought stress in mung bean by strain pseudomonas aeruginosa ggrj21. Plant Soil 377, 111–126. doi: 10.1007/s11104-013-1981-9
Schmidt, J., Messmer, M., and Wilbois, K.-P. (2015). Beneficial microorganisms for soybean (glycine max (l.) merr), with a focus on low root-zone temperatures. Plant Soil 397, 411–445. doi: 10.1007/s11104-015-2546-x
Shapiro, S. S. and Wilk, M. B. (1965). An analysis of variance test for normality (complete samples). Biometrika 52, 591–611. doi: 10.2307/2333709
Singh, J. S., Pandey, V. C., and Singh, D. P. (2011). Efficient soil microorganisms: a new dimension for sustainable agriculture and environmental development. Agric. Ecosyst. Environ. 140, 339–353. doi: 10.1016/j.agee.2011.01.017
Stepanović, S., Vuković, D., Hola, V., Bonaventura, G. D., Djukić, S., Ćirković, I., et al. (2007). Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 115, 891–899. doi: 10.1111/j.1600-0463.2007.apm_630.x
Tchonkouang, R. D., Onyeaka, H., and Nkoutchou, H. (2024). Assessing the vulnerability of food supply chains to climate change-induced disruptions. Sci. Total Environ. 920, 171047. doi: 10.1016/j.scitotenv.2024.171047
Timmusk, S., Abd El-Daim, I., Copolovici, L., Tanilas, T., Kännaste, A., Behers, L., et al. (2014). Droughttolerance of wheat improved by rhizosphere bacteria from harsh environments: enhanced biomass production and reduced emissions of stress volatiles. PloS One 9, e96086. doi: 10.1371/journal.pone.0096086
Turner, S., Pryer, K. M., Miao, V. P., and Palmer, J. D. (1999). Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rrna sequence analysis 1. J. Eukaryotic Microbiol. 46, 327–338. doi: 10.1111/j.1550-7408.1999.tb04612.x
Vasconcelos, J. C. S., Arantes, C. S., Speranza, E. A., Antunes, J. F. G., Barbosa, L. A. F., and Cançado, G.M. d. A. (2025). Predicting sugarcane yield through temporal analysis of satellite imagery during the growth phase. Agronomy 15, 793. doi: 10.3390/agronomy15040793
Vasconcelos, J. C. S., Speranza, E. A., Antunes, J. F. G., Barbosa, L. A. F., Christofoletti, D., Severino, F. J., et al. (2023). Development and validation of a model based on vegetation indices for the prediction of sugarcane yield. AgriEngineering 5, 698–719. doi: 10.3390/agriengineering5020044
Vlamakis, H., Chai, Y., Beauregard, P., Losick, R., and Kolter, R. (2013). Sticking together: building a biofilm the bacillus subtilis way. Nat. Rev. Microbiol. 11, 157–168. doi: 10.1038/nrmicro2960
Vu, B., Chen, M., Crawford, R. J., and Ivanova, E. P. (2009). Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 14, 2535–2554. doi: 10.3390/molecules14072535
Vurukonda, S. S. K. P., Vardharajula, S., Shrivastava, M., and Ali, S. (2016). Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 184, 13–24. doi: 10.1016/j.micres.2015.12.003
Zhang, M., Liao, B.-q., Zhou, X., He, Y., Hong, H., Lin, H., et al. (2015). Effects of hydrophilicity/hydrophobicity of membrane on membrane fouling in a submerged membrane bioreactor. Bioresour. Technol. 175, 59–67. doi: 10.1016/j.biortech.2014.10.058
Keywords: Glycine max (L) merrill., bioinoculants, drought tolerance, statistical modeling, vegetation index, spectral analysis
Citation: Vasconcelos JCS, Arantes CS, Gomes EA, Oliveira-Paiva CAd, de Sousa SM, Speranza EA, Antunes JFG, Lana UGdP and Cançado GMdA (2025) Bacillus-based inoculants enhance drought resilience in soybean: agronomic performance and remote sensing analysis from multi-location trials in Brazil. Front. Plant Sci. 16:1630127. doi: 10.3389/fpls.2025.1630127
Received: 16 May 2025; Accepted: 27 June 2025;
Published: 23 July 2025.
Edited by:
Krzysztof Szczyglowski, Agriculture and Agri-Food Canada (AAFC), CanadaReviewed by:
Mariela I Monteoliva, National Institute of Agricultural Technology (INTA), ArgentinaMalinda S. Thilakarathna, University of Alberta, Canada
Copyright © 2025 Vasconcelos, Arantes, Gomes, Oliveira-Paiva, de Sousa, Speranza, Antunes, Lana and Cançado. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eliane Aparecida Gomes, ZWxpYW5lLmEuZ29tZXNAZW1icmFwYS5icg==; Geraldo Magela de Almeida Cançado, Z2VyYWxkby5jYW5jYWRvQGVtYnJhcGEuYnI=