 Zirui Wang1
Zirui Wang1 Yuanshuo Hao
Yuanshuo Hao Lihu Dong
Lihu Dong Zheng Miao
Zheng Miao Xuehan Zhao
Xuehan Zhao- 1College of Forestry, Northeast Forestry University, Harbin, Heilongjiang, China
- 2Key Laboratory of Sustainable Forest Ecosystem Management-Ministry of Education, School of Forestry, Northeast Forestry University, Harbin, Heilongjiang, China
Introduction: The Da Xing’an Mountains region harbors China's only cold-temperate coniferous forests and serves as a critical ecological barrier, playing a vital role in forest ecosystems and carbon sequestration. Stand age, shaped by population dynamics, disturbance regimes, and management practices, significantly influences the global carbon cycle. Although forest development is known to correlate with productivity shifts, how production varies across specific stand developmental stages and the relative contributions of driving factors remain poorly understood.
Methods: Using data from the National Forest Continuous Inventory (NFCI, 2005–2010) in the eastern Da Xing’an Mountains, we analyzed the effects of stand characteristics, structural diversity, and environmental variables on Net Ecosystem Carbon Change (NECC) across a spectrum of developmental stages, from young to overaged forests.
Results: Our findings demonstrate that: (1) Net Ecosystem Carbon Change (NECC) is co-limited by stand characteristics, structural diversity, and environmental factors, with stand characteristics exerting the strongest influence, primarily via direct effects. (2) As stands develop, the impacts of structural diversity (effect increasing from 8.68% to 16.44%) and soil factors (from 8.80% to 10.30%) on productivity intensify. (3) In contrast, the influence of climate (decreasing from 30.40% to 17.67%) and terrain (from 14.55% to 6.28%) diminishes with advancing growth stages.
Discussion: This study provides a comprehensive, system-level analysis of the determinants of Net Ecosystem Carbon Change (NECC). By integrating multiple drivers, our work establishes a theoretical foundation for predicting Net Ecosystem Carbon Change (NECC) changes under global change scenarios. These insights are crucial for formulating effective forest management strategies to mitigate the challenges of climate change and biodiversity loss.
1 Introduction
Net Ecosystem Carbon Change (NECC) is a key component of carbon fluxes in terrestrial ecosystems and plays a vital role in advancing the understanding of global carbon cycles and land–atmosphere interactions (Shiri et al., 2022). NECC serves as an important metric for assessing the balance of the carbon cycle and is also a critical indicator for evaluating the effects of meteorological variations on ecosystem carbon balance. Therefore, clarifying the factors influencing NECC is essential for scientifically assessing the carbon source/sink capacity of a region. Many studies have evaluated NECC (Shuai et al., 2019; Zhang et al., 2024a). While this static perspective of NECC offers essential information on the present conditions of forest carbon storage, it does not adequately clarify the dynamic temporal changes within forest ecosystems and their responses to environmental variations. NECC is influenced by three processes: the growth of mature trees, the recruitment of new individuals, and carbon loss resulting from mortality (Yuan et al., 2019). For a long time, species diversity (Barrufol et al., 2013), structural diversity (Wang et al., 2021), climate (Jing et al., 2022), and stand characteristics (Lin et al., 2021) have been considered the primary determinants of NECC. However, in forests, the composition of tree species frequently changes as the forest ages, which is a process that might take several decades. During this period, the influence of environmental factors on ecosystem functioning may be altered (Zhao et al., 2024). Understanding the variation in NECC between phases of forest development, from the initial phase of secondary succession to old growth, is essential for restoring ecosystem functionality in damaged environments.
The prevalent view is that environmental elements in forests, including climate and site conditions, along with stand structure, significantly influence NECC (Du et al., 2022; Zhang et al., 2024a). Climate elements are regarded as the principal environmental determinants influencing NECC at a regional scale (Sun et al., 2021), with temperature and precipitation serving as the fundamental drivers of spatiotemporal patterns of NECC (Xi and Yuan, 2022). However, at smaller spatial scales, factors such as stand structure, terrain, and soil significantly influence the spatial variability of NECC (Jian et al., 2022). Furthermore, researchers have reported that in temperate larch forests, the beneficial impact of rising temperatures on the wood supply decreases as the forest matures. Forest production progressively increases, stabilizes, and subsequently decreases as the forest develops (Kira and Shidei, 1967). In northern and mountainous areas, elevated temperatures increase arboreal development and prolong the growing season of vegetation. Under climate change conditions, forests are expected to exhibit accelerated growth, reach maturity sooner, and experience earlier mortality, resulting in varied responses to climate change among forests of different ages (Collalti et al., 2019). Conversely, studies have determined that stand structure is positively connected with NECC (Wang et al., 2021), suggesting that within a stand, trees enhance NECC through resource allocation, facilitation, and biological feedback. Nonetheless, the significance of stand structure in forecasting NECC may evolve over time, as tree interactions intensify with growth during forest succession (Wang et al., 2022). Consequently, it is imperative to account for variations at various ages when forecasting the impacts of environmental conditions and stand structure on NECC.
Moreover, stand variables, including age and density, are crucial determinants of NECC. Stand density indicates the area used by trees and their internal horizontal structure, which affects the forms of tree development, NECC, and ecological stability (Liu et al., 2022). Increased stand densities enhance forest carbon storage and timber output by increasing canopy density, thereby increasing the amount of light captured. Stand age is a significant determinant of biomass and NECC (Liu et al., 2018). Stand age can increase biomass and NECC through increases in tree size (Becknell and Powers, 2014) and variations in size (Zhang and Chen, 2015). Throughout the forest restoration process, tree size, stand age, stand density, and species composition substantially change, whereas NECC markedly increases. Nonetheless, the primary biological processes influencing NECC remain a debated subject.
In recent years, machine learning technology has demonstrated superiority over traditional statistical methods by overcoming the limitations of big data analysis (Christin et al., 2018; Özçelik et al., 2013) and is regarded as a potent tool that is extensively utilized in forestry (Jevšenak and Skudnik, 2021; Lan et al., 2023; Wu et al., 2019). The random forest (RF) algorithm has been widely employed to assess variable significance and facilitate variable selection because of its ability to effectively mitigate overfitting and variation (Lin et al., 2004). Numerous studies have employed linear statistical approaches to examine the correlations between NECC and numerous variables, frequently overlooking the direct or indirect effects of these factors. Furthermore, identifying the ideal range of environmental variables using these methods requires substantial sample sizes and may not adequately elucidate the real mechanisms driving variations in NECC due to stand and environmental factors (Zhang et al., 2024a). Partial least squares structural equation modeling (PLS–SEM) is extensively employed to analyze the direct, indirect, and cumulative impacts of specific variables on other variables. This approach accommodates nonnormal distributions, nonlinear associations, and scenarios with numerous variables (Hair et al., 2021). Integrating the RF model with PLS–SEM to investigate the physiological principles of NECC may enhance the understanding of forest dynamics and serve as a reference in the context of impending climate change.
The Da Xing’an Mountains, situated in Northeast China, form a delicate mountainous region with abundant resources (He et al., 2021). This abundant-resource region maintains an unspoiled cold-temperate coniferous forests ecology, being China’s sole boreal coniferous forest area and one of the few extant biological gene pools (Zhao et al., 2024). The forests of the Da Xing’an Mountains are essential for ecological security in northeastern China and the broader North China region, and their function in the carbon cycle is significant (Jiang et al., 2002). This location has garnered considerable interest from forest managers in recent research (Wang et al., 2008). Nonetheless, prolonged human logging activities have significantly compromised stand structure, functionality, and stability, hence substantially impeding the sustainable growth of the forest ecosystem in the Da Xing’an Mountains (Wu et al., 2022). Consequently, statistically assessing the impacts of stand structure and environmental variables on NECC and providing scientifically valid restoration strategies are imperative. This assessment is crucial for reinstating biological functions, increasing forest quality, and increasing the capacities of carbon sequestration and sinks in the Da Xing’an Mountains.
In this research, 782 natural forest plots were evaluated, and RF and PLS–SEM were utilized to ascertain the direct and indirect influences of stand characteristics, structural diversity, and environmental variables on NECC. Our dataset included forests at various developmental stages, from the initial phases of secondary succession to mature old-growth forests. We categorized these plots into five forest developmental stages on the basis of stand age to investigate the variations in production as the forests matured. Consequently, we propose ways to increase Net Ecosystem Carbon Change in the Da Xing’an Mountains to improve forest quality. The primary aims of this study are as follows:(1) How do stand characteristics, structural diversity, and environmental variables collectively influence NECC in the cold-temperate coniferous forests of the Da Xing’an Mountains? (2) What are the relative impacts and evolutionary trends of stand characteristics, structural diversity, and environmental variables on NECC across different stages of forest development? For these objectives we use the conceptual model outlined in Yuan et al. as a starting point(Figure 1), we tested the following hypotheses: (1)All variables had direct effects on growth, recruitment and mortality, and indirect effects on net ecosystem carbon change. (2)Topography affected climate, structure diversity, soil, stand. (3)Climate affected structure diversity, soil, stand. (3)Structure diversity affected soil, stand. (4)soil affected stand.
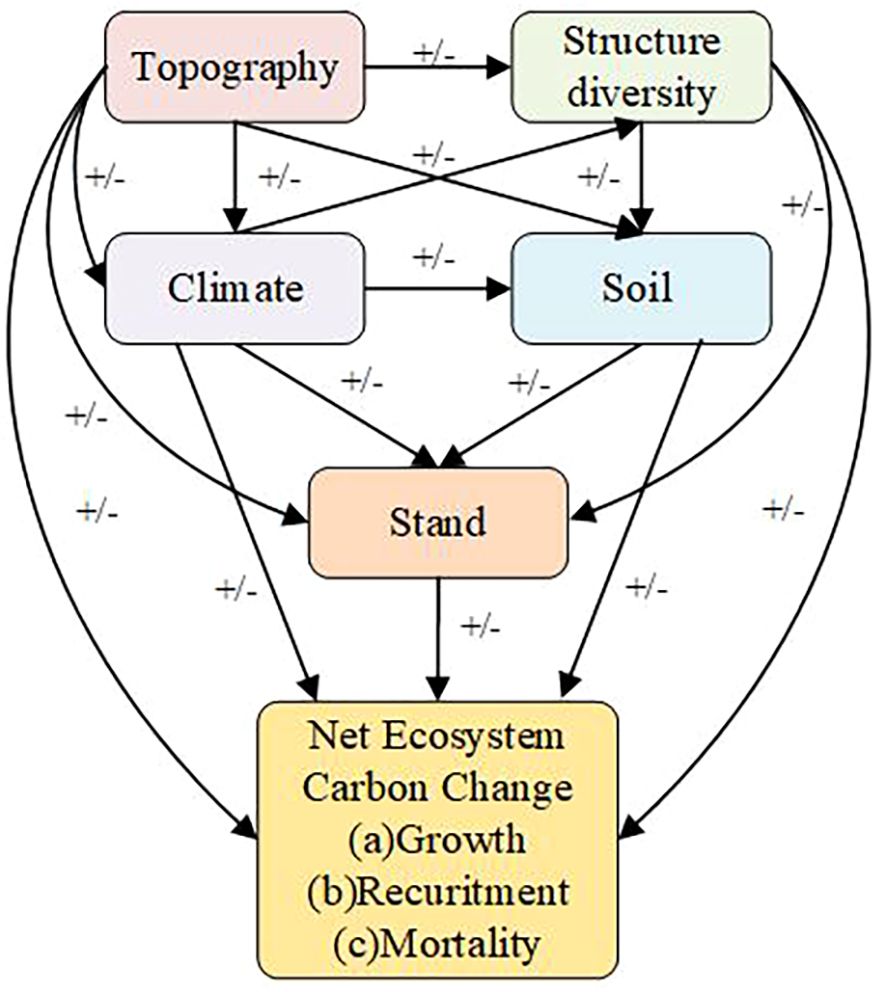
Figure 1. A conceptual model revealing the expected links of environmental factors (topography, climate, and soil nutrients), structure, and stand factors on net ecosystem carbon change demographic processes (recruitment, growth, and mortality). Hypothesized positive, negative, and unknown effects are indicated by +/- signs.
2 Materials and methods
2.1 Study area and sample plot data
The research area is located in the key state-owned forest region of the Da Xing’an Mountains in Heilongjiang Province. Geographically, the area is located in northwestern Heilongjiang Province, northeastern Inner Mongolia Autonomous Region, and on the northeastern slope of Da Xing’an ridge (Figure 2). It spans 121°10’53”-127°01’21”E, 50°07’02”-53°33’42” N and encompasses a total area of 8.02 × 106 hectares. The area has a distinct cold–temperate continental monsoon climate, with an annual average temperature of -2 °C. Annual precipitation values range from 430–460 mm, and precipitation is concentrated from July–September. The primary soil types in the area include brown coniferous forest soil, dark brown soil, gray–black soil, meadow soil, and marsh soil. The climax community of the forest ecosystem in the Da Xing’an Mountains comprises a bright coniferous forest typical of a cold–temperate zone. The dominant tree species include Larix gmelinii, Betula platyphylla, Populus davidiana, Quercus mongolica, and Pinus sylvestris var. mongolica, among others.
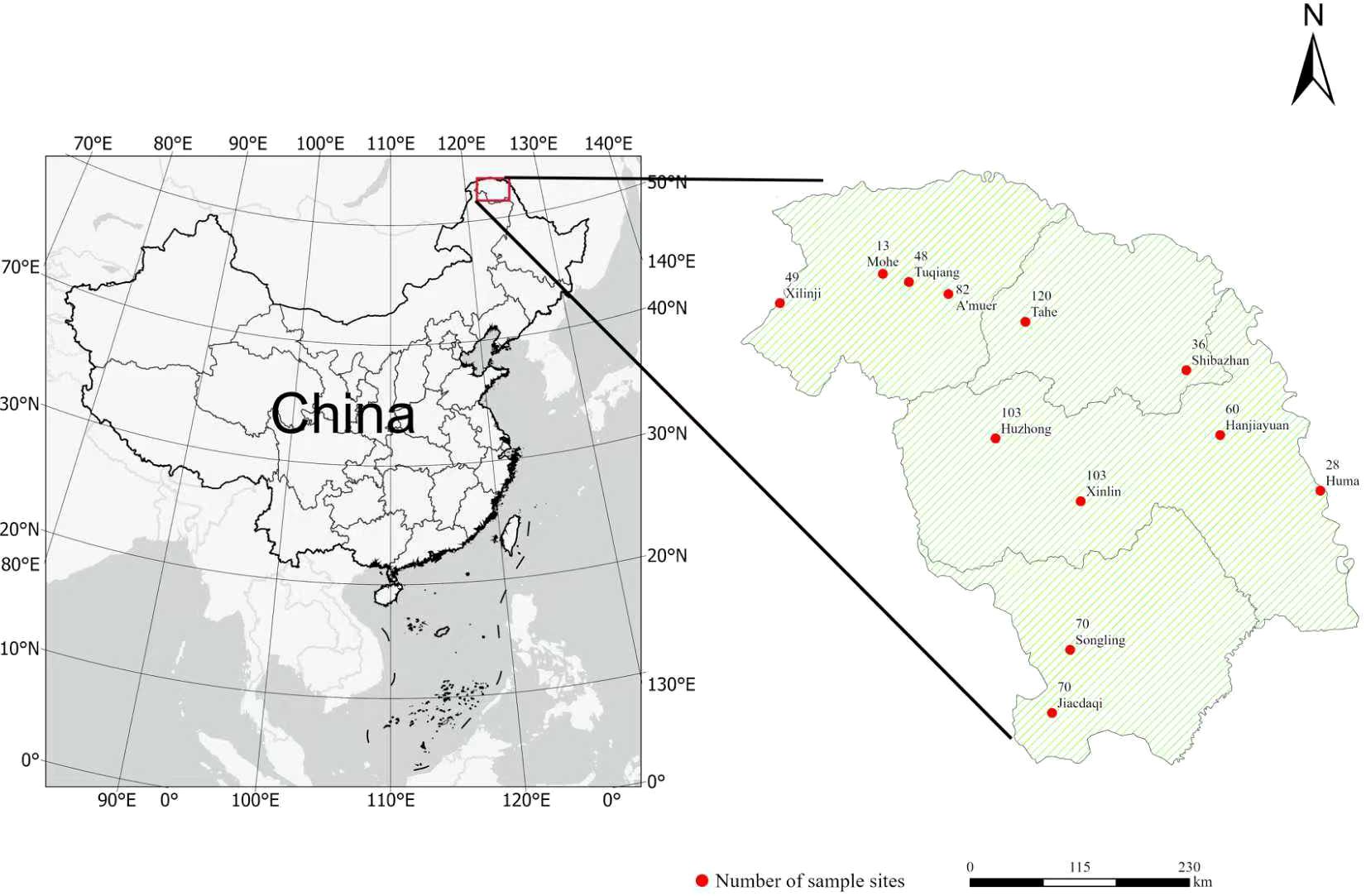
Figure 2. Geographical locations of the study area in the eastern Da Xing’an Mountains, Northeast China.
The data for this study were sourced from the seventh and eighth national forest resource inventories (NFCIs) of the Da Xing’an Mountains. Utilizing systematic sampling and a kilometer grid approach, 782 sample plots were set up over a span of five years. In the Da Xing’an Mountains, the fixed sample areas were organized in a grid measuring 8 km by 8 km, each spanning 0.06 hectares. Measurements were taken for various tree species, breast height diameters (≥ 5 cm), and the mean age of the stands in every fixed plot.
2.2 Calculating carbon sequestration in the forest stands of permanent sample plots
The biomass of each tree was obtained by substituting the tree measurement data into a biomass model. Subsequently, the biomass was multiplied by the ratio of the carbon content of each species to ascertain the carbon storage capacity of every tree. The term ‘growth’ denotes the variation in the carbon sequestration of preserved trees across two survey intervals; ‘recruitment’ indicates the rise in carbon sequestration due to trees having a diameter at breast height (DBH) < 5 cm in the initial survey, increasing to 5 cm in the subsequent survey; ‘mortality’ denotes the reduction in carbon storage resulting from tree death during the survey. This research defines NECC as the aggregate of growth, mortality, and recruitment.
2.3 Stand variables
The variables in our study consisted of stand age, stand density, and DBH. The stand age was ascertained by computing the arithmetic mean of the ages of the typical dominant trees via tree ring analysis. The DBH for each plot was computed for all trees within the plot. The stand density (N·ha−1) was calculated by dividing the total tree count in the plot by the plot area.
2.4 Structural diversity indicators
We employed the diversity index of the tree species composition to delineate structural diversity. This index was utilized to evaluate data, including the quantity of tree species, their relative abundance, and the fraction of species biomass. It also accurately depicts the uniformity and extent of species intermingling within a stand.
2.5 Environmental data
Climate data were obtained from the ClimateAP (v2.30) application, which generates climate variables on the basis of latitude, longitude, and elevation of the sample plots. We obtained soil data from the Harmonized World Soil Database (HWSD) of the Food and Agriculture Organization (http://www.fao.org/faostat/en/#data) (Milovac et al., 2018), which offers extensive soil property and nutrient information at a resolution of 1000 m. Using kriging interpolation, the data were resampled to a 30 m resolution and extractable by the geographic coordinates of a sample plot. Topographic data were acquired during the creation of sample plots, and a GPS was used to identify and document topographic information accurately. In total, there were 16 climate factors, 10 soil variables, and 3 topographic variables, as shown in Table 1.
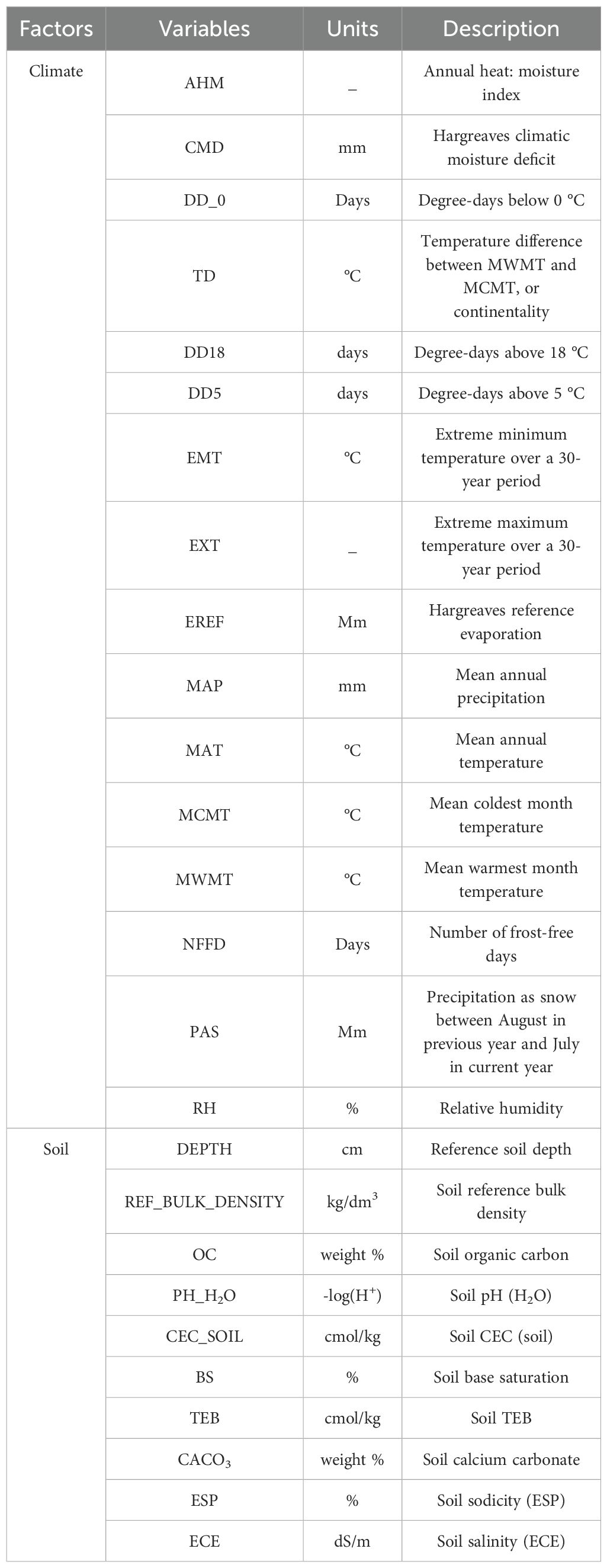
Table 1. Description of the environmental data used in this study.
2.6 Statistical analyses
Pearson correlation analysis was used to initially assess the significant association of prospective predictors associated with NECC (at the 0.05 level) and the threshold for the correlation coefficient between predictor variables and NECC. The predictors were utilised for subsequent analysis only if the absolute values of their correlation coefficients above 0.2 (threshold of 1). Second, the absolute value of the correlation coefficient among the significant candidate variables themselves was guaranteed to be < 0.4 (threshold 1). A predictor was excluded if the absolute value of the correlation coefficient between itself and all other predictors was greater than 0.4, and the variance inflation factor (VIF) was used in stepwise regression analysis to evaluate the multicollinearity. All the VIF values < 10 indicate that collinearity between variables has no significant impact on our results (Graham, 2003). The VIF and Pearson methods were implemented in SPSS software (version 23.0).
The RF algorithm chooses the variables on the basis of the importance scores of the input variables. In the RF algorithm, the importance value was calculated by permuting on out-of-bag (OOB) data: (1) the prediction error (the mean sum of the squares of residuals, MSE) on the OOB portion of the data was recorded for each tree, (2) the same was done after permuting each predictor variable, and (3) the difference between the two was then averaged over all trees as importance scores (Grömping, 2009). The importance scores of all the predictors were normalized to a percentage. The RF method was implemented in the randomForests package in the R platform (version 4.3.2).
Before statistical analysis was conducted, the classical theory of ecology was used to divide the life cycle of a forest into five developmental stages: young forest, middle-aged forest, near-aged forest, mature forest, and overaged forest. During various stages, changes in growth rates, biomass buildup and photosynthetic efficiency result in different degrees of NECC. Consequently, tree species and age were categorized into separate classifications for analysis. Consequently, we categorized the trees on the basis of various tree types and stand ages. Larix gmelinii, Pinus sylvestris, and Picea asperata were categorized on the basis of the following age ranges: ≤ 40, 41–60, 61–80, 81–120 and > 120 a. Populus davidiana and Betula platyphylla were categorized into the following age groups: ≤ 30, 31–50, 51–60, 61–80 and > 80 a. The age classification for Quercus mongolica and Betula davurica was as follows: ≤ 40, 41–60, 61–80, 81–120 and > 120 a. PLS–SEM was employed to investigate the direct, indirect, and interactive links among different variables affecting the response ratio (RR) of agricultural yields. The net effect of one variable on another was determined by integrating all direct and indirect pathways connecting the two variables. The route coefficients and coefficients of determination (R²) were computed using the R package “plspm”. All data analysis was conducted using R version 4.0.2.
3 Results
3.1 Changes in net ecosystem carbon change traits with forest development
The mean carbon reserves in this region’s natural forests stood at 33.35 t·hm-2 in 2010, which was a 4.38 t·hm-2 increase from 2005. On average, NECC was 1.55 t·hm-2·a-1, accompanied by growth, recruitment, and mortality rates of 0.88 t·hm-2·a-1, 0.23 t·hm-2·a-1, and 0.43 t·hm-2·a-1, respectively. Generally, as forests aged, there was a notable reduction in both growth rates and entry rates, alongside an increase in mortality rates. The mortality percentages for forests categorized as young, middle-aged, near-mature, mature, and overaged were 6.73%, 24.66%, 19.28%, 23.77%, and 30.49% of the overall mortality, respectively (Table 2).
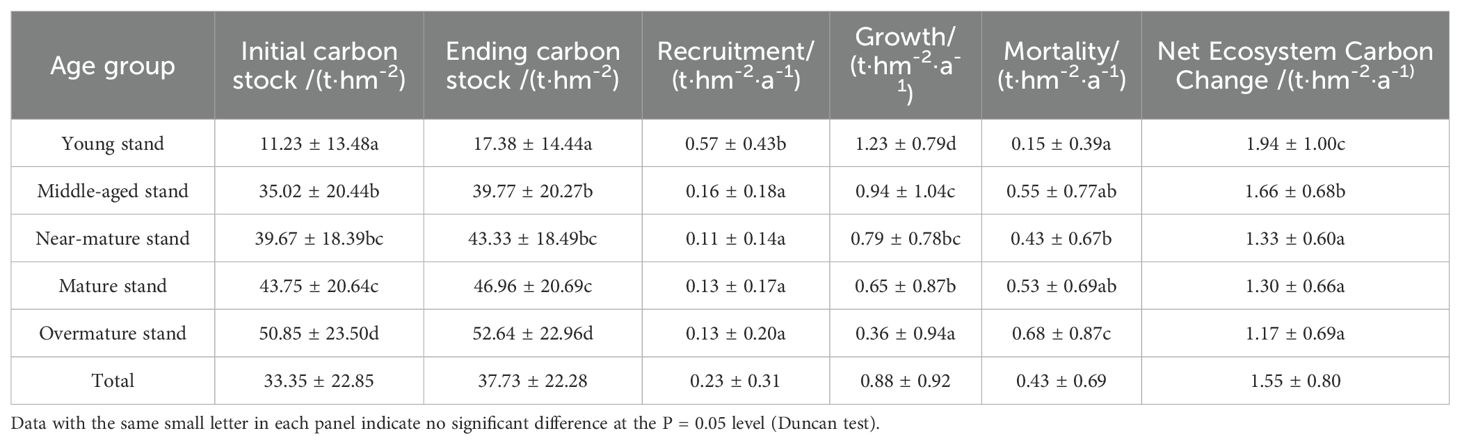
Table 2. Statistics of the basic characteristics of the sample plots.
3.2 Significant variables affecting stand biomass via multiple feature selection methods
In the examination of NECC, considerable discrepancies were observed in the number of variables chosen by the three feature selection approaches, and the selected variables varied among these methods. For example, relative humidity (RH) values were identified as significant using correlation analysis and redundancy analysis but excluded by alternative methods. Moreover, many feature selection strategies exhibited limited consistency in terms of the chosen variables. Age, Dg and density in stand structural diversity were consistently significant variables across all three techniques. Ultimately, five critical biodiversity factors were identified by the intersection approach, as follows: (1) climate: MAT and MAP; (2) stand: age, mean diameter at breast height (DBH, Dg), and density; (3) soil: depth; (4) structural diversity: ISCD; and (5) topography: altitude.
3.3 Results of partial least squares path modeling
The variables identified by the VIF, Pearson, and RF methods were incorporated into the PLS–SEM with NECC (growth, recruitment, and mortality). The PLS–SEM performed well, with a strong explanatory ability for the causal paths. Specifically, the AVE, alpha, CR, and Rho_A values indicated that the model’s fit was within an acceptable range. The goodness-of-fit (GOF) values revealed that the model’s overall quality was quite high (Table 3). The analysis indicated that stand factors, structural diversity, and environmental variables contributed 68.2%, 39.2%, and 35.2%, respectively, to the variability in growth, recruitment, and mortality. Furthermore, growth, recruitment, and mortality jointly accounted for 93.8% of the variation in NECC, underscoring their pivotal role in influencing NECC dynamics (Figure 3).
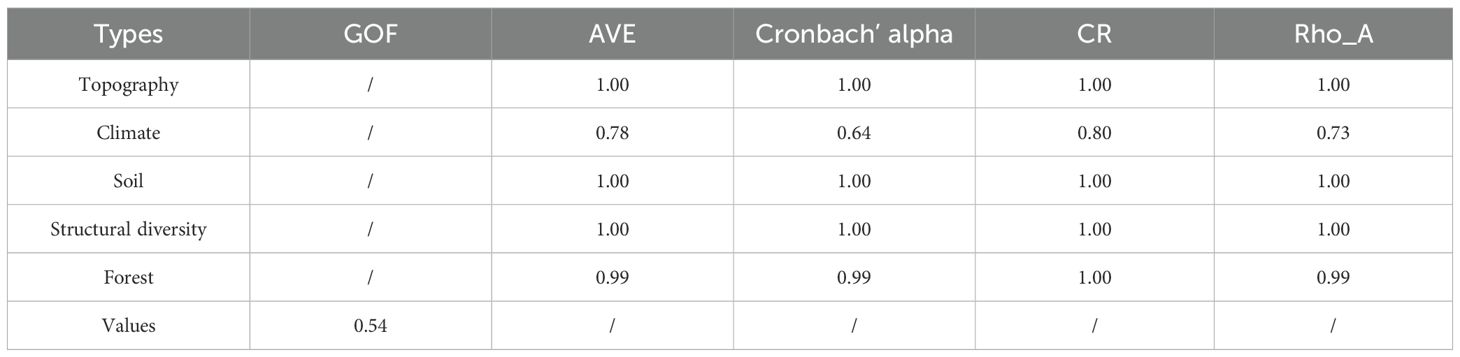
Table 3. Model performance of the PLS–SEM.
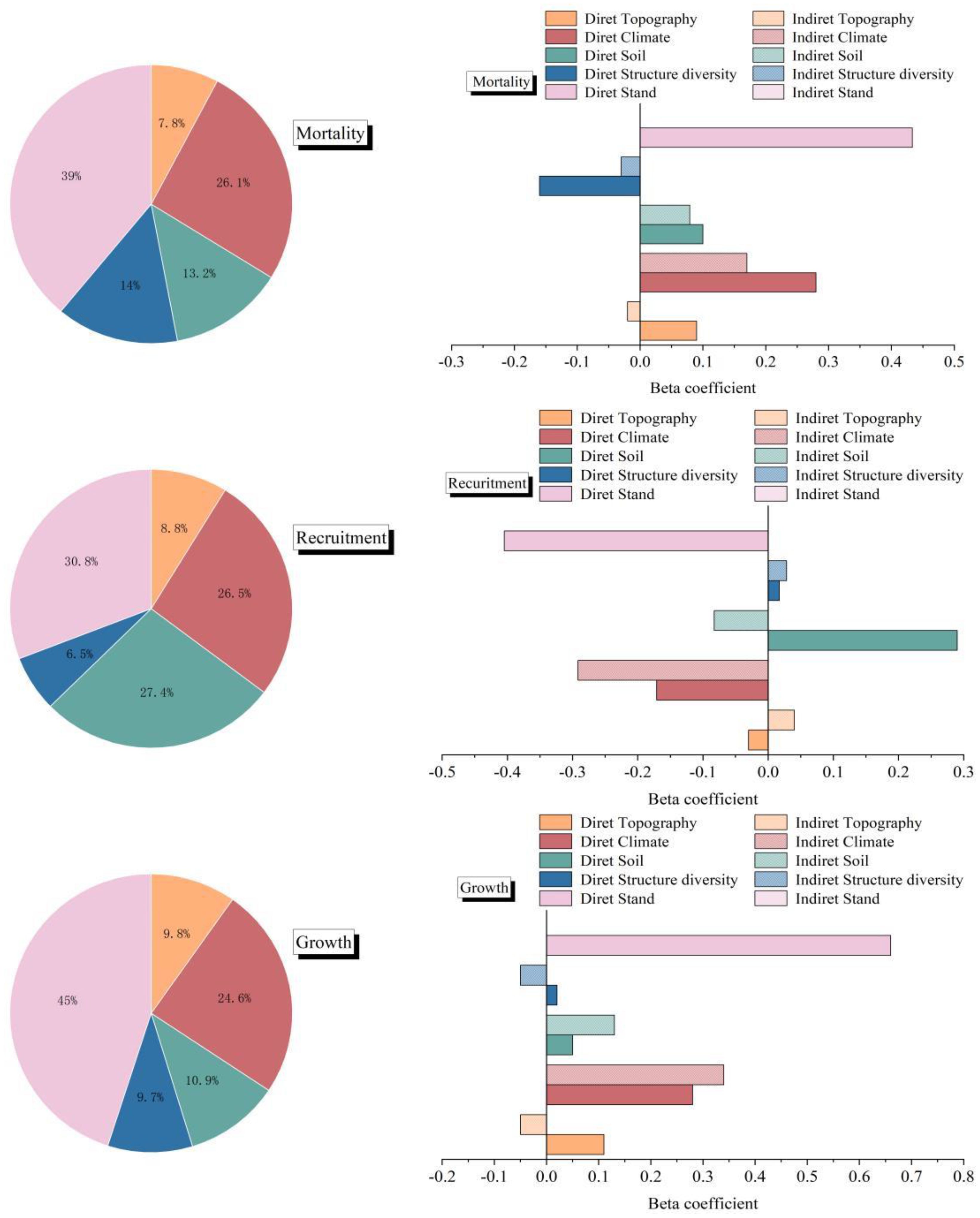
Figure 3. Beta coefficients and the relative contributions of stand factors, structural diversity, and environmental variables to net ecosystem carbon change. The filled bars indicate direct effects, and the striped bars indicate indirect effects of stand factors, structural diversity, and environmental variables on growth, recruitment, and mortality. The pie charts show the relative importance of each predictor for growth, recruitment, and mortality.
The growth of a stand was directly influenced by factors such as stand (b=0.66), climate (b=0.28), topography (b=0.05), structural diversity (b=0.02), and soil (b=0.08; Figure 4a). The increase in stand growth was particularly evident due to stand features, as optimal tree density fosters stand efficacy, facilitates canopy shading, and improves water-use efficiency. Nonetheless, overall, topography constrained stand growth, as elevated altitudes typically result in lower temperatures, which is a critical determinant of plant growth, hence substantially impeding vegetation survival and development. The advantageous influence of climatic circumstances on NECC was significant, particularly as regions with elevated temperatures and more precipitation typically exhibit enhanced forest growth (Figure 4a).
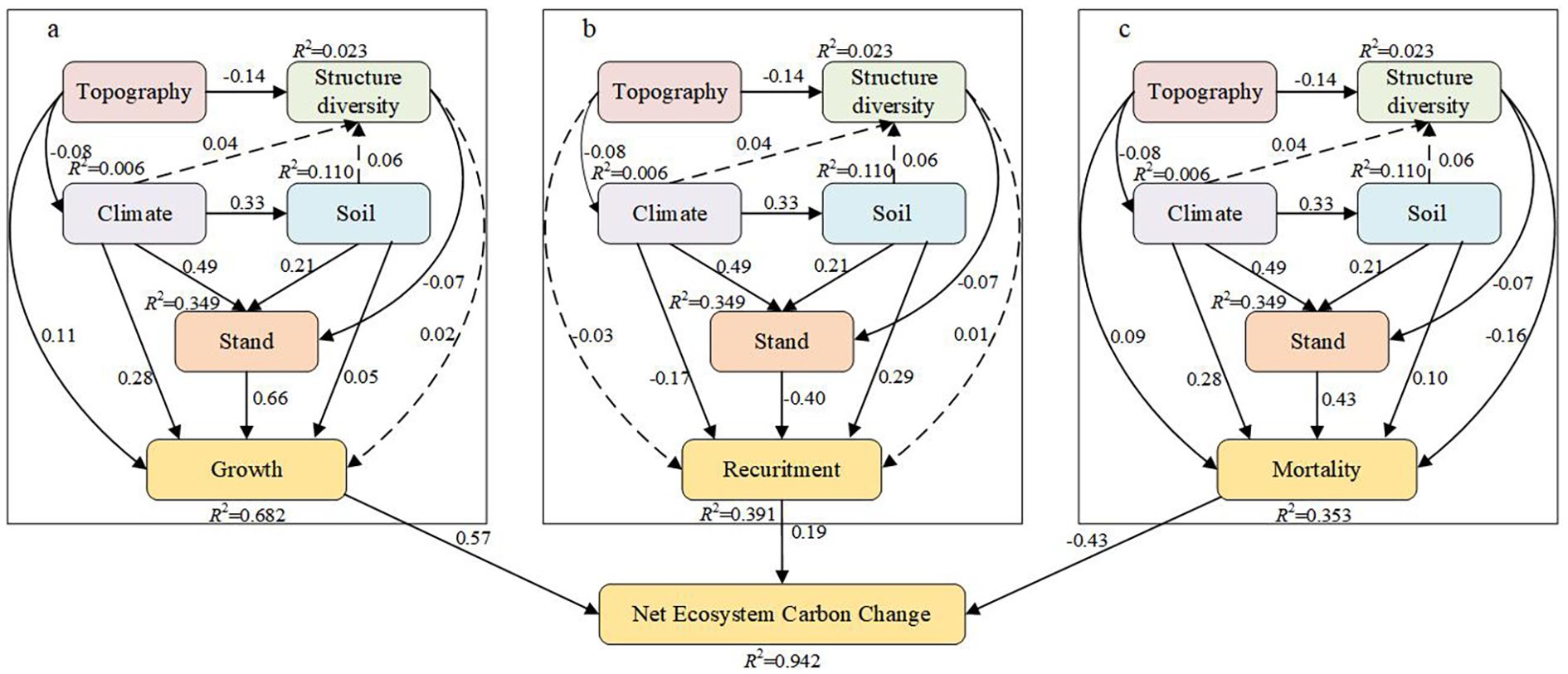
Figure 4. PLS–SEM revealed the effects of stand factors, structural diversity, and environmental variables on Net Ecosystem Carbon Change The climate variables included MAT and MAP; the stand attributes included DBH, stand density, and stand age; the topographic factor was elevation; the soil property was soil depth; and the structural diversity was represented by the diversity index of the tree species composition. (A) The effects of stand factors, structural diversity and environmental factors on Growth. (B) The effects of stand factors, structural diversity and environmental factors on Recuritment. (C) The effects of stand factors, structural diversity and environmental factors on Mortality. The solid and dotted lines indicate that the path is significant (P<0.05) and insignificant (P>0.05), respectively.
The PLS–SEM analysis for recruitment indicated that stand factors exerted the most substantial positive direct influence on recruitment (b = -0.40). Structural variety and soil were positively correlated with recruitment (b=0.02, b=0.21), but climate had a negative impact on recruitment (b=-0.17; Figure 4b). Although topography did not exert a substantial direct influence on recruitment, both factors indirectly affected recruitment because of their negative correlation with climate and positive association with stands (Figure 4b).
The PLS–SEM analysis of mortality revealed the indirect impacts of topography (b = 0.10), climate (b = 0.28), structural variety (b = -0.19), and soil (b = 0.19) on mortality via their interactions with stands (b = 0.43; Figure 4c). In the statistical examination of the three separate carbon pools, growth and death constituted the primary sources of variation in NECC, with recruitment following thereafter (Figure 4c).
3.4 Factors controlling net ecosystem carbon change across different stand ages
We constructed a PLS-SEM to examine the linkages among topography, climate, structural diversity, stand characteristics, soil properties, and NECC. Our analysis revealed significant direct and indirect effects of topography, climate, structural diversity, stand characteristics, and soil on NECC. PLS-SEM analyses were also conducted separately for five distinct stand development stages. We confirmed that the influence of topography on NECC decreased with stand development. The direct effects of topography on growth and mortality were strongest and statistically significant in the seedling stand stage (Figure 5a). These direct effects diminished and became non-significant in subsequent development stages. Similarly, the influence of climate on NECC declined with stand development. The direct effects of climate on growth and recruitment peaked during the seedling stage (Figure 5a). Notably, climate’s effect on recruitment became non-significant in the immature stand stage (Figure 5b). Unexpectedly, climate’s effect on mortality exhibited a strengthening trend as stands developed. Conversely, the influences of soil and structural diversity on NECC increased with stand development. The direct effect of soil on NECC reached its maximum in the mature (Figure 5d) and over-mature stand stages (Figure 5e). Structural diversity’s effect on growth and mortality shifted from negative in the seedling stage (Figure 5a) to positive in the immature stand stage (Figure 5b).
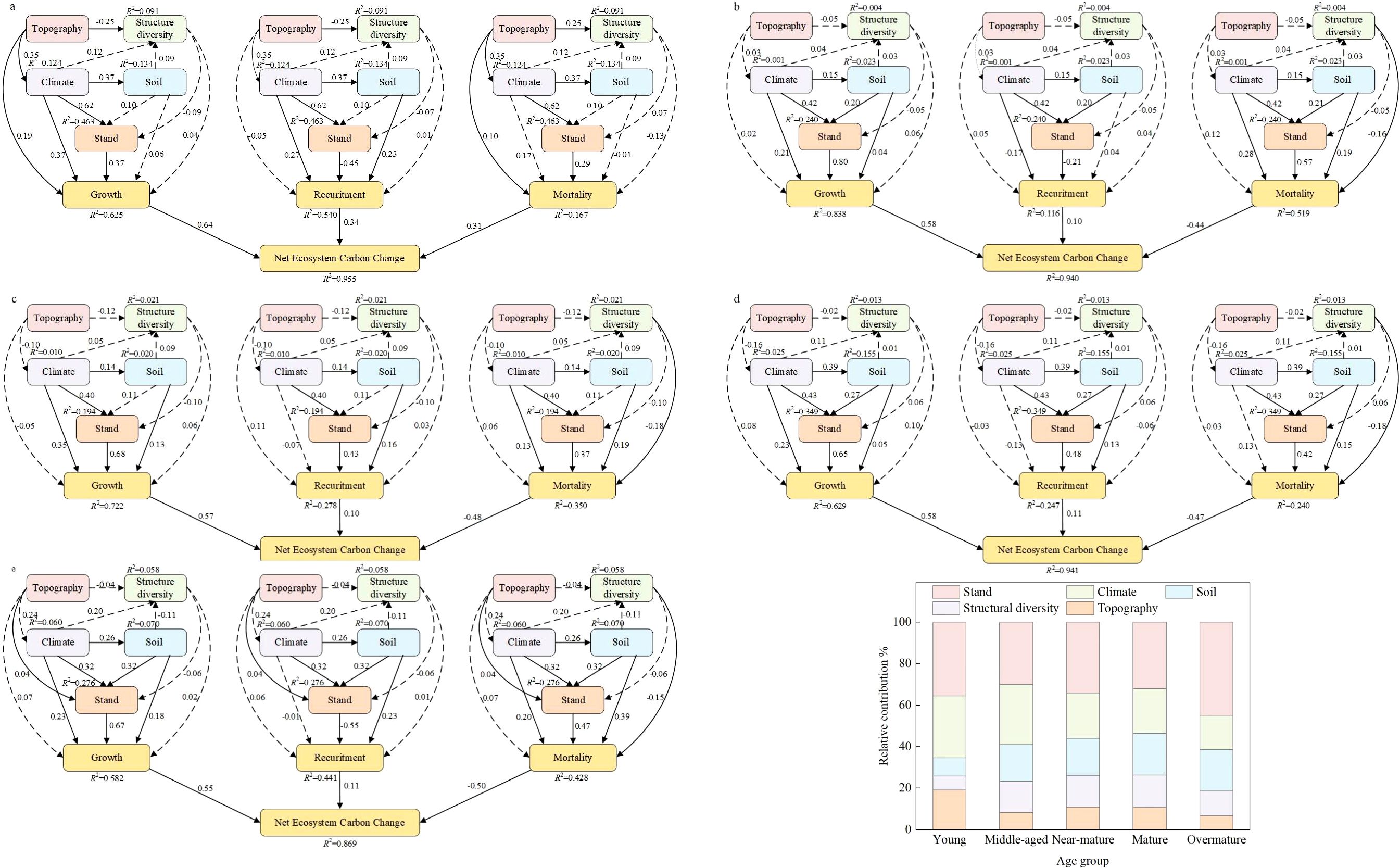
Figure 5. PLS-SEM relating net ecosystem carbon change to the influencing factors at the five development stages. (a) young stand, (b) Middle-ages stand, c near-mature stand, (d) mature stand, (e) Over-mature stand. (f) The relative contribution of the influencing factors on net ecosystem carbon change based on variance partitioning analysis at the five development stages.
Across the five development stages, the influence of climate (from 30.40% to 17.67%) and topography (from 14.55% to 6.28%) on NECC decreased with stand development. Conversely, the influence of structural diversity (from 8.68% to 16.44%) and soil (from 8.80% to 10.30%) increased across the stand development stages (Figure 5f). However, the effect of structural diversity on NECC was most pronounced in the sub-mature stand stage, while the effect of soil properties was most pronounced in the immature stand stage. Furthermore, as stands approached the mature/over-mature stage, the relative importance of structural diversity to NECC exceeded that of the environmental variables (Figure 5f).
4 Discussion
4.1 Impact of stand factors and structural diversity on net ecosystem carbon change
We employed PLS–SEM to analyze the multivariate links driving stand factors and structural diversity. The PLS–SEM effectively elucidated the relationships utilizing the variables chosen through the intersection strategy. The stand variables (age, dg, and density) significantly contributed to net ecosystem carbon change. Researchers have demonstrated that stand density directly influences the environment in which trees develop, including light, heat, temperature, humidity, and soil nutrients (Cui et al., 2022). At low stand densities, tree interactions are minimal or weak, rendering niche complementarity effects insignificant. As stand density escalates, interactions intensify, with trees occupying greater space and utilizing more resources (Forrester et al., 2013; Morin, 2015). Initially, NECC increases as trees grow and the stand matures over time. However, as forests evolve from middle-aged to fully mature, there is an increase in the need for nutrients and water to facilitate the outward growth of trees, simultaneously increasing the rates of transpiration.
The outcomes of our study reveal a beneficial association between NECC and increased structural diversity. Previous studies have validated the influence of structural diversity on forest output, which has been successfully included in predictive models for NECC (LaRue et al., 2023). Importantly, an increase in structural diversity amplifies interactions, such as competition among trees. A varied assortment of structures generates openings in the canopy, enhancing light penetration and promoting a vibrant natural habitat. Layered canopy structures, along with diverse layering of trees, frequently occupy greater spatial areas and exploit a broader range of resources, which is a concept that is elucidated by the principle of niche complementarity (Morin, 2015). The complex design of a stand contributes to minimizing temperature fluctuations, enhancing soil moisture retention, facilitating litter decomposition, and promoting nutrient recycling, thereby improving resource utilization efficiency and increasing both NECC and biomass growth (Crockatt and Bebber, 2015; Schwarz et al., 2014). Although cold–temperate forests lack the characteristic stratification found in subtropical forests, their structural diversity is essential for influencing ecosystem services.
4.2 Impact of environmental factors on net ecosystem carbon change
Observations reveal that climate variables, specifically MAT and MAP, exert the greatest influence on NECC among environmental factors, with increasing temperatures and precipitation increasing NECC. Temperature fluctuations contribute to clarifying the spatial pattern of NECC (Liu et al., 2014; Ni et al., 2022). Moreover, the efficacy of forests is constrained by water availability, as precipitation governs water distribution, subsequently affecting tree habitats. The findings of our study indicate that geographical factors, particularly elevation, significantly affect NECC. This was due mainly to topographic factors affecting the spatial distributions of solar radiation and precipitation, together with soil moisture, nutrients, and depth, resulting in intricate impacts on NECC within different ecosystems (Wu et al., 2021). Nevertheless, elements such as slope and gradient have been identified as critical drivers influencing NECC in some studies (He et al., 2023; Jafarian et al., 2023). However, within the parameters of our research area, the impact of these components is less pronounced than that of height. This may be attributed to the predominant tree species (Larix gmelinii and Betula platyphylla) exhibiting markedly low sensitivity to slope orientation and gradient. Larix gmelinii, recognized for its strong cold resistance and moderate water requirements, can thrive on both steep and shaded slopes. Betula platyphylla necessitates increased moisture; nonetheless, its tolerance to shadow and cold enables it to flourish over diverse slopes and gradients, mitigating the effects of fluctuations in light and temperature.
4.3 Beneficial impact of forest age on net ecosystem carbon change via structural diversity and soil factors
The effects of soil and especially structural diversity on NECC increased with stand age, as predicted. These greater effects may be partly due to increasing tree–tree interactions and complementary effects (Wang et al., 2022), coupled with increased build-up of soil organic matter and heightened microbial activity in the soil, which can intensify the impact on NECC. With respect to forest age, the greater positive effects of structural diversity with stand age illustrate that processes such as resource partitioning, facilitation or trophic interactions may result in greater benefits for tree growth in plots with high stand ages than in plots with low stand ages (Hatami et al., 2020; Zhang et al., 2020). This finding is supported by those of previous studies showing that as a forest advances through its developmental phases, its structure becomes increasingly intricate, and the linkages between resource distribution among trees and biodiversity become more evident. Structural variety enhances resource utilization efficiency and stabilizes ecosystem functioning by affecting tree distribution, density, canopy architecture, and interorganism interactions (Aakala et al., 2013; Buechling et al., 2017; Jiang et al., 2018). In addition, as the trees in the forest undergo maturation, organic matter from litterfall and root systems progressively accumulates, leading to an increase in the soil organic matter content. This accumulation not only augments soil fertility but also improves the physical structure of the soil (Post and Kwon, 2000), including the formation and stabilization of soil aggregates, thereby augmenting soil aeration and water retention capacity. Soil structure directly influences root development and nutrient assimilation. A well-developed soil structure facilitates the efficient cycling of water and nutrients, which is imperative for the optimal growth and vitality of trees (Jandl et al., 2007).
4.4 Role of forest age in mitigating the impact of climate and topographical variations on net ecosystem carbon change
The results of our study reinforce the mitigating effect of forest age in mitigating climate and topographic shifts, showing that mature forests are less susceptible to climate and topographical changes than their younger counterparts are. Research has indicated that mature forests are more sensitive to climate change than younger forests are (Zhang et al., 2024b). This disagreement may arise from variations in the chosen study regions, as the Da Xing’an Mountains are situated in a cold–temperate humid to semihumid zone. The growth dynamics of several species may be affected by habitat factors. Trees in arid environments are more vulnerable to climate change than those in humid environments are, irrespective of their presence in mature or young forests (Xue et al., 2025). The first piece of evidence proves that there is indeed a dampening effect of forest heterogeneity within the ecosystem on the climate sensitivity of forests, which can be related to the fact that multiage forests benefit from increasing structural complexity (de Wergifosse et al., 2022; Jandl et al., 2019). Compared with young forests, overaged forests exhibit greater diversity in the age structure, characterized by a combination of young, middle-aged, and mature trees. The creation of vertical stratification enhances ecosystem complexity and stability. This stratification provides gradients in biomass (deadwood and living aboveground biomass) and different ways to allocate carbon (Maréchaux et al., 2021). This stratification has also enabled the development of unique carbon distribution tactics and distinguished the ecological roles of plants and animals, thus increasing biodiversity and bolstering the ecosystem’s ability to withstand environmental disruptions (Lafond et al., 2014; Pardos et al., 2021). From a functional perspective, a forest of diverse ages exhibits a variety of physiological and life cycle characteristics among its trees. This variety translates into a greater capacity to absorb and store carbon, thereby mitigating the effects of climate change, even in a mosaic of even-aged patches, as we simulated in this study. Young trees grow rapidly and absorb significant amounts of CO2 from the atmosphere, contributing to carbon capture, and are more efficient in converting photosynthates in biomass (Campioli et al., 2015; Collalti et al., 2014; Viet et al., 2023). In contrast, older trees accumulate more biomass and serve as long-term carbon sinks and regenerative shelters. Concurrently, the increased diversity between age groups offsets the beneficial and detrimental effects linked to each age group. Consequently, in fluctuating weather scenarios, the existence of various age groups in forest communities is vital for preserving their functional variety since this diversity offers advantages in terms of resilience and the ability to adapt to climatic shifts (Ehbrecht et al., 2021; Kauppi et al., 2022; Zampieri et al., 2021). Furthermore, the age of forests plays a role in lessening the impact of topographical alterations. Forest age distribution plays a role in shaping how topography impacts forest ecosystems (Durán Zuazo and Rodríguez Pleguezuelo, 2008; Selkimäki et al., 2012). For example, forests of various ages, owing to their intricate structures, might show greater resilience to topographical changes such as soil erosion and landslides. Such varied structures aid in stabilizing the soil, diminishing erosion, and increasing the forest’s ability to retain water.
5 Limitations
The factors influencing NECC were investigated over a relatively short period from 2005 to 2010. An important limitation of this study, which must be acknowledged, is that five years is insufficient to capture the dynamics of slow-growing cold-temperate coniferous forests. Short-term fluctuations caused by extreme climate events, insect outbreaks, or other transient disturbances may disproportionately influence the results. Such events often operate on longer time scales and can profoundly influence forest development pathways; however, short-term data may fail to adequately capture their full effects. Furthermore, although species-specific age classification criteria were applied during data processing, the model was not stratified by individual tree species. It is recognized that significant differences may exist among species in developmental stages—for example, a 40-year-old birch may be considered mature, while a larch of the same age is still young.These differences in growth dynamics and life history strategies could potentially influence the relationships among model variables. Although the current modeling approach aggregated species to maintain sufficient statistical power and model stability, it inherently assumes a certain degree of ecological similarity in stand development across species. Future studies should incorporate extended datasets, integrate assessments of extreme climatic events, and combine species-specific analyses or multi-group structural equation modeling to more accurately evaluate potential impacts on forest ecosystems. Nonetheless, the commonality in stand dynamics and silvicultural practices within the studied forest system provides an ecological rationale for the integrated modeling approach adopted in this study.
6 Conclusions
Using NFCI data from 2005 to 2010, PLS–SEM was employed to examine the correlations among stand variables, structural diversity, and environmental variables in assessing the NECC of natural forests. Our research indicates that stand variables are the principal determinants of NECC, with direct influences being notably substantial. Furthermore, the age of forests enhances the impact of structural variety and soil on NECC while alleviating the effects of climate and topography. In light of impending climate change, forest management practices must be customized to various stages of forest development. In young forests during the early successional stage, the application of moderate and selective thinning techniques can diminish competition among trees, foster the establishment of dominant species, and preserve a degree of species diversity to improve structural complexity. In middle-aged and near-aged forests in the mid-successional and mature phases, implementing moderate thinning and selective logging can foster a multilayered vertical structure, optimize light conditions, promote understory regeneration, and increase ecosystem stability. In mature and overaged forests in the late successional stage, excessive intervention should be curtailed while preserving a proportion of standing dead trees and fallen logs to increase soil nutrient cycling and fertility. Furthermore, artificial regeneration can be effectively executed by introducing seedlings of varying ages to improve the age structure variety within a stand. The implementation of these measures enables forests to significantly contribute to climate change mitigation, improve carbon sequestration, and preserve biodiversity. Our research findings establish a foundational theoretical framework for the development of sustainable forest management practices in the Da Xing’an Mountains of Northeast China.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
ZW: Methodology, Validation, Writing – original draft. LD: Conceptualization, Supervision, Writing – review & editing. YH: Conceptualization, Supervision, Writing – review & editing. ZM: Writing – review & editing. XJ: Writing – review & editing. XZ: Writing – review & editing. SC: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was financially supported by the National Key R&D Program of China (No. 2022YFD2201001), the Joint Funds for Regional Innovation and Development of the National Natural Science Foundation of China (No. U21A20244), and the Heilongjiang Touyan Innovation Team Program (Technology Development Team for High-efficient Silviculture of Forest Resources).
Acknowledgments
We are grateful to the reviewers for their valuable comments that helped improve the manuscript’s quality. This project was funded by the National Key R&D Program of China (No. 2022YFD2201001), the Joint Funds for Regional Innovation and Development of the National Natural Science Foundation of China (No. U21A20244), and the Heilongjiang Touyan Innovation Team Program (Technology Development Team for High-efficient Silviculture of Forest Resources).
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1663271/full#supplementary-material
References
Aakala, T., Fraver, S., D’Amato, A. W., and Palik, B. J. (2013). Influence of competition and age on tree growth in structurally complex old-growth forests in northern Minnesota, USA. For. Ecol. Manage. 308, 128–135. doi: 10.1016/j.foreco.2013.07.057
Barrufol, M., Schmid, B., Bruelheide, H., Chi, X., Hector, A., Ma, K., et al. (2013). Biodiversity promotes tree growth during succession in subtropical forest. PLoS One 8, e81246. doi: 10.1371/journal.pone.0081246
Becknell, J. M. and Powers, J. S. (2014). Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest. Can. J. For. Res. 44, 604–613. doi: 10.1139/cjfr-2013-033
Buechling, A., Martin, P. H., and Canham, C. D. (2017). Climate and competition effects on tree growth in Rocky Mountain forests. J. Ecol. 105, 1636–1647. doi: 10.1111/1365-2745.12782
Campioli, M., Vicca, S., Luyssaert, S., Bilcke, J., Ceschia, E., Chapin Iii, F. S., et al. (2015). Biomass production efficiency controlled by management in temperate and boreal ecosystems. Nat. Geosci. 8, 843–846. doi: 10.1038/ngeo2553
Christin, S., Hervet, É., and Lecomte, N. (2018). Applications for deep learning in ecology. bioRxiv 10, 1632–1644. doi: 10.1111/2041-210X.13256
Collalti, A., Perugini, L., Santini, M., Chiti, T., Nolè, A., Matteucci, G., et al. (2014). A process-based model to simulate growth in forests with complex structure: Evaluation and use of 3D-CMCC Forest Ecosystem Model in a deciduous forest in Central Italy. Ecol. Model. 272, 362–378. doi: 10.1016/j.ecolmodel.2013.09.016
Collalti, A., Thornton, P. E., Cescatti, A., Rita, A., Borghetti, M., Nolè, A., et al. (2019). The sensitivity of the forest carbon budget shifts across processes along with stand development and climate change. Ecol. Appl. 29, e01837. doi: 10.1002/eap.1837
Crockatt, M. E. and Bebber, D. P. (2015). Edge effects on moisture reduce wood decomposition rate in a temperate forest. Global Change Biol. 21, 598–707. doi: 10.1111/gcb.12676
Cui, R., Qi, S., Wu, B., Zhang, D., Zhang, L., Zhou, P., et al. (2022). The influence of stand structure on understory herbaceous plants species diversity of platycladus orientalis plantations in Beijing, China. Forests 13, 1921. doi: 10.3390/f13111921
de Wergifosse, L., André, F., Goosse, H., Boczon, A., Cecchini, S., Ciceu, A., et al. (2022). Simulating tree growth response to climate change in structurally diverse oak and beech forests. Sci. Total Environ. 806, 150422. doi: 10.1016/j.scitotenv.2021.150422
Du, Z., Liu, X., Wu, Z., Zhang, H., and Zhao, J. (2022). Responses of forest net primary productivity to climatic factors in China during 1982–2015. Plants 11, 2932. doi: 10.3390/plants11212932
Durán Zuazo, V. H. and Rodríguez Pleguezuelo, C. R. (2008). Soil-erosion and runoff prevention by plant covers. A review. Agron. Sustain. Dev. 28, 65–86. doi: 10.1051/agro:2007062
Ehbrecht, M., Seidel, D., Annighöfer, P., Kreft, H., Köhler, M., Zemp, D. C., et al. (2021). Global patterns and climatic controls of forest structural complexity. Nat. Commun. 12, 519. doi: 10.1038/s41467-020-20767-z
Forrester, D. I., Kohnle, U., Albrecht, A. T., and Bauhus, J. (2013). Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manage. 304, 233–242. doi: 10.1016/j.foreco.2013.04.038
Graham, M. H. (2003). Confronting multicollinearity in ecological multiple regression. Ecology 84, 2809–2815. doi: 10.1890/02-3114
Grömping, U. (2009). Variable importance assessment in regression: linear regression versus random forest. Am. Statistician 63, 308–319. doi: 10.1198/tast.2009.08199
Hair, J. F., Hult, G. T. M., Ringle, C. M., Sarstedt, M., Danks, N. P., and Ray, S. (2021). “An introduction to structural equation modeling,” in Partial Least Squares Structural Equation Modeling (PLS-SEM) Using R: A Workbook. Eds. Hair, J., Hult, G. T. M., Ringle, C. M., Sarstedt, M., Danks, N. P., and Ray, S. (Springer International Publishing, Cham), 1–29. doi: 10.1007/978-3-030-80519-7
Hatami, N., Lohmander, P., Moayeri, M. H., and Limaei, S. M. (2020). A basal area increment model for individual trees in mixed continuous cover forests in Iranian Caspian forests. J. Forestry Res. 31, 99–106. doi: 10.1007/s11676-018-0862-8
He, J.-S., Dong, S., Shang, Z., Sundqvist, M. K., Wu, G., and Yang, Y. (2021). Above-belowground interactions in alpine ecosystems on the roof of the world. Plant Soil 458, 1–6. doi: 10.1007/s11104-020-04761-4
He, F., Mohamadzadeh, N., Sadeghnejad, M., Ingram, B., and Ostovari, Y. (2023). Fractal features of soil particles as an index of land degradation under different land-use patterns and slope-aspects. Land 12, 615. doi: 10.3390/land12030615
Jafarian, N., Mirzaei, J., Omidipour, R., and Kooch, Y. (2023). Effects of micro-climatic conditions on soil properties along a climate gradient in oak forests, west of Iran: Emphasizing phosphatase and urease enzyme activity. CATENA 224, 106960. doi: 10.1016/j.catena.2023.106960
Jandl, R., Lindner, M., Vesterdal, L., Bauwens, B., Baritz, R., Hagedorn, F., et al. (2007). How strongly can forest management influence soil carbon sequestration? Geoderma 137, 253–268. doi: 10.1016/j.geoderma.2006.09.003
Jandl, R., Spathelf, P., Bolte, A., and Prescott, C. E. (2019). Forest adaptation to climate change—is non-management an option? Ann. For. Sci. 76, 48. doi: 10.1007/s13595-019-0827-x
Jevšenak, J. and Skudnik, M. (2021). A random forest model for basal area increment predictions from national forest inventory data. For. Ecol. Manage. 479, 118601. doi: 10.1016/j.foreco.2020.118601
Jian, Z., Ni, Y., Lei, L., Xu, J., Xiao, W., and Zeng, L. (2022). Phosphorus is the key soil indicator controlling productivity in planted Masson pine forests across subtropical China. Sci. Total Environ. 822, 153525. doi: 10.1016/j.scitotenv.2022.153525
Jiang, H., Apps, M. J., Peng, C., Zhang, Y., and Liu, J. (2002). Modelling the influence of harvesting on Chinese boreal forest carbon dynamics. For. Ecol. Manage. 169, 65–82. doi: 10.1016/S0378-1127(02)00299-2
Jiang, X., Huang, J.-G., Cheng, J., Dawson, A., Stadt, K. J., Comeau, P. G., et al. (2018). Interspecific variation in growth responses to tree size, competition and climate of western Canadian boreal mixed forests. Sci. Total Environ. 631-632, 1070–1078. doi: 10.1016/j.scitotenv.2018.03.099
Jing, X., Muys, B., Baeten, L., Bruelheide, H., De Wandeler, H., Desie, E., et al. (2022). Climatic conditions, not above- and belowground resource availability and uptake capacity, mediate tree diversity effects on productivity and stability. Sci. Total Environ. 812, 152560. doi: 10.1016/j.scitotenv.2021.152560
Kauppi, P. E., Stål, G., Arnesson-Ceder, L., Hallberg, I., Sramek, Hoen, H. F., Svensson, A., et al. (2022). Managing existing forests can mitigate climate change. For. Ecol. Manage. 513, 120186. doi: 10.1016/j.foreco.2022.120186
Kira, T. and Shidei, T. (1967). Primary production and turnover of organic matter in different forest ecosystems of the western pacific. Japanese J. Ecol. 17, 70–87. doi: 10.18960/seitai.17.2_70
Lafond, V., Lagarrigues, G., Cordonnier, T., and Courbaud, B. (2014). Uneven-aged management options to promote forest resilience for climate change adaptation: effects of group selection and harvesting intensity. Ann. For. Sci. 71, 173–186. doi: 10.1007/s13595-013-0291-y
Lan, J., Lei, X., He, X., Gao, W.-Q., and Guo, H. (2023). Stand density, climate and biodiversity jointly regulate the multifunctionality of natural forest ecosystems in northeast China. Eur. J. For. Res. 142, 493–507. doi: 10.1007/s10342-023-01537-0
LaRue, E. A., Knott, J. A., Domke, G. M., Chen, H. Y. H., Guo, Q., Hisano, M., et al. (2023). Structural diversity as a reliable and novel predictor for ecosystem productivity. Front. Ecol. Environ. 21, 33–39 doi: 10.1002/fee.2586
Lin, S., Li, Y., Chen, M., Li, Y., Wang, L., and He, K. (2021). Effects of local neighbourhood structure on radial growth of Picea crassifolia Kom. and Betula platyphylla Suk. plantations in the loess alpine region, China. For. Ecol. Manage. 491, 119195. doi: 10.1016/j.foreco.2021.119195
Lin, N., Wu, B., Jansen, R., Gerstein, M., and Zhao, H. (2004). Information assessment on predicting protein-protein interactions. BMC Bioinf. 5, 154. doi: 10.1186/1471-2105-5-154
Liu, X., Trogisch, S., He, J.-S., Niklaus, P. A., Bruelheide, H., Tang, Z., et al. (2018). Tree species richness increases ecosystem carbon storage in subtropical forests. Proc. R. Soc. B 285, 20181240. doi: 10.1098/rspb.2018.1240
Liu, Y., Yu, G., Wang, Q., and Zhang, Y.-J. (2014). How temperature, precipitation and stand age control the biomass carbon density of global mature forests. Global Ecol. Biogeography 23, 323–333. doi: 10.1111/geb.12113
Liu, D., Zhou, C. Z., He, X., Zhang, X., Feng, L., and Zhang, H. (2022). The effect of stand density, biodiversity, and spatial structure on stand basal area increment in natural spruce-fir-broadleaf mixed forests. Forests. 13, 162 doi: 10.3390/f13020162
Maréchaux, I., Langerwisch, F., Huth, A., Bugmann, H., Morin, X., Reyer, C. P. O., et al. (2021). Tackling unresolved questions in forest ecology: The past and future role of simulation models. Ecol. Evol. 11, 3746–3770. doi: 10.1002/ece3.7391
Milovac, J., Ingwersen, J., and Warrach-Sagi, K. (2018). Global top soil texture data compatible with the WRF model based on the Harmonized World Soil Database (HWSD) at 30 arc-second horizontal resolution Version 1.21 (World Data Center for Climate (WDCC) at DKRZ).
Morin, X. (2015). Species richness promotes canopy packing: a promising step towards a better understanding of the mechanisms driving the diversity effects on forest functioning. Funct. Ecol. 29, 993–994. doi: 10.1111/1365-2435.12473
Ni, Y., Jian, Z., Zeng, L., Liu, J., Lei, L., Zhu, J., et al. (2022). Climate, soil nutrients, and stand characteristics jointly determine large-scale patterns of biomass growth rates and allocation in Pinus massoniana plantations. For. Ecol. Manage. 504, 119839. doi: 10.1016/j.foreco.2021.119839
Özçelik, R., Diamantopoulou, M. J., Crecente-Campo, F., and Eler, U. (2013). Estimating Crimean juniper tree height using nonlinear regression and artificial neural network models. For. Ecol. Manage. 306, 52–60. doi: 10.1016/j.foreco.2013.06.009
Pardos, M., del Río, M., Pretzsch, H., Jactel, H., Bielak, K., Bravo, F., et al. (2021). The greater resilience of mixed forests to drought mainly depends on their composition: Analysis along a climate gradient across Europe. For. Ecol. Manage. 481, 118687. doi: 10.1016/j.foreco.2020.118687
Post, W. M. and Kwon, K. C. (2000). Soil carbon sequestration and land-use change: processes and potential. Global Change Biol. 6, 317–327. doi: 10.1080/02827581.2016.1220615
Schwarz, M. T., Bischoff, S., Blaser, S., Boch, S., Schmitt, B., Thieme, L., et al. (2014). More efficient aboveground nitrogen use in more diverse Central European forest canopies. For. Ecol. Manage. 313, 274–282. doi: 10.1016/j.foreco.2013.11.021
Selkimäki, M., González-Olabarria, J. R., and Pukkala, T. (2012). Site and stand characteristics related to surface erosion occurrence in forests of Catalonia (Spain). Eur. J. For. Res. 131, 727–738. doi: 10.1007/s10342-011-0545-x
Shiri, N., Shiri, J., Kazemi, M. H., and Xu, T. (2022). Estimation of CO2 flux components over northern hemisphere forest ecosystems by using random forest method through temporal and spatial data scanning procedures. Environ. Sci. Pollut. Res. 29, 16123–16137. doi: 10.1007/s11356-021-16501-x
Shuai, O., Xiang, W., Wang, X., Xiao, W., Chen, L., Li, S., et al. (2019). Effects of stand age, richness and density on productivity in subtropical forests in China. J. Ecol. 107, 2266–2277. doi: 10.1111/1365-2745.13194
Sun, J., Jiao, W., Wang, Q., Wang, T., Yang, H., Jin, J., et al. (2021). Potential habitat and productivity loss of Populus deltoides industrial forest plantations due to global warming. For. Ecol. Manage. 496, 119474. doi: 10.1016/j.foreco.2021.119474
Viet, H. D. X., Tymińska-Czabańska, L., and Socha, J. (2023). Modeling the effect of stand characteristics on oak volume increment in Poland using generalized additive models. Forests 14, 123. doi: 10.3390/f14010123
Wang, H.-m., Saigusa, N., Zu, Y.-G., Wang, W.-J., Yamamoto, S., and Kondo, H. (2008). Carbon fluxes and their response to environmental variables in a Dahurian larch forest ecosystem in northeast China. J. Forestry Res. 19, 1–10. doi: 10.1007/s11676-008-0001-z
Wang, Z., Zhang, X., Chhin, S., Zhang, J., and Duan, A. (2021). Disentangling the effects of stand and climatic variables on Net Ecosystem Carbon Change (NECC) of Chinese fir plantations in subtropical China using a random forest algorithm. Agric. For. Meteorology 304-305, 108412. doi: 10.1016/j.agrformet.2021.108412
Wang, Z., Zhang, X., Zhang, J., and Chhin, S. (2022). Effects of stand factors on tree growth of Chinese fir in the subtropics of China depends on climate conditions from predictions of a deep learning algorithm: A long-term spacing trial. For. Ecol. Manage. 520, 120363. doi: 10.1016/j.foreco.2022.120363
Wu, C., Chen, Y., Peng, C., Li, Z., and Hong, X. (2019). Modeling and estimating aboveground biomass of Dacrydium pierrei in China using machine learning with climate change. J. Environ. Manage. 234, 167–179. doi: 10.1016/j.jenvman.2018.12.090
Wu, Z., Fan, C., Zhang, C., Zhao, X., and von Gadow, K. (2022). Effects of biotic and abiotic drivers on the growth rates of individual trees in temperate natural forests. For. Ecol. Manage. 503, 119769. doi: 10.1016/j.foreco.2021.119769
Wu, B., Zhou, L., Qi, S., Jin, M., Hu, J., and Lu, J. (2021). Effect of habitat factors on the understory plant diversity of Platycladus orientalis plantations in Beijing mountainous areas based on MaxEnt model. Ecol. Indic. 129, 107917. doi: 10.1016/j.ecolind.2021.107917
Xi, X. and Yuan, X. (2022). Significant water stress on gross primary productivity during flash droughts with hot conditions. Agric. For. Meteorology 324, 109100. doi: 10.1016/j.agrformet.2022.109100
Xue, R., Jiao, L., Zhang, P., Wang, X., Li, Q., Yuan, X., et al. (2025). Climatic habitat regulates the radial growth sensitivity of two conifers in response to climate change. For. Ecosyst. 12, 100282. doi: 10.1016/j.fecs.2024.100282
Yuan, Z., Ali, A., Jucker, T., Ruiz-Benito, P., Wang, S., Jiang, L., et al. (2019). Multiple abiotic and biotic pathways shape biomass demographic processes in temperate forests. Ecology 100, e02650. doi: 10.1002/ecy.2650
Zampieri, M., Grizzetti, B., Toreti, A., de Palma, P., and Collalti, A. (2021). Rise and fall of vegetation primary production resilience to climate variability anticipated by a large ensemble of Earth System Models’ simulations. Environ. Res. Lett. 16, 105001. doi: 10.1088/1748-9326/ac2407
Zhang, Y. and Chen, H. Y. H. (2015). Individual size inequality links forest diversity and above-ground biomass. J. Ecol. 103, 1245–1252. doi: 10.1111/1365-2745.12425
Zhang, L., Qi, S., Li, P., and Zhou, P. (2024a). Influence of stand and environmental factors on Net Ecosystem Carbon Change (NECC) of Platycladus orientalis plantations in Beijing’s mountainous areas. Ecol. Indic. 158, 111385. doi: 10.1016/j.ecolind.2023.111385
Zhang, X., Wang, Z., Chhin, S., Wang, H., Duan, A., and Zhang, J. (2020). Relative contributions of competition, stand structure, age, and climate factors to tree mortality of Chinese fir plantations: Long-term spacing trials in southern China. For. Ecol. Manage. 465, 118103. doi: 10.1016/j.foreco.2020.118103
Zhang, Z., Zhou, L., Lu, C., Fu, Y., Gu, Z., Chen, Y., et al. (2024b). Drought- induced decrease in tree productivity mainly mediated by the maximum growth rate and growing-season length in a subtropical forest. For. Ecol. Manage. 563, 121985. doi: 10.1016/j.foreco.2024.121985
Keywords: net ecosystem carbon change, environmental factors, stand age, natural forests, the Da Xing’an Mountains
Citation: Wang Z, Hao Y, Dong L, Miao Z, Jin X, Zhao X and Cheng S (2025) Factors influencing net ecosystem carbon change in cold-temperate coniferous forests of the Da Xing’an Mountains: analysis across developmental stages based on stand, structural, and environmental factors. Front. Plant Sci. 16:1663271. doi: 10.3389/fpls.2025.1663271
Received: 10 July 2025; Accepted: 17 October 2025;
Published: 24 November 2025.
Edited by:
Maryam Teimouri, Research Institute of Forests and Rangelands, IranReviewed by:
Chao Huang, Jiangxi Agricultural University, ChinaYunhong Xie, Beijing Forestry University, China
Copyright © 2025 Wang, Hao, Dong, Miao, Jin, Zhao and Cheng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuanshuo Hao, aGFveXVhbnNodW9AbmVmdS5lZHUuY24=; Lihu Dong, bGlodWRvbmdAbmVmdS5lZHUuY24=