 Mathieu Rolland
Mathieu Rolland Aurélie Dupuy
Aurélie Dupuy Aude Pelleray2
Aude Pelleray2 Philippe Delavault
Philippe Delavault- 1GEVES, Beaucouzé, France
- 2Laboratoire de Biologie et Pathologie Végétales, Université de Nantes, Nantes, France
Broomrapes are holoparasitic plants spreading through seeds. Each plant produces hundreds of thousands of seeds which remain viable in the soils for decades. To limit their spread, drastic measures are being taken and the contamination of a commercial seed lot by a single broomrape seed can lead to its rejection. Considering that broomrapes species identification from a single seed is extremely difficult even for trained botanists and that among all the described species, only a few are really noxious for the crops, numerous seed lots are rejected because of the contamination by seeds of non-noxious broomrape species. The aim of this study was to develop and evaluate a High Resolution Melting assay identifying the eight most noxious and common broomrape species (Phelipanche aegyptiaca, Orobanche cernua, O. crenata, O. cumana, O. foetida, O. hederae, O. minor, and P. ramosa) from a single seed. Based on trnL and rbcL plastidial genes amplification, the designed assay successfully identifies O. cumana, O. cernua, O. crenata, O. minor, O. hederae, and O. foetida; P. ramosa, and P. aegyptiaca can be differentiated from other species but not from each other. Tested on 50 seed lots, obtained results perfectly matched identifications performed by sequencing. Through the analysis of common seed lots by different analysts, the reproducibility of the assay was evaluated at 90%. Despite an original sample preparation process it was not possible to extract enough DNA from some seeds (10% of the samples). The described assay fulfills its objectives and allows an accurate identification of the targeted broomrape species. It can be used to identify contaminants in commercial seed lots or for any other purpose. The assay might be extended to vegetative material.
Introduction
Broomrapes (Orobanche and Phelipanche spp.) are angiosperms in the Orobanchaceae which have evolved into obligate root holoparasitic plants (Joel, 2009). Devoid of leaves, of chlorophyll as well as of functional roots, they entirely depend on their host for nutritional requirements (Westwood, 2013). One single broomrape plant can produce hundreds of thousands of extremely small seeds, between 200 and 300 μm, each weighing around 5 μg and composed of only 200 to 300 cells (Joel, 1987). They are easily dispersed mainly by wind and water, and remain viable in the soils for many years until their germination is triggered chemically by exudates released in the soil by the roots of potential host plants (Lopez-Granados and Garcia-Torres, 1996, 1999). At the vicinity of host roots, germinated seeds develop a haustorium that penetrates host tissues and establish a connection with its vascular tissues. This connection will constitute the source of the parasite for water and nutrients. Some species in the Orobanche and Phelipanche genera, the weedy broomrapes, are able to infect a large range of plant species, including many important crops. In severe cases, infection of the host plant can lead in reduction of crop yields up 100%. This makes broomrapes one of the most devastating parasitic weeds in the Mediterranean and western Asian regions but also in many other parts of the world (Parker, 2009). Thus, among these noxious parasitic species are the closely related Phelipanche ramosa L. and P. aegyptiaca Pers. (synonym Orobanche ramosa and O. aegyptiaca) (Joel, 2009), O. cumana Wallr., O. cernua Loefl., O. crenata Forsk., O. foetida Poir., and O. minor Sm., while O. hederae Vaucher ex Duby has no agronomical impact but is extremely common.
Strategies to control parasitic weeds can be classified in chemical, cultural, physical, and biological control methods (Fernández-Aparicio et al., 2016). Among them, breeding for crop resistance seems to be the best approach to manage this issue. However, sources of resistance to most parasitic plants are either scarce or of complex nature (Perez-De-Luque et al., 2009). Despite these difficulties, significant success has been made on some crops. All these approaches allow a control of the parasitic population or permit resistant crops to grow and yield on infested soils, however, the eradication of parasitic weeds remains extremely difficult. Considering that one major mean of field contamination is through contaminated crop seed lots, preventive measures have to be taken to avoid spreading parasite seeds, especially through global scale seed exchanges. This requires to detect efficiently the possible contaminations of crop seeds lots by broomrape seed (Dongo et al., 2012). Visual detection of broomrape seeds in crop seed lots is conducted by sieving and observation of the obtained residues. Characterization of broomrape seeds at the species level in contaminated crop seed lots is important giving the differential host ranges among broomrape weed species and the capacity of some broomrape weeds to thrive in non-parasitic weed species. However, due to their nuanced microscopic morphological features, this identification is extremely difficult and can only can be performed by high qualified specialists (Abu Sbaih and Jury, 1994; Plaza et al., 2004). Molecular tools have been developed to detect and identify broomrape species from soil and crop seed batches. Random amplified polymorphic DNA technique (RAPD) allowed the differentiation between species such as P. aegyptiaca, P. ramosa, O. cernua, O. cumana, and O. crenata (Katzir et al., 1996; Paran et al., 1997). This technique was even used on single seeds (Portnoy et al., 1997), however, the main drawback of RAPDs is their low reproducibility (Harris, 1999). Intersimple sequence repeats (ISSR) were latter used to discriminate closely related species such as O. cumana and O. cernua (Benharrat et al., 2002). A TaqMan assay was developed on internal transcribed spacers (ITS) with the aim of detecting and quantifying P. ramosa and O. cumana seeds in oilseed rape and sunflower seed lots, respectively (Dongo et al., 2012). Microsatellites were also developed to investigate intraspecific variations in O. cumana (Pineda-Martos et al., 2014). Due to its monoparental inheritance, plastid genome has a low intraspecific variability and seems to be an adequate target for species identification. In the case of Orobanche genus, a particular attention was paid to the pseudogene rbcL which showed important sequence divergences among species due to an evolution under purifying selection (Wolfe and dePamphilis, 1997; Benharrat et al., 2000; Manen et al., 2004). Recently, full broomrape plastid genome sequence was made available (Wicke et al., 2013; Cusimano and Wicke, 2015) providing new molecular markers for species identification.
High resolution melting (HRM) is a technique based on the real-time measure of double stranded DNA denaturation at a high resolution. It is suitable for gene scanning and genotyping (Gori et al., 2012) and allows the detection of genetic variations such as single nucleotide polymorphisms (SNP), mutations (Toi and Dwyer, 2008), or methylation (Wojdacz and Dobrovic, 2007). Used on PCR products during a post-PCR denaturation, it requires no tube opening, purification, or product separation. With a minimum manipulation, HRM minimizes the contamination risk, it is cost efficient, suitable for high-throughput, and can be performed in-house by laboratories with no sequencing facility (Reed et al., 2007). This technique has been extensively used on human tissues (Krypuy et al., 2007; Takano et al., 2008), for clinical or phytopathological diagnostic and food analysis (Druml and Cichna-Markl, 2014). It is increasingly used on plant tissues for species and cultivar differentiation (Mackay et al., 2008; Jaakola et al., 2010) or genotyping (Lochlainn et al., 2011).
The objective of this study is to combine the knowledge recently obtained on plastid genome and the HRM technique to develop a new application allowing the differentiation of the seven most noxious and common broomrape weed species (P. aegyptiaca, O. cernua, O. crenata, O. cumana, O. foetida, O. minor, and P. ramosa) and the widely distributed O. hederae species from a single seed. This new application should provide to laboratories, involved in seed certification, a decision-making tool to evaluate crop seed lots potentially contaminated by noxious broomrape species.
Materials and Methods
Plant Material
Broomrape seeds (P. aegyptiaca, O. cernua, O. crenata, O. cumana, O. foetida, O. hederae, O. minor, and P. ramosa) were either obtained from international collections or collected during field sampling by GEVES, Syngenta, Terres Inovia, or University of Nantes. Available data concerning the tested seed lots are summarized in Table 1.
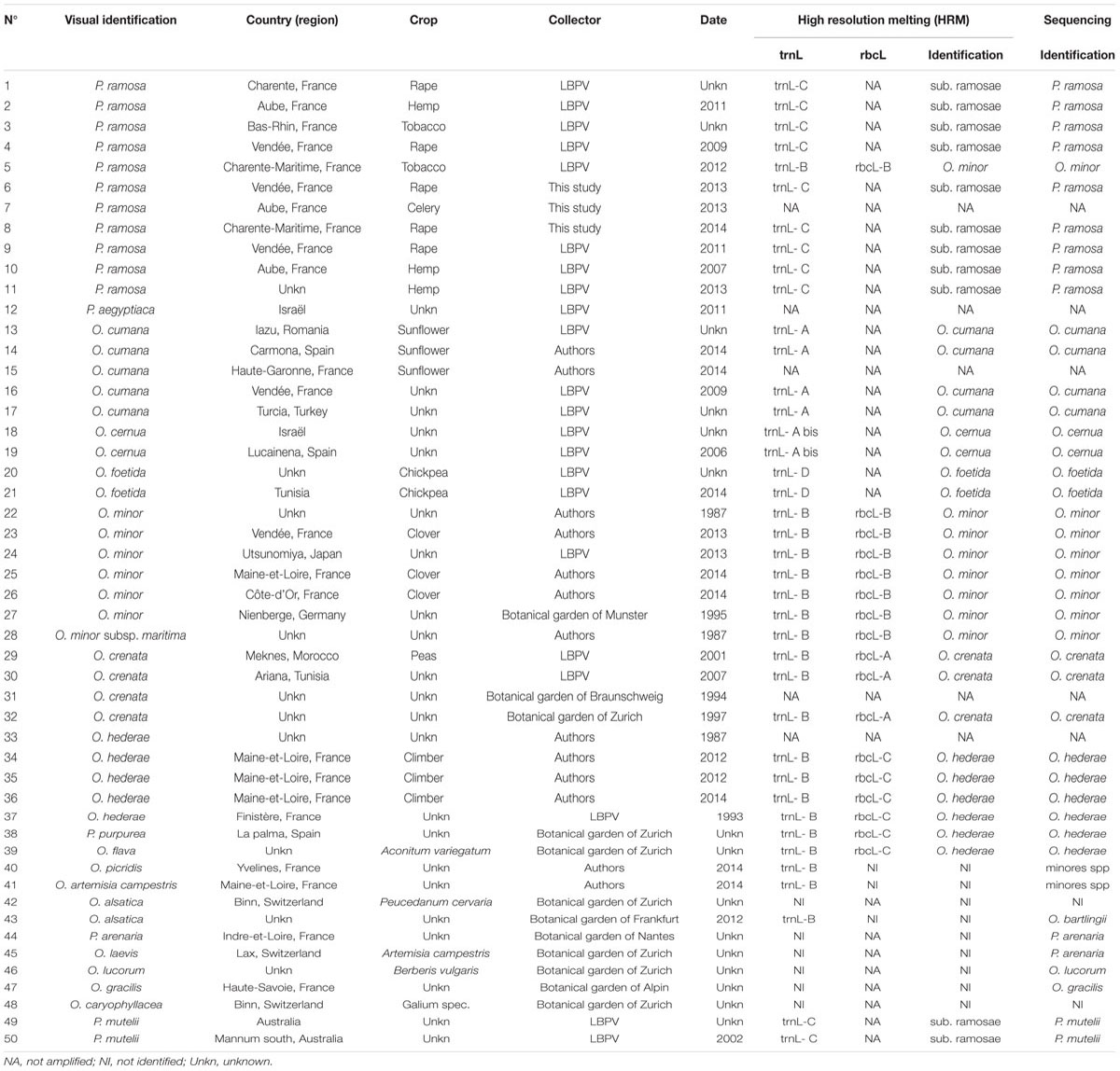
TABLE 1. Origin and identifications of the 50 seed lots tested during the study.
Single Seed Grinding Procedure and DNA Extraction
One of the technical challenges associated with the development of an assay aiming to characterize broomrape single seeds is the ability to obtain enough DNA from seeds weighing an average of 5 μg in a reproducible manner. To this end, each seed was crushed between two microscopy glass slides in presence of 2 μl of ultrapure water and seed tissues were then collected in a microtube. In order to maximize the amount of collected DNA, the slides were rinsed with 400 μl of PL1 extraction buffer (Macherey–Nagel) and the rinse collected in the microtube. Total DNA extraction was then performed using the NucleoSpin® Plant II commercial kit (Macherey–Nagel) following the manufacturer’s instructions (filtration columns were not used). A control of the quantity and quality of the extracted DNA was performed using a NanoVueTM Spectrophotometer (GE Healthcare).
Sequencing
Thanks to previous studies on plastid genome sequences in broomrapes (Wicke et al., 2013; unpublished results), sequences corresponding to eight plastid genes (rbcL, rps7, rps11, rpl36, rpl16, trnQ, trnL, and rrn23) and one nuclear region (ITS) were obtained for the eight studied species. Sequences were aligned using the default alignment algorithm of Geneious v5.6.4. Two markers showing significant sequence divergence among the eight species were selected for subsequent HRM experiments: trnL and rbcL.
To design HRM primers and to control the identification of the species, pseudogenes trnL and rbcL were amplified and sequenced, respectively, using the primers (i) trnL C (F) and trnL HRM R, (ii) 1F and 1352R (Table 2). Amplification was performed on 5 μl of single seed total DNA extract, by 1 U of AmpliTaq Gold® (Life Technologies), in a total volume of 40 μl at the final concentration of 1X of the appropriate Buffer II, 0.3 μM of each primer, 1.5 mM of MgCl2, and 0.2 mM of dNTP. PCR conditions were adjusted as follow, an initial denaturation of 10 min at 95°C, 40 cycles of 30 s at 95°C, 15 s at 58°C, and 1 min at 72°C, and a final extension of 10 min at 72°C. After migration in a 1.5% agarose gel at 180 V for 45 min and ethidium bromide staining, PCR products were visualized under UV light. Purification and sequencing of the PCR products was provided by Genoscreen.

TABLE 2. Primers used for amplification, sequencing, and HRM.
Primer Design
Obtained sequences were aligned using the default alignment algorithm of Geneious v5.6.4 (some alignments are provided as Supplementary Images 1 and 2). Conserved regions and potential markers were identified visually. To achieve HRM identification of the species, primers surrounding the selected markers were designed using primer 31 with an estimated melting temperature of 60°C (Table 2). According to the tested species, the designed primers surround fragments of 315–463 bp for trnL and 345–389 bp for rbcL.
High Resolution Melting Analysis
HRM reactions were performed on 5 μl of single seed DNA extract, in a total volume of 20 μl, using the MeltDoctor Master mix (Life technologies) on a StepOnePlus instrument (Applied Biosystems) following the manufacturers recommendations. TrnL pseudogene was amplified using the primers trnL-Z1-F and trnL-lg-2R at the final concentration of 0.2 μM, rbcL by the primers rbcL-lg-1-F and rbcL-Z2-R at the final concentration of 0.15 μM (Table 2). PCR conditions were adjusted as follow, an initial denaturation of 10 min at 95°C, 45 cycles of 15 s at 95°C, and 1 min at 60°C, a complete denaturation of 10 s at 95°C, 1 min at 60°C, and a continuous melt rising from 60 to 90°C with 0.3% temperature increment every 15 s.
Each extract was run in duplicate, in the presence of the usual positive, negative and process controls and in the presence of reference materials used for HRM profiles analysis. One reference material is required for each HRM profile. These reference materials were previously prepared by extraction of identified seeds using the described protocol and control of the species by sequencing.
Considering the real-time amplification results, only the samples providing cycle threshold (Ct) values below 35 were considered for HRM results analysis. Analysis of the melting profiles was performed using High Resolution Melt Software v3.0 (Applied Biosystems).
Results
DNA Extraction from Single Seeds
A simple methodology was developed to crush individual seeds between two microscopy glass slides and extract total DNA from this crushed material. For the 50 seed lots tested (Table 1), extractions and amplifications were performed separately from two single seeds. DNA concentration of the obtained extracts was too low to be measured using a Nanovolume spectrophotometer. For seed lots number 22, 29, 30, and 37, only one of the extracts allowed a proper amplification. For lots number 7, 12, 15, 31, and 33, respectively, harvested in 2013, 2011, 2014, 1994, and 1987, it was not possible to obtain any amplification. For these five last seed lots, single seed extraction was performed on two more seeds with the similar results. The failure of these seed batches was not species-specific associated. The viability of the different seed lots was not assessed.
Species Identification
For the selected markers trnL and rbcL, respectively, 9 and 11 primer pairs were designed and evaluated for their ability to provide a suitable assay. Results obtained with the best primers are reported. HRM primers were first selected according to their ability to differentiate the eight target species by providing distinct HRM profiles. Discrimination between species was possible because of sequence divergences (SNP and indel) between the amplicons. Targeted species show five different profiles when considering the high resolution melt curves of the trnL PCR product (Figure 1A). O. cumana, O. cernua, and O. foetida are easily identified using this HRM marker since each of these species is the only one associated with a profile (respectively, red, orange, and yellow). O. crenata, O. minor, and O. hederae are associated with the blue profile. They can be then differentiated from other species but not from each other. The same goes for species P. ramosa and P. aegyptiaca associated with the green profile. Among the eight species considered, the rbcL primers amplify only O. crenata, O. minor, and O. hederae. PCR products obtained from these three species show distinct and identifiable HRM profiles (respectively, red, blue, and green; Figure 1A). Considering the obtained results, an identification key is proposed to facilitate the analysis of the results (Table 3).
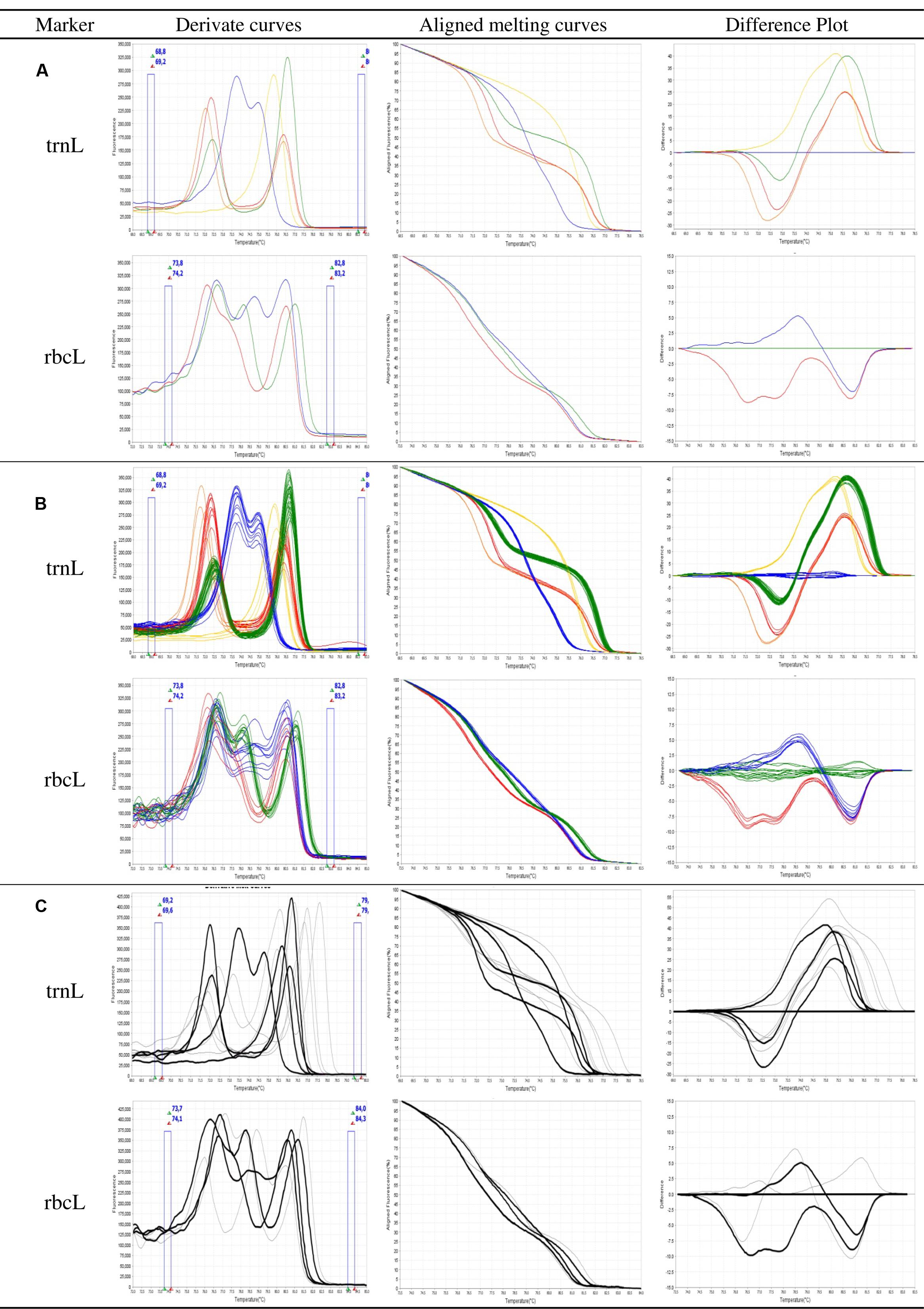
FIGURE 1. High resolution melting (HRM) analysis of broomrape single seeds for the markers trnL and rbcL. (A) Ability of the assay to differentiate the targeted species as different profiles (i) trnL marker: red = Orobanche cumana; orange = O. cernua; blue = O. crenata, O. minor, or O. hederae; green = ramosae clade; yellow = O. foetida; (ii) rbcL marker: red = O. crenata; blue = O. minor; green = O. hederae. (B) Consistency of the profiles between samples of a same species. (C) Specificity of the assay, curves in black correspond to the targeted species, while curves in gray correspond to other tested species. Derivate, aligned melting curves, and difference plots correspond to three representations of the same data: aligned melting curves have been normalized by eliminating fluorescence variance out of the melt regions; difference plots are achieved by subtracting the normalized fluorescence data of a user-defined genotype from that of each of the other samples in the HRM analysis.
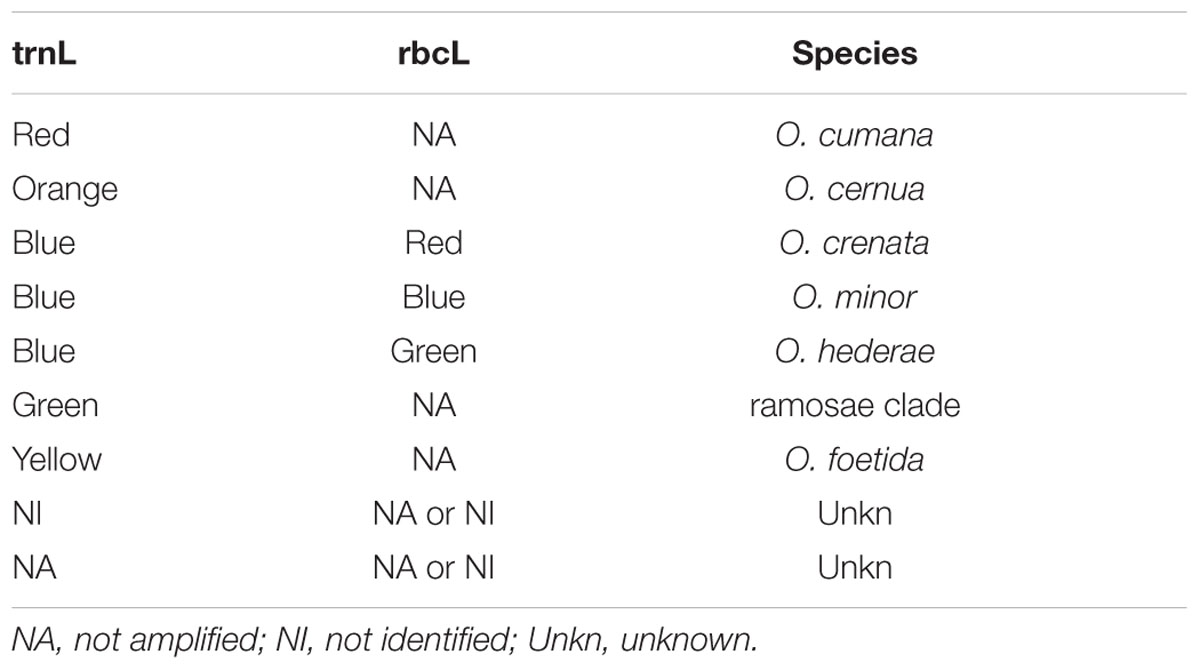
TABLE 3. Correspondence between HRM profiles obtained using the trnL and rbcL markers and broomrape species.
The second primers selection criterion was the consistency of the profiles between lots belonging to the same species. Amplification and HRM were performed on single seed DNA extracts obtained from the 37 (out of 50) available seed lots belonging to the eight targeted species (Table 1). Figure 1B presents the aspect of the obtained melting curve for the trnL and rbcL PCR products. Considering raw (not shown) or derivated melt curves, profiles obtained from samples of identical species show some variability. However, for identical profiles, when considering the aligned melt curves of both trnL and rbcL, the highest relative standard deviation of measured melting temperatures is 3.3%. The different profiles presented above are consistently reproduced between samples of identical species.
Specificity of the Assay
Besides the ability of HRM primers to discriminate among weedy broomrape species, both trnL and rbcL primer pairs amplified single seed DNA extracts obtained from additional 13 seed lots initially identified based on morphological characteristics of adult plant as belonging to eight wild Orobanche species and three wild Phelipanche species (Table 1; Figure 1C). Among the 11 wild species, nine showed original non-identified profiles (NI) which could be easily distinguished from the profiles of the weedy species. Samples 38 and 39, respectively, declared in collection records as P. purpurea and O. flava were identified as O. hederae by HRM analysis. Samples 49 and 50 initially declared in collection records as P. mutelii show the same trnL profile than P. ramosa and P. aegyptiaca.
To control the identification made by HRM, sequencing of the pseudogenes trnL and rbcL was performed on all the analyzed DNA extracts. Sequences were submitted to GenBank and are available with accession numbers KX539159–KX539172. Comparison of the HRM and sequencing interpretations is presented in Table 1. Concerning the targeted species, results show a 100% match between interpretations obtained using both techniques. Furthermore, all the samples identified as non-target by sequencing are designated as non-identifiable by HRM. It is interesting to note that identifications performed by sequencing are not always consistent with primary identification based on visual criteria. For samples 38 and 39 visually identified as O. purpurea and O. flava, HRM identifications as O. hederae are consistent with sequencing results, suggesting that the HRM identification is correct and that the initial morphological identification failed. Sequencing confirmed the visual identity of the samples 49 and 50 as P. mutelii. HRM profile common to P. ramosa and P. aegyptiaca is therefore not specific to these two species but also includes close species.
Reproducibility of the Assay
The HRM profiles obtained with the trnL and rbcL primers can be considered as complex. Furthermore, reading a melting profile is performed by an analyst and is somehow subjective. To question the transferability of the technique, 20 seed lots have been analyzed by three analysts in two different laboratories (GEVES and Terres Inovia). Each analyst performed the experiment on one single seed of each seed lot. Obtained results are shown in Table 4. For 7 out of the 20 seed lots tested, at least one extraction did not allow the amplification. When DNA was properly extracted and amplified, obtained results were in accordance except in the cases of lots 13 and 16. In one case out of three, profiles associated with these seed lots of ramosae had the correct aspect but a different melting temperature and were noted as non-identified. These differences of melting temperature were consistently reproduced, however, sequencing of trnL PCR products showed no difference of sequence between these extracts and others. Excluding the failure of proper DNA extraction on some seeds, the reproducibility of the assay (defined as the percentage of agreements between two identifications performed on a same seed lot) is of 90.9%.

TABLE 4. Data of reproducibility generated by three analysts on 20 seed lots, each analyst analyzing one single seed of each lot.
Discussion
Seed producers may face contaminations of their crop seed lots by seeds of noxious broomrapes. In case of trading, they require international seed lot certificates provided by official seed testing stations. This is mainly carried out through analysis of specific purity of seed lots. However, if this analysis can identify seed lots containing broomrape seeds, it cannot allow a clear identification of the parasite species. Indeed, broomrape species identification can be achieved thanks to seed coat morphological features observed under microscopy (Joel, 1987; Abu Sbaih and Jury, 1994), but this approach is extremely difficult even for trained botanists and requires an extensive expertise usually not available in most laboratories. Molecular markers such as ITS, ISSR, plastid genes, or RAPD were developed for identification of broomrape species, but all required large amount of seeds incompatible with an isolation of few seeds from a specific purity analysis. Thus, protocols allowing DNA extraction from broomrape single seed were developed (Portnoy et al., 1997; Osterbauer and Rehms, 2002) and used with RAPD markers (Katzir et al., 1996). Seeds of five different species could be identified using these methods: P. aegyptiaca, P. ramosa, O. cernua, O. cumana, and O. crenata.
The protocol developed in this study is the first work describing the application of HRM curve analysis for differentiation of broomrape species. Compared to previous technologies, the proposed protocol and markers allow to extend the identification spectrum since it was able to differentiate between eight species, the five above mentioned plus O. foetida, O. hederae, and O. minor. The sequences of the root parasitic plants used in this study present variation generating divergences in the HRM patterns. Deletions, insertions, and several SNPs are responsible for the differences in the observed melting curves between the different species amplicons. The two plastid genes, rbcL, and trnL, have been already used as HRM markers for identification of plants (Madesis et al., 2012; Osathanunkul et al., 2015).
By using the HRM technology and by targeting plastid sequences, it was then possible to develop a simple, reliable, and cost effective assay to identify the seven main weedy species of broomrape potentially found in crop seed lots. In addition, it allows discrimination between these weedy species and 12 species lacking agronomic interest. The high level of divergence between species in the targeted sequences provided more complex profiles than for HRM assays targeting SNP (Toi and Dwyer, 2008) or microsatellites (Mackay et al., 2008). However, in most cases, DNA extracted from single seeds allowed a proper amplification and profiles could be identified by the analyst by comparison with the reference materials introduced in each experiment. On 45 amplified samples, the assay provided results perfectly matching with sequencing. The technique was used by several analysts in two laboratories using different HRM-capable real-time PCR machine and visual analysis of the HRM profiles. In these conditions, the technique shows a reproducibility of 90%. This rate of reproducibility is higher than the one received with RAPD markers, known to be weakly reproducible when employed in different laboratories with different PCR apparatus (Jones et al., 1997). The described assay will make then reliable identification much easier for any diagnostic or research purpose. It is also a fast close tube method not requiring post-PCR manipulation such as DNA gel electrophoresis like in RAPD analysis.
However, the success of the developed assay depends on the concentration and/or quality of the extracted DNA. Indeed for some seed lots it was not possible to reach the minimal concentration and/or quality from some tested seeds or even from any of the tested seeds. HRM technique also requires homogenous DNA extract compositions among samples to compare. Composition may indeed impact the melting temperature of the amplification products. By extracting DNA from single seeds, extracts are relatively homogeneous. However, during the evaluation of the reproducibility, some profiles showed the expected melting profile but with a different melting temperature. In a seed lot, heterogeneity of the seeds may therefore occasionally be an issue for profiles comparison.
Using the identified plastid targets, it was not possible to differentiate the species of the taxonomically difficult ramosa aggregate (P. mutelii, P. ramosa, and P. aegyptiaca) referred to as ramosae clade. Further development of the assay by adding a third marker could provide the ability to differentiate species in the ramosa aggregate. If a species identification is necessary after a ramosae clade or a NI result, the product obtained after the HRM amplification and denaturation can be used for sequencing as any regular PCR product.
The development of an assay able to identify broomrape species from single seeds allows testing of seeds found in commercial seed lots but also identification of mature plants from the field. Broomrape seeds are indeed a material easy to collect and transport, it can be stored at room temperature for many years. For the identification of plants at early stages (before the presence of seeds), the assay can be extended to vegetative material.
Author Contributions
MR planned and designed the research; PD and AP performed the plastid genome sequences analysis; AD performed the experiments. MR wrote the paper with the help of PD.
Funding
Funding for this work was provided by the CASDAR “Orobanche” project (N° C-2012-07) financed by the French Ministère de l’Agriculture, de l’Agroalimentaire et de la Forêt (MAAF).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
The authors want to thank Julien Carpezat (Terres Inovia) for his participation to the evaluation of the reproducibility of the assay.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.01838/full#supplementary-material
IMAGE 1 | Alignment of partial sequences of the trnL gene obtained by amplification using the primers trnL C (F) and trnL HRM R of total DNA extracted from individual seeds belonging to the 8 species of interest.
IMAGE 2 | Alignment of partial sequences of the rbcL gene obtained by amplification of total DNA extracted from individual seeds belonging to 7 of the 8 species of interest.
Footnotes
References
Abu Sbaih, H. A., and Jury, S. L. (1994). Seed micromorphology and taxonomy in Orobanche (Orobanchaceae). Flora Mediterr. 4, 41–48.
Benharrat, H., Delavault, P., Theodet, C., Figureau, C., and Thalouarn, P. (2000). rbcL plastid pseudogene as a tool for Orobanche (subsection Minores) identification. Plant Biol. 2, 34–39. doi: 10.1055/s-2000-9457
Benharrat, H., Veronesi, C., Theodet, C., and Thalouarn, P. (2002). Orobanche species and population discrimination using intersimple sequence repeat (ISSR). Weed Res. 42, 470–475. doi: 10.1046/j.1365-3180.2002.00305.x
Cusimano, N., and Wicke, S. (2015). Massive intracellular gene transfer during plastid genome reduction in nongreen Orobanchaceae. New Phytol. 210, 680–693. doi: 10.1111/nph.13784
Dongo, A., Leflon, M., Simier, P., and Delavault, P. (2012). Development of a high-throughput real-time quantitative PCR method to detect and quantify contaminating seeds of Phelipanche ramosa and Orobanche cumana in crop seed lots. Weed Res. 52, 34–41. doi: 10.1111/j.1365-3180.2011.00891.x
Druml, B., and Cichna-Markl, M. (2014). High resolution melting (HRM) analysis of DNA – Its role and potential in food analysis. Food Chem. 158, 245–254. doi: 10.1016/j.foodchem.2014.02.111
Fernández-Aparicio, M., Reboud, X., and Gibot-Leclerc, S. (2016). Broomrape weeds. Underground mechanisms of parasitism and associated strategies for their control: a review. Front. Plant Sci. 7:135. doi: 10.3389/fpls.2016.00135
Gori, A., Cerboneschi, M., and Tegli, S. (2012). High-Resolution melting analysis as a powerful tool to discriminate and genotype Pseudomonas savastanoi Pathovars and Strains. PLoS ONE 7:e30199. doi: 10.1371/journal.pone.0030199
Harris, S. (1999). RAPDs in systematics: a useful methodology. Mol. Syst. Plant Evol. 57, 211–228. doi: 10.1201/9781439833278.ch11
Jaakola, L., Suokas, M., and Haggman, H. (2010). Novel approaches based on DNA barcoding and high-resolution melting of amplicons for authenticity analyses of berry species. Food Chem. 123, 494–500. doi: 10.1016/j.foodchem.2010.04.069
Joel, D. M. (1987). Detection and identification of Orobanche seeds using fluorescence microscopy. Seed Sci. Technol. 15, 119–124.
Joel, D. M. (2009). The new nomenclature of Orobanche and Phelipanche. Weed Res. 49, 6–7. doi: 10.1111/j.1365-3180.2009.00748.x
Jones, C. J., Edwards, K. J., Castaglione, S., Winfield, M. O., Sala, F., van de Wiel, C., et al. (1997). Reproducibility testing of RAPD, AFLP and SSR markers in plants by a network of European laboratories. Mol. Breed. 3, 381–390. doi: 10.1023/a:1009612517139
Katzir, N., Portnoy, V., Tzuri, G., CastejonMunoz, M., and Joel, D. M. (1996). Use of random amplified polymorphic DNA (RAPD) markers in the study of the parasitic weed Orobanche. Theor. Appl. Genet. 93, 367–372. doi: 10.1007/s001220050290
Krypuy, M., Ahmed, A. A., Etemadmoghadam, D., Hyland, S. J., deFazio, A., Fox, S. B., et al. (2007). High resolution melting for mutation scanning of TP53 exons 5–8. BMC Cancer 7:168. doi: 10.1186/1471-2407-7-168
Lochlainn, S. O., Amoah, S., Graham, N. S., Alamer, K., Rios, J. J., Kurup, S., et al. (2011). High Resolution Melt (HRM) analysis is an efficient tool to genotype EMS mutants in complex crop genomes. Plant Methods 7, 43. doi: 10.1186/1746-4811-7-43
Lopez-Granados, F., and Garcia-Torres, L. (1996). Effects of environmental factors on dormancy and germination of crenate broomrape (Orobanche crenata). Weed Sci. 44, 284–289. doi: 10.2307/4045680
Lopez-Granados, F., and Garcia-Torres, L. (1999). Longevity of crenate broomrape (Orobanche crenata) seed under soil and laboratory conditions. Weed Sci. 47, 161–166. doi: 10.2307/4046192
Mackay, J. F., Wright, C. D., and Bonfiglioli, R. G. (2008). A new approach to varietal identification in plants by microsatellite high resolution melting analysis: application to the verification of grapevine and olive cultivars. Plant Methods 4, 8. doi: 10.1186/1746-4811-4-8
Madesis, P., Ganopoulos, I., Anagnostis, A., and Tsaftaris, A. (2012). The application of Bar-HRM (Barcode DNA-High Resolution Melting) analysis for authenticity testing and quantitative detection of bean crops (Leguminosae) without prior DNA purification. Food Control 25, 576–582. doi: 10.1016/j.foodcont.2011.11.034
Manen, J. F., Habashi, C., Jeanmonod, D., Park, J. M., and Schneeweiss, G. M. (2004). Phylogeny and intraspecific variability of holoparasitic Orobanche (Orobanchaceae) inferred from plastid rbcL sequences. Mol. Phylogenet. Evol. 33, 482–500. doi: 10.1016/j.ympev.2004.06.010
Osathanunkul, M., Madesis, P., and de Boer, H. (2015). Bar-HRM for authentication of plant-based medicines: evaluation of three medicinal products derived from Acanthaceae species. PLoS ONE 10:e0128476. doi: 10.1371/journal.pone.0128476
Osterbauer, N. K., and Rehms, L. (2002). Detecting single seeds of small broomrape (Orobanche minor) with a polymerase chain reaction. Plant Health Progress doi: 10.1094/PHP-2002-1111-01-RS
Paran, I., Gidoni, D., and Jacobsohn, R. (1997). Variation between and within broomrape (Orobanche) species revealed by RAPD markers. Heredity 78, 68–74. doi: 10.1038/hdy.1997.8
Parker, C. (2009). Observations on the current status of Orobanche and Striga problems worldwide. Pest. Manag. Sci. 65, 453–459. doi: 10.1002/ps.1713
Perez-De-Luque, A., Fondevilla, S., Perez-Vich, B., Aly, R., Thoiron, S., Simier, P., et al. (2009). Understanding Orobanche and Phelipanche-host plant interactions and developing resistance. Weed Res. 49, 8–22. doi: 10.1111/j.1365-3180.2009.00738.x
Pineda-Martos, R., Velasco, L., and Perez-Vich, B. (2014). Identification, characterisation and discriminatory power of microsatellite markers in the parasitic weed Orobanche cumana. Weed Res. 54, 120–132. doi: 10.1111/wre.12062
Plaza, L., Fernandez, I., Juan, R., Pastor, J., and Pujadas, A. (2004). Micromorphological studies on seeds of Orobanche species from the Iberian Peninsula and the Balearic Islands, and their systematic significance. Ann. Bot. 94, 167–178. doi: 10.1093/aob/mch124
Portnoy, V. H., Katzir, N., and Joel, D. M. (1997). Species identification of soil-borne Orobanche seeds by DNA fingerprinting. Pestic. Biochem. Physiol. 58, 49–54. doi: 10.1006/pest.1997.2281
Reed, G. H., Kent, J. O., and Wittwer, C. T. (2007). High-resolution DNA melting analysis for simple and efficient molecular diagnostics. Pharmacogenomics 8, 597–608. doi: 10.2217/14622416.8.6.597
Taberlet, P., Gielly, L., Pautou, G., and Bouvet, J. (1991). Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 17, 1105–1109. doi: 10.1007/bf00037152
Takano, E. A., Mitchell, G., Fox, S. B., and Dobrovic, A. (2008). Rapid detection of carriers with BRCA1 and BRCA2 mutations using high resolution melting analysis. BMC Cancer 8:59. doi: 10.1186/1471-2407-8-59
Toi, C. S., and Dwyer, D. E. (2008). Differentiation between vaccine and wild-type varicella-zoster virus genotypes by high-resolution melt analysis of single nucleotide polymorphisms. J. Clin. Virol. 43, 18–24. doi: 10.1016/j.jcv.2008.03.027
Westwood, J. H. (2013). “The physiology of the established parasite–host association,” in Parasitic Orobanchaceae: Parasitic Mechanisms and Control Strategies, eds M. D. Joel, J. Gressel, and J. L. Musselman (Berlin: Springer Berlin Heidelberg), 87–114.
Wicke, S., Muller, K. F., Pamphilis, C. W. D., Quandt, D., Wickett, N. J., Zhang, Y., et al. (2013). Mechanisms of functional and physical genome reduction in photosynthetic and nonphotosynthetic parasitic plants of the broomrape family. Plant Cell 25, 3711–3725. doi: 10.1105/tpc.113.113373
Wojdacz, T. K., and Dobrovic, A. (2007). Methylation-sensitive high resolution melting (MS-HRM): a new approach for sensitive and high-throughput assessment of methylation. Nucleic Acids Res. 35, e41–e41. doi: 10.1093/nar/gkm013
Keywords: Orobanche, Phelipanche, parasitic weed, molecular diagnosis, HRM, trnL, rbcL
Citation: Rolland M, Dupuy A, Pelleray A and Delavault P (2016) Molecular Identification of Broomrape Species from a Single Seed by High Resolution Melting Analysis. Front. Plant Sci. 7:1838. doi: 10.3389/fpls.2016.01838
Received: 27 April 2016; Accepted: 22 November 2016;
Published: 12 December 2016.
Edited by:
Monica Fernandez-Aparicio, Institut National de la Recherche Agronomique (INRA), FranceReviewed by:
Paula Martins-Lopes, University of Trás-os-Montes and Alto Douro, PortugalLeonardo Velasco, Institute for Sustainable Agriculture – CSIC, Spain
Manuel Miller, Helmholtz Zentrum München, Germany
Copyright © 2016 Rolland, Dupuy, Pelleray and Delavault. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mathieu Rolland, bWF0aGlldS5yb2xsYW5kQGFuc2VzLmZy