 Yue Xin
Yue Xin Qiuxian Huang
Qiuxian Huang Hongwei Yu
Hongwei Yu- College of Forestry, Hebei Agricultural University, Baoding, China
Most steppes are experiencing postgrazing succession, coupled with rainfall change and nitrogen (N) deposition. Despite the importance of soil resources in shaping community multifunctionality, little is known about how rainfall increase and N deposition influence steppe composition and production during postgrazing succession. We performed a field factorial experiment, subject to rainfall and N, each with three levels, to understand how simulated rainfall increase and N deposition affect the composition and production of a Leymus chinensis steppe. At the low rainfall increase, the dominance of L. chinensis increased with increasing N, while aboveground community production remained unchanged along the N gradient. At the high rainfall increase, aboveground community production was enhanced due to low N addition, but this facilitation disappeared in the presence of high N addition, and L. chinensis dominance was no longer affected by N. N-induced soil eutrophication but not soil acidification and soil microbes strongly affected steppe composition and production. Our findings suggest that high rainfall increase might weaken the potential of N addition to contribute to steppe composition and production, and also highlight the necessity of investigating interactions among multiple global change drivers.
1 Introduction
Plant communities are fundamentally characterized by their composition and production (Gurevitch et al., 2006), which rely tightly on soil resources (Odum and Barrett, 2005; Blair et al., 2014; Fay et al., 2015). Large amounts of soil resources are required to sustain plant communities (Aerts and Chapin, 2000; Schimel and Bennett, 2004; Odum and Barrett, 2005; Swemmer et al., 2007); on the other hand, available soil resources are commonly scarce (Schimel and Bennett, 2004; Elser et al., 2007; Blair et al., 2014; Fay et al., 2015). Thus, it is a basic challenge for plant communities to acquire sufficient soil resources.
The Earth’s ecosystems are experiencing precipitation change and atmospheric nitrogen (N) deposition (Galloway et al., 2004; IPCC, 2023; Zhang et al., 2024). These changes can strongly alter the availability of soil water and N, and thus influence plant community characteristics. For example, increased rainfall can alter plant community composition (Yang et al., 2011; Prevéy and Seastedt, 2014) and enhance community production (Harpole et al., 2007; DeMalach et al., 2017), and decreased soil water can decrease grassland diversity and production (Collins et al., 2014). Increased soil N can impact species richness and community functions (Bobbink et al., 2010; Yang et al., 2011). Water and N addition can yield synergistic effects on plant communities (Harpole et al., 2007; DeMalach et al., 2017), and also generate independent or even antagonistic effects (Zhang et al., 2014; Shen et al., 2022; Peng et al., 2024). Rainfall and N addition also have significant legacy effects (Yu and He, 2017).
Globally, steppes account for a high proportion of grasslands, occupying 30-40% of land surface and covering more terrestrial area than any other single biome type (Blair et al., 2014). Meanwhile, steppes are currently among the most threatened ecosystems due to overgrazing (Blair et al., 2014; Cao et al., 2024). Because of their ecological importance (Blair et al., 2014; Cao et al., 2024), most steppes are now experiencing postgrazing restoration through ceasing grazing (Yu and He, 2019). However, our current knowledge remains quite limited about how enhanced rainfall and N deposition influence postgrazing succession (Yu and He, 2019).
Our previous work focused on the impacts of rainfall and N addition on species turnover (i.e., species colonization and extinction) during postgrazing steppe succession (Yu and He, 2019). We found that the increases in rainfall and N enhanced species losses and gains at the same time and that the rates of species losses and gains were equal. Here we reported how rainfall and N addition influenced steppe composition and production during postgrazing succession. We proposed the following hypotheses: (1) rainfall increase can amplify the potential of N addition to impact steppe composition and production; (2) if so, this effect might be greater at high rainfall increases than at low rainfall increases. To test these hypotheses, we conducted an experiment with a steppe, which was subject to nine different combinations consisting of rainfall and N. We analyzed experimental data using principal component analysis, linear mixed-effects models, and structural equation modeling simultaneously, providing possible mechanisms explaining how rainfall and N addition alter steppe characteristics via soil acidification, soil eutrophication and soil microbes because previous studies have overlooked the relevant processes determining the effects of rainfall and N.
2 Materials and methods
2.1 Experimental site and designs
This experiment was carried out in a semiarid steppe (49.852° N, 120.348° E, 615 m a.s.l.) in Inner Mongolia, China, which had been severely degraded because of chronic overgrazing. The soil in the study area is categorized as sandy loam. Leymus chinenesis (C3 plant) and Cleistogenes squarrosa (C4 plant) were the dominant plant species in the study site before experiment manipulation. See our paper (Yu and He, 2019) for details about study site and plant species at the beginning of the experiment. During the experimental period (2009-2013), the growing season rainfall (327 mm) was greater than the long-term counterpart (294 mm, 1981-2010).
We conducted a field factorial experiment with rainfall and nitrogen treatments designed according to projections by Galloway et al. (2004) and IPCC (2023). Each factor had three levels, yielding a total of nine treatment combinations. Three N levels were: no N addition (N0); an addition of 5 g N m-2 yr-1 (N5); an addition of 10 g N m-2 yr-1 (N10). Three rainfall levels were: ambient rainfall (R0); a 14% increase in rainfall amount (R14); a 28% increase in rainfall amount (R28). The full-factorial experimental design was replicated in eight blocks at the study site. Within each block, we constructed nine 2 m × 2 m plots, with 3 m wide aisles separating adjacent plots. These plots within each block were then randomly assigned to the nine different rainfall and N treatment combinations. To achieve a 14% or 28% increase in precipitation amount, one or two rainfall collectors were installed adjacent to the designated plots to simulate changes in precipitation. Each collector was made of iron panels with a basal area of 0.56 m², mounted on a 0.3 m tall timber frame, and connected to the plots via drainage pipes. These pipes were evenly distributed within each plot to ensure uniform water delivery. In parallel, nitrogen was added in the form of NH4NO3, in dry form, at the beginning of each growing season (i.e., early June). The precipitation and nitrogen addition treatments were continuously implemented from 2009 to 2013. See our paper (Yu and He, 2017, 2019) for detailed information regarding the experimental plots and treatments of rainfall and N.
2.2 Measurements of plant community and soil properties
In August 2013, a 1.0 × 1.0 m quadrat was positioned at the center of each plot, and community composition was quantified by recording plant species in the 100 0.1 × 0.1 m squares making up a quadrat. We categorized all plant species into two different functional groups (i.e., C3 versus C4 plants), and calculated the ratio of C3 plant biomass to C4 plant biomass to indicate community composition. The aboveground biomass was harvested near the soil surface by clipping within two randomly placed 25 × 25 cm frames in each plot. Meanwhile, we sampled roots (including rhizomes) by taking 25 cm long × 25 cm wide × 15 cm deep soil cores from each clipped area and washed them by hand. The harvested shoots and roots were dried at 75°C for 48 h and weighed.
During plant harvest, soil pH was measured in situ using a portable pH meter, and soil samples were randomly collected at a depth of 0-10 cm from each experimental plot. After sieving through a 2 mm mesh to remove debris, each soil sample was divided into three subsamples. The first subsample was stored at 4 °C for the determination of available phosphorus, and ammonium nitrogen (NH4+-N) and nitrate nitrogen (NO3-–N); the second subsample was air-dried for the measurement of total carbon, total nitrogen and total phosphorus; the third subsample was frozen at -80 °C for soil microbial community analysis. Phospholipid fatty acid (PLFA) profiling was used to characterize microbial communities, which were classified into bacteria, actinomycetes, fungi, arbuscular mycorrhizal fungi, and protozoa. The PLFA biomarkers used for microbial group identification are listed in Supplementary Table S1. Moreover, the fungi/bacteria ratio was calculated to assess changes in microbial community structure.
2.3 Data analyses
We measured one soil acidification variable (i.e., soil pH), six soil nutrient variables and six soil microbial variables in this study. To reduce the dimensionality and collinearity of soil characteristics, principal component analyses (PCA) were conducted on the variables related to soil nutrients (PCAN) and soil microbes (PCAM) separately. The scores of first and second axes of resulting PCA results were used as representative variables and used for the following analyzes.
We first performed linear mixed-effects models to test the effects of N addition, increased rainfall, and their interactions on soil and community characteristics, with block treated as a random term. For variables with significant main or interaction effects, pairwise comparisons were conducted to examine differences among treatment levels or combinations. Then, spearman correlations were performed to explore the associations between plant community characteristics and soil representative variables. In order to improve normality, the ratio of C3 plants to C4 plants and aboveground biomass were log-transformed when necessary. To figure out how N addition and increased rainfall influenced steppe composition and production, we performed structural equation modeling (SEM) to evaluate the relative importance of soil acidification, soil nutrients and soil microbes induced by N addition and increased rainfall on the ratio of C3 plants to C4 plant species, and biomass. We assume that N addition and increased rainfall influence steppe composition and production via affecting soil acidification, soil nutrients and soil microbes. The SEM analyzes were performed with the maximum likelihood estimation method. All analyzes were performed with vegan (Oksanen et al., 2022), lme4 (Bates et al., 2015), car (Fox and Weisberg, 2019), emmeans (Lenth, 2023), lavaan (Rosseel, 2012) and ggcor (Huang et al., 2020) packages using R4.3.1 (R Core Team, 2023).
3 Results
3.1 Effects of rainfall and N addition on steppe performance
Leymus chinensis accounted for 77% of C3 plant biomass and C. squarrosa accounted for 99% of C4 plant biomass. The linear mixed-effects models results showed that increased rainfall did not alter the species richness and the ratio of C3 plant species to C4 plant species (Table 1; Figures 1A, B); N addition increased the ratio of C3 plant species to C4 plant species, but this N-added effect disappeared at higher rainfall increases (Figure 1B). The aboveground biomass was enhanced by N addition rather than increased rainfall; importantly, this facilitation of N addition was substantially offset by higher rainfall increases (Table 1; Figure 1C). The belowground biomass was unaffected by increased rainfall and N addition, but significantly influenced by their interaction (Table 1; Figure 1D).
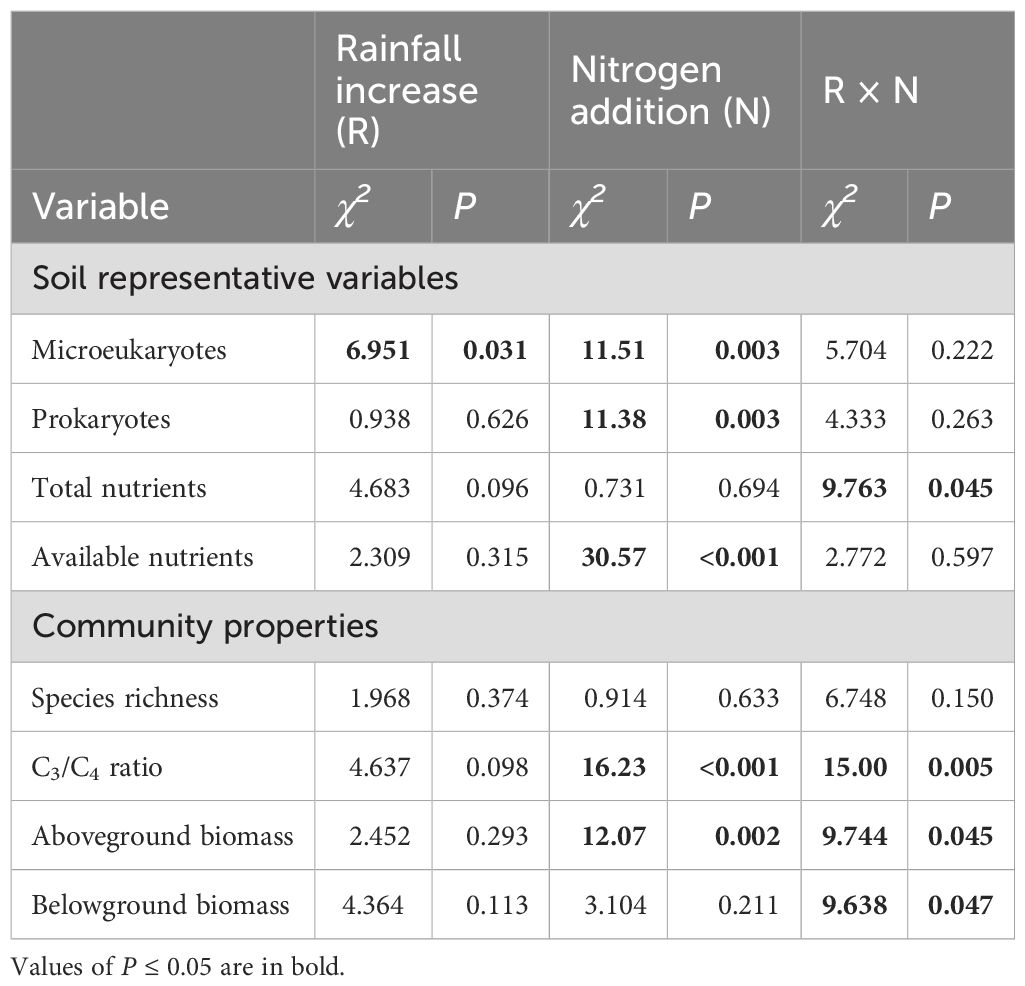
Table 1. The effects of rainfall increase, N addition, and their interactions on soil representative variables and community properties.
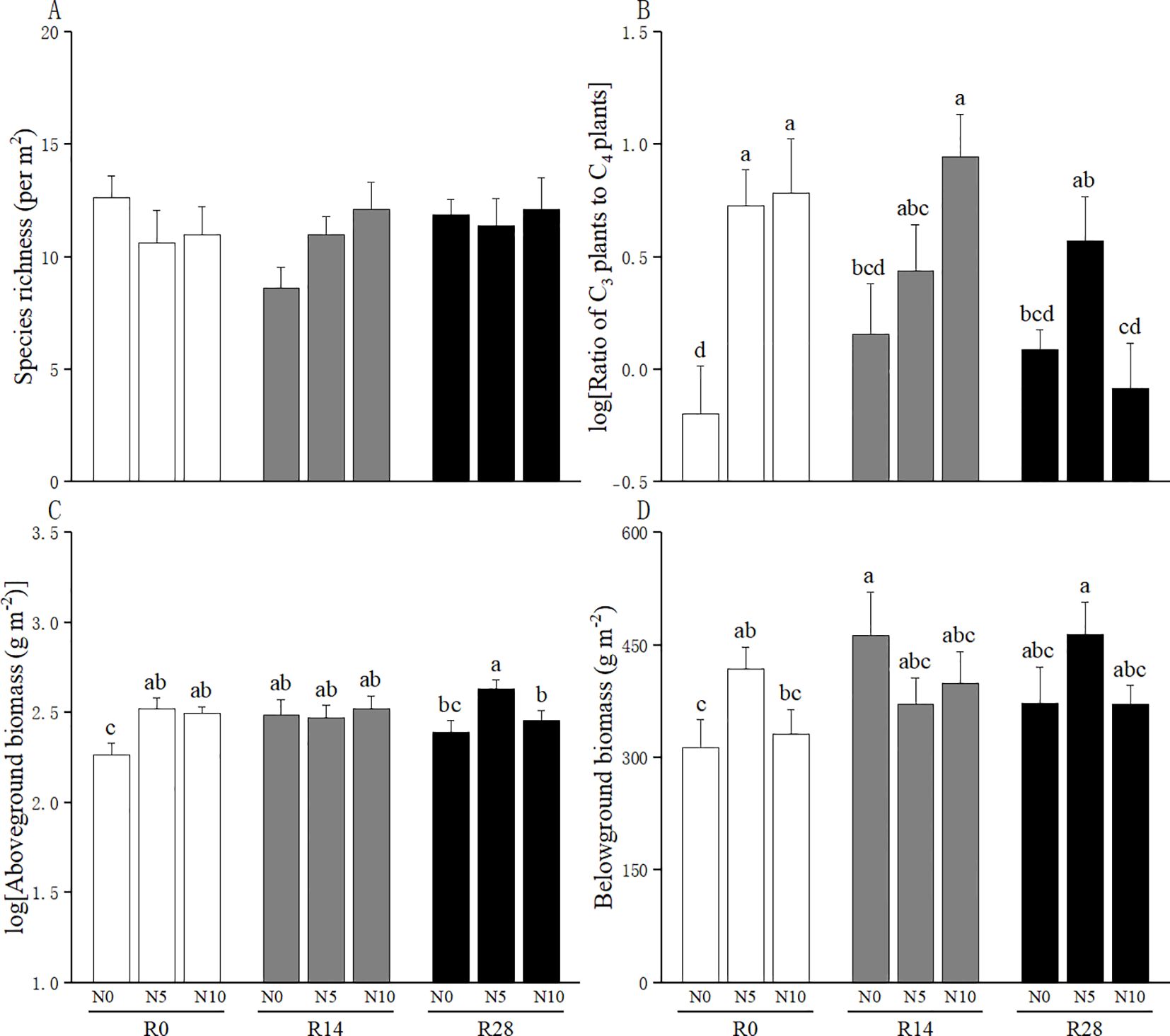
Figure 1. Effects of increased rainfall and N addition on (A) species richness, (B) the ratio of C3 plants to C4 plants, (C) aboveground biomass, and (D) belowground biomass of plant communities. The data are expressed as means + 1 SE. Different lowercase letters indicate significant difference among treatment combinations. N0: no N addition; N5: an addition of 5 g N m-2 yr-1; N10: an addition of 10 g N m-2 yr-1. R0: no rainfall addition; R14: a 14% increase in rainfall amount over the ambient rainfall; R28: a 28% increase in rainfall amount over the ambient rainfall.
3.2 Effects of rainfall and N addition on soil properties
Increased rainfall did not alter soil properties except for soil fungi and arbuscular mycorrhizal fungi (Supplementary Table S2; Supplementary Figure S1). Soil fungi and arbuscular mycorrhizal fungi were more abundant at high rainfall increases than at low rainfall increases. Unlike rainfall enrichment, N addition influenced some soil properties (Supplementary Table S2; Supplementary Figure S2). For example, soil fungi decreased along the N gradient, N addition increased NH4+-N and NO3-–N, and N addition also decreased soil pH (soil acidification) (Supplementary Figure S2, S3). Additionally, increased rainfall and N addition interacted to influence soil total carbon (Supplementary Table S2).
According to the PCAN analysis, the first and second axes explained 36.29% and 28.42% of the variation in soil nutrient, respectively (Figure 2A). The first axis was positively correlated with total carbon, total nitrogen, and total phosphorus and thus was categorized as composite measurements of “total nutrients”. The second axis was categorized as measurements of “available nutrients” because it had a high relevance with available nitrogen and phosphorus. A greater value on the first and second axis indicated a high degree of soil eutrophication. In the PCAM analysis, the first two axes explained 60.95% and 24.74% of the variation in soil microbes, respectively (Figure 2B). The first axis was categorized as measurements of “microeukaryotes” as it most relevant to fungi, arbuscular mycorrhizal fungi and protozoa. The second axis reflected bacteria, actinomycetes and the trade-off between fungi and bacteria, thus was designated as measurements of “prokaryotes”. In these cases, a greater value on the first and second axes corresponds to a higher microbial biomass in microeukaryotes (first axis) and prokaryotes (second axis). The linear mixed-effects models results showed that increased rainfall significantly affected soil microeukaryotes, while it had no effect on other soil representative variables (Table 1; Figure 3). Adding nitrogen, regardless of N5 or N10, significantly enhanced soil prokaryotes compared to N0 (Table 1; Figure 3B). Furthermore, both soil available nutrients and microeukaryotes significantly increased with the increasing nitrogen application rate (Table 1; Figures 3A, D). In addition, increased rainfall and nitrogen addition interactively affected soil total nutrients (Table 1; Figure 3C).
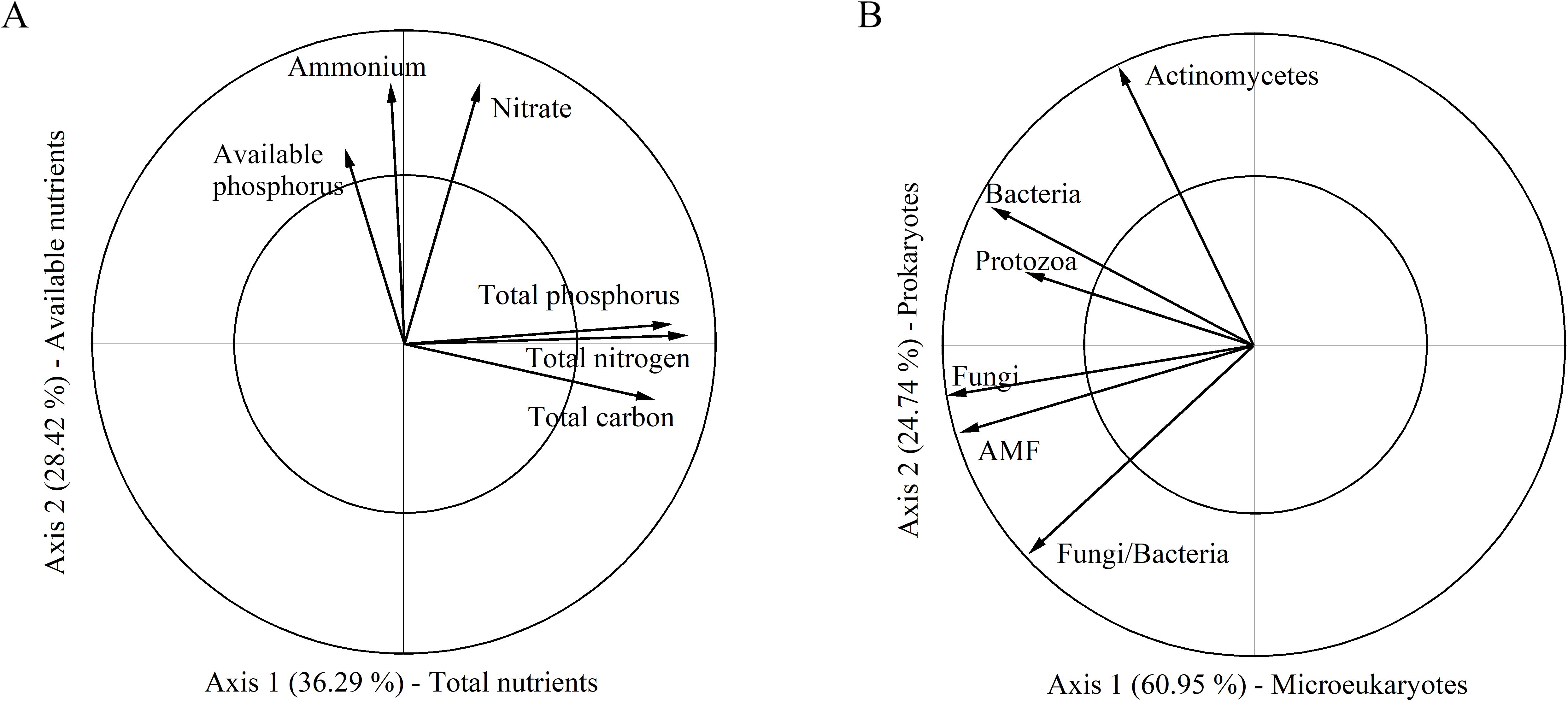
Figure 2. Principal component analysis (PCA) for variables on soil nutrients (PCAN) and soil microbes (PCAM). (A) The first and second axes in the PCAN were categorized as composite measurements of “Total nutrients” and “Available nutrients”, respectively. (B) The first and second axes in the PCAM were categorized as composite measurements of “Microeukaryotes” and “Prokaryotes”, respectively.
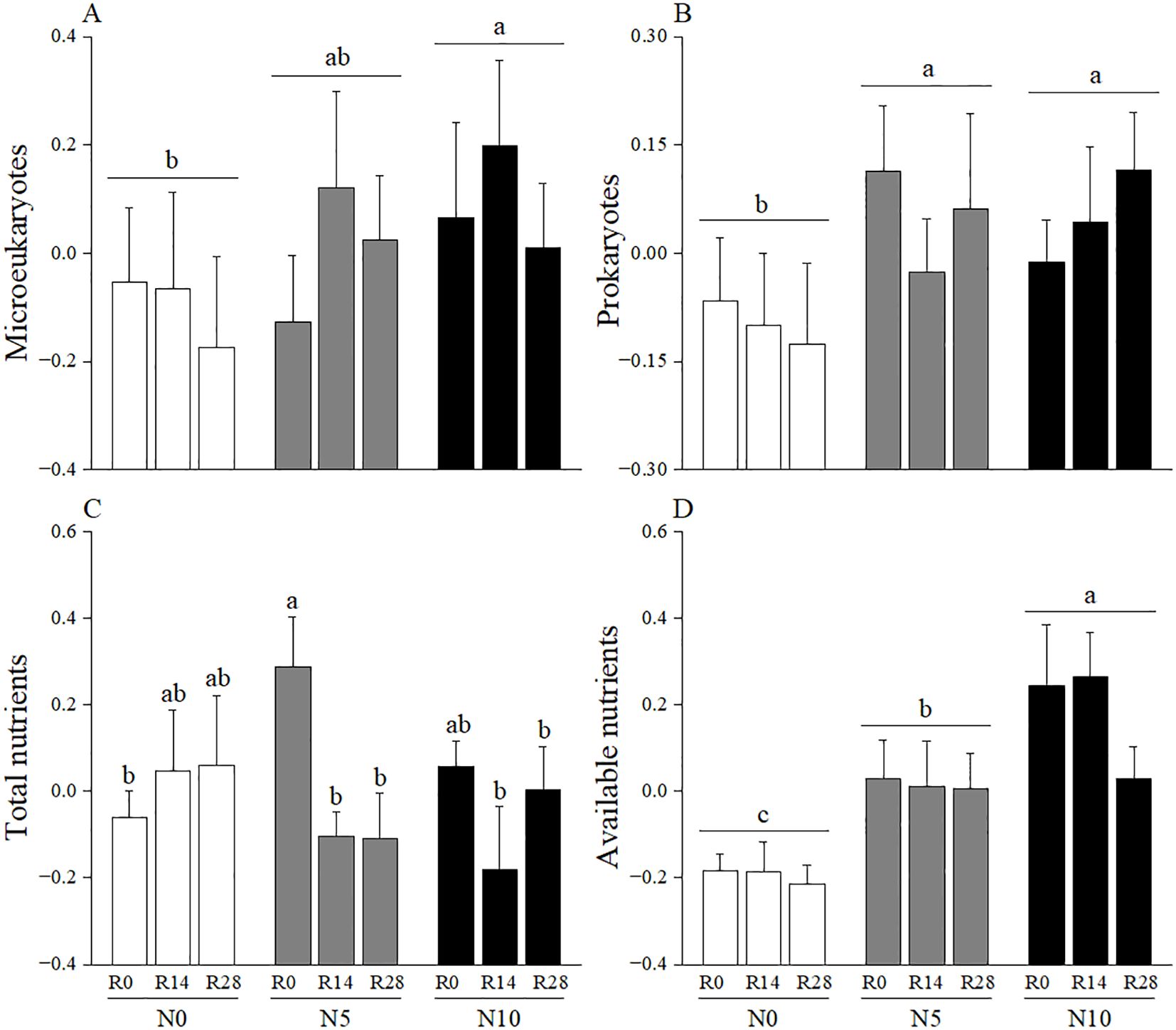
Figure 3. Effects of increased rainfall and N addition on soil representative PCA variables of (A) Microeukaryotes, (B) Prokaryotes, (C) Total nutrients and (D) Available nutrients. The data are expressed as means + 1 SE. Treatment abbreviations are from Figure 1. Different lowercase letters indicate significant difference among N treatment levels or combinations.
3.3 Correlations between steppe performance and soil representative variables
Species composition and community biomass showed different associations with soil representative variables (Figure 4). Soil acidification, as indicated by soil pH, was negatively and significantly correlated with species richness, but no significant associations were detected with other steppe performance indicators (Figure 4). Among soil microbes, soil with higher prokaryotes rather than microeukaryotes tended to enhance the ratio of C3 plant species to C4 plant species and community biomass but decreased species richness (Figure 4). Higher soil eutrophication (i.e., total and available nutrients) exhibited significantly positive correlations with community biomass and the ratio of C3 plant species to C4 plant species but showed no linkage with species richness (Figure 4).
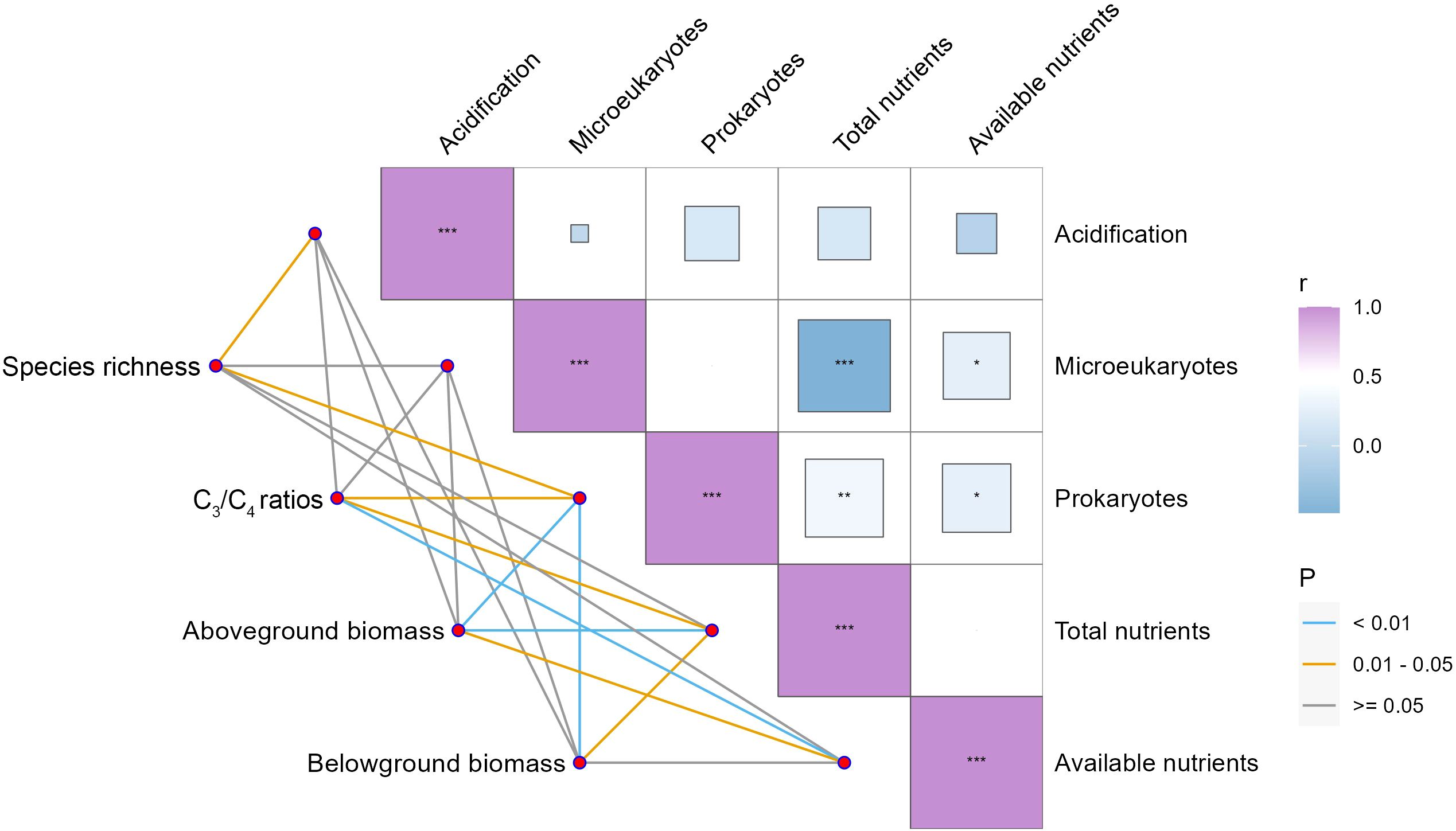
Figure 4. Spearman correlations between steppe performance and soil representative variables. *P < 0.05, **P < 0.01, ***P < 0.001.
3.4 Pathways linking field manipulations and steppe performance
Increased rainfall had no significant effects on steppe composition via affecting soil acidification, eutrophication, and microbes (Figure 5). Nitrogen addition increased the abundance of C3 species indirectly by increasing soil available nutrient availability (Figure 5). Although N addition decreased soil pH (i.e., inducing soil acidification), these alterations had no effects on steppe composition (Figure 5). Additionally, soil total nutrients positively influenced the ratio of C3 plant species to C4 plant species (Figure 5).
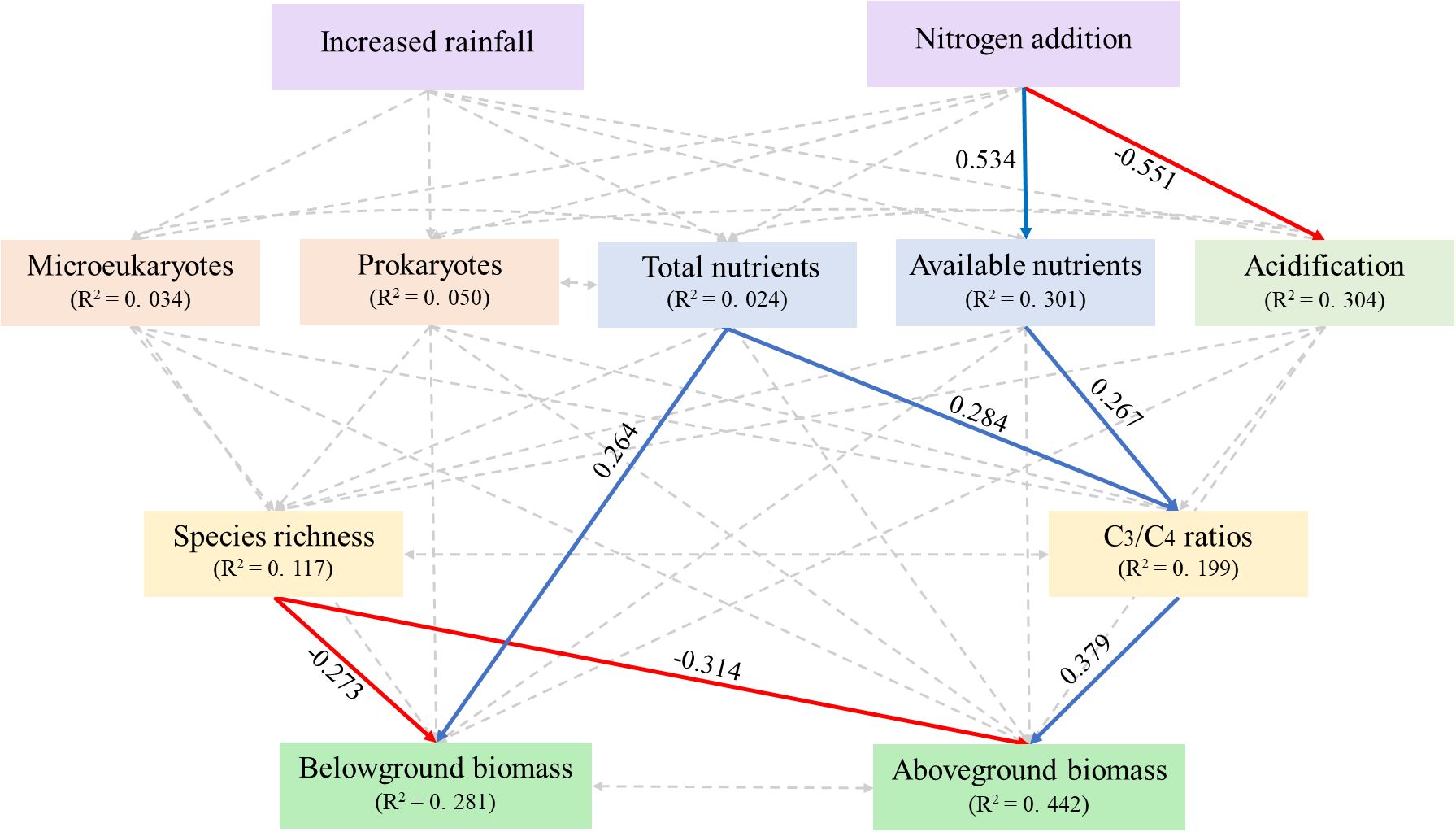
Figure 5. Structural equation modeling examining the effects of increased rainfall and nitrogen addition on plant community composition and production via influencing soil representative variables. Solid red and blue indicate significantly negative and positive effects (P < 0.05); the non-significant pathways are shown in grey dot lines (P > 0.05). Numbers associated with pathways between variables represent standardized path coefficients. Line width is proportional to the strength of the relationship. χ2 = 20.483, P = 0.084.
While the C3/C4 ratios exhibited a significantly positive relationship with aboveground biomass, species richness was negatively and significantly contributed to both aboveground and belowground biomass (Figure 5). The effects of increased rainfall on plant biomass were similar to those on species composition, as no significant pathways were observed in the SEM analysis (Figure 5). Nitrogen exhibited a positive effect on aboveground biomass by influencing available nutrients and the C3/C4 ratios; however, no significant pathways were observed on belowground biomass by influencing soil representative variables and species composition (Figure 5). In addition, soil total nutrients were positively correlated with belowground biomass (Figure 5).
4 Discussion
This work highlighted the influences of simulated rainfall increase and N deposition on steppe composition and production during postgrazing succession. In our study region, the climax plant community is L. chinensis communities and C. squarrosa is a subordinate species in natural vegetation (IGIMVCAS, 1985). When L. chinensis communities degraded due to overgrazing, L. chinensis shifted from a dominant species to a subordinate species whereas C. squarrosa shifted from a subordinate species to a dominant species (Yu and He, 2019). At the scale of our experimental plots, L. chinensis and C. squarrosa accounted for majority of C3 plant biomass (77%) and C4 plant biomass (99%), respectively. Thus, the change in C3/C4 ratios primarily indicated the shift between L. chinensis and C. squarrosa. Here, projected rainfall increases had no significant effects on the composition and primary production of a steppe; a N5 or N10 increased C3/C4 ratios and aboveground primary production (Figure 1). These findings suggest that precipitation increase might not strongly influence steppes but simulated N deposition could improve steppes through shifting local steppes from C. squarrosa communities to L. chinensis communities and increasing their production. As an important global change driver, N deposition has profound impacts on plant community composition, productivity, and biogeochemical cycling of grasslands (Kimmel et al., 2020; Ke et al., 2023; Yuan et al., 2023). However, its ecological efficacy may depend not only on the amount of nitrogen added, but also on the timing of its application (Zhang et al., 2021a; Song et al., 2023). In our study, nitrogen was applied at the beginning of the growing season, aligning with the period of peak plant nutrient demand. This synchrony likely enhanced nitrogen uptake by dominant species such as L. chinensis, while reducing potential losses via microbial immobilization. Such timing may partly explain why nitrogen addition effectively promoted C3 dominance and aboveground productivity during postgrazing succession.
A key finding of our study was that different strengths of rainfall increases had contrasting impacts on the effects of N addition on steppes. More specifically, at the low rainfall increase, C3/C4 plant ratio gradually increased with N levels; in contrast, at the high rainfall increase, low-level N addition substantially increased biomass production compared with the controls, but this facilitation disappeared when high-level N was supplied (Figure 1). Thus, our findings suggest that high rainfall increase might weaken the potential of high-level N to improve steppes, although they are inconsistent with previous findings (Yang et al., 2011; Ma et al., 2020; Ren et al., 2021).
We propose several possibilities that might explain the above-mentioned findings. Intermediate rainfall events are responsible for most of the rainfall effects for steppes (Sala and Lauenroth, 1982; Lauenroth and Sala, 1992). In our study region, precipitation mainly falls in the form of heavy rainfall events during growing seasons. Large rainfall exceeds the field capacity of soil and causes soil erosion and leaching (Kong et al., 2013; Brown and Collins, 2024), thereby decreasing retention of soil nutrients and influencing N availability in the plant root zone. Second, water pulses can result in a higher N mineralization rate and N mobility (Wang et al., 2006; Lü and Han, 2010; Peng et al., 2024), and N addition can decrease soil water availability by increasing plant photosynthetic rate and transpiration (Harpole et al., 2007; Yang et al., 2011). Third, there are counteractive effects among different ecological processes, which could alter interspecific relationships. Although we did not directly measure the availability of water in the soil in this study, it is important to acknowledge its potential role in mediating the effects of rainfall and nitrogen on steppe ecosystems. Our findings might provide insights into the importance of rainfall increase and N deposition in regulating steppes. For example, increased rainfall may play a weak role in improving steppes, particularly in those areas where heavy rainfall events are frequent in growing seasons. Furthermore, increasing rainfall can offset the positive effects of N on steppes.
We also observed that the importance of soil acidification and eutrophication, which were induced by N addition, differed dramatically. We found that soil eutrophication but not soil acidification strongly affected steppe composition and production (Figure 5). These findings disagree with the global synthesis by Bobbink et al. (2010). Interestingly, N-induced soil acidification did not decrease plant species richness (Figure 4). Species loss is commonly viewed to be linked to soil acidification, soil eutrophication, and competitive exclusion (Stevens et al., 2010; Kimmel et al., 2020; Ke et al., 2023). However, these processes can also create new niches for new plant species (Bobbink et al., 2010; Yang et al., 2011; Cai et al., 2023). Thus, soil acidification and eutrophication may not necessarily result in plant species loss. An increase in the abundance of C3 plant species might be related to their high potential to acquire and utilize essential resources (Zhang et al., 2021b), functional types (Harpole et al., 2007; Reich et al., 2001; Cai et al., 2023), and increased ammonium and ammonia availability (Bobbink et al., 2010).
Additionally, steppes had asymmetric sensitivity to experimental manipulations. At the community level, plant species richness was insensitive to N (Yu and He, 2019) whereas steppe composition was sensitive to N. These results are contrary to previous studies (Stevens et al., 2010; Harpole et al., 2016). At the ecosystem level, aboveground biomass was more sensitive to water and N manipulations than belowground biomass. In nature, belowground biomass is determined by root production and turnover (Majdi and Andersson, 2005; Leppälammi-Kujansuu et al., 2014; Keller et al., 2023). N addition enhanced aboveground production, which could be attributed to faster growth in favorable conditions (Ren et al., 2015; Yu and He, 2017; Peng et al., 2024) and the fact that the positive effects of N on aboveground production were stronger than the negative effects of plant species richness.
In summary, the potential of simulated N deposition to improve grasslands was not amplified by increasing rainfall. As opposed to our currently thought, high rainfall increase might weaken the efficacy of high N deposition on steppes. The complex interplay between rainfall changes and N deposition plays a vital role in determining steppe trajectories during postgrazing succession. Recognizing such interactions among multiple global change drivers is essential for predicting and managing grassland ecosystems.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
YX: Methodology, Funding acquisition, Formal analysis, Writing – original draft, Writing – review & editing. QH: Writing – original draft, Visualization, Writing – review & editing. BL: Writing – original draft, Formal analysis, Visualization, Writing – review & editing. ZL: Formal analysis, Writing – original draft, Visualization. YS: Visualization, Writing – original draft, Writing – review & editing. MZ: Writing – original draft, Funding acquisition. HY: Writing – original draft, Writing – review & editing, Funding acquisition, Investigation, Formal analysis, Methodology.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the Natural Science Foundation of Hebei Province (Grant Number: C2024204082, C2022204058), the Central Guided for Local Science and Technology (Grant Number: 246Z7609G), the Hebei Agricultural University Talents Fund (Grant Number: YJ2020052, YJ2024003), and the Hebei Province Higher School Basic Research Business Fee Project (Grant Number: KY2023036, KY2024029).
Acknowledgments
We are grateful to Dr. Wei-Ming He for his help.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1635593/full#supplementary-material
References
Aerts, R. and Chapin, F. S. (2000). The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 30, 1–67. doi: 10.1016/S0065-2504(08)60016-1
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Software 67, 1–48. doi: 10.18637/jss.v067.i01
Blair, J., Nippert, J., and Briggs, J. (2014). “Grassland ecology,” in Ecology and the environment, the plant science, vol. 8 . Ed. Monson, R. K. (Springer, New York), 389–423.
Bobbink, R., Hicks, K., Galloway, J., Spranger, T., Alkemade, R., Ashmore, M., et al. (2010). Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol. Appl. 20, 30–59. doi: 10.1890/08-1140.1, PMID: 20349829
Brown, R. F. and Collins, S. L. (2024). Revisiting the bucket model: long-term effects of rainfall variability and nitrogen enrichment on net primary production in a desert grassland. J. Ecol. 112, 629–641. doi: 10.1111/1365-2745.14258
Cai, J., Weiner, J., Luo, W., Feng, X., Yang, G., Lu, J., et al. (2023). Functional structure mediates the responses of productivity to addition of three nitrogen compounds in a meadow steppe. Oecologia 201, 575–584. doi: 10.1007/s00442-022-05310-9, PMID: 36688977
Cao, F., Li, W., Jiang, Y., Gan, X., Zhao, C., and Ma, J. (2024). Effects of grazing on grassland biomass and biodiversity: A global synthesis. Field Crop Res. 306, 109204. doi: 10.1016/j.fcr.2023.109204
Collins, S. L., Belnap, J., Grimm, N. B., Rudgers, J. A., Dahm, C. N., D’Odorico, P., et al. (2014). A multiscale, hierarchical model of pulse dynamics in arid-land ecosystems. Annu. Rev. Ecol. Evol. S. 45, 397–419. doi: 10.1146/annurev-ecolsys-120213-091650
DeMalach, N., Zaady, E., and Kadmon, R. (2017). Contrasting effects of water and nutrient additions on grassland communities: a global meta-analysis. Global Ecol. Biogeogr. 26, 983–992. doi: 10.1111/geb.12603
Elser, J. J., Bracken, M. E. S., Cleland, E. E., Gruner, D. S., Harpole, W. S., Hillebrand, H., et al. (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135–1142. doi: 10.1111/j.1461-0248.2007.01113.x, PMID: 17922835
Fay, P. A., Prober, S. M., Harpole, W. S., Knops, J. M. H., Bakker, J. D., Borer, E. T., et al. (2015). Grassland productivity limited by multiple nutrients. Nat. Plants 1, 15080. doi: 10.1038/nplants.2015.80, PMID: 27250253
Fox, J. and Weisberg, S. (2019). An R companion to applied regression (Thousand Oaks CA, Sage Publications).
Galloway, J. N., Dentener, F. J., Capone, D. G., Boyer, E. W., Howarth, R. W., Seitzinger, S. P., et al. (2004). Nitrogen cycles: past, present, and future. Biogeochemistry 70, 153–226. doi: 10.1007/s10533-004-0370-0
Gurevitch, J., Scheiner, S. M., and Fox, G. A. (2006). The ecology of plants. 2nd (Sunderland, MA, Sinauer Associates Inc).
Harpole, W. S., Potts, D. L., and Suding, K. N. (2007). Ecosystem responses to water and nitrogen amendment in a California grassland. Global Change Biol. 13, 2341–2348. doi: 10.1111/j.1365-2486.2007.01447.x
Harpole, W. S., Sullivan, L. L., Lind, E. M., Firn, J., Adler, P. B., Borer, E. T., et al. (2016). Addition of multiple limiting resources reduces grassland diversity. Nature 537, 93–96. doi: 10.1038/nature19324, PMID: 27556951
Huang, H., Zhou, L., Chen, J., and Wei, T. (2020). ggcor: Extended tools for correlation analysis and visualization (R package version 0.9.7). Available online at: https://github.com/houyunhuang/ggcor (accessed February 1, 2025)
IGIMVCAS (Investigation Group of Inner Mongolia Vegetation, Chinese Academy of Sciences) (1985). Inner Mongolia vegetation (Beijing: Science Press).
IPCC (2023). “Climate change 2023: synthesis report,” in Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Eds. Core Writing Team, Lee, H., and Romero, J. (IPCC, Geneva, Switzerland), 35–135.
Ke, Y., Yu, Q., Wang, H., Zhao, Y., Jia, X., Yang, Y., et al. (2023). The potential bias of nitrogen deposition effects on primary productivity and biodiversity. Glob. Change Biol. 29, 1054–1061. doi: 10.1111/gcb.16530, PMID: 36408718
Keller, A. B., Walter, C. A., Blumenthal, D. M., Borer, E. T., Collins, S. L., DeLancey, L. C., et al. (2023). Stronger fertilization effects on aboveground versus belowground plant properties across nine US grasslands. Ecology 104, e3891. doi: 10.1002/ecy.3891, PMID: 36208208
Kimmel, K., Furey, G. N., Hobbie, S. E., Isbell, F., Tilman, D., and Reich, P. B. (2020). Diversity-dependent soil acidification under nitrogen enrichment constrains biomass productivity. Glob. Change Biol. 26, 6594–6603. doi: 10.1111/gcb.15329, PMID: 32871613
Kong, D. L., Lü, X. T., Jiang, L. L., Wu, H. F., Miao, Y., and Kardol, P. (2013). Extreme rainfall events can alter inter-annual biomass responses to water and N enrichment. Biogeosciences 10, 8129–8138. doi: 10.5194/bg-10-8129-2013
Lauenroth, W. K. and Sala, O. E. (1992). Long-term forage production of North-American shortgrass steppe. Ecol. Appl. 2, 397–403. doi: 10.2307/1941874, PMID: 27759270
Lenth, R. (2023). emmeans: Estimated marginal means, aka least-squares means (R package version 1.8.8). Available at: https://CRAN.R-project.org/package=emmeans (accessed February 1, 2025).
Leppälammi-Kujansuu, J., Salemaa, M., Kleja, D. B., Linder, S., and Helmisaari, H. S. (2014). Fine root turnover and litter production of Norway spruce in a long-term temperature and nutrient manipulation experiment. Plant Soil. 374, 73–88. doi: 10.1007/s11104-013-1853-3
Lü, X. T. and Han, X. G. (2010). Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia, China. Plant Soil. 327, 481–491. doi: 10.1007/s11104-009-0078-y
Ma, Q. H., Liu, X. D., Li, Y. B., Li, L., Yu, H. Y., Qi, M., et al. (2020). Nitrogen deposition magnifies the sensitivity of desert steppe plant communities to large changes in precipitation. J. Ecol. 108, 598–610. doi: 10.1111/1365-2745.13264
Majdi, H. and Andersson, P. (2005). Fine root production and turnover in a Norway spruce stand in northern Sweden: effects of nitrogen and water manipulation. Ecosystems 8, 191–199. doi: 10.1007/s10021-004-0246-0
Odum, E. P. and Barrett, G. W. (2005). Fundamentals of Ecology. 5th (Belmont, California, USA: Thomson Brooks/Cole).
Oksanen, J., Simpson, G., Blanchet, F., Kindt, R., Legendre, P., Minchin, P., et al. (2022). Vegan: Community Ecology Package (R Package Version 2.6-4). Available online at: https://CRAN.R-project.org/package=vegan (accessed February 1, 2025).
Peng, J. L., Zhang, R. Y., Ma, F. F., Quan, Q., Liao, J. Q., Zhou, Q. P., et al. (2024). Precipitation differentially regulates above- and belowground productivity in response to nitrogen enrichment in an alpine meadow. Agr. For. Meteorol. 358, 110257. doi: 10.1016/j.agrformet.2024.110257
Prevéy, J. S. and Seastedt, T. R. (2014). Seasonality of precipitation interacts with exotic species to alter composition and phenology of a semi-arid grassland. J. Ecol. 102, 1549–1561. doi: 10.1111/1365-2745.12320
R Core Team (2023). R: A Language and Environment for Statistical Computing (Vienna: R Foundation for Statistical Computing).
Reich, P. B., Tilman, D., Craine, J., Ellsworth, D., Tjoelker, M. G., Knops, J., et al. (2001). Do species and functional groups differ in acquisition and use of C, N and water under varying atmospheric CO2 and N availability regimes? A field test with 16 grassland species. New Phytol. 150, 435–448. doi: 10.1046/j.1469-8137.2001.00114.x
Ren, H. Y., Xu, Z. W., Huang, J. H., Lü, X. T., Zeng, D. H., Yuan, Z. Y., et al. (2015). Increased precipitation induces a positive plant-soil feedback in a semi-arid grassland. Plant Soil. 389, 211–223. doi: 10.1007/s11104-014-2349-5
Ren, Z. R., Zhang, Y. Q., and Zhang, Y. H. (2021). Nitrogen deposition magnifies the positive response of plant community production to precipitation: Ammonium to nitrate ratio matters. Environ. pollut. 276, 116659. doi: 10.1016/j.envpol.2021.116659, PMID: 33621734
Rosseel, Y. (2012). lavaan: an R package for structural equation modeling. J. Stat. Software 48, 1–36. doi: 10.18637/jss.v048.i02
Sala, O. E. and Lauenroth, W. K. (1982). Small rainfall events: an ecological role in semi-arid regions. Oecologia 53, 301–304. doi: 10.1007/BF00389004, PMID: 28311731
Schimel, J. P. and Bennett, J. (2004). Nitrogen mineralization: Challenges of a changing paradigm. Ecology 85, 591–602. doi: 10.1890/03-8002
Shen, R., Zhang, Y., Zhu, J., Chen, N., Chen, Y., Zhao, G., et al. (2022). The interactive effects of nitrogen addition and increased precipitation on gross ecosystem productivity in an alpine meadow. J. Plant Ecol. 15, 168–179. doi: 10.1093/jpe/rtab081
Song, C., Zhang, Y., Ren, Z., Lu, H., Chen, X., Liu, R., et al. (2023). Soil nitrate mediates the responses of plant community production to the frequency of N addition in a temperate grassland: a decadal field experiment. Plant Soil. 491, 9–20. doi: 10.1007/s11104-022-05435-z
Stevens, C. J., Thompson, K., Grime, J. P., Long, C. J., and Gowing, D. J. G. (2010). Contribution of acidification and eutrophication to declines in species richness of calcifuge grasslands along a gradient of atmospheric nitrogen deposition. Funct. Ecol. 24, 478–484. doi: 10.1111/j.1365-2435.2009.01663.x
Swemmer, A. M., Knapp, A. K., and Snyman, H. A. (2007). Intra-seasonal precipitation patterns and above-ground productivity in three perennial grasslands. J. Ecol. 95, 780–788. doi: 10.1111/j.1365-2745.2007.01237.x
Wang, C., Wan, S., Xing, X., Zhang, L., and Han, X. (2006). Temperature and soil moisture interactively affected soil net N mineralization in temperate grassland in northern China. Soil Biol. Biochem. 38, 1101–1110. doi: 10.1016/j.soilbio.2005.09.009
Yang, H., Li, Y., Wu, M., Zhang, Z., Li, L., and Wan, S. (2011). Plant community responses to nitrogen addition and increased precipitation: the importance of water availability and species traits. Global Change Biol. 17, 2936–2944. doi: 10.1111/j.1365-2486.2011.02423.x
Yu, H. W. and He, W. M. (2017). Negative legay effects of rainfall and nitrogen amendment on leaf lifespan of steppe species. J. Plant Ecol. 10, 831–838. doi: 10.1093/jpe/rtw090
Yu, H. W. and He, W. M. (2019). Increased rainfall and nitrogen alter colonization and extinction during postgrazing steppe succession. J. Veg. Sci. 30, 75–85. doi: 10.1111/jvs.12698
Yuan, X., Niu, D., Guo, D., and Fu, H. (2023). Responses of soil carbon and nitrogen mineralization to nitrogen addition in a semiarid grassland: The role of season. Catena 220, 106719. doi: 10.1016/j.catena.2022.106719
Zhang, Y., Ren, Z., and Zhang, Y. (2021a). Winter nitrogen enrichment does not alter the sensitivity of plant communities to precipitation in a semiarid grassland. Sci. Total Environ. 790, 148264. doi: 10.1016/j.scitotenv.2021.148264, PMID: 34380248
Zhang, R., Schellenberg, M. P., Tian, D., Ma, F., Zhang, T., Wang, H., et al. (2021b). Shifting community composition determines the biodiversity–productivity relationship under increasing precipitation and N deposition. J. Veg. Sci. 32, e12998. doi: 10.1111/jvs.12998
Zhang, X. M., Wei, H. W., Chen, Q. S., and Han, X. G. (2014). The counteractive effects of nitrogen addition and watering on soil bacterial communities in a steppe ecosystem. Soil Biol. Biochem. 72, 26–34. doi: 10.1016/j.soilbio.2014.01.034
Keywords: community composition and functions, nitrogen addition, offsetting effects, postgrazing steppe succession, rainfall intensity
Citation: Xin Y, Huang Q, Liu B, Li Z, Sun Y, Zhang M and Yu H (2025) Rainfall and nitrogen addition have no synergistic effects on steppe composition and production in postgrazing succession. Front. Plant Sci. 16:1635593. doi: 10.3389/fpls.2025.1635593
Received: 26 May 2025; Accepted: 29 July 2025;
Published: 05 September 2025.
Edited by:
Rogério de Souza Nóia Júnior, INRAE Occitanie Montpellier, FranceReviewed by:
Huailin Zhou, Chinese Academy of Meteorological Sciences, ChinaHongying Yu, Jiangxi Agricultural University, China
Copyright © 2025 Xin, Huang, Liu, Li, Sun, Zhang and Yu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hongwei Yu, aG9uZ3dlaXl1MTk5MEAxMjYuY29t