 Paul Brouwer1‡
Paul Brouwer1‡ Andrea Bräutigam2†‡
Andrea Bräutigam2†‡ Valerie A. Buijs1
Valerie A. Buijs1 Anne O. E. Tazelaar1Adrie van der Werf3
Anne O. E. Tazelaar1Adrie van der Werf3 Urte Schlüter2Gert-Jan Reichart4Anthony Bolger5
Urte Schlüter2Gert-Jan Reichart4Anthony Bolger5 Björn Usadel5
Björn Usadel5 Andreas P. M. Weber2
Andreas P. M. Weber2 Henriette Schluepmann1*
Henriette Schluepmann1*- 1Molecular Plant Physiology, Institute of Environmental Biology, Utrecht University, Utrecht, Netherlands
- 2Institute for Plant Biochemistry, Cluster of Excellence on Plant Sciences, Heinrich Heine University, Düsseldorf, Germany
- 3Plant Research International, Wageningen, Netherlands
- 4Organic Geochemistry, Utrecht University, Utrecht, Netherlands
- 5Institute for Botany and Molecular Genetics, Bioeconomy Science Center, RWTH Aachen University, Aachen, Germany
Sustainable agriculture demands reduced input of man-made nitrogen (N) fertilizer, yet N2 fixation limits the productivity of crops with heterotrophic diazotrophic bacterial symbionts. We investigated floating ferns from the genus Azolla that host phototrophic diazotrophic Nostoc azollae in leaf pockets and belong to the fastest growing plants. Experimental production reported here demonstrated N-fertilizer independent production of nitrogen-rich biomass with an annual yield potential per ha of 1200 kg−1 N fixed and 35 t dry biomass. 15N2 fixation peaked at noon, reaching 0.4 mg N g−1 dry weight h−1. Azolla ferns therefore merit consideration as protein crops in spite of the fact that little is known about the fern’s physiology to enable domestication. To gain an understanding of their nitrogen physiology, analyses of fern diel transcript profiles under differing nitrogen fertilizer regimes were combined with microscopic observations. Results established that the ferns adapted to the phototrophic N2-fixing symbionts N. azollae by (1) adjusting metabolically to nightly absence of N supply using responses ancestral to ferns and seed plants; (2) developing a specialized xylem-rich vasculature surrounding the leaf-pocket organ; (3) responding to N-supply by controlling transcripts of genes mediating nutrient transport, allocation and vasculature development. Unlike other non-seed plants, the Azolla fern clock is shown to contain both the morning and evening loops; the evening loop is known to control rhythmic gene expression in the vasculature of seed plants and therefore may have evolved along with the vasculature in the ancestor of ferns and seed plants.
Introduction
Large yield increases have been achieved by selecting crop varieties particularly responsive to fertilizer applications (Evenson and Gollin, 2003). Yet nitrogen use efficiency of crops when inorganic nitrogen salts are applied to soil is low: as much as 50% of the applied fertilizer is not utilized resulting in often unwanted eutrophication of surrounding ecosystems. Chemical synthesis of nitrogen (N) fertilizer is presently consuming 1–2% of the yearly fossil fuel derived energy and thus is a major contributor to CO2 pollution (Erisman et al., 2008). In addition, denitrification of the wasted N-fertilizer leads to release of N2O, with greenhouse warming effects up to 300-fold those of CO2 (Galloway et al., 2004). Taken together, although spectacular yield increases obtained during the past century with most crops are founded on N-fertilizers, their extensive use is unsustainable, and alternatives must be found to mitigate environmental impact of agriculture.
Reducing N-fertilizer applications without yield penalty may be possible with improved nitrogen use efficiency of crops combined with a more precise application of N-fertilizer (Xu et al., 2012) or by using crops fixing N2 from the atmosphere. N2 fixation is generally carried out by bacteria associated with the plants, as in root nodules of leguminous crops and in extracellular spaces of sugar cane, or by bacteria freely living in the soil and waterways (Dos Santos et al., 2012; Pankievicz et al., 2015). The most established N2-fixing crop symbiosis is the soybean (Glycine max) reaching 230 million metric tons annual production in 2008 ranking fourth in production area after wheat, rice, and maize (Hartman et al., 2011). Soybean harbors rhizobia which, after a complex communication between plant and bacteria is triggered, live in nodules of plant roots in facultative symbiosis (Freiberg et al., 1997). Rhizobia in the nodules are fed by plant sugars and secrete ammonium in return (Udvardi and Poole, 2013). Soybean plants reject colonization by rhizobia if nitrogen fertilizer is available. Modern high-yield cultivars, are generally more responsive to N-fertilizer which contributes some 40% to protein in the seed (Wilson et al., 2014). Similar trends are found with the high yielding N2-fixing forage alfalfa (Medicago sativa), that yields 21 t ha−1 a−1 dry weight (DW) upon addition of 150 kg N ha−1 a−1 (Anglade et al., 2015). To further emancipate plant protein production from N-fertilizer, fixation rates of symbionts in established crops will need to be improved (Oldroyd and Dixon, 2014). Alternatively, plant/bacteria symbioses that fix N2 more efficiently will need to be domesticated. The latter will counter threats associated with increasingly reduced biodiversity on farm land (Stamp et al., 2012). Development of new or orphan crops is timely due to recent advances in genetics that tremendously speed up breeding.
N2-fixing ferns from the genus Azolla have been known as bio-fertilizer in submerged field based taro and rice cultivation in China, Vietnam, and Senegal (Shi and Hall, 1988; Wagner, 1997); Azolla species distribution is global from temperate to tropical regions. Drained fields emit CO2 (Liu et al., 2013). Submerging fields and growing Azolla may contribute to reducing CO2 emissions. Azolla is much easier to contain and harvest in such field situations compared with microalgae or cyanobacteria since growing it does not require sophisticated pumping or mixing to ensure carbonate availability. The N2 fixation of these ferns is generally thought to be carried out by the phototrophic bacterium Nostoc azollae, which was reported to have an eroded genome as a consequence of its symbiotic lifestyle (Ran et al., 2010). The notion that N. azollae are the only symbionts in Azolla has been challenged based on electron microscopic observations and this raises questions concerning their role in N trafficking within the symbiosis (Carrapiço, 1991; Zheng et al., 2009). Azolla/Nostoc symbioses have probably evolved some 90 M years ago (Metzgar et al., 2007), before the legume/rhizobia symbioses dated to have evolved 64 M years ago (Herendeen et al., 1999; Kistner and Parniske, 2002). N. azollae are transmitted vertically during the fern life cycle and thrive inside specialized leaf pockets of the fern that close fully upon maturation after motile hormogonia have infected young leaves (Kaplan and Peters, 1981; Rai et al., 2000; Zheng et al., 2009). Leaves of A. filiculoides are arranged in two rows, each leaf has two lobes, the lower lobe is shaped like a boat and rests on the water surface holding the other upper lobe in an aerial position exposed to light (Supplementary Figure 1). The upper leaf lobe contains a leaf pocket with a single ad-axially located pore (Supplementary Figure 1) presumed to mediate exchange of gasses (Veys et al., 1999).
It is hypothesized that the fern contributes sugars while N. azollae releases some 40% of the fixed nitrogen into the leaf pocket as ammonia; a reduced glutamate synthase activity in the symbiont compared to free-living cyanobacteria may cause this release (Peters and Meeks, 1989; Meeks, 2009). The ammonia is then assimilated by the glutamate synthase and glutamine synthetase (GS/GOGAT) cycle of the fern (Meeks et al., 1987). Label from 15N2 fixed travels from the mature leaves to the shoot tips presumably supplying young leaves and dividing bacterial cells of the shoot tip (Ito and Watanabe, 1983). Molecular mechanisms underpinning the fern responses to N supplied by N. azollae are entirely unknown, yet a gene database was recently established from transcriptome sequencing that may now be used to profile the responses (Brouwer et al., 2014). Azolla species are known to thrive in ditches containing nitrate or/and ammonium, possibly in environments with high P/N ratios. 15N-labeled nitrate is known to be taken up by the root and the label to move slowly to the shoot where it accumulates in the old branches. In contrast, 15N-labeled ammonium is more rapidly taken up and its label rapidly accumulates in the shoot tip, more rapidly that from 15N2 (Ito and Watanabe, 1983). The difference suggests that nitrate reduction occurs at a low rate in the fern. In Azolla unlike in many cyanobacteria, low concentrations (1 mM) of NH4SO4 or potassium nitrate did not inhibit nitrogenase assayed using the acetylene reduction assay which suggests that cyanobacteria in Azolla may not be exposed to medium concentrations of nitrogen salts (Ito and Watanabe, 1983). Particularly nitrate salts did not inhibit nitrogenase activity until 25 mM in 24 h exposure. This could be due to absence of nitrate transporters in N. azollae as reported for A. filiculoides (Ran et al., 2010).
In batch cultures illuminated with incandescent light, Azolla ferns from several species were reported to reach doubling times as low as 2 days in their exponential growth phase without N-fertilizer, yielding an N-content of 4–5% of the DW (Peters et al., 1980). Due to high productivity the ferns ought to be considered as high protein crops in their own right. Data on productivities without N-fertilizer, however, is lacking for ferns grown in the linear growth phase in closed-canopy cultures, the latter being crucial to suppress algae and outgrow fungal and insect pests.
Here we set out to examine the productivity of A. filiculoides under continuous production conditions over 138 days. The effect of various inorganic N-fertilizers on the productivity in terms of biomass, the N-content of the biomass and expression of key fern and bacterial marker genes were analyzed simultaneously. Confocal laser scanning microscopy was used to reveal features of the leaf pocket important for metabolic connectivity of the symbiosis. A. filiculoides clones with and without N. azollae were used to verify the contribution of N. azollae to the fern growth rates. 15N2-fixation rates were measured during the diel cycle and after acclimation to 2 mM NH4NO3. Diel transcriptome analysis was used to probe the fern diel physiological response to different nitrogen sources. RNA sequence assemblies were further used to examine ammonium transporters and markers of vasculature development.
Materials and Methods
Plant Materials
Azolla filiculoides Lam. was collected in Utrecht as described by Brouwer et al. (2014). A. pinnata R. Br. was from the International Rice Research Institute Bio-fertilizer Collection [code PI 0535, originally from Sigiriya in Sri Lanka provided by S. Kulasooriya in 1984; (Watanabe et al., 1992)].
Growth Conditions
The Standard Azolla Medium (SAM) was liquid IRRI medium (Watanabe et al., 1992) with the following modifications: 0.32 μM, CuSO4, 0.835 μM ZnSO4, and 17.9 μM Fe-EDTA. The medium was neither pumped, nor mixed during growth. Light intensity was 300 μmol photons m−2 s−1 over the wave band 400–700 nm supplemented with incandescent lamps for 12 h. Day-time temperature was 22°C whilst night-time temperature was 15°C with the exception in the 15N2-fixation experiment where temperature was kept a constant 21°C. The growth conditions under experimental production were as above but incandescent lamps were turned on for only 2 h at the end of the day. Containers were open to ambient air; water was stagnant. Mat density reached 2–3 kg m−2 fresh weight (FW) biomass before the continuous harvest under linear growth commenced; after each harvest nutrients were added to replace the nutrients removed in the biomass harvested. Full medium replacement was carried out once every 5 weeks.
To obtain sterile cultures of the symbiosis, frond pieces (<1 mm3) of A. filiculoides were surface sterilized using bleach at 1% available chlorine for 40 s and four consecutive rinses in sterile water before culture on agar solidified SAM medium. SAM was solidified with 0.6% agarose (Duchefa, Netherlands). Fern clones without N. azollae were selected on agar solidified SAM with 60 μg ml−1 Erythromycin as previously described (Forni et al., 1991); absence of N. azollae was routinely verified by confocal microscopy using the characteristic bacterial fluorescence as a marker (Brouwer et al., 2014).
Sterile cultures of surface sterilized plants (1 L liquid medium with and without 2 mM NH4NO3) were grown in enclosed glass containers with a stream of air (78 L h−1) pumped through 0.45 μm filters using aquarium pumps (SuperFish Air flow mini); these sterile cultures were used in all of the DNA and RNA sequencing experiments. Sterile cultures were also used for the 15N2-fixation experiments.
Confocal Laser Scanning Microscopy
Propidium iodide/periodic acid staining and mounting of the tissues were essentially as described by Truernit et al. (2008). A Leica SP2 laser scanning confocal fluorescence microscope, equipped with either ×10 or × 40 objectives and a laser with excitation wavelength of 405 nm, was used to visualize propidium iodide at 530–560 nm and auto-fluorescence at 515–530 nm.
RNA Extractions and Quantitative RT-PCR
Plant material was dry blotted, 50 mg FW snap frozen in liquid N2, then ground frozen with two glass beads using the TissueLyser II (Qiagen) set at maximum speed for 1.5 min twice. RNA was extracted with the Spectrum Plant Total RNA kit applying protocol B (Sigma–Aldrich). RNA was then treated with DNase (5 units for 3 μg RNA) 30 min at 37°C and the reaction stopped by 10 min at 65°C in the presence of EDTA (2 mM).
Primers (polyT and random hexamers at 0.030 and 0.074 μg μl−1 final concentrations, respectively, in the reverse transcription reaction) and 1 μg DNase treated RNA were denatured 5 min at 72°C before reverse transcription with MLV reverse transcriptase (5 units, Fermentas) for 10 min at 37°C then 40 min at 42°C. Primers used for q-RT PCR are listed in Supplementary Table 1. Quantitative RT-PCR was performed as described in Brouwer et al. (2014).
RNA Sequencing and Bioinformatic Analyses
Surface sterilized A. filiculoides were grown in excess sterile SAM with and without 2 mM NH4NO3 for 7 days in triplicate replicate cultures. Growth was in a chamber with light set to start at 0600 for 12 h. On day 8 samples were collected, snap frozen at 2, 8, 14, and 20 h of the diel cycle, then total RNA extracted and DNAse treated as above. RNA integrity, sequencing library quality, and fragment size were verified on a 2100 Bioanalyzer (Agilent). Libraries were prepared using the TruSeq RNA Sample Prep Kit v2 (Illumina), and library quantification was performed with a Qubit 2.0 (Invitrogen). All libraries were sequenced on the HISEQ2000 Illumina platform in paired-end mode.
Paired-end reads including public available reads from de Vries et al. (2016) were mapped against the transcriptome database (Brouwer et al., 2014) using CLC genomics workbench®(CLC bio, Denmark) using default parameters (mismatch cost 2, insertion cost 3, deletion cost 3, length fraction 0.8, similarity fraction 0.8, auto-detect paired distance, strand specific both and maximum number of hits for a read 30). Only paired matches were counted. Read counts were used for statistical analysis with edgeR in classic mode (Robinson et al., 2010) using the Bioconductor package. Results were corrected for multiple hypothesis testing using Benjamini–Hochberg FDR correction (Benjamini and Hochberg, 1995) and considered significant if q < 0.01. Read counts were normalized to total mapped reads for each sample and expressed as reads per million reads (rpm; Supplementary Tables 2, 4). Principle component analysis and clustering were conducted on all transcripts which being summed exceeded 10 rpm using the MultiExperimentViewer1.
Clock genes were annotated as the closest homolog to the Arabidopsis thaliana clock genes (Nakamichi, 2011). The diel transcripts were identified by isolating transcripts which display differential expression between at least one pair of two adjacent time points then loaded into Mapman for visualization and enrichment tests (Thimm et al., 2004). Functional annotations were imported using the Mercator Pipeline (Lohse et al., 2014) based on the Azolla transcriptome sequences (Brouwer et al., 2014). K-means clusters were built using the MultiExperimentViewer with Pearson distance metric.
N-Content Determinations and 15N2 Fixation Rates
Ferns (100 mg FW) were grown in pots with 43 ± 4 ml of sterile SAM and a remaining air space of 262 ± 4 ml. To the air space 25 ml of 15N2 was added using air-tight syringes; overpressure was slowly brought to ambient pressure using a release needle. Plants were subsequently grown in these conditions for 2 h at the following time points: 8, 14, 20, 2, and 8 h in the diel cycle where the 12 h day started at 7 h. Ferns devoid of cyanobacteria at the 14 h time point, in the light, were used as the no fixation control. In addition, the effect of 7 day acclimation to 2 mM NH4NO3 on ferns with cyanobacteria was tested at the 14 h time point. Samples were frozen at −20°C, homogenized and freeze-dried before analysis of the dry weights and isotope abundance determinations. All data points represent the average of triplicate biological samples.
Total N-content and stable nitrogen isotopes (δ15N) were analyzed on a ThermoScience Delta Plus isotope ratio mass spectrometer connected on-line to a Carlo Erba Instruments Flash 1112 elemental analyzer. The δ15N of each sample was expressed relative to atmospheric dinitrogen as (‰) and calculated as the following ratio: (([15NSample]-[15NAir]) ∗ 103)/[15NAir]) with [15NSample] the concentration of 15N isotope in the sample, [15NAir] the concentration of 15N in standard air. Instrumental precision was better than 0.2‰. N2 fixation rates were determined using the following calculation: ((([15Nt2] [Nbiomast2])-([15Nt0] [Nbiomast0]))/[15NAirspace] ∗ 2 h) with [15Nti] the concentration of 15N before (i = 0) or after 2 h incubation (i = 2) with 15N2 enriched air in the bottle, [Nbiomasti] the N-content in the biomass before (i = 0) and after 2 h incubation (i = 2) and [15Nairspace] the ratio of 15N over total N in the air space. We assumed no isotope discrimination during the fixation process and therefore rates of fixation calculated may be underestimated.
Results
Azolla filiculoides Yielded 35 t DW ha−1 a−1 in an Experimental Continuous Harvest System
To evaluate the potential of species from the genus Azolla as high protein biomass crops, A. filiculoides was grown under continuous production conditions for 138 days (Figure 1A). When harvesting 33% of the surface twice per week, accumulated biomass increased linearly (Figure 1B). From the slope of dry biomass accumulation, productivity was calculated at 9.72 g biomass DW m−2 day−1, corresponding to an annual yield potential of 35.5 t ha−1 DW for A. filiculoides. Very similar results were obtained with A. pinnata (data not shown). To test whether yield performance of these ferns was influenced by N-fertilizers often encountered in runoff from agricultural land, we examined the effect of several inorganic nitrogen sources in the media used for continuous production. Growth of A. filiculoides was in SAM without nitrogen or with 4 mM NaNO3, NH4Cl or 2 mM NH4NO3. We harvested 33% of the culture area twice weekly over a period of 4 weeks, whilst replenishing nutrients weekly. Cumulative harvest over 30 days indicated slight benefits with N-fertilizer (up to 15%, Figure 2A). N-fertilizers moreover led to N accumulation in the biomass, from 3.41 up to maximally 4.36% w/w N with NaNO3 (Figure 2B) and to induced expression of nitrate reductase from the fern (Figure 2C). NaNO3 as well as NH4NO3 did not affect transcript abundance of the bacterial housekeeping gene N. azollae secA suggesting that bacterial growth was not affected by NO3− containing media (Figure 2D). In contrast, NH4Cl reduced A. filiculoides productivity and led to a reduction of N. azollae secA and nifH transcript detection (Figures 2D,E). nifH encodes the catalytic subunit of the nitrogenase enzyme and is intact in N. azollae (Ran et al., 2010). The above results demonstrate that A. filiculoides does not require N-fertilizer to sustain high productivity but that it reacts to and benefits from additional N sources in the medium.
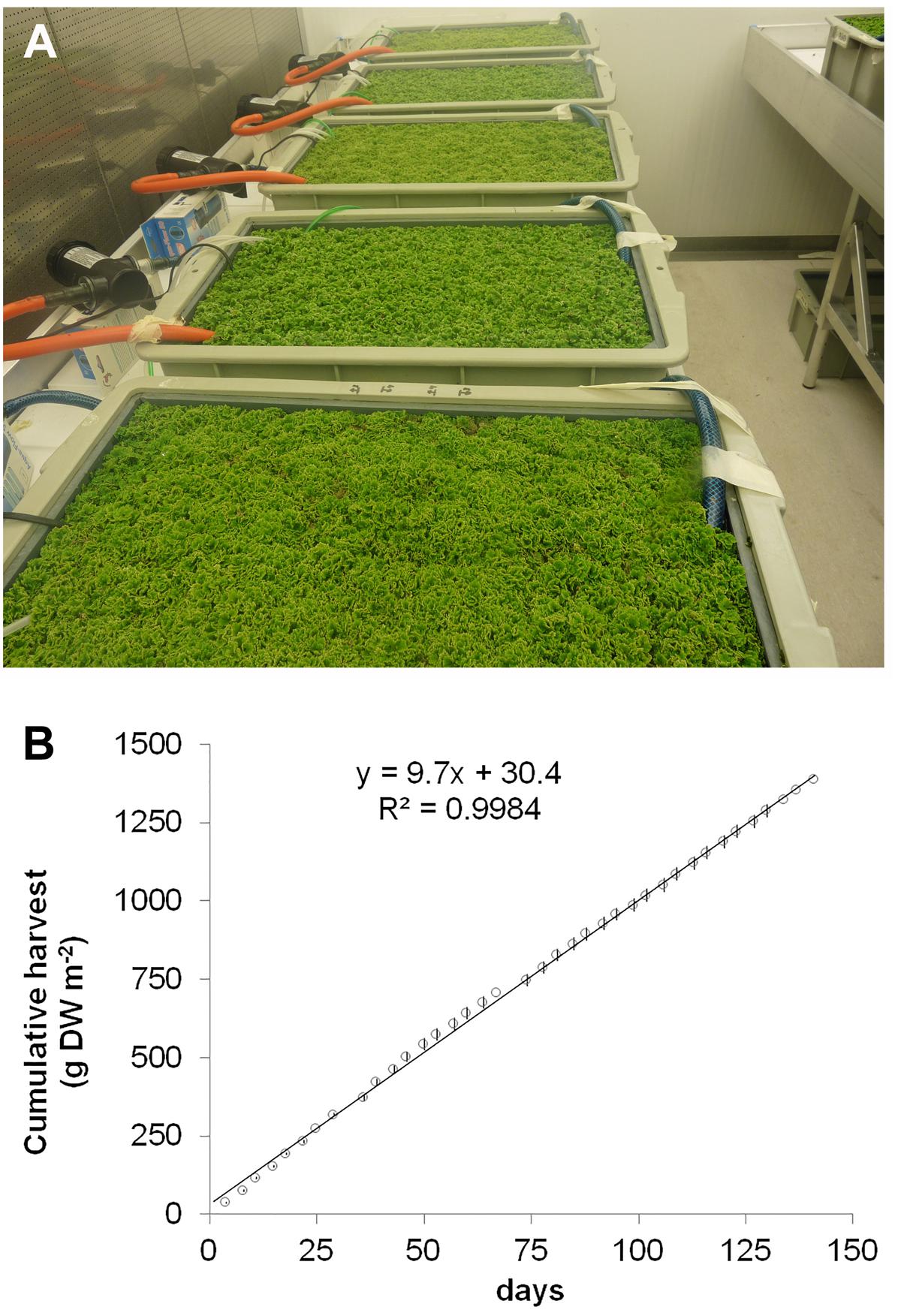
FIGURE 1. Azolla filiculoides yield potential under continuous production conditions. Ferns were grown with 300 μmol s−1 cm−2 photosynthetic active radiation at 25°C for the 12 h day and at 22°C during the 12 h night. (A) Closed-canopy cultures with plants in the linear growth phase for continuous harvest. (B) Cumulative dry biomass harvested. Once standing crop formed a closed mat at 160 g m2 dry weight, continuous harvest was at a rate of 33% of the biomass twice per week. Linear regression equation is shown along with its fit; n = 6, standard deviations were smaller than the labels.
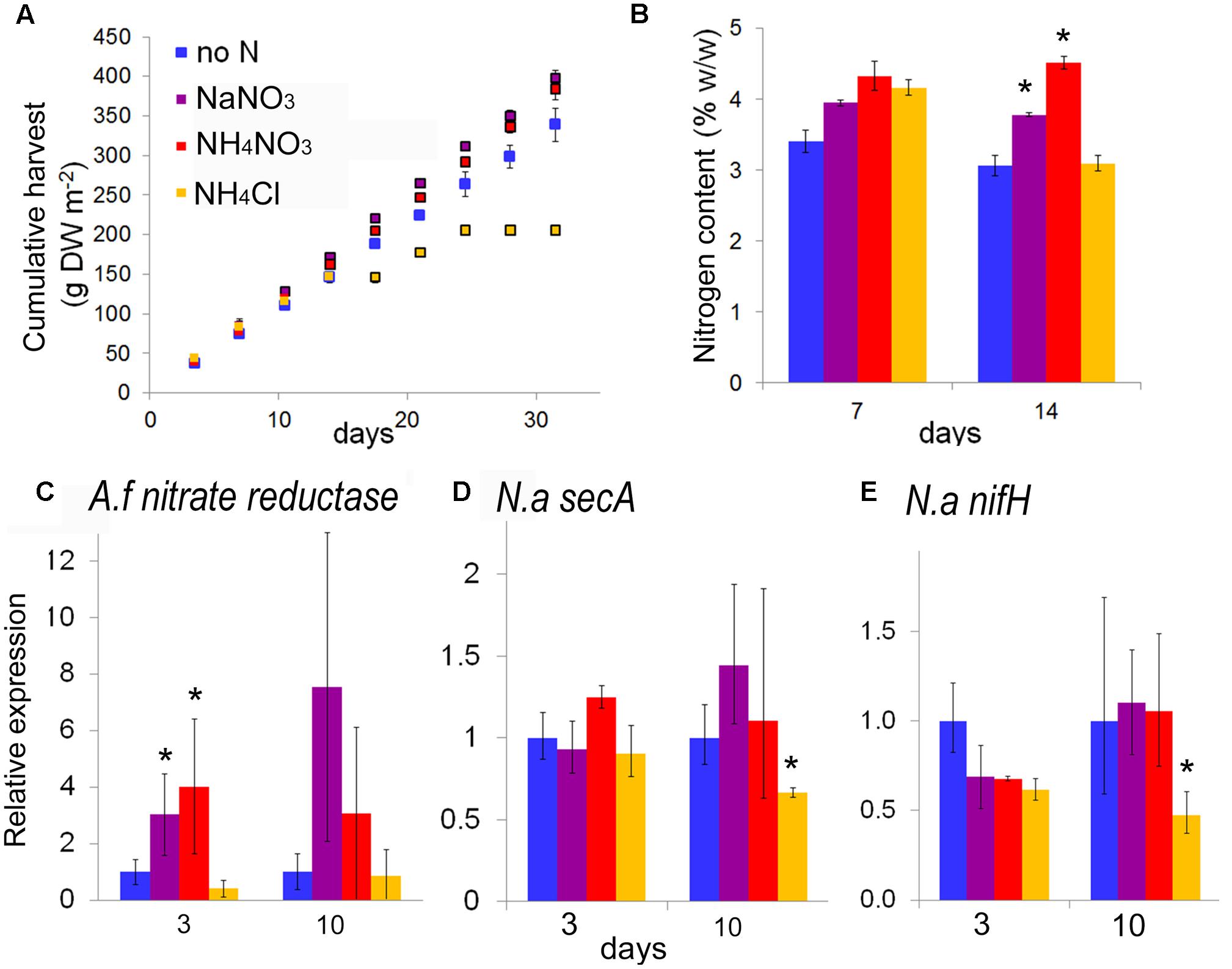
FIGURE 2. Azolla filiculoides response to N-fertilizer under continuous production conditions. Ferns were grown as in Figure 1 but this time in medium without (no N) or with 4 mM NaNO3, or 2 mM NH4NO3, or 4 mM NH4Cl. Biomass harvested was split to determine cumulative harvest (A) nitrogen content (B) and to extract mRNA. Reverse transcription of the RNA was with an excess random hexamers to analyze levels of A. filiculoides and Nostoc azollae transcripts by quantitative PCR: A. filiculoides nitrate reductase Afcontig 35782 (C), N. azollae secA (D) and nifH (E). Standard deviation is for n = 3, T-test significance with reference to no N for P < 0.05 is marked with ∗.
Growth of A. filiculoides without N. azollae Entirely Relied on Exogenous N-Fertilizer
To evaluate the contribution from N. azollae to fern productivity we raised surface sterilized A. filiculoides, then for every fern, fronds were grown without and with erythromycin so as to obtain clonal material with and without cyanobacteria. Complete absence of N. azollae was verified using confocal microscopy and quantitative PCR on the extracted plant DNA (Supplementary Figure 2). A. filiculoides without N. azollae failed to grow without a source of reduced N and with 2 mM NH4Cl (Figure 3A). Growth in all other conditions was exponential. Optimum supplementation of ferns without N. azollae was using 4 or 8 mM NaNO3, or 2 mM NH4NO3, and yielded growth rates approaching those of ferns with N. azollae (Figures 2A, 3A). Given the possibility of amino acids as exchange substrates between fern and symbiont in addition to ammonia (Kaplan and Peters, 1981; Meeks et al., 1987), amino acids were tested for their ability to restore growth keeping the total N supplied in the amino acids at 2 mM in the medium (Figure 3B). Whilst arginine and proline supported highest average growth rates that were not significantly different from ferns with N. azollae, growth rates on amino acids mostly correlated with those from the seed plant A. thaliana (Forsum et al., 2008) and, therefore, unlikely reflected trafficking inside leaf pockets. Close-up inspection confirmed that the leaf pockets were well removed from the medium and enclosed entities except for the leaf-pocket pore open to air (Figure 3C and Supplementary Figure 1). Confocal microscopy further revealed a conspicuous vasculature curving around the pockets that was closely connected to the pocket fluid (Figure 3C). This vasculature differed from that in leaf lobes without leaf pocket: it was made of several layers of tracheid cells with re-enforced cell walls containing pectin brightly stained by the propidium iodide/periodic acid stain (Supplementary Figure 3).
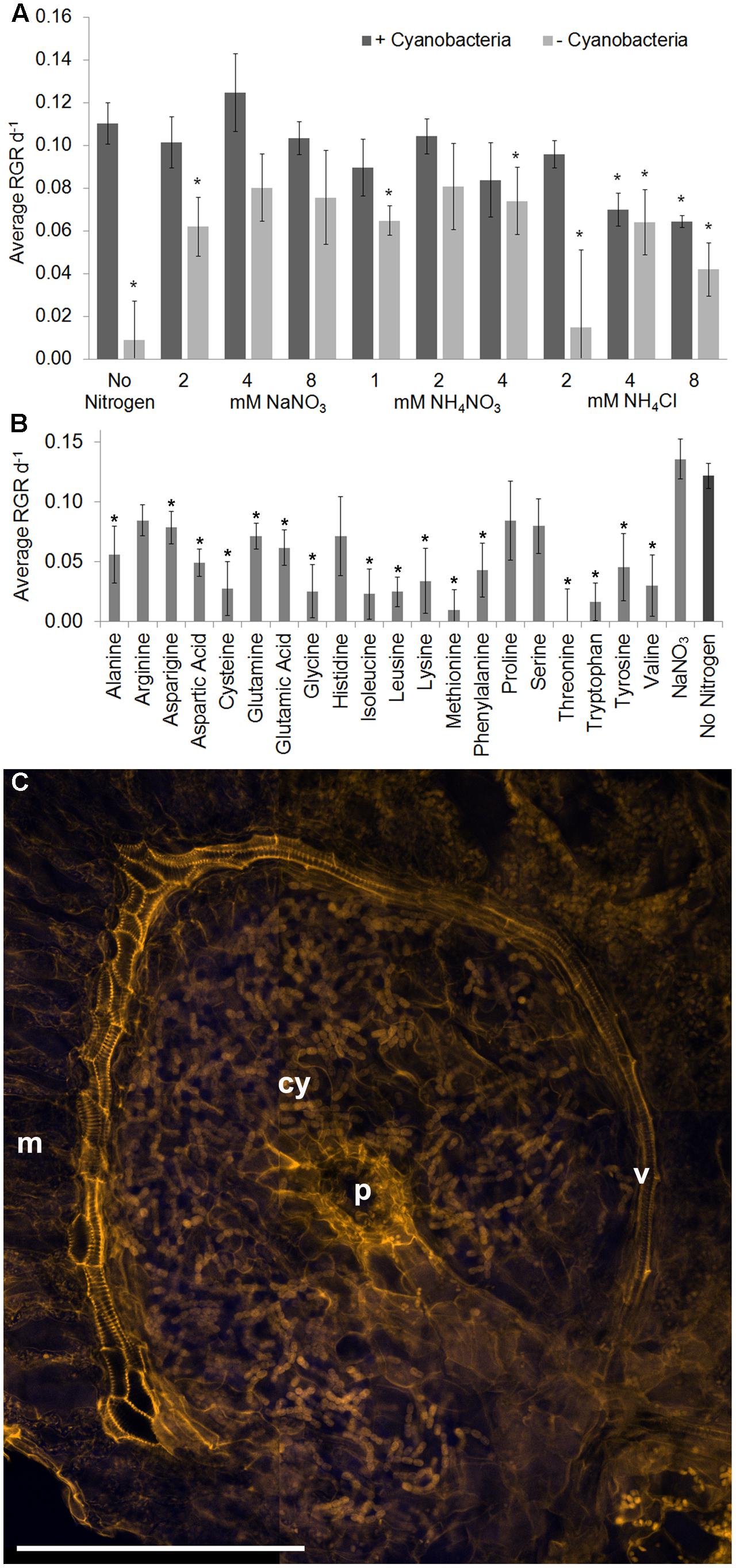
FIGURE 3. Role of N. azollae in sustaining fern growth and specific structures of the leaf pocket. Clonal ferns with (+cyano) and without N. azollae (-cyano) were grown on agar solidified Standard Azolla Medium (SAM). (A) Growth in inorganic N-fertilizer ranging from 2 to 8 mM. (B) Growth with 2 mM nitrogen in differing amino acids, or with 4 mM NaNO3 (NO3), and +cyano on SAM for comparison. Growth was measured as leaf area. Ferns without N. azollae failed to grow on SAM but in all other conditions growth fitted an exponential curve. Standard deviations with n = 3, T-test significance with reference to ferns +cyano on SAM for P < 0.05 is marked with ∗. (C) Structures revealed by propidium iodide staining using confocal microscopy 405 nm excitation, 560 nm emission (orange); auto-fluorescence at 515–530 nm emission (blue). v, vasculature; p, pore; cy, N. azollae bacteria; m, mesophyll cells; scale bar: 200 μm.
N2-Fixation Rates Reached 0.4 mg N g−1dw h−1 at Noon and Dropped 7.5 Times in the Dark
Nostoc azollae are phototrophic bacteria, therefore, we set out to measure diel 15N2 fixation in Azolla. We first established that N2-fixation rates were constant over at least 12 h when keeping light, temperature, and CO2 concentrations constant (data not shown). Ferns in excess medium were exposed to 15N2-enriched air for 2 h starting at 8, 14, 18, 20, and 8 h in the diel cycle of a growth chamber with 12 h light starting at 7 h. Ferns without N. azollae constituted the control non-fixing ferns at the 14 h time point. The relative enrichment of 15N over air (δ15N) in Azolla biomass at 20 and 2 h in the dark was up to 7.5 times lower than that measured at noon (Figure 4A). N. azollae, therefore, fixed the bulk amounts of N2 during day-time.
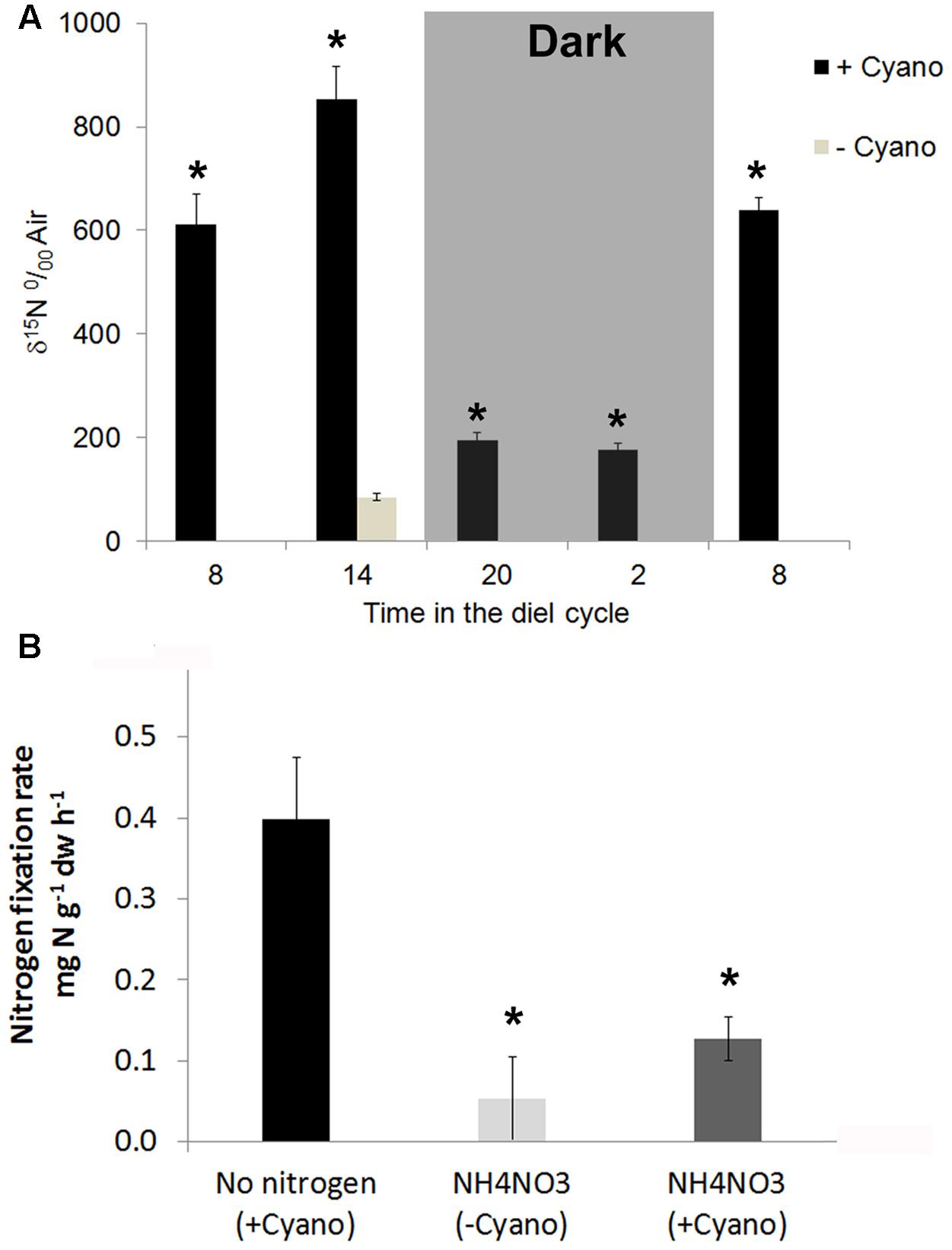
FIGURE 4. 15N2 fixation during the diel cycle and after acclimation to 2 mM NH4NO3. (A) Raw 15N enrichment after 2 h exposure to 15N2 averaged for the time points 8, 14, 20, and 2h during the diel cycle. Ferns were on SAM without nitrogen, the 12 h day started at 6 h. Temperature was kept a constant 21°C. Ferns with N. azollae (+Cyano, black) and without (–Cyano, light gray). (B) 15N2-fixation rates averaged over 2 h at the timepoint 14 h in the diel cycle after a week acclimation on SAM without [No nitrogen (+Cyano), black and with 2 mM NH4NO3 (NH4NO3 (+Cyano), dark gray]; control ferns without N. azollae [NH4NO3 (–Cyano), light gray]. Standard deviations with n = 3. T-test significance marked with ∗ was P < 0.05 for +Cyano data points with reference to ferns –Cyano on SAM for (A); and with reference to +Cyano ferns on no N in (B).
We further assessed 15N2-fixation rates in ferns acclimated for a week to +N (2 mM NH4NO3) or –N (SAM) during peak N2 fixation at 14 h (Figure 4B). Whilst the N2-fixation rate in ferns grown without nitrogen reached 0.394 mg N g−1 DW h−1, N-fertilizer reduced 15N2-fixation rates fourfold. The dramatic change further indicated that responses studied in ferns +N resulted from N-uptake rather than the combination of fixation and uptake.
Transcriptional Patterns of the Diel Cycle Dominate over the Response to N-Fertilizer
The fern’s ability to react to N-fertilizer prompted an analysis of the transcriptional mechanisms underpinning these responses; the specificity with which cDNA libraries were generated afforded stringent spatial resolution thus distinguishing for the first time processes in the fern from those in the cyanobacteria. Given the diel behavior of nitrogen assimilation genes in the angiosperm A. thaliana (Cheng et al., 1991; Gutiérrez et al., 2008), we chose to compare the diel transcript profiles of ferns acclimated for a week to SAM without nitrogen (-N) or with 2 mM NH4NO3 (+N). Ferns were sampled from triplicate cultures at 2, 8, 14, and 20 h within the 24 h cycle of a growth cabinet with 12 h light starting at 6 h in the morning. RNA-seq resulted in 17.3–37.8 million read pairs per sample of which between 47–55% mapped as read pairs and 63–65% mapped as single end reads to the transcriptome database [Brouwer et al., 2014; Supplementary Table 2_mapping_stats]. Transcriptional investments in functional categories were similar in A. filiculoides and A. thaliana (Supplementary Figure 4). Principle component analysis indicated four groups that corresponded to the different time points during the day which separated in both the first and second dimension and explained 38% of the variation in the dataset (Figure 5A). The first two dimensions also resolved the N-fertilized samples: +N samples projected apart from the –N samples in the same direction at all time points.
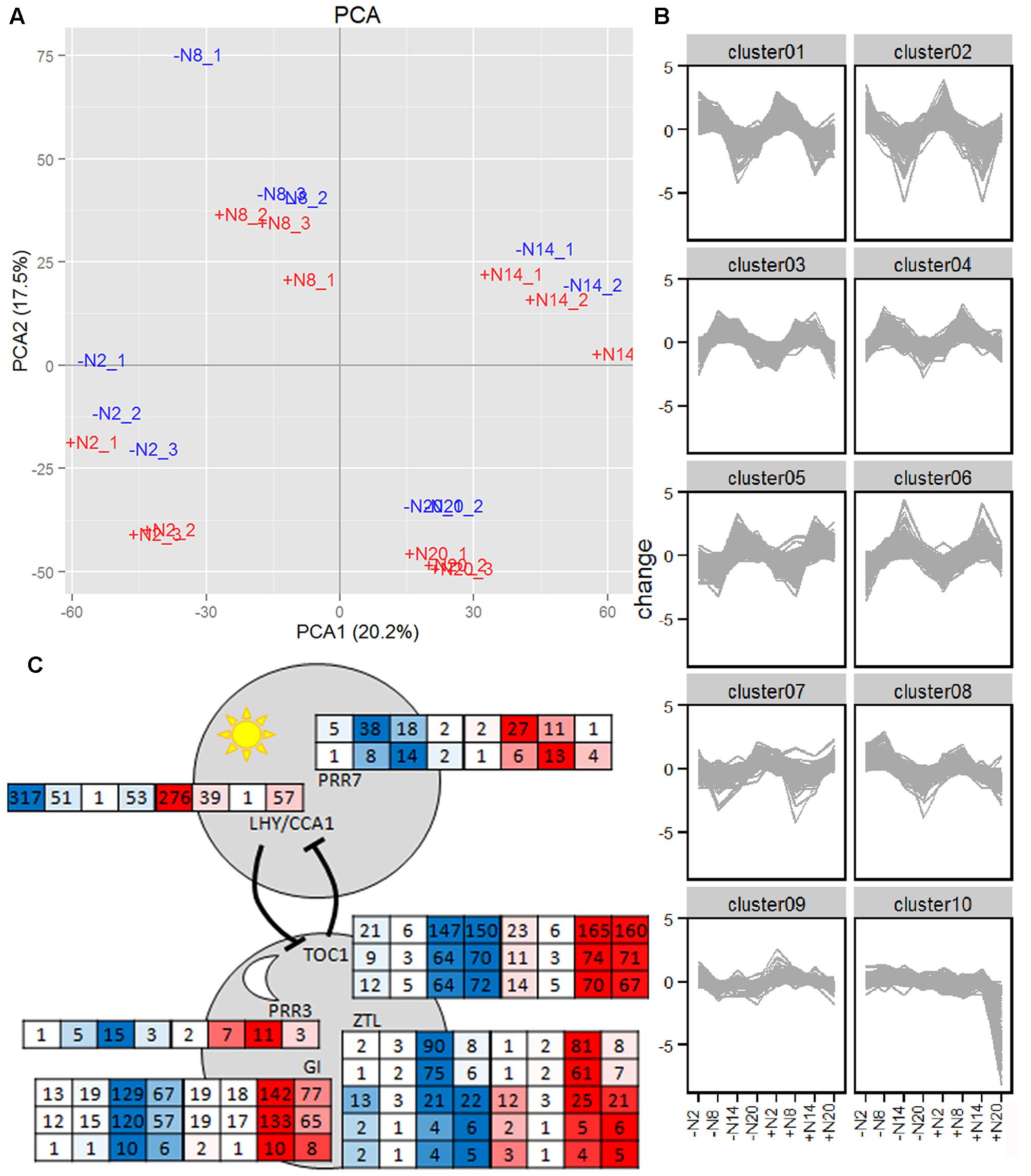
FIGURE 5. Diel transcript profiles in A. filiculoides grown with and without 2 mM NH4NO3. Ferns were acclimated to SAM without (–N, in blue) and with 2 mM NH4NO3 (+N, in red) for a week then harvested from triplicate cultures at 2, 8, 14, and 20 h with 12 h day-light starting at 6 h. RNA was extracted, sequenced and sequences analyzed as described in the section “Materials and Methods.” (A), Principal Component Analysis (PCA) and (B), K-means clustering included all genes with a read count > 10 rpm for all time points and conditions. K-means clusters were built with Pearson distance metric. (C) Clock components from the morning (above) and evening (below) loops with their respective read counts in rpm, on medium without N (in blue) and with N (in red) at 2, 8, 14, and 20 h in each box, respectively.
Diel transcripts were defined as transcripts with read counts significantly changed in at least two adjacent time points in either the +N or –N series. Of all transcripts detected, 4872 or 13% were diel (Supplementary Table 2_diel_transcripts). K-means clustering of diel genes identified clusters (Figure 5B and Supplementary Table 2_clusters) with genes peaking at 2 h (8 h into the dark, clusters 1 and 2), genes peaking at 8 h (2 h into the light, clusters 3 and 4), genes peaking at 14 h (8 h into the light, clusters 5 and 6), and genes peaking at 20 h (2 h into the dark, cluster 7) with no difference between the –N and +N samples, indicating that diel rhythms of transcription were unresponsive to N-fertilizer, with the exception of genes in cluster 10 (369 contigs) that sharply decreased 2 h into dark if N was supplemented (Figure 5B and Supplementary Table 2_cluster_10). Cluster 10 genes included genes from the Calvin-Benson cycle, PS1 and PS2 light harvesting complexes, and glycine decarboxylase P-protein 2 (Af_7035) from photorespiration; in addition it included the amino acid sensor ACT domain repeat 3-like protein (Af_11070) and three sugar transporters (Af_3233, Af_17210, and Af_14354).
Clock-gene expression patterns corresponded to those in A. thaliana with the morning loop peaking at 2 and 8 h and the evening loop peaking at 14 and 20 h although the number of isoforms did not correspond to the numbers in A. thaliana; clock-gene transcripts did not respond to N-fertilizer (Figure 5C and Supplementary Table 2_clock_genes). Diel transcripts were enriched in starch and sucrose metabolism, pathways of photosynthesis, nutrient assimilation and selected pathways in secondary, lipid, and amino acid metabolism (Supplementary Table 3). Diel rhythms of gene expression and clock components in A. filiculoides ferns were hence similar to those in angiosperms. In the fern, moreover, diel rhythms of gene expression dominated over the response to N-fertilizer.
Transcriptional Profiles of Ferns Grown without N-Fertilizer Reflected Early Morning Recovery from Night-Time N Deficiency
Acclimation to +N (2 mM NH4NO3) compared to –N changed the amounts of 526 transcripts significantly when all contigs were counted that were changed in at least one time point (Supplementary Table 2_N_responsive). The changes were, however, not uniform throughout the day: most changes occurred during darkness at 20 and 2 h with 250 and 240 changed contigs, respectively (Figure 6A). In contrast only 28 contigs were changed at noon (14 h; Figure 6A).
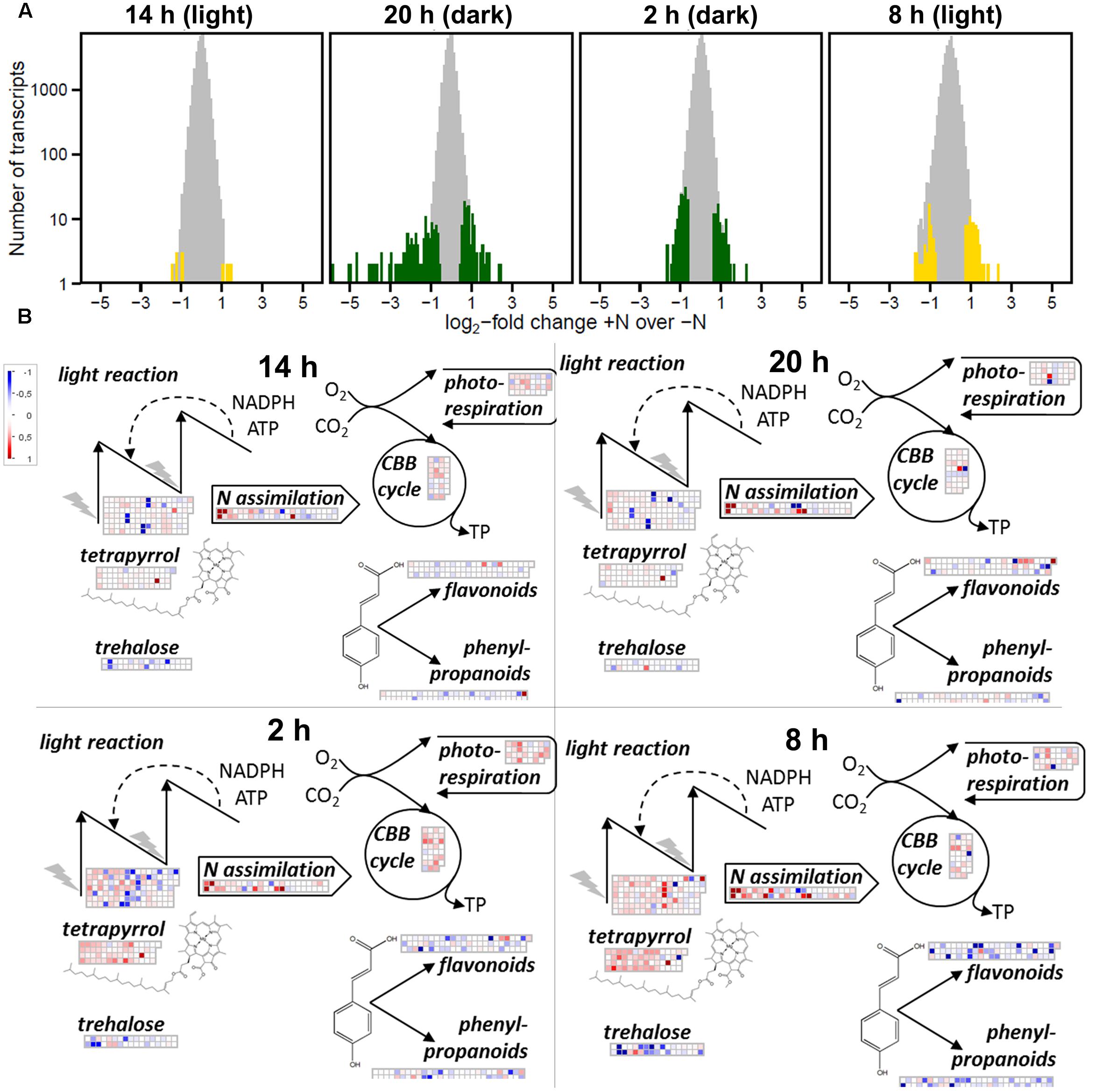
FIGURE 6. Metabolic responses of A. filiculoides to diurnal supply of reduced N from N. azollae. (A) Number of genes differentially transcribed comparing log2-fold change of the read-count ratios in ferns with 2 mM NH4NO3 (+N) over without (–N). Yellow, time points in the light; green, time points in the night. (B) Mapman overview of transcriptional changes in metabolism comparing transcription +N over –N. Noon (14 h), evening (20 h; 2 h into dark), night (2 h), and morning (8 h; 2 h into the 12 h day). Upregulation in –N is depicted in red, downregulation is depicted in blue, maximal coloration is set at twofold changes.
To visualize N-dependent transcriptional investments in central metabolic pathways, transcript read counts were submitted to the Mapman software [Figure 6B; (Thimm et al., 2004)]. In the middle of the night (2 h) and in the morning (8 h), +N conditions led to accumulation of transcripts involved in tetrapyrrole synthesis (q < 10−9), the Calvin-Benson cycle (q < 10−20), and in the morning transcripts of the light reaction (q < 10−9). In contrast, transcripts from flavonoid (q < 10−3), phenylpropanoid synthesis (q < 10−3), and trehalose metabolism (q < 10−2) accumulated in –N conditions, most strongly in the morning at 8 h (Figure 6B and Supplementary Table 4). This pattern essentially reflected the pattern reported in N-starved A. thaliana upon nitrate addition (Scheible et al., 2004) and therefore suggested N deficit at night in ferns grown in the absence of N.
Pathways for the Assimilation of N into Amino Acids Remained Unchanged but Ureic Acid Metabolism Changed with N Supply
Transcriptional investment in N-assimilation pathways into amino acids, remained unchanged when comparing +N with –N with very few exceptions: transcripts accumulated for two contigs of glutamine dehydrogenase (Af2747 and Af3257) in ferns on +N but were much less abundant than transcripts of GOGAT (Af4692) and GS (Af31780) enzymes (Table 1 and Supplementary Figure 5), and therefore may represent enzymes in specific tissues exposed to +N from the medium.
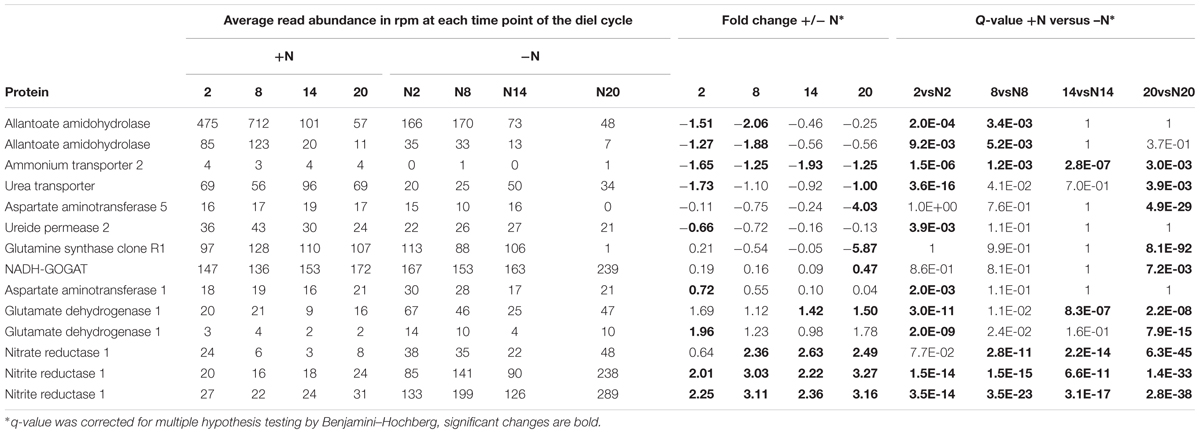
TABLE 1. Transcripts associated with N metabolism that responded to +/-N conditions at the time points 2, 8, 14, or 20 h of the diel cycle.
At night in plants grown –N, stored ureides may have been remobilized by allantoate amidohydrolase (Af8889) as reads from this enzyme were abundant. In contrast at night in ferns grown +N, reads of this enzyme were low (Table 1). Consistently in ferns grown +N at night, N-remobilization likely was decreased since transcripts of a transporter of urea (Af13203) and ureide permease (Af5356) were fewer whilst urea biosynthesis likely was maintained since transcripts of the aspartate aminotransferase 1 (Af3010) and NADH-dependent GOGAT (Af4692) accumulated (Table 1).
Genes affected by +N throughout the day included the nitrite and nitrate reductase genes (Af1086, Af13194, and Af36782, respectively; Table 1) and confirm that fern metabolism responded to nitrate in the medium. Similarly increased transcript of the HPP family nitrite transporter (Af1275; Supplementary Table 2_N_responsive) in ferns +N suggested increased nitrite transport to chloroplasts in fern cells; this fern gene is of cyanobacterial origin and was reported absent in mosses (Maeda et al., 2014) In contrast, increased transcript abundance in ferns –N of an AMT2 type ammonium transporter (Af12800; Table 1), two phosphate transporters homologous to AtPHT1 and AtPHT3 (Af380, Af1508; Supplementary Table 2_N_responsive), and a Nodule INception (NIN) transcription factor with homology to AtNLP7 (Af7964; Supplementary Table 2_N_responsive) suggest that ferns with active N2 fixation regulate P and N transport coordinately as known from the angiosperm symbioses with arbuscular mycorrhiza (Breuillin-Sessoms et al., 2015). Manual assembly of AMT transporter sequences allowed construction of a phylogenetic tree showing that the fern AMT transporters are both of the MEB (AMT2 type in A. thaliana which is electroneutral) and AMT (AMT1 type in A. thaliana) clades as defined by [Neuhäuser et al., 2009; McDonald and Ward, 2016; Figure 7]; AMT2 had increased read counts in ferns with actively N2-fixing cyanobacteria (Table 1). N. azollae AMT clustered with the bacterial AmtB clade, yet the N. azollae AmtB that is functional was not in the genome location of closely related strains which was inactivated by a mobile element (Supplementary Figure 6).
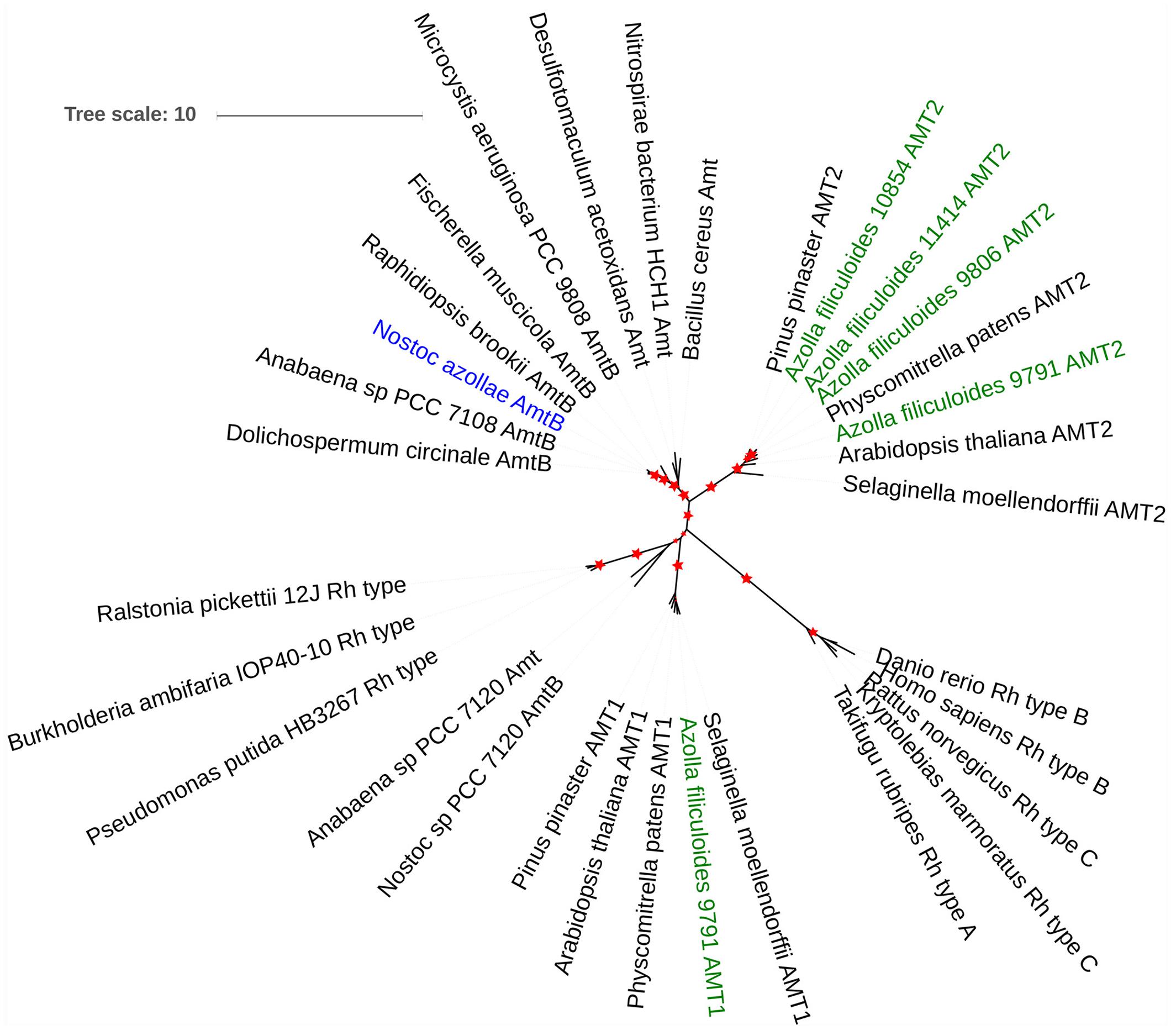
FIGURE 7. The origin of ammonium transporters from A. filiculoides and N. azollae. Sequences were manually assembled (Supplementary List of AMT Protein Sequences) then aligned with MUSCLE, the phylogenetic tree calculated using PhyML with 100 bootstraps (Dereeper et al., 2008) and then visualized using iTol (Letunic and Bork, 2016), bootstrap values ranging from 70 to 100% are represented by the increasing red star sizes.
Key Genes of the Vasculature Were Upregulated under –N Growth Conditions
Xylem was a prominent feature of the vasculature curving around the leaf pocket (Figure 3C). This specialized vasculature may have as much a structural function as a nutrient and water transport function. In ferns –N compared to +N, transcripts of fern homologs of the xylem cysteine peptidase accumulated (XCP3, Af1829) and of the CYTOKININ OXIDASE decreased (Af34233), suggesting active xylem formation (Supplementary Table 2_N_responsive). Further investigation of xylem specific genes revealed homologs of VND6 and LBD15 as well as IRX3 in the A. filiculoides sequences. Phloem cells lined the xylem cells for most of the vasculature curving around the leaf pocket (Figure 3C and Supplementary Figures 3B,C). To test a possible role of vasculature in the response to differential N supply, full length gene-homologs characteristic for the vasculature were manually assembled then reads mapped to quantify differential accumulation of the mRNA with more accuracy. Transcripts of a fern homolog to AtGL22 were amongst the most differentially accumulating transcripts (13 × averaged over all time points) under conditions −N compared to +N, GL22 is known from A. thaliana to regulate allocation of phloem nutrient contents.
Discussion
Light-Driven N2 Fixation by N. azollae Entirely Supported the High Yields of Azolla Biomass
Clones of A. filiculoides without N. azollae required N-fertilizer for growth and inorganic nitrogen sufficed to reach growth rates nearing those of clones with symbionts (Figure 3B). N. azollae supplied sufficient N to support high biomass yields with little gain from N-fertilizer (Figure 1B). 15N2-fixation rates for Azolla on medium without N-fertilizer peaked at 0.4 mg N g−1 DW h−1 during midday but averaged 0.17 mg g−1 DW h−1 over the diel cycle; this corresponded to the N increase in the biomass, reaching 0.33 g m−2 d−1 in our experiments or 0.15 g−1 DW h−1. To compare, 15N2-fixation rates in soybean averaged 0.08 mg g−1 DW h−1 (Hung et al., 2013). Diel variation of N2 fixation in clover (Medicago truncatula/Sinorhizobium meliloti) was small and entirely dependent upon temperature (Cabeza et al., 2015). The diel variation reported here for A. filiculoides was measured under constant temperature and therefore the high N2-fixation was diurnal, and depended on light.
Nostoc azollae is a filamentous cyanobacterium. Typically N2-fixation is sequestered to heterocysts in filamentous cyanobacteria and occurs mostly during the day when, firstly, cells of the filament supply carbon to fuel heterocyst glucose-6-phosphate dehydrogenase (G6PDH) providing reductant NADPH and, secondly, light energy captured by heterocyst photosystem I supplies ATP; G6PDH was required for N2-fixation and growth of Nostoc sp. in the dark (Gallon, 1981; Summers et al., 1995). N. azollae lack glycolysis pathway enzymes and therefore may channel all carbon through G6PDH to generate NADPH required to reduce N2; the N. azollae genome retained PS1 for ATP production in light (Ran et al., 2010). In contrast, heterotrophic Sinorhizobium meliloti fixes N2 in the darkness of root nodules and therefore requires carbohydrates to synthesize NADPH as well as ATP: malate provided by the plant provides NADPH via malic enzyme, and is further respired via oxidative phosphorylation specialized for micro-aerobic environments. The malic enzyme gene is required for N2 fixation by Rhizobium meliloti (Driscoll and Finan, 1993). These differences may underlie the higher N2-fixation rates observed here with phototrophic diazotrophic symbionts in Azolla compared to heterotrophic diazotrophic symbionts in legumes.
Azolla Yields without N-fertilizer Input Competed Favorably with Established Plant Protein Crops
Nitrate fertilizer at 4 mM increased productivity of Azolla by merely 15% (Figure 2A) compared to 175 and 132% increase reported when 5 mM NO3− was administered to elite breed soybeans 40 and 60 days after planting (Yashima et al., 2003). RNAseq and quantitative RT-PCR revealed that the fern responded to exogenous NO3− with the accumulation of transcripts from nitrite and nitrate reductase (Figures 2A,C and Table 1), and a nitrite transporter (Supplementary Table 2) consistent with previous data of A. pinnata showing that labeled NO3− is incorporated in roots and then is seen to move slowly to shoots (Ito and Watanabe, 1983). Exogenous NH4NO3 at 2 mM resulted in a 78.4% decrease in 15N2-fixation by the symbiont (Figure 4B). Similarly, N2 fixation decreased by 20–70% in 99 different strains of Azolla on 2.86 mM (NH4)2SO4 (Okoronkwo et al., 1989). This may be due to diffusion of external NH4 into the leaf pocket: complete inhibition of N2 fixation took place with 1 mM NH4+ but not NO3− in the related free-living Anabaena cylindrica (Ohmori and Hattori, 1974). Alternatively, the fern may contribute to actively regulate N2-fixation in the symbiont by controlling gas exchange via the leaf-pocket pore (Veys et al., 1999) and O2 release. Increased O2 exposure as evidenced by increased PS2 and hemoglobin expression in ferns +N further suggest this (Supplementary Table 2); NO3− or urea from the fern unlikely affect N2 fixation in the symbiont directly since genes for import and assimilation of NO3− and urea were reported missing in N. azollae (Ran et al., 2010). N2 fixation by rhizobia in the nodules of legumes is also inhibited by NH4+: NH4SO4 at 5 mM rapidly decreased N2 fixation in alfalfa roots exposed over a period of 5 days (Cabeza et al., 2015). In plant symbioses with either cyanobacteria or rhizobia, therefore, the N-substrate exchanged when present in the surrounding medium inhibits N2-fixation of the bacteria.
In contrast to the large majority of agricultural crops, Azolla species grow in shallow surface freshwater. Once the surface of the water was covered by exponential growth, our experimental farming system produced biomass linearly and stably over periods exceeding 130 days (Figure 1B). Given a measured N-content of 3.5% w/w biomass DW and 35 t ha−1 annual productivity potential without any N-input (Figures 1, 2), A. filiculoides has the potential to fix over 1200 kg N ha−1 a−1 and therefore should be considered as a sustainable plant protein crop in temperate regions. Whilst freshwater aquaculture of macro- and microalgae exceed the yield potential in biomass and protein of the floating Azolla, reaching over 100 t ha−1 a−1 with similar nitrogen content, they have not been studied for their N-fixation capacity, with exception of the cyanobacteria (Neveux et al., 2015). They are generally cultured for wastewater treatment because they require high nutrient loads to reach these productivities. As in the case of Azolla more ecological work is needed to be able to reap the benefit of their characteristics and make the most of associated microbiomes for sustainable aquaculture in specific settings (Ramanan et al., 2016). Azolla aquaculture appears particularly suited to halt land subsidence in the Dutch lowlands, for example: the dried peat-richt lands now used as grazing and cropping fields emit large amounts of CO2 from respiration causing “double trouble” (Erkens et al., 2016). Azolla biomass grown on the flooded lowlands has the potential to substitute forage as well as other crops currently grown at lower productivities and be more sustainable stopping N-fertilizer pollution, CO2 emmisions and land subsidence. To compare, established forage crops that still require some 150 kg ha−1 a−1 N-fertilizer to reach 25 t ha−1 a−1 fix some 600 kg N ha−1 a−1 in the harvested material (de Visser et al., 2014; Anglade et al., 2015). Modern soybean varieties yielding 5 t beans ha−1 with no added fertilizer to the soil fixed up to 300 kg N ha−1 per crop (Gelfand and Robertson, 2015). Our experimental system aimed at industrialization and thus was closed and entirely controlled. Promising yields, however, have been reported in an open system of 468 m2 in the tropics (Colombia), where A. filiculoides yielded 39 t ha−1 a−1; albeit in this study nitrogen-containing chicken manure was supplied to the fern (Becerra et al., 1990). Feeding Azolla biomass to pigs as protein-rich admixtures to soymeal supported growth as well as control soymeal diets (Becerra et al., 1990; Leterme et al., 2009, 2010). Continuous production is particularly suited for on-farm processing as it requires processing units of smaller capacity. Azolla ferns efficiently use run-off water from fields and remove both nitrogen and phosphate (Shilton et al., 2012), thereby preventing eutrophication if harvested regularly and contributing to closing the nitrogen and phosphate cycles. If grown as dense mats the ferns may help re-solve iron oxides, while releasing adsorbed phosphate and therefore be an alternative to mining increasingly rare phosphate fertilizer from sediments of shallow freshwater (Cordell et al., 2011). The findings reported here warrant a detailed assessment of potential (agro-) ecosystem benefits and services of Azolla farming, including potential threats to protected wetland ecosystems.
Specific Physiological Adaptations of the Ferns to the Diurnal Cyanobacterial N2 Fixation
If hosting cyanobacteria is such an efficient solution to fixing large amounts of N2, we wondered why taxa hosting cyanobacterial N2 fixation are comparatively rare and what adaptations would be required by the host plants.
Since N2 fixation was mostly light driven, the fern host alternated between high N availability during the day and low N availability during the night; low N availability may be restricted to the later part of the night due to buffering by the high NH4+ concentrations in the N. azollae and leaf-pocket fluid (Kaplan and Peters, 1981; Meeks et al., 1987). Consequently, the transcriptional patterns differed in A. filiculoides supplemented with NH4NO3 and without. The morning time point without NH4NO3 (Figure 6B) displayed transcripts highly reminiscent of those in A. thaliana when supplied with nitrate after N-starvation (Scheible et al., 2004). The responses to low N availability were therefore conserved between ferns and seed plants. The transcriptional response to low nitrogen availability disappeared during the day with the noon time point having the fewest changes between +N and –N conditions and the highest N. azollae N2-fixation rates. The magnitude of the fern transcriptional response to N-fertilizer was therefore much lower than that reported for spermatophytes entirely dependent on externally available N (Wang et al., 2003; Vidal et al., 2013). As N supply was short during the night, it is likely that Azolla possess a storage system for N which balances availability during the day and the night, this could have been reflected by the prominence of urea transporters in the list of genes accumulating in ferns without N-fertilizer. Night-time accumulation of transcripts encoding enzymes of allantoin metabolism in ferns without N-fertilizer points to allantoin as an intermediate in N storage. To host phototrophic diazotrophic cyanobacteria, therefore, ferns adapted existing responses required to cope with irregular supply of N-fertilizer from the environment that evolved in ancestors common to ferns and seed plants.
Features That Ensure Metabolic Connectivity between Host and Cyanobacteria
Streptophyte algae and bryophytes were found to regularly associate with N2-fixing cyanobacteria as well as rhizobia, suggesting that evolution of land plants, including the fern Azolla, happened after mechanisms important for both these interactions evolved (Knack et al., 2015). Recognition of specific cyanobacteria evolved with chloroplast endosymbiosis, but recognition competency may have been lost and subsequently acquired again with a novel mechanism by the common ancestor of vascular plants. Mechanisms employed by the seed plants Gunnera and Blasia to control development of N. punctiforme hormogonia may therefore also occur in Azolla ferns (Liaimer et al., 2015). Specific regulators and transporters for the exchange of nutrients with intracellular organelles such as the chloroplasts may not have been recruited for extracellular symbiosis, but instead have evolved from extracellular interactions such as those with arbuscular mycorrhiza, rhizobia or, Nostoc species. This would allow differential regulation of the transport processes. The AMT2 and PHT1 family transporters and the NIN transcription factor transcripts seen to accumulate in Azolla actively fixing N2 could therefore play a role in the interaction with N. azollae. Active mobile elements inside N. azollae likely contributed to novel regulation of the ammonium transporter expression required for the symbiosis (Supplementary Figure 5) in addition to the metabolic adaptations such as the lacking glycolysis pathway reported earlier (Ran et al., 2010).
The most striking adaptation of the fern to N. azollae is the enclosed leaf-pocket organ with its specific pore, leaf-pocket hair cells, and extensive curving xylem-rich vasculature (Figure 3 and Supplementary Figures 3B,C). The pore structure is likely important for gas exchange; it is lined with specific teat cells possibly having a role in defense against invading organisms and water repulsion (Veys et al., 1999, 2002). The hair cells have been proposed to resemble transfer cells based on EM-cytology detection of dense cytoplasm adjacent to wall ingrowths and abundant ER with ribosomes and mitochondria, and therefore play a role in nutrient exchange (Duckett et al., 1975) consistent with their close contact with the phloem (Supplementary Figure 3B). Finally, the prominent and specific development of xylem cells surrounding the leaf pockets was consistent with a water conducing function; stark re-enforcement of the tracheids furthermore revealed a structural role of the xylem cells not generally described in ferns (Figure 3C). Presence of VND6 in Azolla suggested that these NAC factors control secondary wall thickening in ferns as in seed plants (Lucas et al., 2013). Stark accumulation of transcripts related to vasculature, and nutrient transport under conditions requiring N2 fixation (manually assembled homologs to AtGL22, 13x and AtXCP1, 9x) suggested that vasculature functions are essential for nutrient supply to cyanobacteria, given that the leaf pockets are enclosed structures that do not communicate with surrounding medium. Hosting cyanobacteria in a leaf therefore may require exquisite control over water and nutrient supply as well as structural re-enforcements and a specialized pore structure so as to hold the heavy pool with bacteria in the air whilst at the same time ensuring light and gas supply.
The Evening Loop of the Clock is Ancestral to Ferns and Seed Plants
Differential diel transcript accumulation was found for 13% of the fern genes. The types of genes which changed and the magnitude of changes observed here were comparable to those changing in diel cycles in spermatophytes (Bläsing et al., 2005) and occurred in all diel patterns (Figure 6), hence the combined output of clock and diel signals such as temperature and light results in similar outputs among vascular plants. Among the transcription factors, the CONSTANS-like group of C2C2 transcription factors was enriched in both A. filiculoides and A. thaliana diurnal transcripts confirming their ancient role in diurnal gene expression. The best blast match to the C2C2 CONSTANS type transcription factor of Chlamydomonas reinhardtii Cre06.g278159, is also diurnal [Zones et al., 2015 in Supplementary Table 16]. Nitrogen fertilizer does not influence the diurnal gene expression in A. thaliana (Bläsing et al., 2005) and did so only for a very limited number of transcripts in A. filiculoides (Figure 5B), which may have been a result of diel variability in N-supply in –N ferns rather than less N availability per se (Figure 4). The similarity also points to an evolutionary origin of the diel patterns before the split of fern and seed plants. While no diel transcriptome analyses from mosses were available for comparison, the analyses of the single celled C. reinhardtii show both different and overlapping patterns. Gene expression in the alga was largely diel with ∼80% of genes in diel rhythms (Zones et al., 2015) compared to only 13% in this study and 10–30% depending on criteria in A. thaliana (Bläsing et al., 2005); the difference may reflect asymmetric coupling of tissue specific clocks in multicellular organisms (Endo et al., 2014).
Azolla filiculoides harbored the classical clock of seed plants (Figure 5C). Mosses have been shown to only contain a single loop of the clock comprising the morning but not the evening loop (Holm et al., 2010), while some algae contain various clock components including those of the evening loop (Noordally and Millar, 2014). In A. thaliana vasculature enriched transcripts were mostly controlled by the evening loop, whilst the mesophyll cell transcripts were mostly controlled by the morning loop; also the vasculature clock was shown to override that of mesophyll cells (Endo et al., 2014). In the ferns, whilst the evening loop was complete with TOC1, ZTL, GI, and PRR3, some detected with multiple transcripts in the original assembly (Brouwer et al., 2014), the morning loop was less complex compared to A. thaliana (Figure 5C). Possibly, therefore, the ancestor of vascular plants evolved an evening loop with a function in the vasculature.
Author Contributions
PB, ABr, VB, AvdW, AT, ABo, and HS planned, designed the research and carried it out; ABr, US, and PB carried out the bioinformatics analyses. Confocal images were from VB. All authors, including G-JR, BU, and AW discussed the results and commented on the manuscript that HS, PB, and ABr drafted. The final version of the manuscript was approved by all authors.
Funding
We acknowledge funding from the European Institute of Technology for the Climate KIC Pathfinder project AzoFaST and from the Deutsche Forschungsgemeinschaft (grants EXC 1028 and WE 2231/9-2).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Arnold van Dijk for expert technical support at the Organic Geochemistry Department, Utrecht University. We further thank Dr. U Ganeteg at the Umeå Plant Science Centre, Department or Forest Genetics and Plant Physiology, Swedish University of Agricultural Sciences, SE-901 83 Umeå, Sweden, for help with the interpretation of Azolla growth rates on amino acids.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.00442/full#supplementary-material
Footnotes
References
Anglade, J., Billen, G., and Garnier, J. (2015). Relationships for estimating N2 fixation in legumes: incidence for N balance of legume-based cropping systems in Europe. Ecosphere 6, art37. doi: 10.1890/ES14-00353.1
Becerra, M., Murgueitio, E., Reyes, G., and Preston, T. R. (1990). Azolla filiculoides as partial replacement for traditional protein supplements in diets for growing-fattening pigs based on sugar cane juice. Livest. Res. Rural Dev. 2, 15–22.
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. 57, 289–300.
Bläsing, O. E., Gibon, Y., Günther, M., Höhne, M., Morcuende, R., Osuna, D., et al. (2005). Sugars and circadian regulation make major contributions to the global regulation of diurnal gene expression in Arabidopsis. Plant Cell 17, 3257–3281. doi: 10.1105/tpc.105.035261
Breuillin-Sessoms, F., Floss, D. S., Gomez, S. K., Pumplin, N., Ding, Y., Levesque-Tremblay, V., et al. (2015). Suppression of arbuscule degeneration in Medicago truncatula phosphate transporter4 mutants is dependent on the ammonium transporter 2 family protein AMT2; 3. Plant Cell 27, 1352–1366. doi: 10.1105/tpc.114.131144
Brouwer, P., Bräutigam, A., Külahoglu, C., Tazelaar, A. O. E., Kurz, S., Nierop, K. G. J., et al. (2014). Azolla domestication towards a biobased economy? New Phytol. 202, 1069–1082. doi: 10.1111/nph.12708
Cabeza, R. A., Liese, R., Fischinger, S. A., Sulieman, S., Avenhaus, U., Lingner, A., et al. (2015). Long-term non-invasive and continuous measurements of legume nodule activity. Plant J. 81, 637–648. doi: 10.1111/tpj.12751
Carrapiço, F. (1991). Are bacteria the third partner of the Azolla-Anabaena symbiosis? Plant Soil 137, 157–160. doi: 10.1007/BF02187448
Cheng, C. L., Acedo, G. N., Dewdney, J., Goodman, H. M., and Conkling, M. (1991). Differential expression of the two Arabidopsis nitrate reductase genes. Plant Physiol. 96, 275–279. doi: 10.1104/pp.96.1.275
Cordell, D., Rosemarin, A., Schröder, J. J., and Smit, A. L. (2011). Towards global phosphorus security: a systems framework for phosphorus recovery and reuse options. Chemosphere 84, 747–758. doi: 10.1016/j.chemosphere.2011.02.032
de Visser, C. L. M., Schreuder, R., and Stoddard, F. (2014). The EU’s dependency on soya bean import for the animal feed industry and potential for EU produced alternatives. OCL 21, D407. doi: 10.1051/ocl/2014021
de Vries, J., Fischer, A.M., Roettger, M., Rommel, S., Schluepmann, H., Brautigam, A., et al., (2016). Cytokinin-induced promotion of root meristem size in the fern Azolla supports a shoot-like origin of euphyllophyte roots. New Phytol. 209, 705–720. doi: 10.1111/nph.13630
Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., et al. (2008). Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 1, W465–W469. doi: 10.1093/nar/gkn180
Dos Santos, P. C., Fang, Z., Mason, S. W., Setubal, J. C., and Dixon, R. (2012). Distribution of nitrogen fixation and nitrogenase-like sequences amongst microbial genomes. BMC Genomics 13:162. doi: 10.1186/1471-2164-13-162
Driscoll, B. T., and Finan, T. M. (1993). NAD+-dependent malic enzyme of Rhizobium meliloti is required for symbiotic nitrogen fixation. Mol. Microbiol. 7, 865–873. doi: 10.1111/j.1365-2958.1993.tb01177.x
Duckett, J. G., Toth, R., and Soni, S. L. (1975). An ultrastructural study of the Azolla, Anabaena azollae relationship. New Phytol. 75, 111–118. doi: 10.1111/j.1469-8137.1975.tb01377.x
Endo, M., Shimizu, H., Nohales, M. A., Araki, T., and Kay, S. A. (2014). Tissue-specific clocks in Arabidopsis show asymmetric coupling. Nature 515, 419–422. doi: 10.1038/nature13919
Erisman, J. W., Sutton, M. A., Galloway, J., Klimont, Z., and Winiwarter, W. (2008). How a century of ammonia synthesis changed the world. Nat. Geosci. 1, 636–639. doi: 10.1038/ngeo325
Erkens, G., Meulen, M. J., and Middelkoop, H. (2016). Double trouble: subsidence, and CO2 respiration due to 1,000 years of Dutch coastal peatlands cultivation. Hydrogeol. J. 24, 551–568. doi: 10.1007/s10040-016-1380-4
Evenson, R. E., and Gollin, D. (2003). Assessing the impact of the green revolution, 1960 to 2000. Science 300, 758–762. doi: 10.1126/science.1078710
Forni, C. I., Tel-Or, E., Bar, E., and Grilli-Caiola, M. (1991). “Effects of antibiotic treatments on Azolla-Anabaena and Arthrobacter,” in Nitrogen Fixation, eds M. Polsinelli, R. Materassi, and M. Vincenzini (Dordrecht: Springer), 151–156. doi: 10.1007/978-94-011-3486-6_95
Forsum, O., Svennerstam, H., Ganeteg, U., and Näsholm, T. (2008). Capacities and constraints of amino acid utilization in Arabidopsis. New Phytol. 179, 1058–1069. doi: 10.1111/j.1469-8137.2008.02546.x
Freiberg, C., Fellay, R., Bairoch, A., Broughton, W. J., Rosenthal, A., and Perret, X. (1997). Molecular basis of symbiosis between Rhizobium and legumes. Nature 387, 394–401. doi: 10.1038/387394a0
Gallon, J. R. (1981). The oxygen sensitivity of nitrogenase: a problem for biochemists and micro-organisms. Trends Biochem. Sci. 6, 19–23. doi: 10.1016/0968-0004(81)90008-6
Galloway, J. N., Dentener, F. J., Capone, D. G., Boyer, E. W., Howarth, R. W., Seitzinger, S. P., et al. (2004). Nitrogen cycles: past, present, and future. Biogeochemistry 70, 153–226. doi: 10.1007/s10533-004-0370-0
Gelfand, I., and Robertson, G. P. (2015). A reassessment of the contribution of soybean biological nitrogen fixation to reactive N in the environment. Biogeochemistry 123, 175–184. doi: 10.1007/s10533-014-0061-4
Gutiérrez, R., Stokes, T. L., Thum, K., Xu, X., Obertello, M., Katari, M. S., et al. (2008). Systems approach identifies an organic nitrogen-responsive gene network that is regulated by the master clock control gene CCA1. Proc. Natl. Acad. Sci. U.S.A. 105, 4939–4944. doi: 10.1073/pnas.0800211105
Hartman, G. L., West, E. D., and Herman, T. K. (2011). Crops that feed the World 2. Soybean—worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 3, 5–17. doi: 10.1007/s12571-010-0108-x
Herendeen, P. S., Magallon-Puebla, S., Lupia, R., Crane, P. R., and Kobylinska, J. (1999). A Preliminary conspectus of the allon flora from the late cretaceous (Late Santonian) of Central Georgia, U.S.A. on JSTOR. Ann. Mo. Bot. Gard. 86, 407–471. doi: 10.2307/2666182
Holm, K., Källman, T., Gyllenstrand, N., Hedman, H., and Lagercrantz, U. (2010). Does the core circadian clock in the moss Physcomitrella patens (Bryophyta) comprise a single loop? BMC Plant Biol. 10:109. doi: 10.1186/1471-2229-10-109
Hung, N. V. P., Watanabe, S., Ishikawa, S., Ohtake, N., Sueyoshi, K., Sato, T., et al. (2013). Quantitative analysis of the initial transport of fixed nitrogen in nodulated soybean plants using 15N as a tracer. Soil Sci. Plant Nutr. 59, 888–895. doi: 10.1080/00380768.2013.838742
Ito, O., and Watanabe, I. (1983). The relationship between combined nitrogen uptakes and nitrogen fixation in Azolla-Anabaena symbiosis. New Phytolo. 95, 647–654. doi: 10.1111/j.1469-8137.1983.tb03528.x
Kaplan, D., and Peters, G. A. (1981). The Azolla-Anabaena azollae relationship. X. 15N2 fixation and transport in main stem axes. New Phytol. 89, 337–346. doi: 10.1111/j.1469-8137.1981.tb07494.x
Kistner, C., and Parniske, M. (2002). Evolution of signal transduction in intracellular symbiosis. Trends Plant Sci. 7, 511–518. doi: 10.1016/S1360-1385(02)02356-7
Knack, J. J., Wilcox, L. W., Delaux, P. M., Ané, J. M., Piotrowski, M. J., Cook, M. E., et al. (2015). Microbiomes of streptophyte algae and bryophytes suggest that a functional suite of microbiota fostered plant colonization of land. Int. J. Plant Sci. 176, 405–420. doi: 10.1086/681161
Leterme, P., Londoño, A. M., Muñoz, J. E., Súarez, J., Bedoya, C. A., Souffrant, W. B., et al. (2009). Nutritional value of aquatic ferns (Azolla filiculoides Lam. and Salvinia molesta Mitchell) in pigs. Ani. Feed Sci. Technol. 149, 135–148. doi: 10.1016/j.anifeedsci.2008.04.013
Leterme, P., Londoño, A. M., Ordoñez, D. C., Rosales, A., Estrada, F., Bindelle, J., et al. (2010). Nutritional value and intake of aquatic ferns (Azolla filiculoides Lam. and Salvinia molesta Mitchell.) in sows. Anim. Feed Sci. Technol. 155, 55–64. doi: 10.1016/j.anifeedsci.2009.10.002
Letunic, I., and Bork, P. (2016). Interactive tree of life (iTOL) v3: an online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 44, W242–W245. doi: 10.1093/nar/gkw290
Liaimer, A., Helfrich, E. J., Hinrichs, K., Guljamow, A., Ishida, K., Hertweck, C., et al. (2015). Nostopeptolide plays a governing role during cellular differentiation of the symbiotic cyanobacterium Nostoc punctiforme. Proc. Natl. Acad. Sci. U.S.A. 112, 1862–1867. doi: 10.1073/pnas.1419543112
Liu, Y., Wan, K. Y., Tao, Y., Li, Z. G., Zhang, G. S., Li, S. L., et al. (2013). Carbon dioxide flux from rice paddy soils in central china: effects of intermittent flooding and draining cycles. PLoS ONE 8:e56562. doi: 10.1371/journal.pone.0056562
Lohse, M., Nagel, A., Herter, T., May, P., Schroda, M., Zrenner, R., et al. (2014). Mercator: a fast and simple web server for genome scale functional annotation of plant sequence data. Plant Cell Environ. 37, 1250–1258. doi: 10.1111/pce.12231
Lucas, W. J., Groover, A., Lichtenberger, R., Furuta, K., Yadav, S. R., Helariutta, Y., et al. (2013). The plant vascular system: evolution, development and Functions. J. Integr. Plant Biol. 55, 294–388. doi: 10.1111/jipb.12041
Maeda, S. I., Konishi, M., Yanagisawa, S., and Omata, T. (2014). Nitrite transport activity of a novel HPP family protein conserved in cyanobacteria and chloroplasts. Plant Cell Physiol. 55, 1311–1324. doi: 10.1093/pcp/pcu075
McDonald, T. R., and Ward, J. M. (2016). Evolution of electrogenic ammonium transporters (AMTs). Front. Plant Sci. 7:352. doi: 10.3389/fpls.2016.00352
Meeks, J. C. (2009). Physiological adaptations in nitrogen fixing Nostoc-plant symbiotic associations. Microbiol. Monogr. 8, 181–205. doi: 10.1007/7171_2007_101
Meeks, J. C., Steinberg, N. A., Enderlin, C. S., Joseph, C. M., and Peters, G. A. (1987). Azolla-Anabaena relationship: XIII. Fixation of [13N]N2. Plant Physiol. 84, 883–886. doi: 10.1104/pp.84.3.883
Metzgar, J. S., Schneider, H., and Pryer, K. M. (2007). Phylogeny and divergence time estimates for the fern genus Azolla (Salviniaceae). Int. J. Plant Sci. 168, 1045–1053. doi: 10.1086/519007
Nakamichi, N. (2011). Molecular mechanisms underlying the Arabidopsis circadian clock. Plant Cell Physiol. 52, 1709–1718. doi: 10.1093/pcp/pcr118
Neuhäuser, B., Dynowski, M., and Ludewig, U. (2009). Channel-like NH3 flux by ammonium transporter AtAMT2. FEBS Lett. 583, 2833–2838. doi: 10.1016/j.febslet.2009.07.039
Neveux, N., Magnusson, M., Maschmeyer, T., Nys, R., and Paul, N. A. (2015). Comparing the potential production and value of high-energy liquid fuels and protein from marine and freshwater macroalgae. Global Change Biol. Bioenergy 7, 673–689. doi: 10.1111/gcbb.12171
Noordally, Z. B., and Millar, A. J. (2014). Clocks in algae. Biochemistry 54, 171–183. doi: 10.1021/bi501089x
Ohmori, M., and Hattori, A. (1974). Effect of ammonia on nitrogen fixation by the blue-green alga Anabaena cylindrica. Plant Cell Physiol. 15, 131–142.
Okoronkwo, N., VanHove, C., and Eskew, D. L. (1989). Evaluation of nitrogen-fixation by different strains of the Azolla-Anabaena symbiosis in the presence of a high-level of ammonium. Biol. Fertil. Soils 7, 275–278. doi: 10.1007/BF00709661
Oldroyd, G. E. D., and Dixon, R. (2014). Biotechnological solutions to the nitrogen problem. Curr. Opin. Biotechnol. 26, 19–24. doi: 10.1016/j.copbio.2013.08.006
Pankievicz, V. C. S., do Amaral, F. P., Santos, K. F. D. N., Agtuca, B., Xu, Y., Schueller, M. J., et al. (2015). Robust biological nitrogen fixation in a model grass-bacterial association. Plant J. Cell Mol. Biol. 81, 907–919. doi: 10.1111/tpj.12777
Peters, G. A., and Meeks, J. C. (1989). The Azolla-Anabaena symbiosis - basic biology. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 193–210. doi: 10.1146/annurev.pp.40.060189.001205
Peters, G. A., Toia, Jr. R. E, Evans, W. R., Crist, D. K., Mayne, B. C., and Poole, R. E. (1980). Characterization and comparison of five N2-fixing Azolla-Anabaena associations, I. Optimization of growth conditions for biomass increase and N content in a controlled environment. Plant Cell Environ. 3, 261–269.
Rai, A. N., Soderback, E., and Bergman, B. (2000). Cyanobacterium-plant symbioses. New Phytol. 147, 449–481. doi: 10.1046/j.1469-8137.2000.00720.x
Ramanan, R., Kim, B. H., Cho, D. H., Oh, H. M., and Kim, H. S. (2016). Algae–bacteria interactions: evolution, ecology and emerging applications. Biotechnol. Adv. 34, 14–29. doi: 10.1016/j.biotechadv.2015.12.003
Ran, L., Larsson, J., Vigil-Stenman, T., Nylander, J. A. A., Ininbergs, K., Zheng, W. W., et al. (2010). Genome erosion in a nitrogen-fixing vertically transmitted endosymbiotic multicellular cyanobacterium. PLoS ONE 5:e11486. doi: 10.1371/journal.pone.0011486
Robinson, M. D., McCarthy, D. J., and Smyth, G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Scheible, W.-R., Morcuende, R., Czechowski, T., Fritz, C., Osuna, D., Palacios-Rojas, N., et al. (2004). Genome-wide reprogramming of primary and secondary metabolism, protein synthesis, cellular growth processes, and the regulatory infrastructure of Arabidopsis in response to nitrogen. Plant Physiol. 136, 2483–2499. doi: 10.1104/pp.104.047019
Shi, D. J., and Hall, D. O. (1988). The Azolla-Anabaena association: historical perspective, symbiosis and energy metabolism. Bot. Rev. 54, 353–386. doi: 10.1007/BF02858416
Shilton, A. N., Powell, N., and Guieysse, B. (2012). Plant based phosphorus recovery from wastewater via algae and macrophytes. Curr. Opin. Biotechnol. 23, 884–889. doi: 10.1016/j.copbio.2012.07.002
Stamp, P., Messmer, R., and Walter, A. (2012). Competitive underutilized crops will depend on the state funding of breeding programmes: an opinion on the example of Europe. Plant Breed. 131, 461–464. doi: 10.1111/j.1439-0523.2012.01990.x
Summers, M. L., Wallis, J. G., Campbell, E. L., and Meeks, J. C. (1995). Genetic evidence of a major role for glucose-6-phosphate dehydrogenase in nitrogen fixation and dark growth of the cyanobacterium Nostoc sp. strain ATCC 29133. J. Bacteriol. 177, 6184–6194. doi: 10.1128/jb.177.21.6184-6194.1995
Thimm, O., Bläsing, O., Gibon, Y., Nagel, A., Meyer, S., Krüger, P., et al. (2004). MAPMAN: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 37, 914–939. doi: 10.1111/j.1365-313X.2004.02016.x
Truernit, E., Bauby, H., Dubreucq, B., Grandjean, O., Runions, J., Barthélémy, J., et al. (2008). High-resolution whole-mount imaging of three-dimensional tissue organization and gene expression enables the study of phloem development and structure in Arabidopsis. Plant Cell 20, 1494–1503. doi: 10.1105/tpc.107.056069
Udvardi, M., and Poole, P. S. (2013). Transport and metabolism in legume-rhizobia symbioses. Annu. Rev. Plant Biol. 64, 781–805. doi: 10.1146/annurev-arplant-050312-120235
Veys, P., Lejeune, A., and Van Hove, C. (2002). The pore of the leaf cavity of Azolla species: teat cell differentiation and cell wall projections. Protoplasma 219, 31–42. doi: 10.1007/s007090200003
Veys, P., Waterkeyn, L., Lejeune, A., and Van Hove, C. (1999). The pore of the leaf cavity of Azolla: morphology, cytochemistry and possible functions. Symbiosis 27, 33–57.
Vidal, E. A., Moyano, T. C., Krouk, G., Katari, M. S., Tanurdzic, M., McCombie, W. R., et al. (2013). Integrated RNA-seq and sRNA-seq analysis identifies novel nitrate-responsive genes in Arabidopsis thaliana roots. BMC Genomics 14:701. doi: 10.1186/1471-2164-14-701
Wagner, G. M. (1997). Azolla: a review of its biology and utilization. Bot. Rev. 63, 1–26. doi: 10.1007/BF02857915
Wang, R., Okamoto, M., Xing, X., and Crawford, N. M. (2003). Microarray analysis of the nitrate response in Arabidopsis roots and shoots reveals over 1,000 rapidly responding genes and new linkages to glucose, trehalose-6-phosphate, iron, and sulfate metabolism. Plant Physiol. 132, 556–567. doi: 10.1104/pp.103.021253
Watanabe, I., Roger, P. A., Ladha, J. K., and van Hove, C. (1992). Biofertilizer Germplasm Collections at IRRI. Los Baños: International Rice Research Institute.
Wilson, E. W., Rowntree, S. C., Suhre, J. J., Weidenbenner, N. H., Conley, S. P., Davis, V. M., et al. (2014). Genetic Gain × Management Interactions in Soybean: II. Nitrogen Utilization. Crop Sci. 54, 340. doi: 10.2135/cropsci2013.05.0339
Xu, G., Fan, X., and Miller, A. J. (2012). Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 63, 153–182. doi: 10.1146/annurev-arplant-042811-105532
Yashima, H., Fujikake, H., Sato, T., Ohtake, N., Sueyoshi, K., and Ohyama, T. (2003). Systemic and local effects of long-term application of nitrate on nodule growth and N2 fixation in soybean (Glycine max [L.] Merr.). Soil Sci. Plant Nutr. 49, 825–834. doi: 10.1080/00380768.2003.10410344
Zheng, W., Bergman, B., Chen, B., Zheng, S., Xiang, G., and Rasmussen, U. (2009). Cellular responses in the cyanobacterial symbiont during its vertical transfer between plant generations in the Azolla microphylla symbiosis. New Phytol. 181, 53–61. doi: 10.1111/j.1469-8137.2008.02644.x
Keywords: dinitrogen fixation, heterocystous cyanobacteria, aquatic ferns, vasculature, diel cycling, clock evolution, Azolla/Nostoc azollae symbiosis, RNA-seq
Citation: Brouwer P, Bräutigam A, Buijs VA, Tazelaar AOE, van der Werf A, Schlüter U, Reichart G-J, Bolger A, Usadel B, Weber APM and Schluepmann H (2017) Metabolic Adaptation, a Specialized Leaf Organ Structure and Vascular Responses to Diurnal N2 Fixation by Nostoc azollae Sustain the Astonishing Productivity of Azolla Ferns without Nitrogen Fertilizer. Front. Plant Sci. 8:442. doi: 10.3389/fpls.2017.00442
Received: 02 December 2016; Accepted: 14 March 2017;
Published: 31 March 2017.
Edited by:
Robert Edward Sharwood, Australian National University, AustraliaReviewed by:
Axel Tiessen, Center for Advanced Research, The National Polytechnic Institute, MexicoSean Cutler, University of California, Riverside, USA
Copyright © 2017 Brouwer, Bräutigam, Buijs, Tazelaar, van der Werf, Schlüter, Reichart, Bolger, Usadel, Weber and Schluepmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Henriette Schluepmann, aC5zY2hsdXBtYW5uQHV1Lm5s
†Present address: Andrea Bräutigam, Leibniz Institute of Plant Genetics and Crop Plant Research, Gatersleben, Germany
‡These authors have contributed equally to this work