 Panpan Jing
Panpan Jing Dan Wang
Dan Wang Chunwu Zhu2
Chunwu Zhu2 Jiquan Chen
Jiquan Chen- 1International Center for Ecology, Meteorology, and Environment, School of Applied Meteorology, Nanjing University of Information Science and Technology, Nanjing, China
- 2State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing, China
- 3CGCEO/Geography, Michigan State University, East Lansing, MI, USA
Land surface temperature over the past decades has shown a faster warming trend during the night than during the day. Extremely low night temperatures have occurred frequently due to the influence of land-sea thermal difference, topography and climate change. This asymmetric night temperature change is expected to affect plant ecophysiology and growth, as the plant carbon consumption processes could be affected more than the assimilation processes because photosynthesis in most plants occurs during the daytime whereas plant respiration occurs throughout the day. The effects of high night temperature (HNT) and low night temperature (LNT) on plant ecophysiological and growing processes and how the effects vary among different plant functional types (PFTs) have not been analyzed extensively. In this meta-analysis, we examined the effect of HNT and LNT on plant physiology and growth across different PFTs and experimental settings. Plant species were grouped according to their photosynthetic pathways (C3, C4, and CAM), growth forms (herbaceous, woody), and economic purposes (crop, non-crop). We found that HNT and LNT both had a negative effect on plant yield, but the effect of HNT on plant yield was primarily related to a reduction in biomass allocation to reproduction organs and the effect of LNT on plant yield was more related to a negative effect on total biomass. Leaf growth was stimulated at HNT and suppressed at LNT. HNT accelerated plants ecophysiological processes, including photosynthesis and dark respiration, while LNT slowed these processes. Overall, the results showed that the effects of night temperature on plant physiology and growth varied between HNT and LNT, among the response variables and PFTs, and depended on the magnitude of temperature change and experimental design. These findings suggest complexities and challenges in seeking general patterns of terrestrial plant growth in HNT and LNT. The PFT specific responses of plants are critical for obtaining credible predictions of the changes in crop production, plant community structure, vegetation dynamics, biodiversity, and ecosystem functioning of terrestrial biomes when asymmetric night temperature change continues.
Introduction
The increased intensity of human activities has been magnifying the climate change and its consequences in recent decades (IPCC, 2013). A remarkable feature of climate change is global warming, caused by anthropogenic emissions of key greenhouse gasses that absorb infrared radiation, such as CO2, CH4, and N2O, deforestation and urbanization. The global temperature is forecasted to continuously increase 1–3.7°C by the end of the 21st century (IPCC, 2013). Compared with day temperature, night temperature has increased faster at local (Peng et al., 2004), country (Zhou et al., 2004; Rao et al., 2014), and global scales (Vose et al., 2005). On average, the lowest land nighttime temperature increased about 0.2°C per decade between 1950 and 1993, which is double the increased highest daytime temperature (IPCC, 2001). It is probably due to the incremental cloudiness, which leads to less radiant heat loss (Alward et al., 1999). Night temperature increased 1.13°C in the Philippines from 1979 to 2003 (Peng et al., 2004), whereas night temperature in Lybia over a period of 45 years (1950–1995) increased at a rate of 0.18°C per decades (Jones et al., 1999). Based on the prediction of multi-model ensembles, asymmetric warming between day and night is going to continue in the future (Christensen et al., 2007; Sillmann et al., 2013). Therefore, plants in the future will be exposed to warmer nights, which could greatly influence crop yield and vegetation dynamics as well as ecosystem biodiversity, structure and productivity.
Due to the influence of land-sea thermal differences, topography and climate change, extremely low temperatures have also occurred frequently around the world (Yang et al., 2006). Low temperature is one of the major environmental factors impacting plant growth, development and ecological distribution (Allen and Ort, 2001). A variety of crops from tropical and sub-tropical regions, such as maize, tomato, cucumber, and mango, are sensitive to cold when cultivated in temperate environments (Jones and Ort, 1998; Allen and Ort, 2001; Meng et al., 2008). As people have begun introducing plants from warm climates into cool climates, it has become important to understand the effects of LNT stress, which needs substantially more research.
Studies on plant response mechanisms to warming or chilling temperatures serve a great purpose in understanding agriculture and natural ecosystems. Increased research efforts have used manipulated field experiments across the world to investigate the potential impacts of climate warming on terrestrial plants and ecosystems (Rustad, 2008). However, the majority of these previous studies have focused on the increase of daily or monthly mean temperature, assuming no difference in the impact of day versus night temperature (Peng et al., 2004). Rustad et al. (2001) conducted a meta-analysis of experimental data from ecosystem warming studies and found that elevated temperatures significantly increased above ground productivity by 19%. In fact, the effects of night temperature are different from that of day temperature (Xia et al., 2014) and produced a relatively greater challenge in estimating global change impact on crop yield and ecosystem functions (Jagadish et al., 2015). Previous studies on night temperatures have focused either on the effects of HNT and LNT alone (Friend, 1981; Seddigh and Jolliff, 1984a,b,c; Koscielniak, 1993; Bertamini et al., 2005) or the mixed effects of night temperatures and CO2 concentration (Mortensen and Moe, 1992; Volder et al., 2004; Cheng et al., 2008, 2009, 2010), light period (Gimenez and Rumi, 1988; Turner and Ewing, 1988; Lee et al., 1991; Verheul et al., 2007), intensity (Bunce, 1985; Mortensen, 1994; Rapacz, 1998; Flexas and Osmond, 1999; Davies et al., 2002) as well as other environmental factors (Schoppach and Sadok, 2013) and growth regulators (Shah et al., 2011; Mohammed et al., 2013; Zhang et al., 2014). These experiments had been conducted on pineapple (Neales et al., 1980), peanut (Bagnall et al., 1988; Wang, 2007; Lin et al., 2011) and shrub-grass ecosystems (Beier et al., 2004). Although the interest in the influence of night temperatures on many aspects of plants is growing, studies are scattered and there lacks a synthetic study on how and to what extent night temperature change impacts terrestrial plant growth and biomass accumulation. To accurately predict the effects of climatic change and develop sound adaptive agricultural systems and land management practices, it is imperative to understand how night temperature affects photosynthetic carbon gain, loss and allocation through a comprehensive analysis of HNT and LNT studies.
Night temperature has both direct and indirect effects on plant physiology, morphology, growth and yield. HNT and LNT impact plant physiology in many aspects, of which photosynthesis is the most severely affected process (Berry and Bjorkman, 1980; Damian and Donald, 2001; Yu et al., 2002; Liu et al., 2010, 2011). There was a consistent suppression on Anet (net CO2 assimilation rate) at LNT for both C3 (Flexas and Osmond, 1999; Bange and Milroy, 2004; Zhang et al., 2010; Sao et al., 2013b) and C4 species (Sao et al., 2013a), but a stimulation for CAM species (Chen et al., 2008; Pollet et al., 2011). HNT had a positive (Seddigh and Jolliff, 1984c; Prieto et al., 2009; Darnell et al., 2013), negative (Teragishi et al., 2001; Mohammed et al., 2013; Narayanan et al., 2015; Peraudeau et al., 2015), or no effect (Veatch et al., 2007; Ibrahim et al., 2010; Cheesman and Klaus, 2013) on Anet for C3 species and a negative (Prasad and Djanaguiraman, 2011) effect for C4 species. The effect of HNT and LNT on photosynthesis was related to leaf chlorophyll content (Prasad and Djanaguiraman, 2011), fluorescence parameters including photochemical efficiency of PSII (Fv/Fm), PSII quantum yield (ΦPSII) and ETR (Liu et al., 2011, 2012; Zhang et al., 2014), nitrogen (N) concentration (Mohammed and Tarpley, 2009a), gs (stomatal conductance) (Farquhar and Sharkey, 1982) and enzyme activities related to carbon fixation (Noctor and Foyer, 1998). Among different PFTs, a positive correlation between HNT and plant height was reported (Patterson, 1990; Papadopoulos and Hao, 2000; Cheng et al., 2009; Lucidos et al., 2013). However, LNT had a negative effect on plant height for C3 (Zieslin et al., 1986; Pressman et al., 2006; Kjær et al., 2008) and C4 species (Uehara et al., 2009), but a positive effect for CAM species (Serra and Carrai, 1988). The responses of biomass accumulation to different night temperature conditions were not identical among different species. HNT had a positive effect on stem dry weight for woody plants (Malek et al., 1992; Cheesman and Klaus, 2013), a positive (Cheng et al., 2008, 2009; Darnell et al., 2013) or negative (Seddigh and Jolliff, 1984a; Lee and Myeongwhan, 2011) effect for herbaceous plants. However, LNT had a positive (Lepage et al., 1984), negative (Kjær et al., 2008; Uehara et al., 2009; Rehmani et al., 2014) or no effect (Dejong and Smeets, 1982) for herbaceous plants. Both HNT and LNT resulted in a reduction in crop yield, such as rice (Ziska and Manalo, 1996; Kanno and Makino, 2010; Mohammed and Tarpley, 2010; Shi et al., 2013), winter wheat (Zhang Y. H. et al., 2013; Narayanan et al., 2015), sorghum (Prasad and Djanaguiraman, 2011) and tomato (Khayat et al., 1985; Zhang et al., 2010; Qi et al., 2011; Zhang Y. et al., 2013). Clearly, lessons from previous studies are not all consistent and sometimes, contradictory. It is essential to conduct a comprehensive review on the responses of different plant functional groups to different night temperatures.
In addition to species functional groups and night temperature treatments, experimental design (e.g., treatment duration and growth facility) may also matter in understanding plant responses to night temperature change. A significant reduction in rice yield at HNT was associated with the reduction of N and non-structural content translocation after flowering in a field experiment (Shi et al., 2013). In a pot-growing experiment, yield loss was concerned with decreased dry matter allocation to grain due to reduced spikelet fertility during the reproductive stage (Cheng et al., 2009). The variation of Anet deduction due to HNT and LNT was dependent on experimental durations (Teragishi et al., 2001; Zhu et al., 2005; Ibrahim et al., 2010; Sao et al., 2010; Prasad and Djanaguiraman, 2011). However, the difference between responses to different treatment durations or to growing facilities is ambiguous. Confirming the effects of experimental methodology is of key theoretical and practical significance to help agriculture to choose the right cultivation practice to mitigate adverse effects caused by HNT or LNT.
A meta-analysis of plant responses to temperature indicated that CO2 elevation affected plant ecophysiology and growth, with different magnitudes at different temperature treatments (Wang et al., 2012). Not only daily temperature, but also the magnitude of night temperature variation caused different impacts. Elevated night temperature by 5°C had a negative effect on Anet and gs but no effect on intercellular CO2 concentration (Ci) (Mohammed et al., 2013), while elevated night temperature by 8°C significantly increased Anet, gs, and Ci of bell peppers (Darnell et al., 2013). With more night temperature reduction, the decrease in total dry weight, number of leaves and leaf area for goatsrue were increased gradually (Patterson, 1993). Although the effects of different magnitude of night temperature variation on plant physiology and growth varied significantly, the comparisons between these effects are missing and a quantitative review would reveal the optimal night temperature for different ecophysiological processes and growth.
The primary objective of this study was to investigate the effects of high and LNTs on various aspects of plant responses, including physiological, morphological, and growth characteristics. Specifically, our objectives were to: (1) assess the difference and magnitude of HNT and LNT effects on plant physiology, morphology and yield-related responses. The physiological characters included Anet, PSII function, gs, dark respiration (Rd), maximum carboxylation rate (Vcmax), maximum ETR (Jmax), tissue N and TNC. The morphology responses included plant height and leaf characteristics (number of leaves, LAI, SLA and LAR). Yield-related parameters included dry biomass, number of reproductive organs and yield; (2) detect differences among different PFTs based on photosynthetic pathways, growth forms and economic value; (3) investigate the effect of the magnitude of the night temperature changes on different responses; (4) tease apart the effect of growth facilities or treatment durations on affecting these responses. Accordingly, the specific hypotheses were proposed as: (1) HNT and LNT would have similar negative effects on plant physiological performance and growth; (2) LNT would have a stronger negative effect on C4 species than on C3 species; HNT would have a stronger negative effect on C3 than on C4 species. To test these hypotheses, we conducted a comprehensive meta-analysis of night-temperature manipulated studies published from 1980 through 2015, using the response ratio lnr as an estimate of the effect size of night-temperature relative to control plots.
Materials and Methods
Data Selection
Peer-reviewed journal publications were searched with the key word “night temperature” on the Web of Science to build a comprehensive database. The list of papers were then cross-checked with a list of references cited in review articles that were relevant to night temperature effects in order to assure that all articles available for this meta-analysis were included. Any article published in English from 1980 to 2015 that met the following two criteria were included: (1) plants were treated at ANT as a control group, and HNT or LNT as treatment groups; (2) measurements on physiology, morphology, and yield were carried out on both control and treatment groups. The following two criteria were applied to exclude studies: (1) day and night temperatures were treated at the same time; (2) studies focused on extreme temperature values, which resulted in the death of plants. Eventually, 112 papers were selected in this study (Supplementary Material S1). Data were extracted directly from the tables in the articles or were obtained by using the software GetData Graph Digitizer when presented in graphical formats. In these studies, night temperature was 1–20°C above or below ANT, with only four studies more than 20°C above or below ANT (Supplementary Figure S2). Response variables extracted from these articles contained physiological characters including net photosynthesis (Anet), PSII efficiency (Fv/Fm), stomatal conductance (gs), dark respiration (Rd), non-structural carbohydrate content (TNC), tissue nitrogen (N) content (i.e., stem, leaf, panicle, spike, grain, shoot, root, and total N) and tissue carbon (C) content (i.e., stem, leaf, shoot, root, and total C), morphological features (i.e., plant height, stem diameter, internode length, number of leaves, SLA, LAI, LAR) and yield-related parameters (i.e., dry weight, number of reproductive organs, days to flowering and yield). For multi-year studies on annual species, results from different seasons were considered independent and all observations were included in this analysis. To ensure the independent nature of the data, we excluded duplicate results found in different publications. However, our analyses were not completely independent because individual papers often provided data with more than one treatment (e.g., different HNT or LNT magnitudes) and/or different response variables. To examine the influence of non-independence of data, we first averaged those data from the same published study by PFTs so that only one comparison was used from a published study for each PFT. Nonetheless, we found that most of the response patterns were unchanged; therefore, all data were used in our study.
Categorization of the Studies
Night temperature was categorized into three levels: ANT, HNT, and LNT. In addition to the response variables and night temperature categories described above, plant species, sample sizes, growth facilities and treatment durations under each temperature treatment were also collected. Following the categorization of Wang et al. (2012), plant species were classified based on photosynthetic pathways (C3, C4, or CAM), growth forms (herbaceous or woody) and economic values (crop or non-crop). We listed the species, PFTs and associated references used in this study (Supplementary Table S3). The experiments conducted in these studies were either indoors (growth chamber and greenhouse) or field studies. Due to relatively less data in the field studies, growth facilities used in these experiments were categorized as two levels of pot size: <10L and >10L. Because the treatment duration varied from hours to years, we grouped then into two levels: short-term (hours–days) and long-term (months–years).
Meta-Analysis Methods
We employed a similar method from Hedges et al. (1999). To avoid the adverse effects of different units, we used the response ratio r = Xt/Xc to estimate the effect size of night temperature treatments, where Xt is the treatment mean and Xc is the control mean. In order to compare expediently, we calculated the natural logarithm of the response ratio (lnr). In addition to the mean value, standard deviation (SD) and sample size (n) for each individual observation were also collected to calculate the variance of effect size. Using METAWIN software 2.1 (Sinauer Associates, Inc. Sunderland, MA, USA), we calculated the effect size of the targeted variables and used a weighted, fixed-effects model to evaluate the categorical effects on night temperature treatments, plant species, pot sizes and treatment durations. If the 95% confidence interval (CI) of lnr generated by the fixed-effects model overlapped 0, the temperature treatment was considered to have no significant impact on the response variables. If the upper bound of the 95% CI was smaller than 0, the response was considered significantly negative. Conversely, it indicated that the treatment had a significantly positive effect on variables if the lower bound of the 95% CI was greater than 0. Although total difference among groups was divided into within-group and between group difference, the significance level of the latter revealed whether the response was different among groups (Hedges et al., 1999). The response of plants was considered significantly disparate between HNT and LNT overall or for different species, pot size or treatment duration if their 95% CIs did not overlap. Significance was established at p < 0.05 unless otherwise noted.
Publication bias of the effect size (lnr) in this meta-analysis was determined with METAWIN software 2.1 (Sinauer Associates, Inc. Sunderland, MA, USA). We calculated Spearman’s rank-order correlation (rs) which indicates the relationship between the effect size (lnr) and the sample size (Begg and Mazumdar, 1994), and Rosenthal’s fail-safe number which represents the number of additional studies with a mean effect size of zero needed to eliminate the significance of a significant effect (Rosenthal, 1979). Publication bias was significant if p-value of rs was smaller than 0.05. However, the publication bias may be safely ignored if the fail-safe number is larger than a critical value of 5n+10 where n is the number of studies (Rosenberg, 2005).
Statistical Analysis
Original data collected from these studies were arranged into a database in which the value of response variables was lnr. The relationship between lnr of all the variables and the magnitude of night temperature treatments were evaluated by a second-degree polynomial or linear regression analysis with the R statistical programming language (R 3.2.2 for Windows GUI front-end).
Results
Significance of HNT and LNT
Across all of the studies, HNT increased Anet, gs, Rd, and tissue N content on average by 2.56, 11.37, 27.02, and 26.87%, respectively, decreased Fv/Fm, chlorophyll content, starch, sucrose and TNC content by 0.98, 8.08, 22.26, 13.77, and 13.97%, but unaffected Tr (transpiration rate), Ci, PSII quantum yield, ETR and tissue C content (Figure 1A). LNT had negative effects on most physiological response variables by different magnitudes, but increased chlorophyll (4.81%), C (1.11%), starch (5.73%), sucrose (4.71%) and TNC content (3.32%). HNT decreased stem diameter and internode length by 1.61%, and 15.97%, which were unchanged by LNT (Figure 1B). HNT and LNT had an opposite effect on plant height, number of leaves, SLA, LAI, and LAR (Figure 1B). HNT had positive effects on total dry weight and number of productive tillers, negative effects on leaf, stem, and fruit dry weight, number of reproductive organs, flowering time and yield, and no effects on above-ground, below-ground dry weight and fruit size (Figure 1C). LNT decreased leaf (13.69%), fruit (15.18%), above-ground (6.7%), below-ground (23.8%), and total dry weight (55.33%), reproductive organs number (6.82%) and yield (37.66%), respectively, but had no effects on stem dry weight, number of productive tillers, anthesis and fruit size (Figure 1C). Among all the variables, there was publication bias for chlorophyll content (rs = -0.399, p = 0.02), leaf (rs = 0.346, p = 0.002), stem (rs = 0.339, p = 0.0006), above-ground (rs = 0.235, p = 0.006), and below-ground dry weight (rs = 0.22, p = 0.07), which could not be ignored based on Rosenthal’s value.
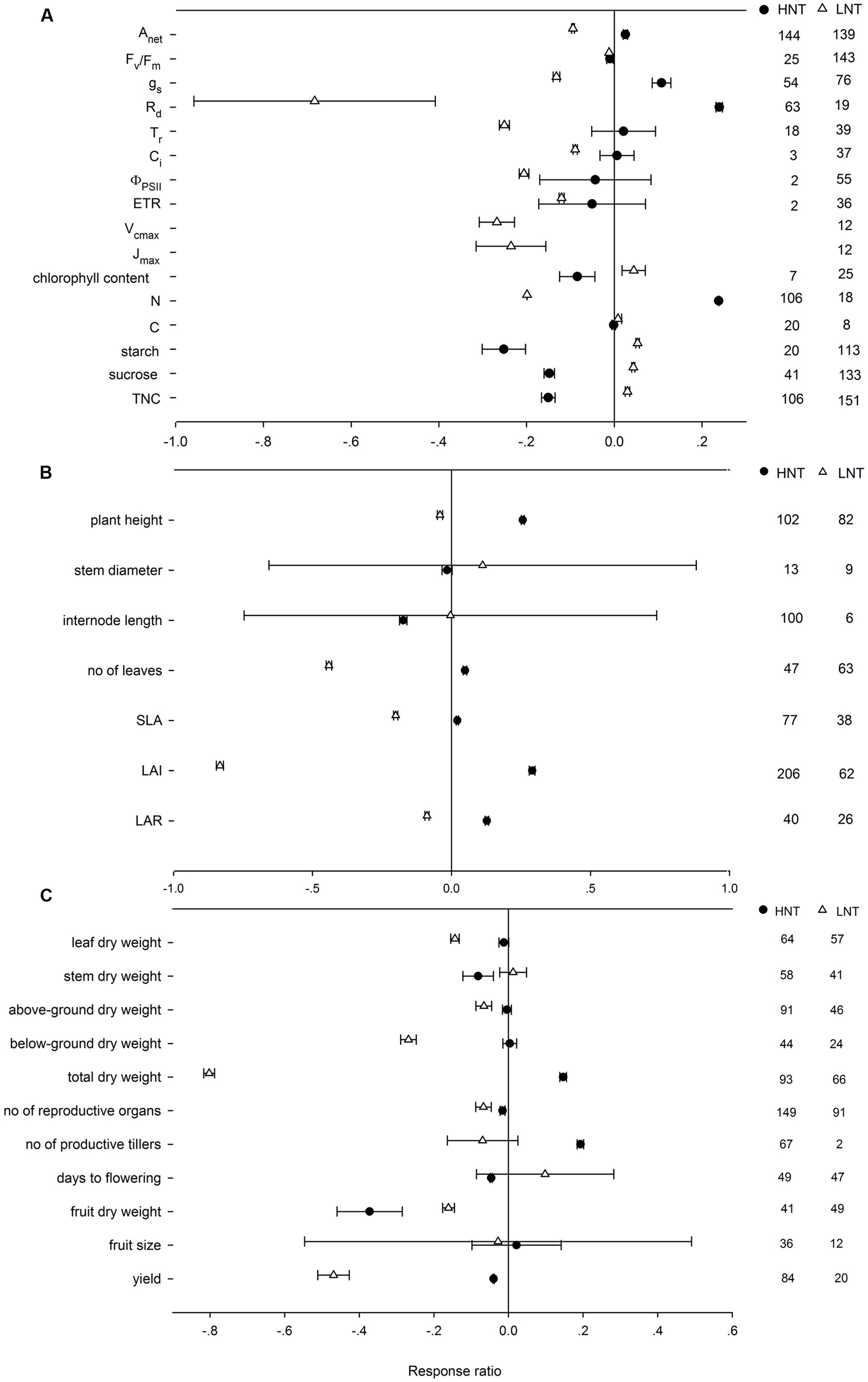
FIGURE 1. Plant (A) physiological, (B) morphological, and (C) yield-related responses to HNT (filled circles) and LNT (open triangle). Each data point represents the mean ± 95% confidence interval (CI). The number of observations for each variable is given on the right of the graph. Note that N is tissue nitrogen content including stem, leaf, panicle, spike, grain, shoot, root, and total N. C is tissue carbon content including stem, leaf, shoot, root, and total C.
Variable Responses among Plant Functional Types (PFTs)
HNT stimulated Anet by 3.43% for C3 species, but suppressed it by 35.57% for CAM species (Figure 2). Note that there were not enough publications for a summary on C4 species. LNT suppressed Anet more for C4 species than for C3 and CAM species. HNT increased plant height differently for C3 and C4 species by 6.41 and 150%, respectively. For woody species, Anet, Rd and biomass (stem and below-ground) responded more positively, while gs and plant height responded less positively to HNT than for herbaceous species (Figure 3). The LNT effect on woody and herbaceous species was significant for Anet, gs, Tr, stem dry weight and plant height. LNT had a less negative effect on Anet, Tr, and gs but a larger negative effect on stem dry weight and plant height in herbaceous species than in woody species.
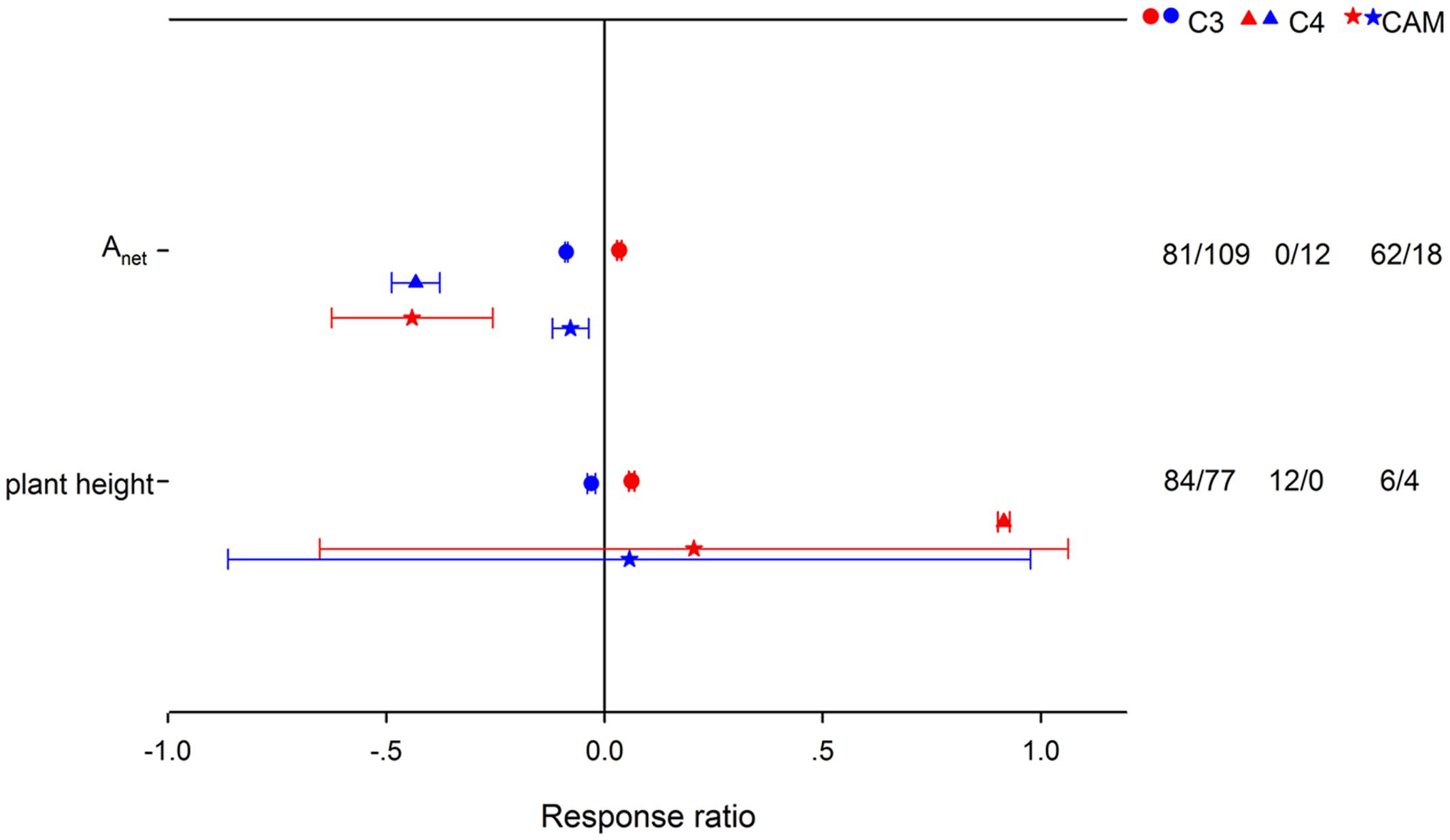
FIGURE 2. Photosynthetic rate (Anet) and plant height responses to HNT (red) and LNT (blue) in C3 (circles), C4 (triangles), and CAM (stars) species. Each data point represents the mean ± 95% CI. The number of observations for each variable is given on the right of the graph.
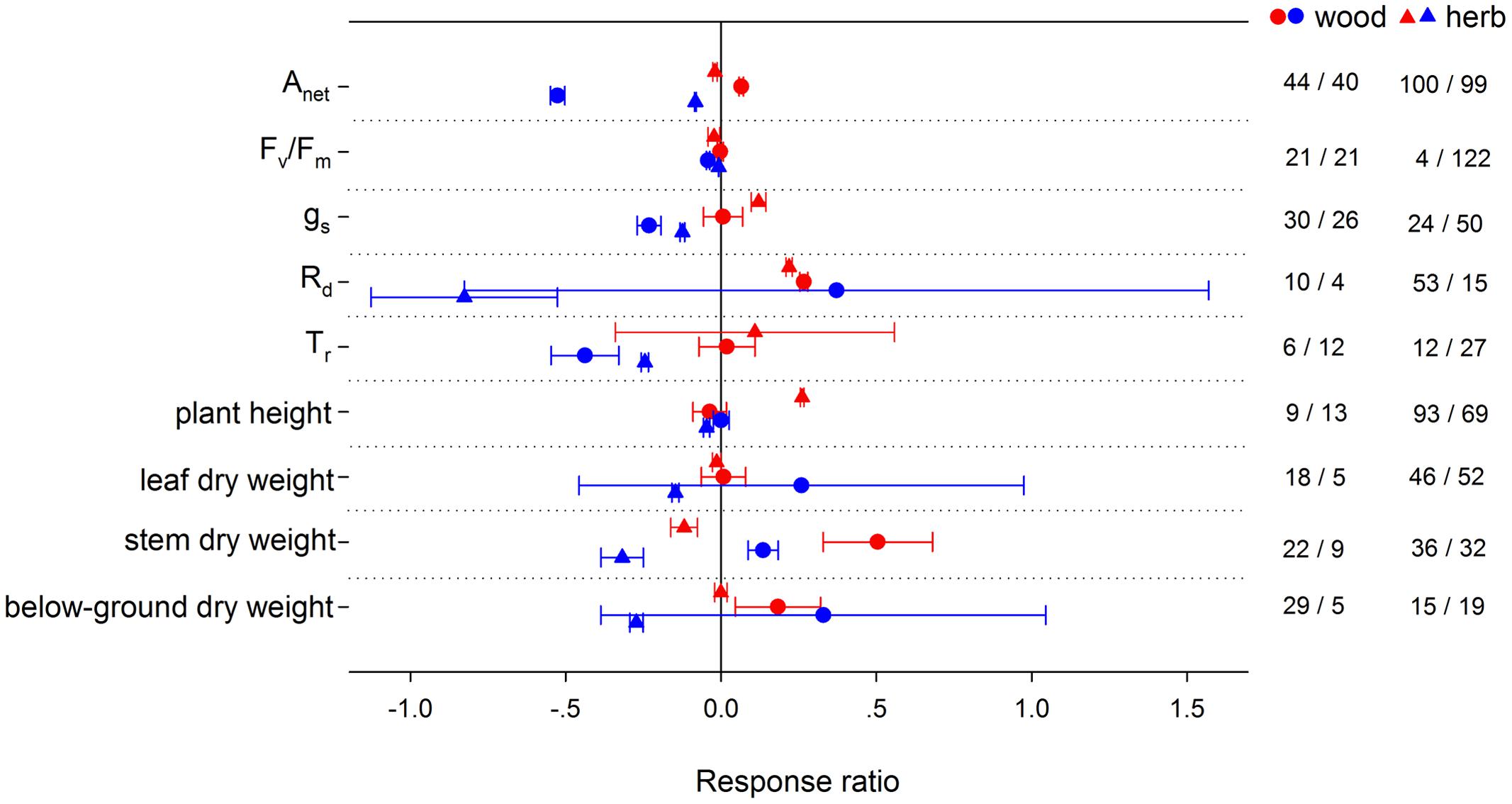
FIGURE 3. Plant physiological, morphological and yield-related responses to HNT (red) and LNT (blue) in woody (circles) and herbaceous (triangles) species. Each data point represents the mean ± 95% CI. The number of observations for each variable is given on the right of the graph.
High night temperature had a greater positive effect for gs and Rd in crops and Anet in non-crops (Figure 4). Positive effects of LNT on gs and TNC were significantly greater for non-crops than for crops. Positive effects of HNT on plant height, number of leaves and LAR were greater in crops, but the effects on SLA and LAI were greater in non-crops (Figure 5). LNT decreased plant height and number of leaves more in crops, but decreased SLA and LAI more in non-crops. HNT had positive effects on leaf, stem and total dry weight for crops but negative effects on non-crops (Figure 6). For below ground, dry weight and number of reproductive organs, non-crops responded more negatively to HNT than crops. LNT had positive effects on above-ground and total dry weight for crops and negative effects for non-crops, while stem dry weight in crops and non-crops responded to LNT oppositely. LNT had a greater negative effect on leaf dry weight and anthesis for crops than for non-crops.
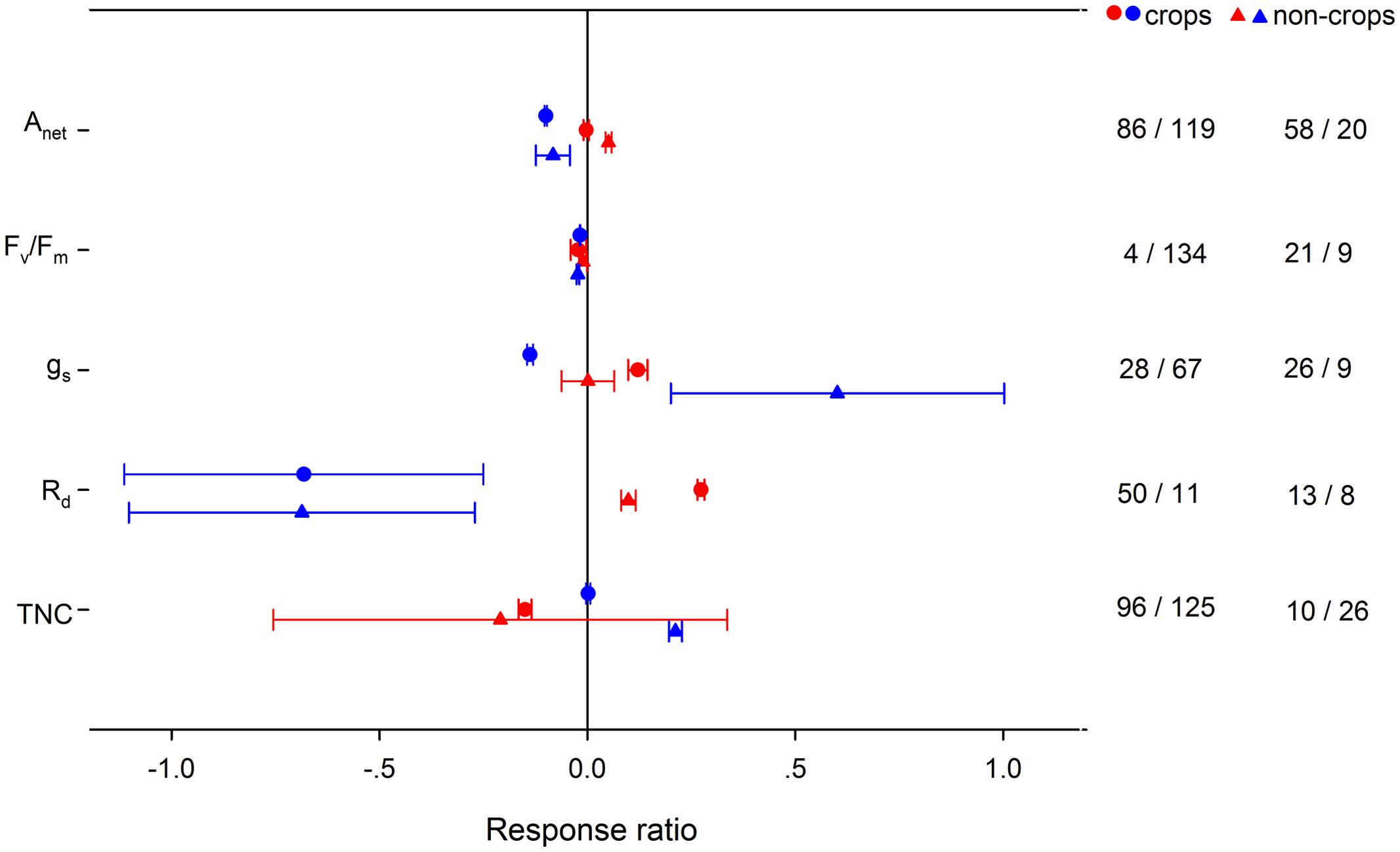
FIGURE 4. Plant physiological responses to HNT (red) and LNT (blue) in crops (circles) and non-crops (triangles) species. Each data point represents the mean ± 95% CI. The number of observations for each variable is given on the right of the graph.
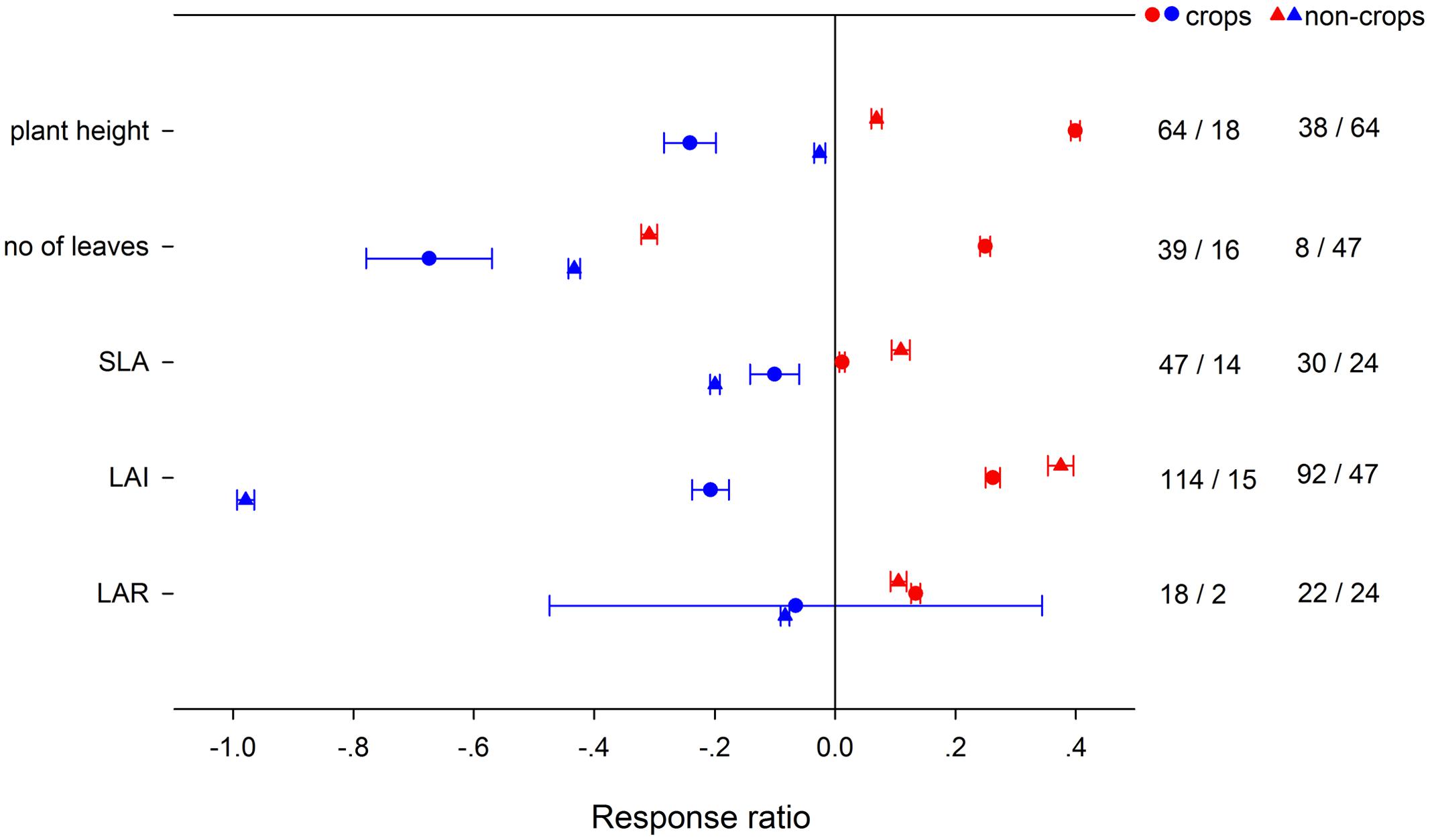
FIGURE 5. Plant morphological responses to HNT (red) and LNT (blue) in crops (circles) and non-crops (triangles) species. Each data point represents the mean ± 95% CI. The number of observations for each variable is given on the right of the graph.
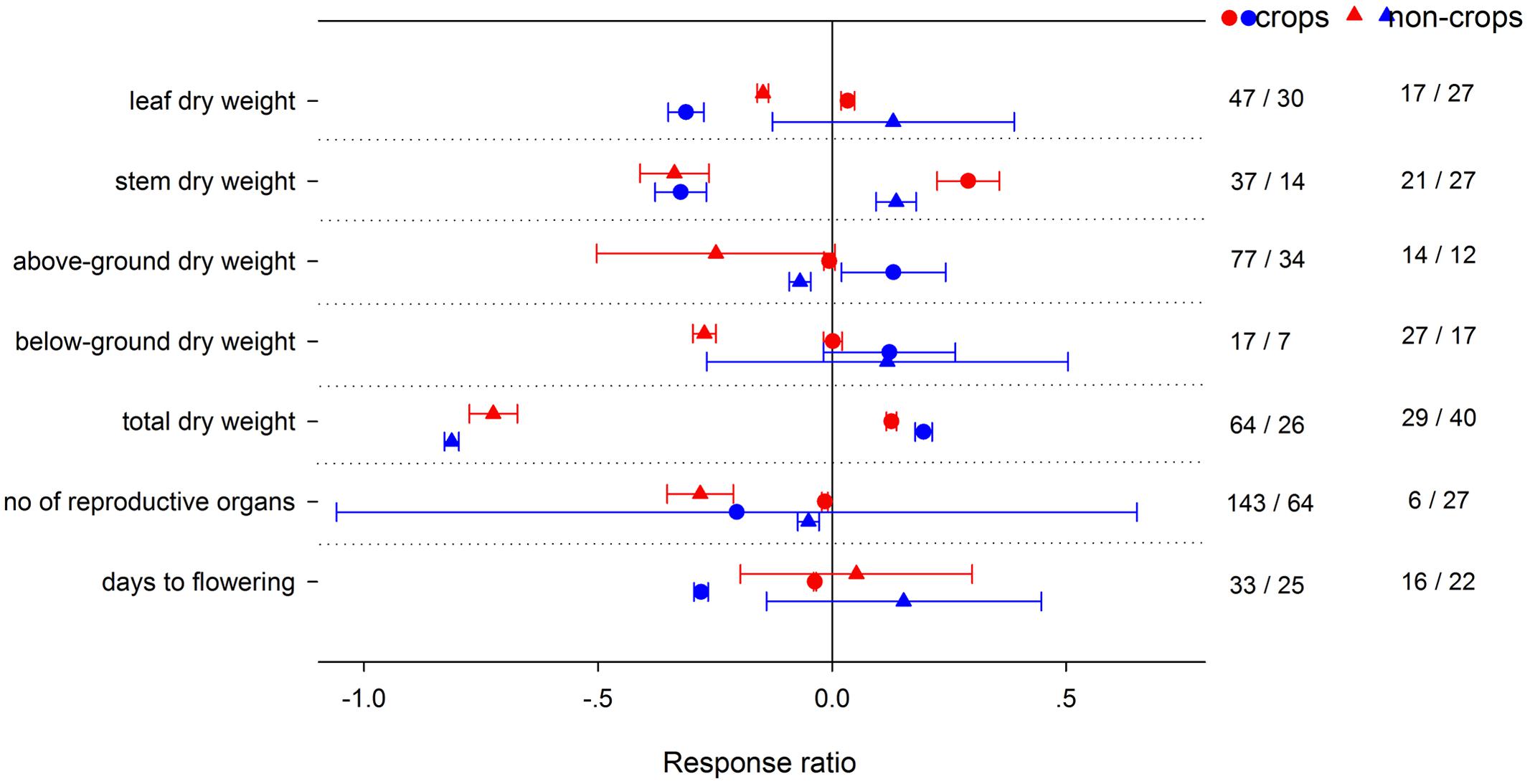
FIGURE 6. Plant yield-related responses to HNT (red) and LNT (blue) in crops (circles) and non-crops (triangles) species. Each data point represents the mean ± 95% CI. The number of observations for each variable is given on the right of the graph.
Magnitude-Introduced Uncertainty
Most ecophysiological and growth parameters formed a quadratic relationship, except for Rd, which responded linearly, to night temperature treatment (Figures 7–9). Anet, gs, and tissue N were the highest when NT was 0.675, 5.43, and 2.1°C above ambient temperature, respectively (Figure 7). Morphological parameters, including number of leaves, LAI, SLA, and LAR, formed downward-opening parabola relationships with night temperature change, while plant height, on the other hand, formed an upward-opening parabola relationship with night temperature change (Figure 8). Yield-related parameters including leaf, stem, above-ground and below-ground dry biomass as well as the number of reproductive organs, days to flowering, fruit size and fruit weight had downward-opening quadratic relationships with night temperature change (Figure 9).
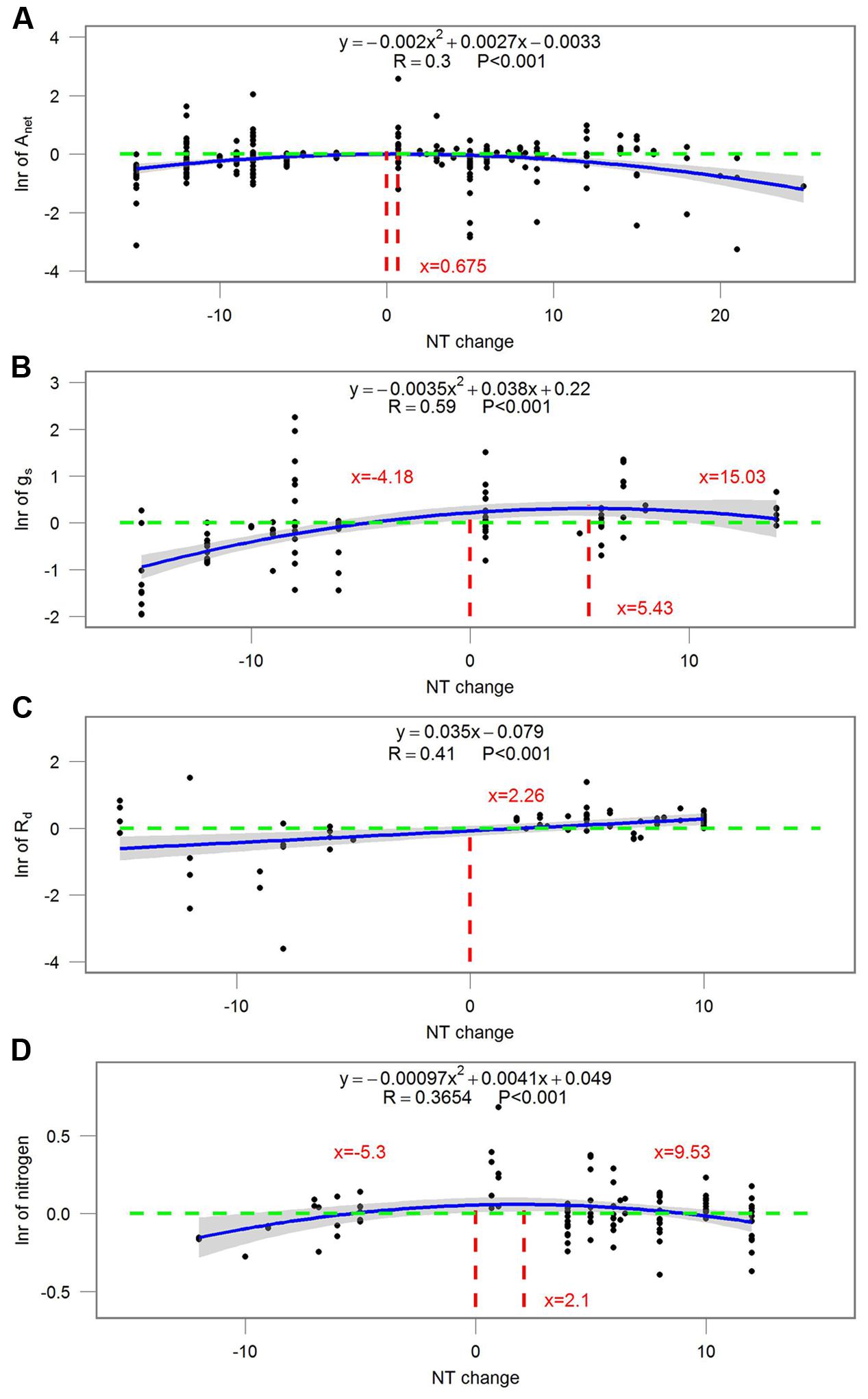
FIGURE 7. Correlations between the magnitude of NT treatment and the response ratio of (A) net photosynthetic rate (Anet); (B) stomatal conductance (gs); (C) dark respiration rate (Rd) and (D) tissue nitrogen content (N). Each point represents response ratio (lnr) to HNT or LNT. Regression function, variation coefficient and p-value are presented in the middle of each graph. Different lines indicate X = 0 (red line), x-value when y is the maximum, crossing points of y = 0 (green line) and regression relationships. Note that N is tissue nitrogen content including stem, leaf, panicle, spike, grain, shoot, root, and total N.
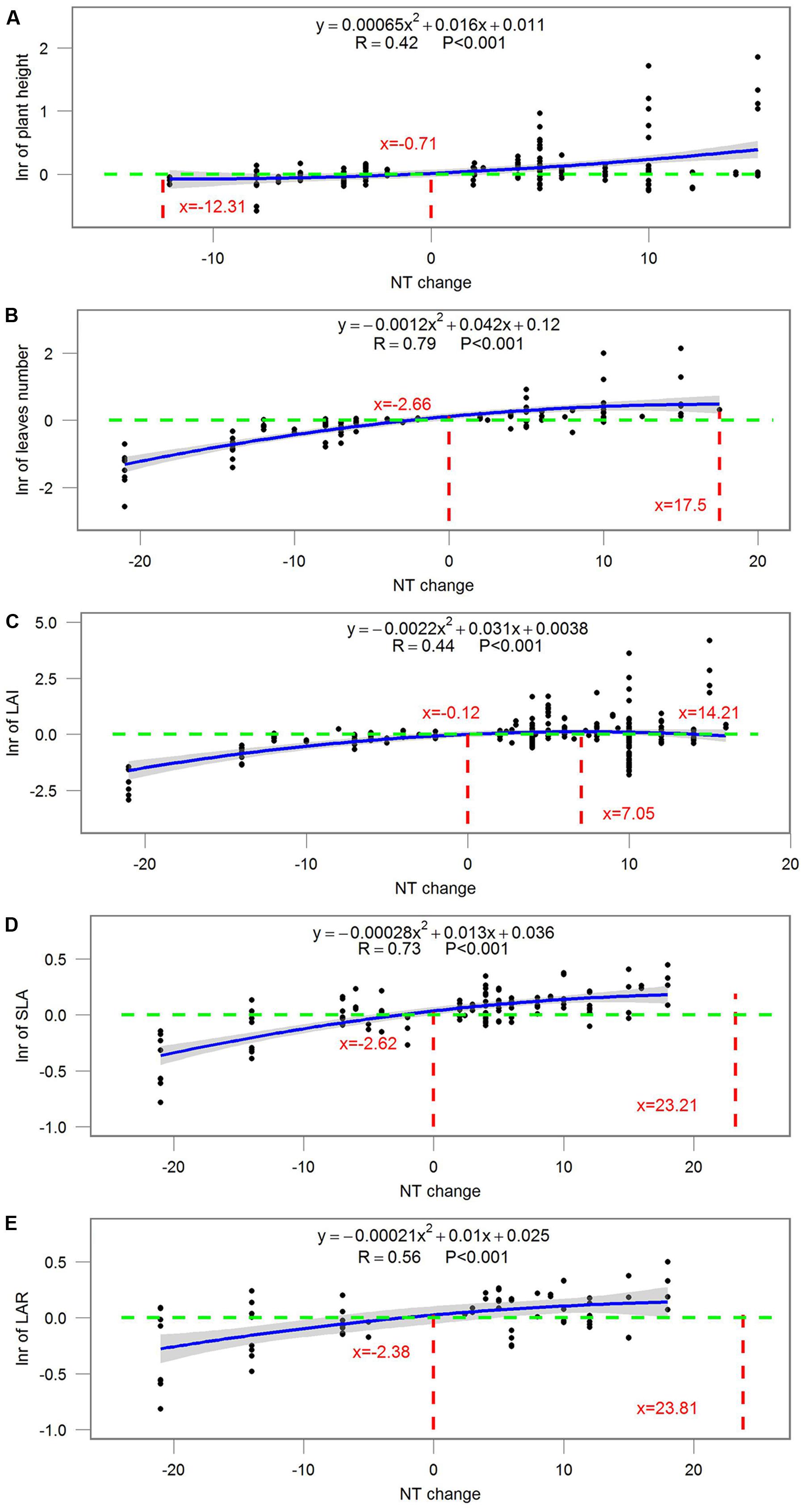
FIGURE 8. Correlations between the magnitude of NT treatment and the response ratio of (A) plant height; (B) number of leaves; (C) leaf area index (LAI); (D) specific leaf area (SLA); (E) leaf area ratio (LAR). Each point represents response ratio (lnr) to HNT or LNT. Regression function, variation coefficient and p-value are presented in the middle of each graph. Different lines indicate X = 0 (red line), x-value when y is the maximum, crossing points of y = 0 (green line) and regression relationships.
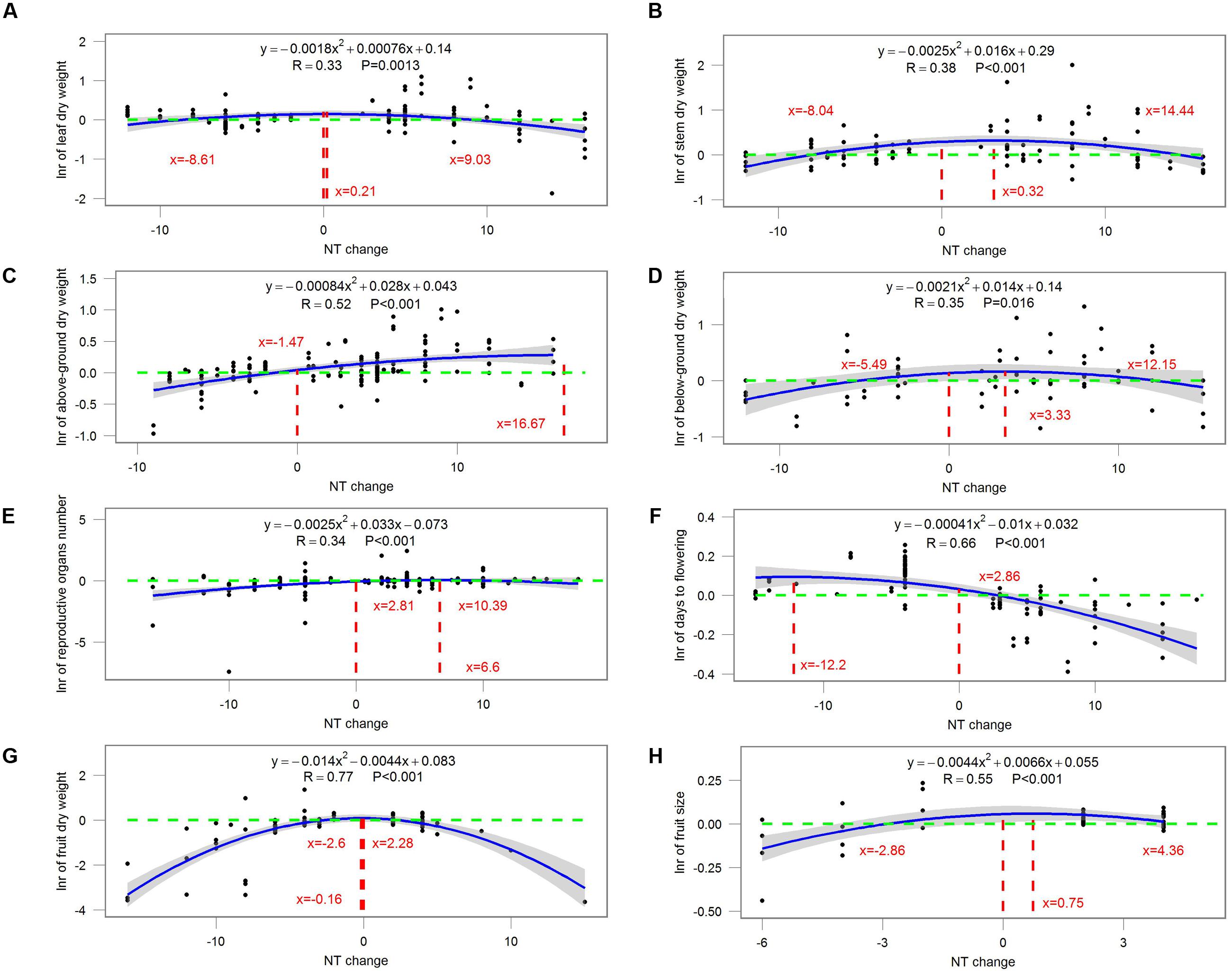
FIGURE 9. Correlations between the magnitude of NT treatment and the response ratio of (A) leaf dry weight; (B) stem dry weight; (C) above-ground dry weight; (D) below-ground dry weight; (E) number of reproductive organs; (F) days to flowering; (G) fruit dry weight and (H) fruit size. Each point represents response ratio (lnr) to HNT or LNT. Regression function, variation coefficient and p-value are presented in the middle of each graph. Different lines indicate X = 0 (red line), x-value when y is the maximum, crossing points of y = 0 (green line) and regression relationships.
Uncertainty Related to the Experimental Design
Pot size involved in the experiments was a significant factor influencing the effects on Anet, tissue N, TNC, total dry weight, number of reproductive organs, and plant height’s responses to HNT (Figure 10). Plants in smaller pots (<10L) responded more positively for Anet, N, wt, and plant height to HNT. TNC, however, responded more negatively at HNT in larger pots. N, wt, number of reproductive organs and plant height were reduced more in smaller pots at LNT. Anet was decreased more and TNC was increased more in larger pots with LNT.
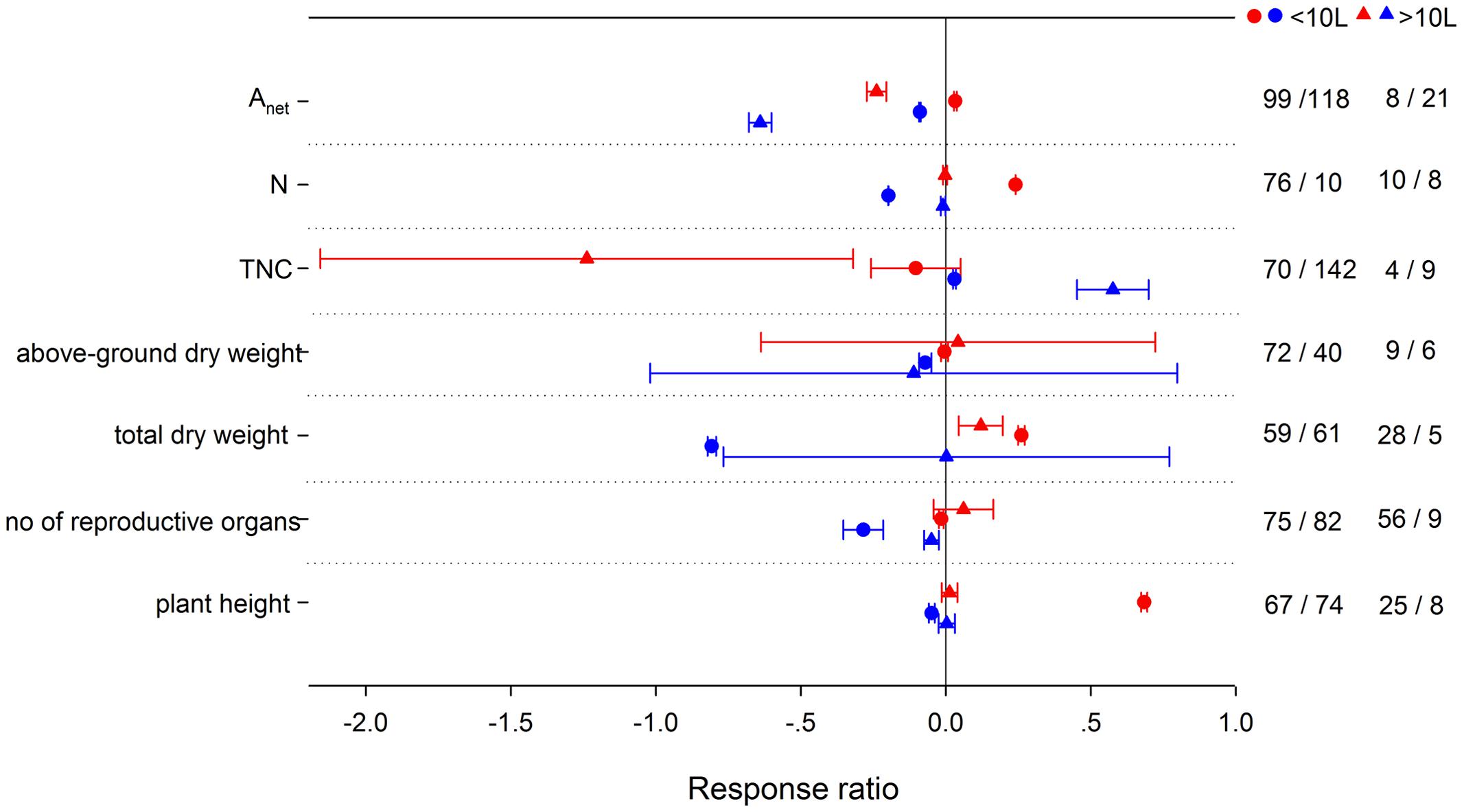
FIGURE 10. Plant physiological, morphological and yield-related responses to HNT (red) and LNT (blue) grown in <10 L pots (circles) or >10L pots (triangles). Each data point represents the mean ± 95% CI. The number of observations for each variable is given on the right of the graph. Note that N is tissue nitrogen content including stem, leaf, panicle, spike, grain, shoot, root, and total N.
Experimental duration also played an important role in affecting plant responses to HNT and LNT. HNT increased respiration less in short-term treatments than that in long-term treatments, while LNT duration had insignificant effects (Figure 11). Stomatal conductance was significantly increased at short-term HNT but decreased at long-term HNT. Compared with short-term duration, long-term LNT caused more reduction on Anet and gs. Experimental duration had no effects on the responses of SLA to HNT or to LNT, but generated different effects on plant height and LAI at both HNT and LNT. Long-term HNT increased plant height and LAI more, whereas short-term LNT reduced plant height more and LAI less. For total dry weight, long-term HNT and LNT had greater effects than short-term. Yield was decreased more at short-term HNT and different durations had no significant effects in affecting yield responses to LNT.
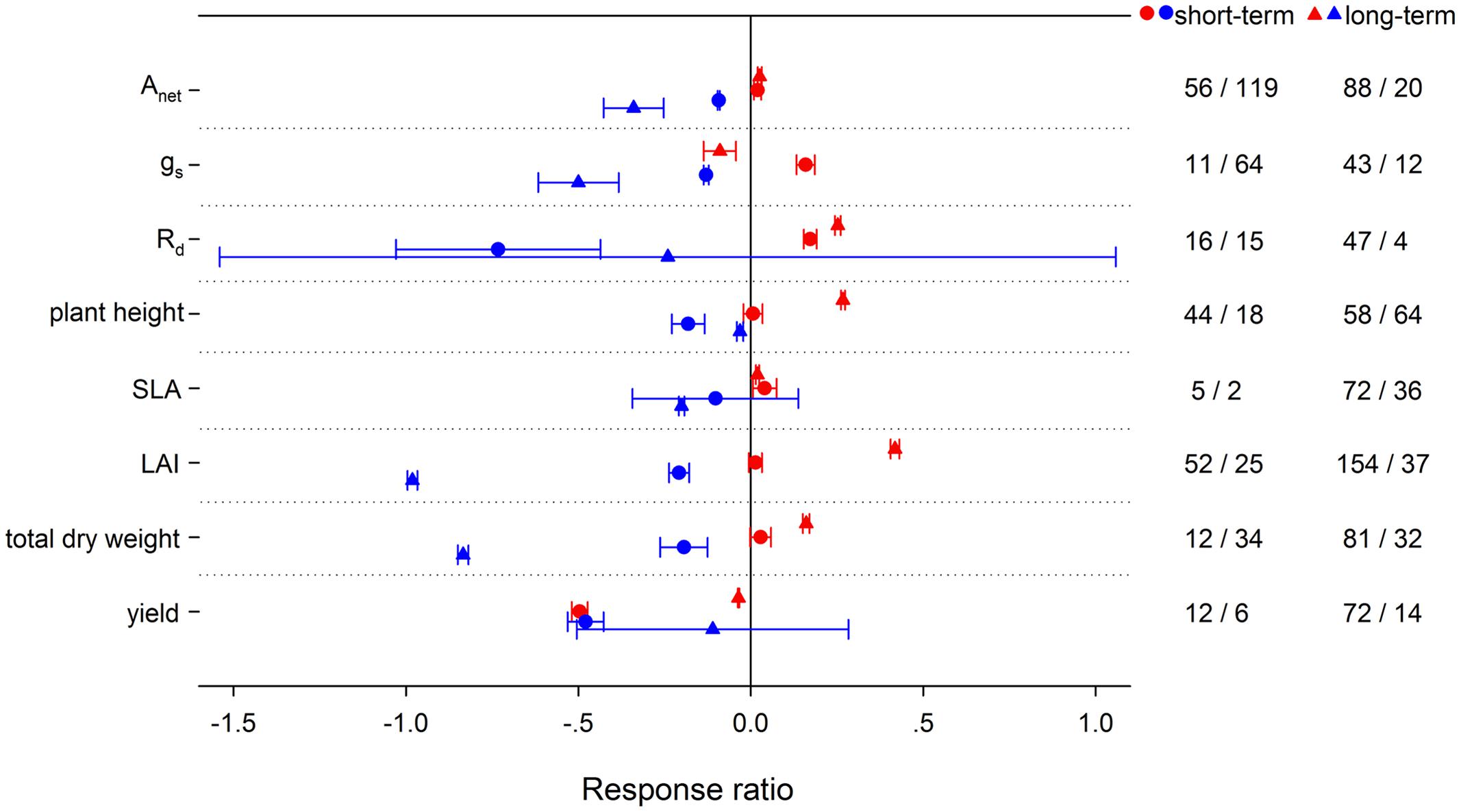
FIGURE 11. Plant physiological, morphological and yield-related responses to HNT (red) and LNT (blue) for different treatment durations (circles, short-term; triangles, long-term). Each data point represents the mean ± 95% CI. The number of observations for each variable is given on the right of the graph.
Discussion
Asymmetric night warming and chilling have long been acknowledged as a universal phenomenon in recent years and caused great agricultural, economic and ecological consequences (Xia et al., 2014). At the leaf and organism level, however, comprehensive quantitative analysis of the effect of HNT and LNT on plants ecophysiology and growth remain unclear. In this study, we collected data from night temperature manipulative studies and analyzed the changes in ecophysiological and whole-plant responses due to changes in night temperatures. Overall, we found that: (1) the significance and degree of the effect of HNT and LNT and the causes of yield reduction at HNT and LNT were different; (2) there existed significant variations among different PFTs in responding HNT and LNT; (3) there was an optimal night temperature for important processes of plants physiology and growth; (4) the responses to HNT and LNT appeared dependent of the experimental designs.
Plant Responses to HNT and LNT
Consistent with our hypothesis, both HNT and LNT had a negative effect on plants yield, with a greater negative effect at LNT, probably due to a greater night temperature reduction for LNT treatments (Supplementary Figure S2). HNT and LNT are considered great threats to plants production, especially for crops (Hall, 2000; Maali and Heidarvand, 2010). The impacts of temperature change on plant yield have been assessed directly through greenhouse (Cheng et al., 2009; Mohammed and Tarpley, 2009a,b; Kanno and Makino, 2010; Zhang et al., 2010; Qi et al., 2011) and field experiments (Peng et al., 2004; Nagarajan et al., 2010; Shi et al., 2013; Zhang Y. H. et al., 2013). The long-term effect of climate change on crops was also estimated through crop-growth models, such as CERES (Ritchie and Otter, 1985; Jones et al., 1986), ORYZA2000 (Bouman and Laar, 2006), and CropSyst (Stockle et al., 1994). Warming stress triggered a significant loss of crop yield worldwide, particularly in nations such as China (Li et al., 2004), Japan (Hasegawa et al., 2009), Philippines (Peng et al., 2004), as well as nations in south and southeast Asia (Welch et al., 2010). The deduction of yield was often associated with the decrease in the number of panicles (Peng et al., 2004), grain maturity (Suzuki and Moroyu, 1962; Ziska and Manalo, 1996) and final grain weight (Morita, 2005; Kanno and Makino, 2010), spikelet number per plant (Morita et al., 2002, 2004; Peng et al., 2004) and spikelet fertility (Cheng et al., 2009; Mohammed and Tarpley, 2009a, 2010), accelerative respiration rates (Mohammed and Tarpley, 2010), grave membrane leakage (Mohammed and Tarpley, 2009b), lower pollen germination (Mohammed and Tarpley, 2009a), and poor assimilates and N translocation to grains (Morita, 2005; Cheng et al., 2009; Kanno and Makino, 2010; Shi et al., 2013).
We found that the negative effect of HNT on yield was associated with a reduction in number of reproductive organs, fruit dry weight, and time for flowering. The reproductive process was regarded as most susceptible to heat stress (Prasad et al., 2006; Jagadish et al., 2007, 2008, 2010), with limited pollen viability as the major cause of yield reduction (Yoshida et al., 1981; Ziska and Manalo, 1996; Jagadish et al., 2010; Zhang et al., 2010; Zhang Y. H. et al., 2013; Fang et al., 2013; Rehmani et al., 2014). Decreased seed yield and lower seed-set under HNT were also reported in wheat (Prasad et al., 2008), rice (Mohammed and Tarpley, 2009b), cowpea (Ahmed et al., 1993), and tomato (Peet and Bartholemew, 1996). Low temperature is one of the most important abiotic stresses for plant growth, development, energy distribution (Xiong et al., 2002; Oufir et al., 2008), and yield (Kasuga et al., 1999; Lang et al., 2005). The negative effect of LNT on yield in this study appeared to be associated with a decline in the number of reproductive organs, fruit dry weight, as well as above-ground, below-ground, and total dry weight. Clearly, the mechanisms for the negative effect of HNT and LNT on plant yield were different (Figure 1). The negative effect of LNT on plant yield was primarily related to the negative effect of LNT on the total biomass accumulation, but the negative effect of HNT on plant yield was caused mostly by the reduced allocation of biomass to reproductive organs, as the total biomass was even stimulated by HNT. HNT had been shown to cause no change, or indeed an increase, in total biomass accumulation in crop plants such as rice (Ziska and Manalo, 1996; Cheng et al., 2009), sorghum and sunflower (Manunta and Kirkham, 1996), tobacco (Camus and Went, 1952), and cotton (Königer and Winter, 1993).
The balance between photosynthesis and respiration controls plant growth. Several recent meta-analyses of plant responses to increasing temperature had highlighted how plants may be particularly vulnerable to increases in both HNT and LNT (Lin et al., 2010; Way and Oren, 2010). In contrast to the prediction that Anet is constrained at both LNT and HNT (Berry and Bjorkman, 1980), HNT increased Anet by 2.56% and LNT decreased Anet by 8.73% among the plants included in this study (Figure 1A). The positive effect of HNT on Anet could be associated with the increment of gs and tissue N, instead of PSII quantum yield and ETR (Figure 1A). HNT increased gs of two rice genotypes (Shah et al., 2011) and wheat (Lu et al., 1998). HNT enhanced nitrogen soil mineralization (Sardans et al., 2008; Patil et al., 2010) and therefore increased leaf N concentrations (Rustad et al., 2001), which was closely related to photosynthetic capacity (Mae, 1997; Llorens et al., 2003). The loss of chlorophyll owing to HNT has been reported in many crops (Reynolds et al., 1994; Guo et al., 2006). The negative effect of LNT on Anet was associated with the negative effect on gs, PSII function (ΦPSII, ETR, and Fv/Fm), ribulose-1,5-bisphosphate carboxylase (RuBP) inactivation and leaf N (Figure 1A), consistent with other studies (Allen and Ort, 2001; Huang and Guo, 2005; Bertamini et al., 2007). Photosystem II has been regarded as the most sensitive to LNT (Huang et al., 2010), as LNT induced PSII photoinhibition and caused reversible inhibition of photosynthetic capacity (Dongsansuk et al., 2013; Zhang et al., 2014).
Our meta-analysis indicated that dark respiration (Rd) was increased with HNT but was decreased with LNT. Increased dark respiration in HNT was reported in rice (Mohammed and Tarpley, 2009b) and Stipa krylovii Roshev (Chi et al., 2013a,b) at leaf scale and in rice (Cheng et al., 2009), lettuce, tomato, soybean (Frantz et al., 2004), and cotton (Loka and Oosterhuis, 2010) at organism scale. Different from the hypothesis proposed by Peng et al. (2004) that HNT increased biomass loss by enhancing respiration, our study concluded that HNT increased total biomass but LNT decreased the total, above-ground and below-ground biomass. Total biomass was stimulated by HNT in rice (Cheng et al., 2008, 2009), Ficus insipida and Ochroma pyramidale (Cheesman and Klaus, 2013), Rosebay rhododendron (Starrett et al., 1993), panicum (Patterson, 1990) and soybean (Hewitt et al., 1985). Studies in natural systems have seen impacts on plant phenological development. For example, the large herbaceous perennial, Phytolacca americana (Phytolaccaceae) demonstrated no difference in biomass accumulation but had flower set faster as a result of HNT (Wolfe-Bellin et al., 2006). Studies in temperate North America demonstrated that HNT in urban environments increased growth rates in seedlings of Quercus rubra (Searle et al., 2012). Similarly, changes in Rd of non-photosynthetic tissues (Saveyn et al., 2008), or a change in carbon-use efficiency (Hansen et al., 2009), could contribute to increased growth rate under HNT.
The balance between photosynthesis and respiration also controls carbohydrate accumulation, which is essential for plant growth and development (Azcón-Bieto and Osmond, 1983; Guy et al., 1992). The processes of photosynthesis and respiration responded independently to temperature and are linked by leaf carbon status (Turnbull et al., 2002). TNCs including starch and sucrose responded differently to HNT and LNT. Although HNT stimulated both photosynthesis and respiration, carbohydrate content was significantly decreased, probably due to the imbalance of the stimulation of HNT on Rd and Anet. Leaf carbohydrates synthesized during the daytime were observed to be consumed more quickly during warmer nights because of enhanced leaf respiration, which depletes foliar carbohydrates and may produce a rebound effect of compensatory stimulated photosynthesis during the following day. Evidence supporting this hypothesis has been reported in both greenhouse and field experiments (Beier et al., 2004; Lin et al., 2010). Turnbull et al. (2002) found that leaf starch concentration, soluble sugar and total non-structural carbohydrate declined significantly with the increase of nocturnal temperature. However, no compensatory effect was found between respiration and photosynthesis under nocturnal warming in Mediterranean grassland (Xia et al., 2014). Further investigation is required to discover whether the compensatory effect between respiration and photosynthesis under night warming is related to other environmental conditions, such as water and nitrogen availability. LNT increased carbohydrate content via a lesser negative effect on Rd than on Anet, which was approved in many studies (Siminovitch and Briggs, 1953; Steponkus and Lanphear, 1968; Guy et al., 1980; Kaurin et al., 1981; Miao et al., 2009). The increased carbohydrate content posed a negative effect on the day-time photosynthesis at LNT.
Plants adapt to climate stresses via multiple strategies, such as the adjustments of phenology and morphology (Wahid et al., 2007). Leaf morphology was particularly sensitive to HNT as leaf expansion often reached its peak during the night (Schurr et al., 2000; Pantin et al., 2011). HNT had positive effects on SLA, LAR, number of leaves and LAI. When exposed to HNT, the expansion of leaf area and plant height benefited capturing more light for photosynthesis (Seddigh and Jolliff, 1984c; Darnell et al., 2013). Elevated water temperature, in addition to air temperature, can also stimulate leaf expansion and elongation (Tsunoda, 1964; Cutler et al., 1980). Kanno et al. (2009) attributed the positive effect of HNT on plant biomass to an increase in leaf area in rice plants, which was found in tomatoes and Galega officinalis (Hussey, 1965; Patterson, 1993) as well. LNT, on the other hand, suppressed plant height, number of leaves, LAI, SLA, and LAR, corresponding to the previous studies (Cockshull, 1979; Dejong and Smeets, 1982; Patterson, 1993; Kjær et al., 2007, 2008; Kjaer et al., 2010).
Variable Responses among Plant Functional Types (PFTs)
Previous studies indicated that increased daytime temperature had stronger effects on Anet of C3 species than C4 species (Wahid et al., 2007). No consistent results for HNT effect on Anet were found due to insufficient data for C4 species. However, we did find that LNT decreased Anet more in C4 species than in C3 species, which was closely associated with relatively more reduction of gs, ETR, Vcmax, and Jmax for C4 species (Figure 2; Supplementary Figure S4). Low temperature effects on C4 photosynthesis have been frequently examined (Long, 1998). C4 photosynthesis has been suggested to be inherently sensitive to chilling because of the cold lability of the C4 cycle enzymes (Long, 1983; Sage, 2002; Sage and Kubien, 2007).
High night temperature stimulated Anet in woody plants but suppressed it in herbaceous plants. The same pattern was found in stem biomass. Though studies have reported a greater warming-induced stimulation in woody biomass than in herbaceous biomass (Lin et al., 2010), the result that woody plants were more favored under night warming than herbaceous plants has not been reported. The stimulation of HNT on stem biomass is greater than that on below-ground biomass for woody species, however, for herbaceous species, HNT had no effect on leaf- or below-ground biomass, yet suppressed stem biomass significantly. Our results imply that more resources will be allocated to aboveground growth, and therefore above-ground competition for resources, such as light, will be more important for woody species under night warming (Suding et al., 2005; Lin et al., 2010). In ecosystems where herbaceous and woody plants coexist, a greater biomass stimulation of woody than of herbaceous species may increasingly suppress growth, especially above-ground growth, of herbaceous species via a shading effect (Castro and Freitas, 2009). LNT also had a positive effect on woody stem biomass, which might be related to a stronger suppression in Rd than Anet. The negative effect of HNT on Anet for herbaceous plant was caused primarily by damage to PSII efficiency (Fv/Fm) (Figure 3). LNT decreased Anet more in woody plants than in herbaceous plants, along with a greater decrease of Fv/Fm and gs in woody plants, which was consistent with previous studies (Sao et al., 2010, 2013b; Liu et al., 2011). It is important, however, to note that more research is needed on the effect of night temperature on plants biomass allocation, since the publication bias on this effect could not be ignored in this meta-analysis. Low sample sizes for some functional groups used in this analysis (e.g., C4 and woody species) require that some results in this analysis be interpreted with caution. Nevertheless, results from these under-represented groups demonstrate that further study of these groups is critical for this untested hypothesis in the future.
Teasing apart the variations of the responses among crops and non-crops is important to guide future research in agricultural practice and genetic engineering. HNT had a positive effect on Anet for non-crops but had no effect for crops, while HNT had a positive effect on Rd for both crops and non-crops. Due to different responses of Anet and Rd to HNT between crops and non-crops, TNC was decreased in crops but unchanged in non-crops. Accordingly, HNT stimulated leaf-, stem-, total dry biomass for crops but decreased leaf-, stem-, below-ground, total dry biomass for non-crops. Similarly, crops better adapted to LNT than non-crops, as LNT had a positive effect on total dry biomass for crops but negative for non-crops. The fact that crops coped with HNT and LNT better than non-crops could imply an improved stress-tolerant ability for improved crops through breeding, genetic engineering, and management practices. Changes in HNT and LNT could influence vegetation dynamics and ecosystem structure through shifting competitive interactions among different functional groups in natural or agricultural systems.
Uncertainties
The magnitude of night temperature treatment did have an impact on most of the parameters that were investigated in the study. The negative effects of HNT and LNT on plants ecophysiological parameters (Anet, gs, and tissue N), morphological parameters (SLA, LAI, and LAR), and yield related parameters (above-ground and below-ground biomass, fruit size and dry weight) were more evident with the increasing magnitude of HNT and LNT treatment (Figures 7–9). Consistent with the results discussed before, plant peak physiology and growth mostly occurred at night temperatures higher than the ambient, especially for leaf growth. Plant height, on the other hand, was even stimulated by HNT and not much affected by LNT. Whether plants at HNT tend to be thinner and taller requires further investigation. In all cases, photosynthesis shows an optimum temperature that roughly corresponds to the middle of the non-harmful range and drops off with an increased slope as temperatures rise above the thermal optimum.
It is essential that potential confounding factors be considered in a meta-analysis, which synthesizes results from a large number of studies conducted under a variety of growing conditions on different plant species. In our analysis, studies in plant responses to other environmental stresses (e.g., drought, low nutrients, light deficiency, or elevated ozone) were excluded. In addition to the variation caused by PFTs, different experimental design can be also responsible for the inconsistent responses (Mohammed and Tarpley, 2009a,b; Nagarajan et al., 2010; Cheesman and Klaus, 2013; Rehmani et al., 2014). Here we focused on the effects of pot size (<10L vs. >10L) and treatment duration (short vs. long term) on plant responses. Responses of plant growth under HNT and LNT may vary with time because thermal sensitivity of plants may differ among growth stages. Long-term LNT treatment had a stronger negative effect on Anet than short-term treatment, whereas HNT treatment of different durations had no significant effect on Anet. On the contrary, Rd was increased greater in long-term HNT but not different between different LNT durations. Several studies had reported the long-term acclimatization of dark respiration on tropical trees (Atkin and Tjoelker, 2003; Cheesman and Klaus, 2013). Total biomass responded differently between different treatment durations of HNT and LNT, though no significant experiment duration effect was found in above- and below-ground dry matter. Plant growth and yield were decreased less at long-term HNT, which might be related to plant acclimation ability (Hare et al., 1998; Wahid et al., 2007). Pot size significantly altered the responses of Anet, non-structural carbohydrates and total biomass to HNT and LNT. Small pots constrained below-ground growth along with more limitation on above-ground growth (Thomas and Strain, 1991; Loh et al., 2003; Climent et al., 2011), given that a skimpy supply of nutrients and water which might induce strong nutrient or water inhibition (Walters and Reich, 1989).
Conclusion
We found that both HNT and LNT had a negative effect on plants yield, with the HNT effect primarily related to reduced biomass allocation to reproductive organs, flower development and seed maturation and the LNT effect more related to a negative effect on total biomass. HNT accelerated plants ecophysiological processes, including photosynthesis and dark respiration, while LNT slowed these processes. The responses to LNT and HNT varied significantly among different PFTs. HNT stimulated photosynthesis in C3, woody and non-crop species, but inhibited it in herbaceous, and had no effect in crops. LNT caused more reduction of Anet in woody than in herbaceous species but decreased it for both crops and non-crops with no significant difference. Both experimental settings and durations had significant effects in plants responses to night temperature change. Long-term HNT led to a relatively smaller loss of yield while the response of yield to LNT was unchanged by different treatment durations. The magnitude of night temperature did have an impact on most of the parameters that were investigated in the study. Plants peak physiology and growth mostly occurred at night temperatures higher than the ambient, especially for leaf growth. Our results suggest that the diurnal variations in vegetation responses to night temperature changes are important for understanding the changes in vegetation photosynthetic activity and growth in future climates. Such diurnal variations, however, have rarely been incorporated into current modeling studies on vegetation responses to global warming—which are overwhelmingly based on daily or growing season mean air temperature and will not capture the response of vegetation to asymmetric diurnal temperature changes. New field experiments with different elevated temperatures during day vs. night, and across different seasons, are urgently needed for different plant functional groups. Such experiments will shed new light on the ecophysiological effects of night-time temperature change during different seasons, and can be used to improve the performance of current land surface models. The functional type specific response patterns of plants to night temperature changes are critical for obtaining credible predictions of the changes in food production, carbon sequestration and climate regulation.
Author Contributions
DW proposed the research idea. PJ collected and analyzed the data and then wrote the paper. DW, CZ, and JC help modify the paper.
Funding
Funding for this research was provided by Nanjing University of Information Science and Technology, the Jiangsu Natural Science Foundation (BK20150894), The National Natural Science Foundation of China (31500503), and the International S&T Cooperation Program of China (2012DFA60830) through DW.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
We thank Gabriela Shirkey of Michigan State University for editing the language.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.01774/full#supplementary-material
Abbreviations
ANT, ambient night temperature; Anet, net CO2 assimilation rate (μmol m-2 s-1); C, carbon; Ci, intercellular CO2 concentration (μmol mol-1); ETR, electron transport rate (μmol m-2 s-1); Fv/Fm, Photosystem II (PSII) efficiency; gs, stomatal conductance (mol m-2 s-1); HNT, high night temperature; LNT, low night temperature; LAI, leaf area index; LAR, leaf area ratio (cm2 g-1); N, nitrogen; Jmax, maximum electron transport rate (μmol m-2 s-1); PFTs, plant functional types; ΦPSII, PSII quantum yield; Rd, dark respiration rate (μmol m-2 s-1); RuBP, ribulose-1,5-bisphosphate carboxylase; SLA, specific leaf area (cm2 g-1); TNC, total non-structural carbohydrate (mg g-1); Tr, transpiration rate (mmol m-2 s-1); Vcmax, maximum carboxylation rate (μmol m-2 s-1).
References
Ahmed, F. E., Hall, A. E., and Madore, M. A. (1993). Interactive effects of high temperature and elevated carbon dioxide concentration on cowpea [Vigna unguiculata (L.) Walp.]. Plant Cell Environ. 16, 835–842. doi: 10.1016/j.jphotobiol.2010.05.013
Allen, D. J., and Ort, D. R. (2001). Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 6, 36–42. doi: 10.1016/S1360-1385(00)01808-2
Alward, R. D., Detling, J. K., and Milchunas, D. G. (1999). Grassland vegetation changes and nocturnal global warming. Science 283, 229–231. doi: 10.1126/science.283.5399.229
Atkin, O. K., and Tjoelker, M. G. (2003). Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 8, 343–351. doi: 10.1016/S1360-1385(03)00136-5
Azcón-Bieto, J., and Osmond, C. B. (1983). Relationship between photosynthesis and respiration. Plant Physiol. 71, 574–581. doi: 10.1104/pp.71.3.574
Bagnall, D. J., King, R. W., and Farquhar, G. D. (1988). Temperature-dependent feedback inhibition of photosynthesis in peanut. Planta 175, 348–354. doi: 10.1007/BF00396340
Bange, M. P., and Milroy, S. P. (2004). Impact of short-term exposure to cold night temperatures on early development of cotton (Gossypium hirsutum L.). Aust. J. Agric. Res. 55, 655–664. doi: 10.1071/AR03221
Begg, C. B., and Mazumdar, M. (1994). Operating characteristics of a rank correlation test for publication bias. Biometrics 50, 1088–1101. doi: 10.2307/2533446
Beier, C., Emmett, B., Gundersen, P., Tietema, A., Peñuelas, J., Estiarte, M., et al. (2004). Novel approaches to study climate change effects on terrestrial ecosystems in the field: drought and passive nighttime warming. Ecosystems 7, 583–597. doi: 10.1007/s10021-004-0178-8
Berry, J., and Bjorkman, O. (1980). Photosynthetic response and adaptation to temperature in higher plants. Ann. Rev. Plant Physiol. 31, 491–543. doi: 10.1146/annurev.pp.31.060180.002423
Bertamini, M., Muthuchelian, K., Rubinigg, M., Zorer, R., and Nedunchezhian, N. (2005). Low-night temperature (LNT) induced changes of photosynthesis in grapevine (Vitis vinifera L.) plants. Plant Physiol. Bioch. 43, 693–699. doi: 10.1016/j.plaphy.2005.06.001
Bertamini, M., Zulini, L., Muthuchelian, K., and Nedunchezhian, N. (2007). Low night temperature effects on photosynthetic performance on two grapevine genotypes. Biol. Plant. 51, 381–385. doi: 10.1007/s10535-007-0080-2
Bouman, B. A. M., and Laar, H. H. V. (2006). Description and evaluation of the rice growth model ORYZA2000 under nitrogen-limited conditions. Agric. Syst. 87, 249–273. doi: 10.1016/j.agsy.2004.09.011
Bunce, J. A. (1985). Effects of day and night temperature and temperature variation on photosynthetic characteristics. Photosynth. Res. 6, 175–181. doi: 10.1007/BF00032791
Camus, G. C., and Went, F. W. (1952). The thermoperiodicity of three varieties of Nicotiana tabacum. Am. J. Bot. 39, 521–528. doi: 10.2307/2438697
Castro, H., and Freitas, H. (2009). Above-ground biomass and productivity in the Montado: from herbaceous to shrub dominated communities. J. Arid Environ. 73, 506–511. doi: 10.1016/j.jaridenv.2008.12.009
Cheesman, A. W., and Klaus, W. (2013). Elevated night-time temperatures increase growth in seedlings of two tropical pioneer tree species. New Phytol. 197, 1185–1192. doi: 10.1111/nph.12098
Chen, W. H., Tseng, Y. C., Liu, Y. C., Chuo, C. M., Chen, P. T., Tseng, K. M., et al. (2008). Cool-night temperature induces spike emergence and affects photosynthetic efficiency and metabolizable carbohydrate and organic acid pools in Phalaenopsis aphrodite. Plant Cell Rep. 27, 1667–1675. doi: 10.1007/s00299-008-0591-0
Cheng, W., Hasegawa, T., Sakai, H., and Yagi, K. (2010). Combined effects of elevated [CO2] and high night temperature on carbon assimilation, nitrogen absorption, and the allocations of C and N by rice (Oryza sativa L.). Agric. For. Meteorol. 150, 1174–1181. doi: 10.1016/j.agrformet.2010.05.001
Cheng, W., Sakai, H., Hartley, A., Yagi, K., and Hasegawa, T. (2008). Increased night temperature reduces the stimulatory effect of elevated carbon dioxide concentration on methane emission from rice paddy soil. Glob. Change Biol. 14, 644–656. doi: 10.1111/j.1365-2486.2007.01532.x
Cheng, W., Sakai, H., Yagi, K., and Hasegawa, T. (2009). Interactions of elevated [CO2] and night temperature on rice growth and yield. Agric. For. Meteorol. 149, 51–58. doi: 10.1016/j.agrformet.2008.07.006
Chi, Y., Ming, X., Shen, R., and Wan, S. (2013a). Acclimation of leaf dark respiration to nocturnal and diurnal warming in a semiarid temperate steppe. Funct. Plant Biol. 40, 1159–1167. doi: 10.1071/FP12369
Chi, Y., Xu, M., Shen, R., Yang, Q., Huang, B., and Wan, S. (2013b). Acclimation of foliar respiration and photosynthesis in response to experimental warming in a temperate steppe in northern China. PLoS ONE 8:e56482. doi: 10.1371/journal.pone.0056482
Christensen, J. H., Hewitson, B., Busuioc, A., Chen, A., Gao, X., Held, R., et al. (2007). “Regional climate projections,” in Climate Change, 2007: The Physical Science Basis. Contribution of Working group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, eds S. D. Solomon, M. Qin, Z. Manning, M. Chen, K. B. Marquis, M. Averyt et al. (Cambridge: Cambridge University Press), 847–940.
Climent, J., Chambel, M. R., Pardos, M., Lario, F., and Villar-Salvador, P. (2011). Biomass allocation and foliage heteroblasty in hard pine species respond differentially to reduction in rooting volume. Eur. J. For. Res. 130, 841–850. doi: 10.1007/s10342-010-0476-y
Cockshull, K. E. (1979). Effects of irradiance and temperature on flowering of chrysanthemum morifolium ramat in continuous light. Ann. Bot. 44, 451–460.
Cutler, J. M., Steponkus, P. L., Wach, M. J., and Shahan, K. W. (1980). Dynamic aspects and enhancement of leaf elongation in rice. Plant Physiol. 66, 147–152. doi: 10.1104/pp.66.1.147
Damian, J. A., and Donald, R. O. (2001). Impact of chilling temperatures on photosynthesis in warm climate plants. Trends Plant Sci 6, 36–42. doi: 10.1016/S1360-1385(00)01808-2
Darnell, R. L., Cruz-Huerta, N., and Williamson, J. G. (2013). Night temperature and source-sink effects on growth, leaf carbon exchange rate, and carbohydrate accumulation in bell pepper ovaries. J. Am. Soc. Hortic. Sci. 138, 331–337.
Davies, L. J., Brooking, I. R., Catley, J. L., and Halligan, E. A. (2002). Effects of day/night temperature differential and irradiance on the flower stem quality of Sandersonia aurantiaca. Sci. Hortic. 95, 85–98. doi: 10.1016/S0304-4238(02)00026-2
Dejong, J., and Smeets, L. (1982). Effect of day and night temperatures during long photoperiods on the vegetative growth and flowering of Chrysanthemum-morifolium Ramat. Sci. Hortic. 17, 271–275. doi: 10.1016/0304-4238(82)90050-4
Dongsansuk, A., Lütz, C., and Neuner, G. (2013). Effects of temperature and irradiance on quantum yield of PSII photochemistry and xanthophyll cycle in a tropical and a temperate species. Photosynthetica 51, 13–21. doi: 10.1007/s11099-012-0070-2
Fang, S., Ren, S., and Tan, K. (2013). Responses of winter wheat to higher night temperature in spring as compared within whole growth period by controlled experiments in North China. J. Food Agric. Environ. 11, 777–781.
Farquhar, G. D., and Sharkey, T. D. (1982). Stomatal conductance and photosynthesis. Ann. Rev. Plant Physio. 33, 317–345. doi: 10.1146/annurev.pp.33.060182.001533
Flexas, J., and Osmond, C. B. (1999). Analysis of the relative increase in photosynthetic O2 uptake when photosynthesis in grapevine leaves is inhibited following low night temperatures and/or water stress. Plant Physiol. 121, 675–684. doi: 10.1104/pp.121.2.675
Frantz, J. M., Cometti, N. N., and Bugbee, B. (2004). Night temperature has a minimal effect on respiration and growth in rapidly growing plants. Ann. Bot. 94, 155–166. doi: 10.1093/aob/mch122
Friend, D. J. C. (1981). Effect of night temperature on flowering and fruit size in pineapple (Ananas comosus [L.] Merrill). Bot. Gaz. 142, 188–190. doi: 10.1086/337211
Gimenez, D. O., and Rumi, C. P. (1988). Interaction of night temperature and photoperiod on Bromus unioloides H.B.K. aerial biomass components. Environ. Exp. Bot. 28, 1–7. doi: 10.1016/0098-8472(88)90039-1
Guo, Y. P., Zhou, H. F., and Zhang, L. C. (2006). Photosynthetic characteristics and protective mechanisms against photooxidation during high temperature stress in two citrus species. Sci. Hortic. 108, 260–267. doi: 10.1016/j.scienta.2006.01.029
Guy, C. L., Huber, J. L., and Huber, S. C. (1992). Sucrose phosphate synthase and sucrose accumulation at low temperature. Plant Physiol. 100, 502–508. doi: 10.1104/pp.100.1.502
Guy, C. L., Yelenosky, G., and Sweet, H. C. (1980). Light exposure and soluble sugars in citrus frost hardiness. Fla. Sci. 43, 268–273.
Hansen, L. D., Thomas, N. R., and Arnholdt-Schmitt, B. (2009). Temperature responses of substrate carbon conversion efficiencies and growth rates of plant tissues. Physiol. Plant. 137, 446–458. doi: 10.1111/j.1399-3054.2009.01287.x
Hare, P. D., Cress, W. A., and Staden, J. V. (1998). Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 21, 535–553. doi: 10.1046/j.1365-3040.1998.00309.x
Hasegawa, T., Kuwagata, T., Nishimori, M., Ishigooka, Y., Murakami, M., Yoshimoto, M., et al. (2009). “Recent warming trends and rice growth and yield in Japan,” in Proceedings of the MARCO Symposium on Crop Production under Heat Stress: Monitoring, Impact Assessment and Adaptation, eds T. Hasegawa and H. Sakai (Tsukuba: National Institute for Agro-Environmental Studies).
Hedges, L. V., Gurevitch, J., and Curtis, P. S. (1999). The meta-analysis of response ratios in experimental ecology. Ecology 80, 1150–1156. doi: 10.1890/0012-9658(1999)080[1150:TMAORR]2.0.CO;2
Hewitt, J. D., Casey, L. L., and Zobel, R. W. (1985). Effect of day length and night temperature on starch accumulation and degradation in soybean. Ann. Bot. 56, 513–522.
Huang, M., and Guo, Z. (2005). Responses of antioxidative system to chilling stress in two rice cultivars differing in sensitivity. Biol. Plant. 27, 768–779.
Huang, W., Zhang, S. B., and Cao, K. F. (2010). The different effects of chilling stress under moderate light intensity on photosystem II compared with photosystem I and subsequent recovery in tropical tree species. Photosynth. Res. 103, 175–182. doi: 10.1007/s11120-010-9539-7
Hussey, G. (1965). Growth and development in the young tomato: III. The effect of night and day temperatures on vegetative growth. J. Exp. Bot. 16, 373–385. doi: 10.1093/jxb/16.3.373
Ibrahim, M. A., Mäenpää, M., Hassinen, V., Kontunen-Soppela, S., Malec, L., Rousi, M., et al. (2010). Elevation of night-time temperature increases terpenoid emissions from Betula pendula and Populus tremula. J. Exp. Bot. 61, 1583–1595. doi: 10.1093/jxb/erq034
IPCC (2001). “Climate change 2001: the scientific basis,” in Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change, eds J. T. Houghton, Y. Ding, D. J. Griggs, M. Noguer, P. J. van der Linden, X. Dai, et al. (Cambridge: Cambridge University Press).
IPCC (2013). “Climate change 2013: the physical science basis,” in Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds T. F. Stocker, D. Qin, G. K. Plattner, M. Tignor, S. K. Allen, J. Boschung, et al. (Cambridge: Cambridge University Press).
Jagadish, S. V., Craufurd, P. Q., and Wheeler, T. R. (2007). High temperature stress and spikelet fertility in rice (Oryza sativa L.). J. Exp. Bot. 58, 1627–1635. doi: 10.1093/jxb/erm003
Jagadish, S. V., Craufurd, P. Q., and Wheeler, T. R. (2008). Phenotyping parents of mapping populations of rice for heat tolerance during anthesis. Crop Sci. 48, 1140–1146. doi: 10.2135/cropsci2007.10.0559
Jagadish, S. V., Murty, M., and Quick, W. (2015). Rice responses to rising temperatures–challenges, perspectives and future directions. Plant Cell Environ. 38, 1686–1698. doi: 10.1111/pce.12430
Jagadish, S. V., Muthurajan, R., Oane, R., Wheeler, T. R., Heuer, S., Bennett, J., et al. (2010). Physiological and proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.). J. Exp. Bot. 61, 143–156. doi: 10.1093/jxb/erp289
Jones, C. A., Kiniry, J. R., and Dyke, P. (1986). CERES-Maize: A Simulation Model of Maize Growth and Development. College Station, TX: Texas A&M University Press.
Jones, P. D., New, M., Parker, D. E., Martin, S., and Rigor, I. G. (1999). Surface air temperature and its changes over the past 150 years. Rev. Geophys. 37, 173–199. doi: 10.1029/1999RG900002
Jones, T. L., and Ort, D. R. (1998). Chilling delays circadian pattern of sucrose phosphate synthase and nitrate reductase activity in tomato. Plant Physiol. 118, 149–158. doi: 10.1104/pp.118.1.149
Kanno, K., Mae, T., and Makino, A. (2009). High night temperature stimulates photosynthesis, biomass production and growth during the vegetative stage of rice plants. Soil Sci. Plant Nutr. 55, 124–131. doi: 10.1111/j.1747-0765.2008.00343.x
Kanno, K., and Makino, A. (2010). Increased grain yield and biomass allocation in rice under cool night temperature. Soil Sci. Plant Nutr. 56, 412–417. doi: 10.1111/j.1747-0765.2010.00473.x
Kasuga, M., Liu, Q., Miura, S., Yamaguchi-Shinozaki, K., and Shinozaki, K. (1999). Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 17, 287–291. doi: 10.1038/7036
Kaurin, A., Junttila, O., and Hanson, J. (1981). Seasonal changes in frost hardiness in cloudberry (Rubus chamaemorus) in relation to carbohydrate content with special reference to sucrose. Physiol. Plant. 52, 310–314. doi: 10.1111/j.1399-3054.1981.tb08512.x
Khayat, E., Ravad, D., and Zieslin, N. (1985). The effects of various night-temperature regimes on the vegetative growth and fruit production of tomato plants. Sci. Hortic. 27, 9–13. doi: 10.1016/0304-4238(85)90049-4
Kjaer, K. H., Thorupkristensen, K., Rosenqvist, E., and Aaslyng, J. M. (2010). Leaf carbohydrate contents and growth of Chrysanthemum morifolium in response to low night temperatures and high CO2. Eur. J. Hortic. Sci. 275, 118–122.
Kjær, K. H., Hansson, I. M., Thorup-Kristensen, K., Rosenqvist, E., and Aaslyng, J. M. (2008). Root-zone heating at a night air temperature of 8°C does not decrease starch accumulation in Chrysanthemum morifolium. J. Hortic. Sci. Biotechnol. 83, 381–387. doi: 10.1080/14620316.2008.11512395
Kjær, K. H., Thorupkristensen, K., Rosenqvist, E., and Mazanti, J. (2007). Low night temperatures change whole-plant physiology and increase starch accumulation in Chrysanthemum morifolium. J. Hortic. Sci. Biotechnol. 82, 1452–1455. doi: 10.1080/14620316.2007.11512320
Königer, M., and Winter, K. (1993). Growth and photosynthesis of Gossypium hirsutum L. at high photon flux densities: effects of soil temperatures and nocturnal air temperatures. Agronomie 13, 423–431. doi: 10.1051/agro:19930507
Koscielniak, J. (1993). Effect of low night temperatures on photosynthetic activity of the maize seedlings (Zea-mays l). J. Agron. Crop Sci. 171, 73–81. doi: 10.1111/j.1439-037X.1993.tb00116.x
Lang, P., Zhang, C. K., Ebel, R. C., Dane, F., and Dozier, W. A. (2005). Identification of cold acclimated genes in leaves of Citrus unshiu by mRNA differential display. Gene 359, 111–118. doi: 10.1016/j.gene.2005.06.013
Lee, C. C., Bilderback, T. E., and Thomas, J. F. (1991). Growth responses of Heptacodium miconioides to various photoperiods and day/night temperatures. J. Am. Soc. Hortic. Sci. 116, 339–399.
Lee, C. H., and Myeongwhan, C. (2011). Control of unseasonable flowering in Chrysanthemum ‘Baekma’ by 2-chloroethylphosphonic acid and night temperature. Korean J. Hortic. Sci. 29, 539–548.
Lepage, I., Jong, J. D., and Smeets, L. (1984). Effect of day and night temperatures during short photoperiods on growth and flowering of Chrysanthemum morifolium Ramat. Sci. Hortic. 22, 373–381. doi: 10.1016/S0304-4238(84)80009-6
Li, C. Y., Peng, C. H., Zhao, Q. B., Xie, P., and Chen, W. (2004). Characteristic analysis of the abnormal high temperarure in 2003’s midsummer in Wuhan city. J. Central China Normal Univ. 38, 379–382.
Lin, D. L., Xia, J. Y., and Wan, S. Q. (2010). Climate warming and biomass accumulation of terrestrial plants: a meta-analysis. New Phytol. 188, 187–198. doi: 10.1111/j.1469-8137.2010.03347.x
Lin, L. Q., Li, L., Bi, C., Zhang, Y. L., Wan, S. B., Meng, J. J., et al. (2011). Damaging mechanisms of chilling- and salt stress to Arachis hypogaea L. leaves. Photosynthetica 49, 37–42. doi: 10.1007/s11099-011-0005-3
Liu, Y., Li, T., Qi, H., Li, J., and Yin, X. (2010). Effects of grafting on carbohydrate accumulation and sugar-metabolic enzyme activities in muskmelon. Afr. J. Biotechnol. 9, 025–035.
Liu, Y. F., Li, T. L., Xu, T., Qi, M. F., Xu, C. Q., and Qi, H. Y. (2011). Effect of low night temperature treatment and recovery on photosynthesis and the allocation of absorbed light energy in tomato (Lycopersicon esculentum Mill.) leaves. J. Hortic. Sci. Biotechnol. 86, 91–96. doi: 10.1080/14620316.2011.11512731
Liu, Y. F., Qi, M. F., and Li, T. L. (2012). Photosynthesis, photoinhibition, and antioxidant system in tomato leaves stressed by low night temperature and their subsequent recovery. Plant Sci. 196, 8–17. doi: 10.1016/j.plantsci.2012.07.005
Llorens, L., Peñuelas, J., and Estiarte, M. (2003). Ecophysiological responses of two Mediterranean shrubs, Erica multiflora and Globularia alypum, to experimentally drier and warmer conditions. Physiol. Plant. 119, 231–243. doi: 10.1034/j.1399-3054.2003.00174.x
Loh, F. C. W., Grabosky, J. C., and Bassuk, N. L. (2003). Growth response of Ficus benjamina to limited soil volume and soil dilution in a skeletal soil container study. Urban For. Urban Gree. 2, 53–62. doi: 10.1078/1618-8667-00023
Loka, D. A., and Oosterhuis, D. M. (2010). Effect of high night temperatures on cotton respiration, ATP levels and carbohydrate content. Environ. Exp. Bot. 68, 258–263. doi: 10.1016/j.envexpbot.2010.01.006
Long, S. (1998). “Ecology of C4 photosynthesis: environmental responses,” in C4 Plant Biology, eds R. R. Sage and R. K. Monson (New York, NY: Academic Press).
Long, S. P. (1983). C4 photosynthesis at low temperatures. Plant Cell Environ. 6, 345–363. doi: 10.1111/1365-3040.ep11612141
Lu, Z., Percy, R. G., Qualset, C. O., and Zeiger, E. (1998). Stomatal conductance predicts yields in irrigated Pima cotton and bread wheat grown at high temperatures. J. Exp. Bot. 49, 453–460. doi: 10.1093/jexbot/49.suppl_1.453
Lucidos, J. G., Ryu, K. B., Younis, A., Kim, C. K., Hwang, Y. J., Son, B. G., et al. (2013). Different day and night temperature responses in Lilium hansonii in relation to growth and flower development. Hort. Environ. Biotechnol. 54, 405–411. doi: 10.1007/s13580-013-1241-1
Maali, A. R., and Heidarvand, L. (2010). What happens in plant molecular responses to cold stress? Acta Physiol. Plant. 32, 419–431. doi: 10.1007/s11738-009-0451-8
Mae, T. (1997). Physiological nitrogen efficiency in rice: nitrogen utilization, photosynthesis, and yield potential. Plant Soil 196, 201–210. doi: 10.1023/A:1004293706242
Malek, A. A., Blazich, F. A., Warren, S. L., and Shelton, J. E. (1992). Initial growth of seedlings of Flame azalea in response to day/night temperature. J. Am. Soc. Hortic. Sci. 117, 216–219.
Manunta, P., and Kirkham, M. B. (1996). Respiration and growth of sorghum and sunflower under predicted increased night temperatures1. J. Agron. Crop Sci. 176, 267–274. doi: 10.1111/j.1439-037X.1996.tb00471.x
Meng, F. Z., Hu, L. P., Wang, S. H., Sui, X. L., Wei, L., Wei, Y. X., et al. (2008). Effects of exogenous abscisic acid (ABA) on cucumber seedling leaf carbohydrate metabolism under low temperature. Plant Growth Regul. 56, 233–244. doi: 10.1007/s10725-008-9303-6
Miao, M., Zhang, Z., Xu, X., Wang, K., Cheng, H., and Cao, B. (2009). Different mechanisms to obtain higher fruit growth rate in two cold-tolerant cucumber (Cucumis sativus L.) lines under low night temperature. Sci. Hortic. 119, 357–361. doi: 10.1016/j.scienta.2008.08.028
Mohammed, A. R., Cothren, J. T., and Tarpley, L. (2013). High night temperature and abscisic acid affect rice productivity through altered photosynthesis, respiration and spikelet fertility. Crop Sci. 53, 2603–2612. doi: 10.2135/cropsci2013.01.0060
Mohammed, A. R., and Tarpley, L. (2009a). High nighttime temperatures affect rice productivity through altered pollen germination and spikelet fertility. Agric. Forest Meteorol. 149, 999–1008. doi: 10.1016/j.agrformet.2008.12.003
Mohammed, A. R., and Tarpley, L. (2009b). Impact of high nighttime temperature on respiration, membrane stability, antioxidant capacity, and yield of rice plants. Crop Sci. 138, 398–399.
Mohammed, A. R., and Tarpley, L. (2010). Effects of high night temperature and spikelet position on yield-related parameters of rice (Oryza sativa L.) plants. Eur. J. Agron. 33, 117–123. doi: 10.1016/j.eja.2009.11.006
Morita, S. (2005). Grain growth and endosperm cell size under high night temperatures in rice (Oryza sativa L.). Ann. Bot. 95, 695–701. doi: 10.1093/aob/mci071
Morita, S., Shiratsuchi, H., Takanashi, J. I., and Fujita, K. (2002). Effect of high temperature on ripening in rice plants: comparison of the effects of high night temperatures and high day temperatures (Crop Physiology and Cell Biology). Jpn. J. Crop Sci. 71, 102–109. doi: 10.1626/jcs.71.102
Morita, S., Shiratsuchi, H., Takahashi, J. I., and Fujita, K. (2004). Effect of high temperature on grain ripening in rice plants – Analysis of the effects of high night and high day temperatures applied to the panicle and other parts of the plant. Jpn. J. Crop Sci. 73, 77–83. doi: 10.1626/jcs.73.77
Mortensen, L. M. (1994). Effects of day/night temperature variations on growth, morphogenesis and flowering of Kalanchoe blossfeldiana v. Poelln at different CO2 concentrations, day lengths and photon flux densities. Sci. Hortic. 59, 233–241. doi: 10.1016/0304-4238(94)90016-7
Mortensen, L. M., and Moe, R. (1992). Effects of CO2 enrichment and different day/night temperature combinations on growth and flowering of Rosa L. and Kalanchoe blossfeldiana v. Poelln. Sci. Hortic. 51, 145–153. doi: 10.1016/0304-4238(92)90113-Q
Nagarajan, S., Jagadish, S. V. K., Prasad, A. S. H., Thomar, A. K., Anand, A., Pal, M., et al. (2010). Local climate affects growth, yield and grain quality of aromatic and non-aromatic rice in northwestern India. Agric. Ecosyst. Environ. 138, 274–281. doi: 10.1016/j.agee.2010.05.012
Narayanan, S., Prasad, P. V. V., Fritz, A. K., Boyle, D. L., and Gill, B. S. (2015). Impact of high night-time and high daytime temperature stress on winter wheat. J. Agron. Crop Sci. 201, 206–218. doi: 10.1111/jac.12101
Neales, T. F., Sale, P. J. M., and Meyer, C. P. (1980). Carbon dioxide assimilation by pineapple plants, Ananas comosus (L.) Merr. II. Effects of variation of the day/night temperature regime. Funct. Plant Biol. 7, 375–385.
Noctor, G., and Foyer, C. H. (1998). Ascorbate and glutathione: keeping active oxygen under control. Annu. Rev. Plant Biol. 49, 249–279. doi: 10.1146/annurev.arplant.49.1.249
Oufir, M., Legay, S., Nicot, N., Moer, K. V., Hoffmann, L., Renaut, J., et al. (2008). Gene expression in potato during cold exposure: changes in carbohydrate and polyamine metabolisms. Plant Sci. 175, 839–852. doi: 10.1016/j.plantsci.2008.08.010
Pantin, F., Simonneau, T., Rolland, G., Dauzat, M., and Muller, B. (2011). Control of leaf expansion: a developmental switch from metabolics to hydraulics. Plant Physiol. 156, 803–815. doi: 10.1104/pp.111.176289
Papadopoulos, A. P., and Hao, X. M. (2000). Effects of day and night air temperature on growth, productivity and energy use of long English cucumber. Can. J. Plant Sci. 80, 143–150. doi: 10.4141/P99-021
Patil, R. H., Laegdsmand, M., Olesen, J. E., and Porter, J. R. (2010). Effect of soil warming and rainfall patterns on soil N cycling in Northern Europe. Agric. Ecosyst. Environ. 139, 195–205. doi: 10.1016/j.agee.2010.08.002
Patterson, D. T. (1990). Effects of day and night temperature on vegetative growth of Texas panicum (Panicum texanum). Weed Sci. 38, 365–373.
Patterson, D. T. (1993). Effects of day and night temperature on Goatsrue (Galega officinalis) and Alfalfa (Medicago sativa) growth. Weed Sci. 41, 38–45.
Peet, M. M., and Bartholemew, M. (1996). Effect of night temperature on pollen characteristics, growth, and fruit set in tomato. J. Am. Soc. Hortic. Sci. 121, 514–519.
Peng, S., Huang, J., Sheehy, J. E., Laza, R. C., Visperas, R. M., Zhong, X., et al. (2004). Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. U.S.A. 101, 9971–9975. doi: 10.1073/pnas.0403720101
Peraudeau, S., Roques, S., Quiñones, C. O., Fabre, D., Rie, J. V., Ouwerkerk, P. B. F., et al. (2015). Increase in night temperature in rice enhances respiration rate without significant impact on biomass accumulation. Field Crops Res. 171, 67–78. doi: 10.1016/j.fcr.2014.11.004
Pollet, B., Vanhaecke, L., Dambre, P., Lootens, P., and Steppe, K. (2011). Low night temperature acclimation of Phalaenopsis. Plant Cell Rep. 30, 1125–1134. doi: 10.1007/s00299-011-1021-2
Prasad, P. V. V., Boote, K. J., Allen, L. H., Sheehy, J. E., and Thomas, J. M. G. (2006). Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res. 95, 398–411. doi: 10.1016/j.fcr.2005.04.008
Prasad, P. V. V., and Djanaguiraman, M. (2011). High night temperature decreases leaf photosynthesis and pollen function in grain sorghum. Funct. Plant Biol. 38, 993–1003. doi: 10.1071/FP11035
Prasad, P. V. V., Pisipati, S. R., Ristic, Z., Bukovnik, U., and Fritz, A. K. (2008). Impact of nighttime temperature on physiology and growth of spring wheat. Crop Sci. 48, 2372–2380. doi: 10.2135/cropsci2007.12.0717
Pressman, E., Shaked, R., and Firon, N. (2006). Exposing pepper plants to high day temperatures prevents the adverse low night temperature symptoms. Physiol. Plant. 126, 618–626.
Prieto, P., Peñuelas, J., Llusià, J., Asensio, D., and Estiarte, M. (2009). Effects of long-term experimental night-time warming and drought on photosynthesis, Fv/Fm and stomatal conductance in the dominant species of a Mediterranean shrubland. Acta Physiol. Plant. 31, 729–739. doi: 10.1007/s11738-009-0285-4
Qi, H., Hua, L., Zhao, L., and Zhou, L. (2011). Carbohydrate metabolism in tomato (Lycopersicon esculentum Mill.) seedlings and yield and fruit quality as affected by low night temperature and subsequent recovery. Afr. J. Biotechnol. 10, 5743–5749.
Rao, B. B., Chowdary, P. S., Sandeep, V. M., Rao, V. U. M., and Venkateswarlu, B. (2014). Rising minimum temperature trends over India in recent decades: Implications for agricultural production. Glob. Planet. Change 117, 1–8. doi: 10.1016/j.gloplacha.2014.03.001
Rapacz, M. (1998). The effects of day and night temperatures during early growth of winter oilseed rape (Brassica napus L. var. oleifera cv. Górczañski) seedlings on their morphology and cold acclimation responses. Acta Physiol. Plant. 20, 67–72. doi: 10.1007/s11738-998-0045-x
Rehmani, M. I. A., Wei, G., Hussain, N., Ding, C., Li, G., Liu, Z., et al. (2014). Yield and quality responses of two indica rice hybrids to post-anthesis asymmetric day and night open-field warming in lower reaches of Yangtze River delta. Field Crops Res. 156, 231–241. doi: 10.1016/j.fcr.2013.09.019
Reynolds, M. P., Balota, M., Delgado, M., Amani, I., and Fischer, R. A. (1994). Physiological and morphological traits associated with spring wheat yield under hot, irrigated conditions. Funct. Plant Biol. 21, 717–730.
Ritchie, J. R., and Otter, S. (1985). Description and Performance of CERES – Wheat: A User-Oriented Wheat Yield Model. Washington, DC: Agricultural Research Service.
Rosenberg, M. S. (2005). The file-drawer problem revisited: a general weighted method for calculating fail-safe numbers in meta-analysis. Evolution 59, 464–486. doi: 10.1111/j.0014-3820.2005.tb01004.x
Rosenthal, R. (1979). The file drawer problem and tolerance for null results. Psychol. Bull. 86, 638–641. doi: 10.1037/0033-2909.86.3.638
Rustad, L., Campbell, J., Marion, G., Norby, R., Mitchell, M., Hartley, A., et al. (2001). A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 126, 543–562. doi: 10.1007/s004420000544
Rustad, L. E. (2008). The response of terrestrial ecosystems to global climate change: towards an integrated approach. Sci. Total Environ. 404, 222–235. doi: 10.1016/j.scitotenv.2008.04.050
Sage, R. F. (2002). Variation in the kcat of Rubisco in C3 and C4 plants and some implications for photosynthetic performance at high and low temperature. J. Exp. Bot. 53, 609–620. doi: 10.1093/jexbot/53.369.609
Sage, R. F., and Kubien, D. S. (2007). The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 30, 1086–1106. doi: 10.1111/j.1365-3040.2007.01682.x
Sao, P. M. D. F., Machado, E. C., Machado, R. S., and Ribeiro, R. V. (2010). Effects of low night temperature and rootstocks on diurnal variation of leaf gas exchange rates and photochemical activity of ‘valencia’ sweet orange plants. Rev. Bras. Frutic. 32, 351–359.
Sao, P. M. D. F., Magalhaes, A. L. A. M., Ribeiro, R. V., Ribeiro Marchiori, P. E., Machado, R. S., and Machado, E. C. (2013a). Low night temperature and water deficit on photosynthesis of sugarcane. Pesqui. Agropecu. Bras. 48, 487–495.
Sao, P. M. D. F., Ribeiro, R. V., da Silveira, J. A., Magalhaes Filho, J. R., and Machado, E. C. (2013b). Rootstocks induce contrasting photosynthetic responses of orange plants to low night temperature without affecting the antioxidant metabolism. Theor. Exp. Plant Physiol. 25, 26–35. doi: 10.1590/S2197-00252013000100004
Sardans, J., Peñuelas, J., Prieto, P., and Estiarte, M. (2008). Drought and warming induced changes in P and K concentration and accumulation in plant biomass and soil in a Mediterranean shrubland. Plant Soil 306, 261–271. doi: 10.1007/s11104-008-9583-7
Saveyn, A., Steppe, K., McGuire, M. A., Lemeur, R., and Teskey, R. O. (2008). Stem respiration and carbon dioxide efflux of young Populus deltoides trees in relation to temperature and xylem carbon dioxide concentration. Oecologia 154, 637–649. doi: 10.1007/s00442-007-0868-y
Schoppach, R., and Sadok, W. (2013). Transpiration sensitivities to evaporative demand and leaf areas vary with night and day warming regimes among wheat genotypes. Funct. Plant Biol. 40, 708–718. doi: 10.1071/FP13028
Schurr, U., Heckenberger, U., Herdel, K., Walter, A., and Feil, R. (2000). Leaf development in Ricinus communis during drought stress: dynamics of growth processes, of cellular structure and of sink–source transition. J. Exp. Bot. 51, 1515–1529. doi: 10.1093/jexbot/51.350.1515
Searle, S. Y., Turnbull, M. H., Boelman, N. T., Schuster, W. S., Yakir, D., and Griffin, K. L. (2012). Urban environment of New York City promotes growth in northern Red oak seedlings. Tree Physiol. 32, 389–400. doi: 10.1093/treephys/tps027
Seddigh, M., and Jolliff, G. D. (1984a). Effects of night temperature on dry-matter partitioning and seed growth of indeterminate field-grown soybean. Crop Sci. 24, 704–710. doi: 10.2135/cropsci1984.0011183X002400040019x
Seddigh, M., and Jolliff, G. D. (1984b). Night temperature effects on morphology, phenology, yield and yield components of indeterminate field-grown soybean. Agron. J. 76, 824–828. doi: 10.2134/agronj1984.00021962007600050026x
Seddigh, M., and Jolliff, G. D. (1984c). Physiological-responses of field-grown soybean leaves to increased reproductive load induced by elevated night temperature. Crop Sci. 24, 952–957. doi: 10.2135/cropsci1984.0011183X002400050031x
Serra, G., and Carrai, C. (1988). Effect of prolonged day temperature in night-time and different levels of night temperature on growth of Cordyline “lord robertson”. Acta Hortic. 229, 393–398. doi: 10.17660/ActaHortic.1988.229.44
Shah, F., Huang, J., Cui, K., Nie, L., Shah, T., Wu, W., et al. (2011). Physiological and biochemical changes in rice associated with high night temperature stress and their amelioration by exogenous application of ascorbic acid (vitamin C). Aust. J. Crop Sci. 5, 1810.
Shi, W., Muthurajan, R., Rahman, H., Selvam, J., Peng, S., Zou, Y., et al. (2013). Source-sink dynamics and proteomic reprogramming under elevated night temperature and their impact on rice yield and grain quality. New Phytol. 197, 825–837. doi: 10.1111/nph.12088
Sillmann, J., Kharin, V. V., Zwiers, F. W., Zhang, X., and Bronaugh, D. (2013). Climate extremes indices in the CMIP5 multimodel ensemble: Part 2. Future climate projections. J. Geophys. Res. Atmos. 118, 2473–2493. doi: 10.1002/jgrd.50188
Siminovitch, D., and Briggs, D. R. (1953). Studies on the chemistry of the living bark of the Black Locust in relation to its frost hardiness. V. Seasonal transformations and variations in the carbohydrates: starch-sucrose interconversions. Plant Physiol. 28, 383–400. doi: 10.1104/pp.28.3.383
Starrett, M. C., Blazich, F. A., Warren, S. L., Starrett, M. C., Blazich, F. A., and Warren, S. L. (1993). Initial growth of Rosebay Rhododendron seedlings as influenced by day and night temperatures. HortScience 28, 705–707.
Steponkus, P. L., and Lanphear, F. (1968). The relationship of carbohydrates to cold acclimation of Hedera helix L. cv. Thorndale. Physiol. Plant. 21, 777–791. doi: 10.1111/j.1399-3054.1968.tb07301.x
Stockle, C. O., Martin, S. A., and Campbell, G. S. (1994). CropSyst, a cropping systems simulation model: Water/nitrogen budgets and crop yield. Agric. Syst. 46, 335–359. doi: 10.1016/0308-521X(94)90006-2
Suding, K. N., Collins, S. L., Gough, L., Clark, C., Cleland, E. E., Gross, K. L., et al. (2005). Functional-and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Natl. Acad. Sci. U.S.A. 102, 4387–4392. doi: 10.1073/pnas.0408648102
Suzuki, S., and Moroyu, H. (1962). Effect of high-night-temperature treatment on the growth of rice plants with special reference to its nutritional condition. Bull. Chugoku Agric. Exp. Stn. 8, 269–290.
Teragishi, A., Kanbara, Y., and Ono, H. (2001). The effects of different night temperatures and the amount of hydroponic solution on the growth and fruiting of own-rooted Fig cuttings. Engei Gakkai Zasshi 70, 1–6. doi: 10.2503/jjshs.70.1
Thomas, R. B., and Strain, B. R. (1991). Root restriction as a factor in photosynthetic acclimation of cotton seedlings grown in elevated carbon dioxide. Plant Physiol. 96, 627–634. doi: 10.1104/pp.96.2.627
Tsunoda, K. (1964). Studies on the effects of water-temperature on the growth and yield in rice plants. Bull. Natl. Inst. Agric. Sci. A 11, 75–174.
Turnbull, M. H., Murthy, R., and Griffin, K. L. (2002). The relative impacts of daytime and night-time warming on photosynthetic capacity in Populus deltoides. Plant Cell Environ. 25, 1729–1737. doi: 10.1046/j.1365-3040.2002.00947.x
Turner, A. D., and Ewing, E. E. (1988). Effects of photoperiod, night temperature, and irradiance on flower production in the potato. Potato Res. 31, 257–268. doi: 10.1007/BF02365534
Uehara, N., Sasaki, H., Aoki, N., and Ohsugi, R. (2009). Effects of the temperature lowered in the daytime and night-time on sugar accumulation in sugarcane. Plant Prod. Sci. 12, 420–427. doi: 10.1626/pps.12.420
Veatch, B. M. E., Ray, D. T., and Gehrels, A. (2007). Night temperature, rubber production, and carbon exchange in guayule. Ind. Crop Prod. 25, 34–43. doi: 10.1016/j.indcrop.2006.06.019
Verheul, M. J., Sønsteby, A., and Grimstad, S. O. (2007). Influences of day and night temperatures on flowering of Fragaria x ananassa Duch., cvs. Korona and Elsanta, at different photoperiods. Sci. Hortic. 112, 200–206. doi: 10.1016/j.scienta.2006.12.022
Volder, A., Edwards, E. J., Evans, J. R., Robertson, B. C., Schortemeyer, M., and Gifford, R. M. (2004). Does greater night-time, rather than constant, warming alter growth of managed pasture under under ambient and elevated atmospheric CO2? New Phytol. 162, 397–411. doi: 10.1111/j.1469-8137.2004.01025.x
Vose, R. S., Easterling, D. R., and Gleason, B. (2005). Maximum and minimum temperature trends for the globe: an update through 2004. Geophys. Res. Lett. 32: L23822. doi: 10.1029/2005GL024379
Wahid, A., Gelani, S., Ashraf, M., and Foolad, M. R. (2007). Heat tolerance in plants: an overview. Environ. Exp. Bot. 61, 199–223. doi: 10.1016/j.envexpbot.2007.05.011
Walters, M. B., and Reich, P. B. (1989). Response of Ulmus americana seedlings to varying nitrogen and water status. 1 Photosynthesis and growth. Tree Physiol. 5, 159–172. doi: 10.1093/treephys/5.2.159
Wang, D., Heckathorn, S. A., Wang, X., and Philpott, S. M. (2012). A meta-analysis of plant physiological and growth responses to temperature and elevated CO2. Oecologia 169, 1–13. doi: 10.1007/s00442-011-2172-0
Wang, Y. T. (2007). Average daily temperature and reversed day/night temperature regulate vegetative and reproductive responses of a Doritis pulcherrima Lindley hybrid. HortScience 42, 68–70.
Way, D. A., and Oren, R. (2010). Differential responses to changes in growth temperature between trees from different functional groups and biomes: a review and synthesis of data. Tree Physiol. 30, 669–688. doi: 10.1093/treephys/tpq015
Welch, J. R., Vincent, J. R., Auffhammer, M., Moya, P. F., Dobermann, A., and Dawe, D. (2010). Rice yields in tropical/subtropical Asia exhibit large but opposing sensitivities to minimum and maximum temperatures. Proc. Natl. Acad. Sci. U.S.A. 107, 14562–14567. doi: 10.1073/pnas.1001222107
Wolfe-Bellin, K. S., He, J. S., and Bazzaz, F. A. (2006). Leaf-level physiology, biomass, and reproduction of Phytolacca americana under conditions of elevated carbon dioxide and increased nocturnal temperature. Int. J. Plant Sci. 167, 1011–1020. doi: 10.1086/506154
Xia, J., Chen, J., Piao, S., Ciais, P., Luo, Y., and Wan, S. (2014). Terrestrial carbon cycle affected by non-uniform climate warming. Nat. Geosci. 7, 173–180. doi: 10.1038/ngeo2093
Xiong, L., Schumaker, K. S., and Zhu, J.-K. (2002). Cell signaling during cold, drought, and salt stress. Plant Cell 14(Suppl. 1), S165–S183.
Yang, D. H., Yang, X. X., and Liu, C. (2006). Global low temperature, earthquake and tsunami (Dec.26, 2004) in Indonesia. Prog. Geophys. 21, 1023–1027.
Yoshida, S., Satake, T., and Mackill, D. (1981). High-Temperature Stress in Rice [Study Conducted at IRRI, Philippines]. Los Baños: IRRI.
Yu, C. W., Murphy, T. M., Sung, W. W., and Lin, C. H. (2002). H2O2 treatment induces glutathione accumulation and chilling tolerance in mung bean. Funct. Plant Biol. 29, 1081–1087. doi: 10.1071/PP01264
Zhang, G., Liu, Y., Yang, N., Meng, Z., Lu, T., and Li, T. (2014). Exogenous calcium alleviates low night temperature stress on the photosynthetic apparatus of tomato leaves. PLoS ONE 9:97322. doi: 10.1371/journal.pone.0097322
Zhang, L., Hao, X. M., Li, Y. G., and Jiang, G. M. (2010). Response of greenhouse tomato to varied low pre-night temperatures at the same daily integrated temperature. HortScience 45, 1654–1661.
Zhang, Y., Jiang, J., and Yang, Y. (2013). Acetyl salicylic acid induces stress tolerance in tomato plants grown at a low night-time temperature. J. Hortic. Sci. Biotechnol. 88, 490–496. doi: 10.1080/14620316.2013.11512996
Zhang, Y. H., Li, R., and Wang, Y. (2013). Night-time warming affects N and P dynamics and productivity of winter wheat plants. Can. J. Plant Sci. 93, 397–406. doi: 10.4141/cjps2012-044
Zhou, L., Dickinson, R. E., Tian, Y., Fang, J., Li, Q., Kaufmann, R. K., et al. (2004). Evidence for a significant urbanization effect on climate in China. Proc. Natl. Acad. Sci. U.S.A. 101, 9540–9544. doi: 10.1073/pnas.0400357101
Zhu, J., Bartholomew, D. P., and Goldstein, D. (2005). Photosynthetic gas exchange and water relations during drought in ‘Smooth Cayenne’ pineapple (Ananas comosus (L.) Merr.) grown under ambient and elevated CO2 and three day/night temperatures. Acta Hortic. 666, 161–173. doi: 10.17660/ActaHortic.2005.666.15
Zieslin, N., Khayat, E., and Mor, Y. (1986). The response of rose plants to different night-temperature regimes. Acta Hortic. 112, 86–89.
Keywords: high night temperature, low night temperature, photosynthesis, respiration, biomass, plant functional types
Citation: Jing P, Wang D, Zhu C and Chen J (2016) Plant Physiological, Morphological and Yield-Related Responses to Night Temperature Changes across Different Species and Plant Functional Types. Front. Plant Sci. 7:1774. doi: 10.3389/fpls.2016.01774
Received: 14 September 2016; Accepted: 10 November 2016;
Published: 24 November 2016.
Edited by:
Raquel Esteban, University of the Basque Country, SpainReviewed by:
M. Teresa Sanchez-Ballesta, Spanish National Research Council, SpainNobuhiro Suzuki, Sophia University, Japan
Copyright © 2016 Jing, Wang, Zhu and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dan Wang, d2FuZ2Rhbi5udWlzdEBvdXRsb29rLmNvbQ== Jiquan Chen, anFjaGVuQG1zdS5jb20=