 Pasqualina Colasuonno1†
Pasqualina Colasuonno1† Ilaria Marcotuli1†
Ilaria Marcotuli1† Antonio Blanco1
Antonio Blanco1 Marco Maccaferri2
Marco Maccaferri2 Giuseppe Emanuele Condorelli2
Giuseppe Emanuele Condorelli2 Roberto Tuberosa2
Roberto Tuberosa2 Roberto Parada3
Roberto Parada3 Adriano Costa de Camargo3
Adriano Costa de Camargo3 Andrés R. Schwember3*
Andrés R. Schwember3* Agata Gadaleta1
Agata Gadaleta1- 1Department of Agricultural and Environmental Science (DISAAT), University of Bari “Aldo Moro”, Bari, Italy
- 2Department of Agricultural and Food Sciences (DISTAL), University of Bologna, Bologna, Italy
- 3Facultad de Agronomía e Ingeniería Forestal, Pontificia Universidad Católica de Chile, Santiago, Chile
Carotenoid pigment content is an important quality trait as it confers a natural bright yellow color to pasta preferred by consumers (whiteness vs. yellowness) and nutrients, such as provitamin A and antioxidants, essential for human diet. The main goal of the present review is to summarize the knowledge about the genetic regulation of the accumulation of pigment content in durum wheat grain and describe the genetic improvements obtained by using breeding approaches in the last two decades. Although carotenoid pigment content is a quantitative character regulated by various genes with additive effects, its high heritability has facilitated the durum breeding progress for this quality trait. Mapping research for yellow index and yellow pigment content has identified quantitative trait loci (QTL) on all wheat chromosomes. The major QTL, accounting for up to 60%, were mapped on 7L homoeologous chromosome arms, and they are explained by allelic variations of the phytoene synthase (PSY) genes. Minor QTL were detected on all chromosomes and associated to significant molecular markers, indicating the complexity of the trait. Despite there being currently a better knowledge of the mechanisms controlling carotenoid content and composition, there are gaps that require further investigation and bridging to better understand the genetic architecture of this important trait. The development and the utilization of molecular markers in marker-assisted selection (MAS) programs for improving grain quality have been reviewed and discussed.
Introduction
Durum wheat (Triticum turgidum L. ssp. durum) is a cereal crop grown around the world on about 17 million hectares and with about 37 million tons produced annually during the last decade, with wide variation from 32 to 42 million tons caused mainly by drought and heat stresses (data FAO, 2017). Globally durum wheat represents only 8% of the whole area cultivated with wheat and about 5% of world wheat production. The principal durum-producing countries are the European Union, Canada, Turkey, USA, Algeria, Kazakhstan, and Mexico, whereas minor production countries encompass Syria, Morocco, Tunisia, India, Australia, and Argentina and Chile, among others. Major producers in the EU are Italy, France, Greece, and Spain. Although durum wheat is a relatively minor crop worldwide, it is the main crop for many areas of the Mediterranean basin and makes up the raw material for various finished products such as pasta and couscous consumed all over the world (Kabbaj et al., 2017).
Grain protein content and conformation together with yellow color are the most valued wheat quality traits, which are important in the commercial, nutritional, and technological values of grain and end products of both common and durum wheat (Sissons, 2008; Nazco et al., 2012; Subira et al., 2014; Mazzeo et al., 2017).
Semolina and pasta color are the consequence of two distinct constituents: yellow (desirable) and brown (undesirable) pigments. The yellow color, principally explained by carotenoid accumulation in kernels, has been considered a source of significant nutrients/antioxidant compounds and a factor for the commercial value since consumers prefer a bright yellow color of semolina and the pasta products.
It is a typical quantitative trait controlled by a complex genetic system (quantitative trait loci, QTL) and influenced by environmental factors. As confirmed by the high value of heritability, the genetic component is predominant, and this has facilitated the success of breeding programs (Elouafi et al., 2001). A consequence of this intense breeding activity has been proved by the higher carotenoid concentration in durum wheat cultivated varieties compared to the wild ones (Digesù et al., 2009).
Genetic analyses based on molecular markers have mapped major QTL for carotenoid content on homoeologous group 7. Minor QTL, associated to significant markers, were detected on almost all chromosomes of the durum wheat genome. Significant marker-trait associations for carotenoid content have been detected on all of the chromosomes by linkage mapping. Association mapping has been used as a new strategy for the dissection of this trait and highlighted its complexity.
Due to the importance of this quality trait, the aim of the present review is to summarize the information available on the detection of QTL for carotenoid content and individual components and the identification of candidate genes in durum wheat.
Carotenoids and Nutritional Aspects
In the first years of the 21st century, major breeding programs were focused on improving the durum productivity traits of wheat, such as grain yield and biotic and abiotic stress resistance. Recently, the attention on food quality over quantity has switched the research aims at increasing wheat nutritional value estimated through different parameters, like protein content, water absorption, and flour color. The latter one is due to the carotenoid pigments, whose nutritional benefits in human health is worldwide recognized (Sommer and Davidson, 2002).
Over 600 carotenoids have been identified in plants and microorganisms. They are one of the most studied groups of natural pigments, because of their broad distribution, structural variety, and multiple functions. All fruit and plant color, ranging from yellow to red, are good sources of carotenoids (Britton, 1998).
Bendich and Olson were the first scientists that characterize more than 750 carotenoid compounds (Bendich and Olson, 1989; Olson and Krinsky, 1995; DellaPenna and Pogson, 2006). Carotenoids is the generic term indicating the majority of red, orange, and yellow pigments naturally encountered in photosynthetic organisms and in certain fungi and bacteria (Britton 1995; Khoo et al., 2011).
Most carotenoids are tetraterpenoids (C40 compounds), which are composed of eight isoprenic units linked in a linear and symmetrical structure. The basic cyclic structure can be changed by dehydrogenation, hydrogenation, cyclization, and oxidation reactions, while the high chemical reactivity has been induced by a complex system of double bonds (Oliver and Palou, 2000). Two classes of carotenoids are found in nature: (a) the carotenes, linear tetraterpenoid hydrocarbons (i.e., β-carotene) that can be cyclized at one or both ends of the molecule, and (b) the xanthophylls, composed by one or more oxygen groups (i.e., lutein, violaxanthin, neoxanthin, and zeaxanthin) (Van den Berg et al., 2000). Overall, carotenoids possess general properties common to all carotenoids (i.e., antioxidant nature) from specific ones (i.e., provitamin A, only particular ones) (Young and Lowe, 2018).
Food, animal feed, and pharmaceutical industries are using carotenoids for their color properties (in fruit juices, pasta, candies, cheese, chicken skin, and egg yolk) or for their contribution in the flavor and fragrances of some foods (Landrum, 2009). In addition, in a context of increasing interest in improving health through the consumption of natural products, they are considered also in food fortification. It has been demonstrated that they are precursors of vitamin A, generating health advantages, such as antioxidant properties, reinforcing the immune system, decreasing the risk of degenerative and cardiovascular ailments, anti-obesity/hypolipidemic properties, and defense of the macula region of the retina (Mezzomo et al., 2015).
Among the carotenoid compounds, following the presence/absence of the provitamin A in the molecule structure, fewer than 10% show a significant biological activity and act as vitamin A precursors. The precursors of vitamin A have a minimum of one β-ionone ring and a polienic chain with 11 carbons at least (Kelly et al., 2018).
Bioactive compounds and vitamin A are categorized as antioxidants, playing a crucial function in humans’ health such as in growth, in the development and maintenance of epithelial tissues, in the immune system strength, and in the first protection mechanism against oxidative stress. Antioxidants reduce the singlet oxygen in the human body and scavenge free radicals, i.e., reactive nitrogen species (RNS) and reactive oxygen species (ROS) (Choe and Min, 2005; Leong et al., 2018). The oxidation of carotenoids by ROS causes the loss of their characteristic color inducing cell protection and the prevention of degenerative diseases (Halliwell, 2011; Guaadaoui et al., 2014).
Carotenoids are transformed to vitamin A to satisfy the body requirements, changing the levels of conversion efficiency (Mezzomo and Ferreira, 2016). A prolonged deficiency of vitamin A can produce skin modifications, corneal ulcers, and night blindness, while a surplus is toxic and may be associated to congenital malformation in pregnancy, bone diseases in patients with chronic renal malfunction, blindness, xerophthalmia, and death (Mezzomo et al., 2015).
According to the high unsaturation degree of carotenoids, light, heat, acids, and enzymatic oxidation can change their structure from the trans-isomers (the most stable type in nature) to the cis-structure, producing a minor decrease of color and provitamin activity (Schroeder and Johnson, 1995).
The Carotenoids’ Value in Yellow Color of Wheat Products
The yellow-amber color of semolina is caused by the carotenoid (yellow) pigment content (YPC) in the entire grain, which is known as the yellow index (YI) of semolina at a commercial level (CIE, 1986). The average carotenoid concentration in durum wheat is 6.2 ± 0.13 mg/kg in dry weight, determining the pasta color (Beleggia et al., 2011; Brandolini et al., 2015).
In wheat kernel, a wide range of carotenoids have been detected such as lutein, β-carotene, zeaxanthin, β-cryptoxanthin, β-apocarotenal, antheraxanthin, taraxanthin (lutein-5,6-epoxide), flavoxanthin, and triticoxanthin (Lachman et al., 2017).
The pigments are variable: α- and β-carotene (7.7%) are mainly located in the germ, while lutein, the most abundant pigment (86–94%) (Konopka et al., 2004; Digesù et al., 2009), is equally distributed across the layers (Borrelli et al., 2008). Specifically, aleurone layer, starchy endosperm, and germ contain 0.425, 0.557, and 2.157 mg/kg of lutein, respectively. In parallel, aleurone and germ contain 0.776 and 3.094 mg/kg zeaxanthin.
During the milling process, a large amount of these components are gradually reduced, depending on the extraction rate (Paznocht et al., 2019). Lutein, and a small amount of zeaxanthin, has higher cooking stability compared to other carotenoids commonly present in foods, for example, β-cryptoxanthin and β-carotene (Britton and Khachik, 2009; Kean et al., 2011).
In the wheat end products, steady-state level of carotenoids is dependent on the equilibrium between biosynthesis and degradation in the processing phase. This last process has been principally attributed to oxidative enzymes, such as the lipoxygenases (LOX), polyphenol oxidase (PPO), and peroxidases (PER), that can cause the loss of yellow color of flour and pasta (Hessler et al., 2002; Leenhardt et al., 2006; Garbus et al., 2013; Morris, 2018).
Carotenoid Biosynthesis and Degradation Pathways
The carotenoid metabolic biosynthetic pathway has been thoroughly investigated in some plants, including Arabidopsis, rice, maize, pepper, tomato, and orange (see studies by Colasuonno et al., 2017a; Rodriguez-Concepcion et al., 2018; Sun et al., 2018). This biosynthetic route has been examined in depth (Figure 1), and all genes and enzymes involved have been isolated and well characterized. It starts with the condensation of two molecules of geranylgeranyl diphosphate (GGPP) by phytoene synthase (PSY) to generate phytoene, which normally is not accumulated in tissues. This step is a key rate-limiting stage of carotenoid biosynthesis, since it might affect the carotenoid pool content (Cazzonelli and Pogson, 2010; Ke et al., 2019). Following a sequence of desaturation and isomerization reactions catalyzed by phytoene desaturase (PDS), zeta-carotene isomerase (Z-ISO), zeta-carotene desaturase (ZDS), and carotenoid isomerase (CRTISO), the lycopene biosynthesis, a red-colored carotenoid, takes place. Double lycopene cyclization produces orange β-carotene (branch β-β) or α-carotene (branch β-ε). Further hydroxylation of α-carotene generates yellow zeinoxanthin and lutein, while the modification of β-carotene produces β-cryptoxanthin, zeaxanthin, antheraxanthin, violaxanthin, and neoxanthin (Figure 1). These steps are catalyzed by two non-heme β-carotene hydroxylases (BCH1 and BCH2) and two heme hydroxylases (CYP97A and CYP97C), respectively (Sun et al., 2018). The consequent epoxidation and de-epoxidation of zeaxanthin by zeaxanthin epoxidase (ZEP) and violaxanthin de-epoxidase (VDE) induce the production of xanthophylls, molecules involved in plant protection’s mechanisms (Jahns and Holzwarth, 2012). The last phase of carotenoid biosynthesis consists in the transformation of violaxanthin into neoxanthin by neoxanthinsynthase (NXS) (Sun et al., 2018). Further oxidative cleavage reactions of violaxanthin and neoxanthin produce xanthoxin, transformed to the abscisic acid (ABA) plant hormone by ABA-aldehyde enzymes (Seo and Koshiba, 2002).
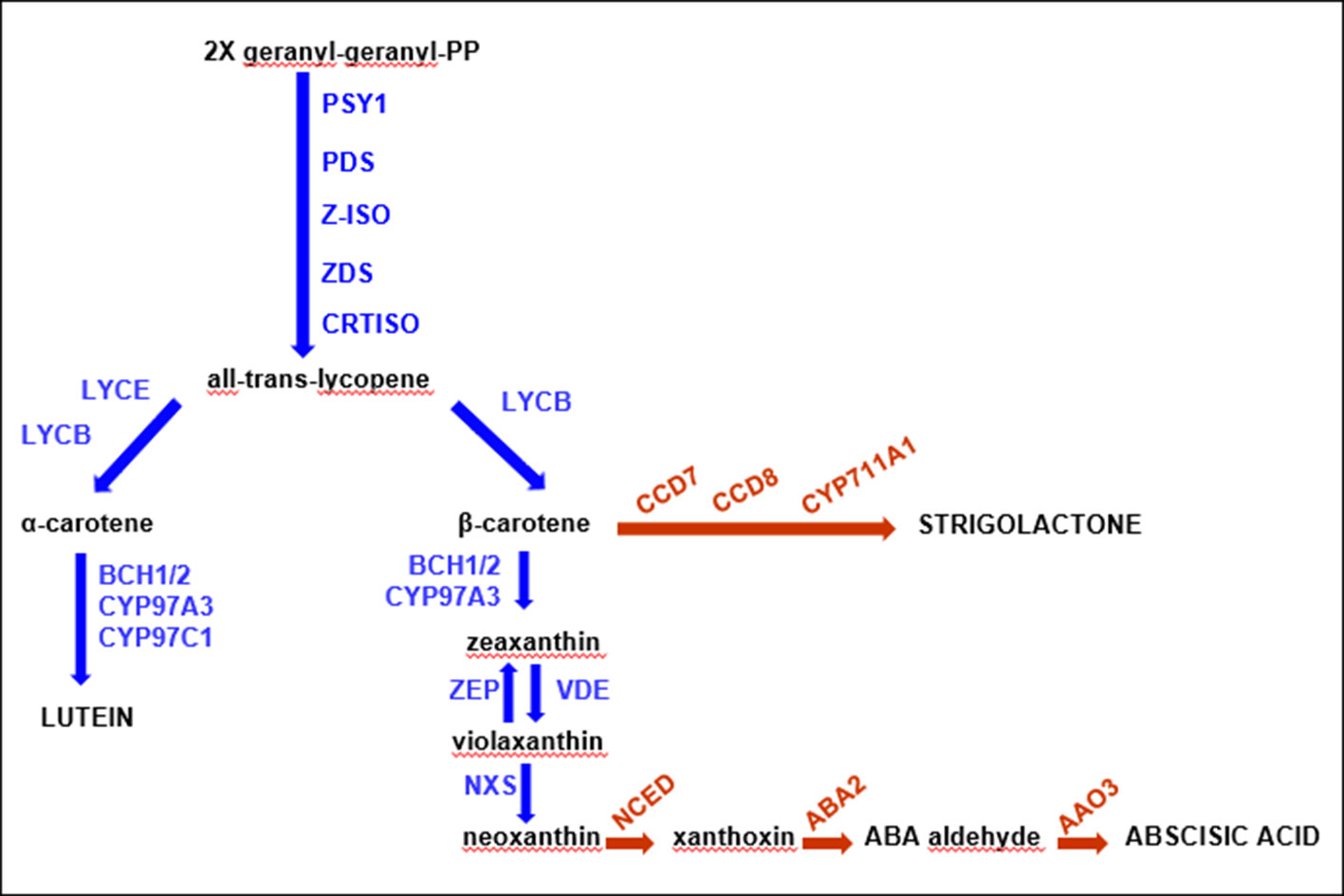
Figure 1 Schematic carotenoid pathway. The main components of the biosynthetic chain have been reported in the figure in black font, while all the enzymes involved are blue. The proteins from the related pathways are indicated in red.
Strigolactones are carotenoid derivatives and originated from the functioning of carotenoid cleavage dioxygenases (CCDs), contributing to the regular accumulation and balanced levels of pigments (Al-Babili and Bouwmeester, 2015; Nisar et al., 2015).
Numerous studies focused on all the carotenoid enzymes have shown that the inheritance of this trait in wheat is of quantitative nature, and it is highly heritable (Digesù et al., 2009; Blanco et al., 2011; Roncallo et al., 2012; Schulthess and Schwember, 2013; Schulthess et al., 2013). Consequently, the correct approach to study carotenoid pigments is to conduct a QTL mapping strategy.
Determination of Yellow Pigments Content and Their Single Components
One of the principal problems of carotenoid analyses is testing for their composition and concentration through reproducible and accurate methods, often using only small amounts of seeds. The detection of carotenoid levels is technically restricted by several limitations such as the interference with some regulation/degradation processes and the product storage (Sandmann, 2001). Carotenoid content is a complex trait, and there are several procedures for measuring total pigment concentration and individual pigment compounds.
The reference methods for the total YPC are Standard Method 152 (ICC, 1990) of the International Association for Cereal Science and Technology (ICC) and AACC International Official Method 14-50.01 (AACC International, 2013). These two procedures rely on the extraction of total pigmwwents in water-saturated n-butanol followed by a spectrophotometric quantification of the absorbance of the alcoholic extract at 435.8 nm (the wavelength of maximum absorption of lutein, the dominant carotenoid in durum wheat), using USA/Canada standards (Fu et al., 2017).
Alternatively, pigment content can be measured by the YI determination based on the quantification of flour light reflectance. Analysis with Minolta CR-300 Chroma Meter (Konica Minolta Pty Ltd, Macquarie Park, NSW) geared up with a pulsed xenon arc lamp is the most used instrument for YI analysis. It provides the brown and yellow indexes, after calibration with standard flour control samples. Specially, the measurements consist of the L* (lightness, ranging from 0 for darkness to 100 for total light), a* (red–green chromaticity), and b* (yellow–blue chromaticity) coordinates of the Munsell color system, employing D65 lightning (CIE, 1986). The b* value is directly linked to the lutein and carotene contents, determining the variation in the yellow intensity (Rodriguez-Amaya and Kimura, 2004). Near-infrared (NIR) spectroscopy is a fast, non-destructive, and economic technique, useful for prediction of individual carotenoid pigments in maize and wheat semolina (McCaig et al., 1992).
Accurate measurements of carotenoids and their components can be exclusively obtained by high-performance liquid chromatography (HPLC) analysis (Brandolini et al., 2008; Fu et al., 2017). HPLC, with grade solvents of methyl tert-butyl ether and methanol, and photodiode array detector, allows the identification and separation of carotenoid compounds (Panfili et al., 2004). Detailed HPLC protocols to identify carotenoids in cereals and end products have been designed by several research groups (Burkhardt and Böhm, 2007; Digesù et al., 2009; Irakli et al., 2011; Shen et al., 2015).
Qtl Detection in Durum Wheat
YPC is controlled by various genes with additive effects that are affected by different environmental conditions (Schulthess et al., 2013). In classic quantitative genetics research, the creation of linkage maps in biparental populations allowed studying the number of loci regulating the trait, the phenotypic and pleiotropic effects, as well as the epistatic interactions with other QTL, enabling the identification and characterization of candidate genes. Mapping studies for YPC and YI in various biparental populations have led to QTL detection across all wheat chromosomes (Table 1).
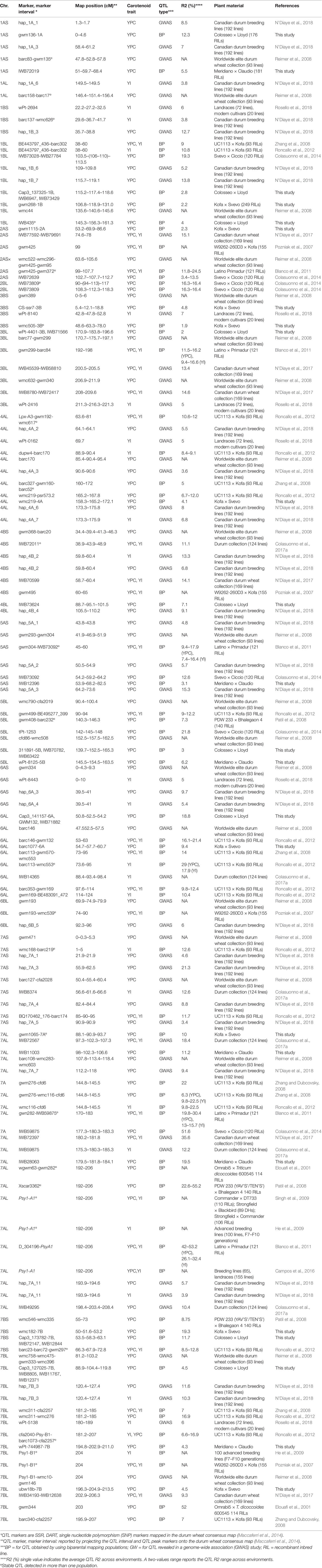
Table 1 Summary of quantitative trait loci (QTL) clusters for grain yellow pigment content (YPC) and yellow index (YI) reported in tetraploid wheat from the literature and from this study.
A suitable population for carotenoid analysis are the recombinant inbred lines (RILs) in advanced selfing generations, doubled haploid (DH) populations, or populations derived from backcrosses (Elouafi et al., 2001; Pozniak et al., 2007; Singh et al., 2009; Tsilo et al., 2011; Colasuonno et al., 2014).
When wheat germplasm, including cultivars, advanced breeding lines, or germplasm collections, are considered, mapping methods of genome-wide association study (GWAS) have been applied to link some marker haplotypes with trait expression (Vargas et al., 2016; Colasuonno et al., 2017a; Fiedler et al., 2017). The principle behind the method is to estimate correlations among the genotypes and the phenotypes in panels of lines, based on the linkage disequilibrium between the allelic variants of molecular markers and causal genes (Gupta et al., 2005; Bush and Moore, 2012). This approach has been the official method for many years. In the last decade, it has been extensively used for different traits owing to the availability of high numbers of DNA-based markers uniformly distributed in the genome (such as the high-density maps obtained from single nucleotide polymorphisms, SNPs, Wang et al., 2014 and Maccaferri et al., 2014) and the improvement of statistical tools. These included improved mixed models that effectively take into account the interfering panel population structure effects (Breseghello and Sorrells, 2006; Maccaferri et al., 2006; Zhang et al., 2010; Maccaferri et al., 2011; Lipka et al., 2012). Although several reports of SNP markers, associated to QTL for carotenoid pigments, have been published to date (Table 1), they mostly relied on linkage mapping and biparental RIL populations. Only four studies were deeply focused on SSR and SNP-based association mapping in durum wheat (Reimer et al., 2008; Colasuonno et al., 2017a; N’Diaye et al., 2017; Rosello et al., 2018), and only one considered haplotypes instead of single bi-allelic markers (N’Diaye et al., 2018).
Carotenoid QTL Regions
The results of recent peer-reviewed studies and additional studies provided by the authors reporting QTL for carotenoid pigments in durum wheat are compiled in Table 1 (QTL clusters) and Table S1 (all QTL), encompassing information on mapping population, examined phenotypic traits, and markers associated to carotenoid QTL. Map position of the major QTL clusters listed in Tables 1 and S1 are reported in Figure 2. The schematic representation is based upon the durum wheat consensus map published by Maccaferri et al. (2014), and revised for the carotenoid composition by Colasuonno et al. (2017a). Considering that each QTL/MTA identified by different studies was located in the same position of the durum wheat consensus map, we used the cM reported by Maccaferri et al. (2014) to identify the genome location.
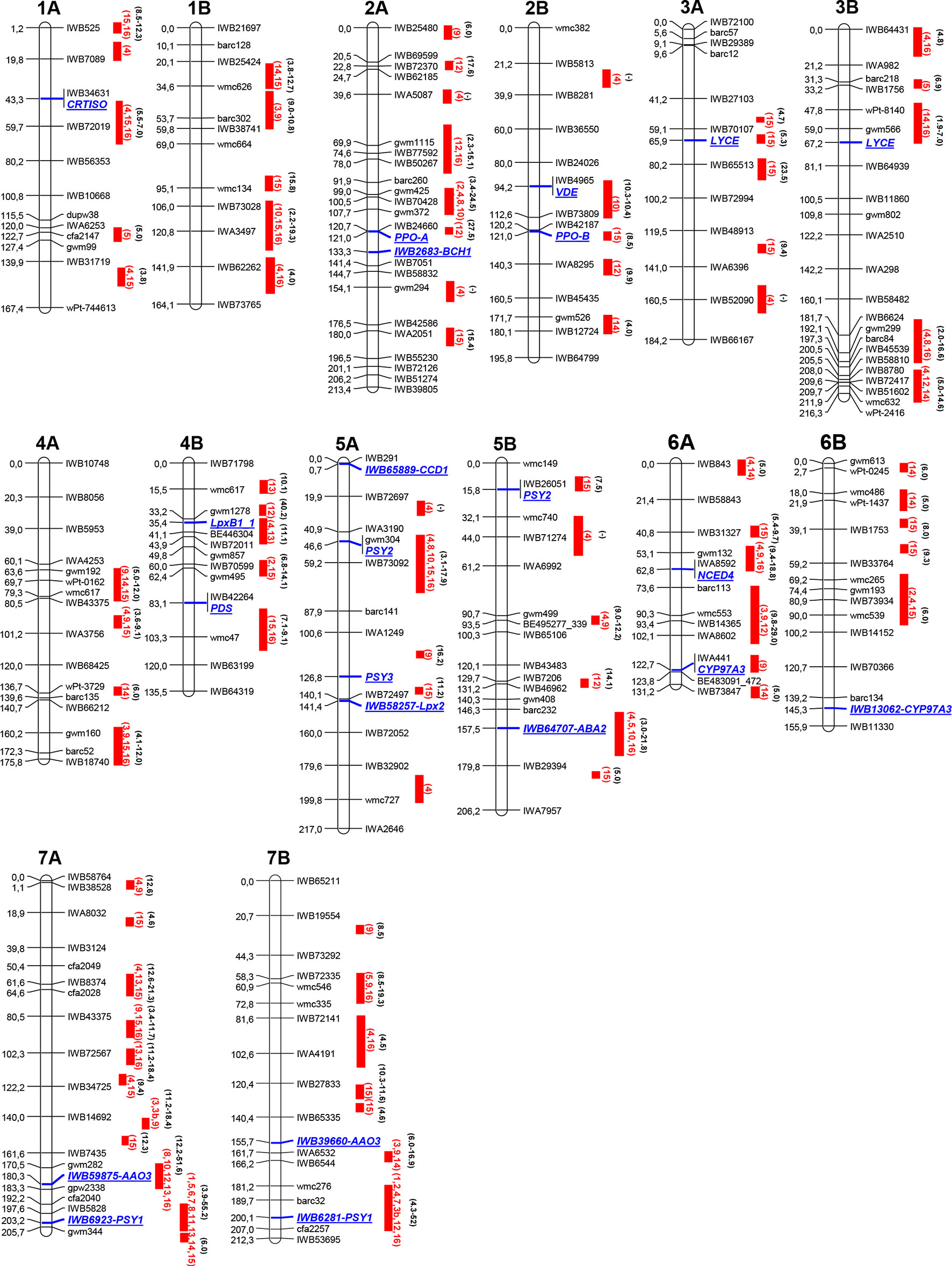
Figure 2 Schematic representation of durum wheat chromosomes [based on theMaccaferri et al. (2014) map] and quantitative trait loci (QTL) summary for yellow pigment content (YPC) or yellow index (YI) trait detected in references in Table 1. Markers, on the right chromosome side, are reported every 20 cM approximately. cM distances are indicated on the left side of the bar. Red solid bars indicate the QTL confidence interval regions. The red bars report the reference publication numbers in red (as in Table 1 and Table S1) and the QTL R2 range value in black. 1, Elouafi et al. (2001); 2, Pozniak et al. (2007); 3, Zhang et al. (2008); 3b, Zhang and Dubcovsky (2008); 4, Reimer et al. (2008); 5, Patil et al. (2008); 6, Singh et al. (2009); 7, He et al. (2009); 8, Blanco et al. (2011); 9, Roncallo et al. (2012); 10, Colasuonno et al. (2014); 11, Campos et al. (2016); 12, N’Diaye et al. (2017); 13, Colasuonno et al. (2017a); 14, Rosello et al. (2018); 15, N’Diaye et al. (2018); 16, this study.
Every chromosome is depicted with the first and the last SNP marker, and one marker every 20 cM. SSR markers have also been incorporated to anchor the actual consensus map with the previous ones. For most of the detected QTL, the connection between the different maps has been quite simple to be achieved.
The QTL mapping study highlights the differences in number and map position of the QTL identified in several analyses. This could be due to the specific plant material and/or the analytical and statistical procedures adopted in each study (e.g., linkage mapping vs. GWAS). In fact, the presence of numerous genes with an additive effect on the trait, the parental influence on the genotypes of the mapping populations, the genotype × environment interaction, and the number of markers used may affect the results. Moreover, a different carotenoid measurement approach and statistical procedures adopted might influence the reproducibility of each QTL analysis.
Mapping studies for YPC and YI identified 81 QTL (including singletons and QTL clusters) distributed on all wheat chromosomes (Table S1). Some of these QTL have been detected in more than one map/population, indicating the presence of stable alleles valuable for enhancing color and nutritional value of wheat grain (QTL clusters, listed in Table 1). Twenty stable QTL (highlighted in Table 1) were detected on chromosomes 1A (two), 1B (two), 2A, 2B, 3B, 4A (two), 4B, 5A, 5B (two), 6A, 6B, 7A (four), and 7B (two). Therefore, these QTL can be considered useful for MAS in breeding programs. Major, moderate, and minor QTL were defined basing on the percentage of the phenotypic variation (> 40%, 10–40%, and <10%, respectively) as reported in the different studies. The major QTL have been mapped on chromosomes 7AL (Elouafi et al., 2001; Patil et al., 2008; Zhang and Dubcovsky, 2008) and 7BL distal regions (Elouafi et al., 2001; Pozniak et al., 2007; Zhang and Dubcovsky, 2008). In particular, two different QTL have been identified on 7AL (Zhang and Dubcovsky, 2008; Blanco et al., 2011; Crawford and Francki, 2013): the first one (QTL-72) associated with allelic variations of the AO gene (Colasuonno et al., 2014; Colasuonno et al., 2017b) with a negative effect (R2 22.1% for YPC and 18.4% for YI) on carotenoid content, and the second one (QTL-73) in the region of the PSY1 gene (He et al., 2008; Zhang and Dubcovsky, 2008; He et al., 2009) with a positive and consistent effect up to 60% on YPC. The same QTL–gene associations are reported on chromosome 7BL with R2 up to 29.1% for the first QTL (QTL-80, at 180–189 cM) and 52% for the one at 181–211 cM (QTL-81). All the loci detected on chromosome group 7 resulted as negatively correlated with grain yield per spike (GYS) and thousand grain weights (TKWs) but positively correlated with protein content. The same association highlighted on chromosomes 7A and 7B between QTL and PSY2 gene has been reported on chromosome 5AS with moderate effect (R2 9.4–17.9% for YPC and 7.4–16.4 for YI) and negative correlation with GYS and TKW.
Various studies have confirmed the role of the violaxanthin de-epoxidase (VDE) on 2B (Tsilo et al., 2011; Roncallo et al., 2012) in YPC and YI content. The association between VDE and the QTL on 2BL at 94–113 cM has been validated by many results gained thanks to biparental mapping populations. Colasuonno et al. (2014) identified a QTL on 2BL at 94–113 cM with moderate and negative effect from the “Ciccio” alleles (16.4%).
Minor QTL have been detected on chromosomes 1A, 1B, 2A, 3A, 3B, 4A, 4B, 5A, 5B, and 6B (Pozniak et al., 2007; Zhang et al., 2008). Among them, 2A, 4B, and 6A showed variable effect with R2 of 3.5–15% for YPC and 12.3–21% for YI, 4–11.9% for YPC and 26.2 for YI, and 14–17% for YPC and 28.3% for YI, respectively (Table 1 and Table S1). Lipoxygenase (LpxB1.1) gene is located on chromosome 4B, in the same site of the QTL described in literature by several authors (QTL-40; Carrera et al., 2007; Reimer et al., 2008; Verlotta et al., 2010; Colasuonno et al., 2017a).
QTL for individual carotenoid constituents in wheat were outlined by Howitt et al. (2009) and Blanco et al. (2011). Considering wheat rice synteny, two genes, phytoene synthase (Psy-A1) and ε-cyclase (ε-LCY), were identified by Howitt et al. (2009) as candidate genes for two of the QTL involved lutein content in wheat endosperm. A segregant population, achieved from crossing the durum wheat cultivars Primadur and Latino, was used by Blanco et al. (2011) to detect QTL for individual carotenoid compounds (lutein, zeaxanthin, β-cryptoxanthin, α-carotene, and β-carotene), YI, and yellow pigment concentration. Total carotenoid concentration accounted for 30–50% of the yellow pigment quantities in durum wheat (Hentschel et al., 2002), reflecting unknown color-producing substances in the durum extracts. Lutein was the most prominent carotenoid compound identified, followed by zeaxanthin, α-carotene, and β-carotene, whereas β-cryptoxanthin was a secondary component. QTL mapping identified clusters of QTL for total and/or one or more carotenoid compounds (α-carotene and β-carotene) in the same chromosomal zones (2A, 3B, 5A, and 7A) where QTL for yellow pigment concentration and YI were detected. The existence of molecular markers related to the main QTL previously indicated is a valuable tool for marker-assisted selection (MAS) programs to increase high carotenoid concentration and the nutritional value of wheat grains.
Candidate Genes for Endosperm Yellowness
Candidate Genes for Carotenoid Biosynthesis
The “candidate gene approach” has been used in QTL or association mapping to test SNPs within a candidate gene for a significant association with the yellow color character. Although most studies were focused in increasing the carotenoid content or altering the relative components through conventional breeding (Schulthess and Schwember, 2013), some carotenoid genes have been characterized and/or linked to QTL for carotenoids. In wheat, significant attention has been given to the carotenoid biosynthesis genes (PSY (Pozniak et al., 2007; He et al., 2008; Dibari et al., 2012; Campos et al., 2016; Vargas et al., 2016), lycopene ε-cyclase (LCYE) (Howitt et al., 2009; Crawford and Francki, 2013), lycopene β-cyclase (LYCB) (Zeng et al., 2015), carotenoid β-hydroxylase (HYD) (Qin et al., 2012), carotene desaturase (PDS), and ZDS (Cong et al., 2010), while the degradation of carotenoids has been studied by some catabolic genes, such as aldehyde oxidase (AO) (Colasuonno et al., 2017b), polyphenol oxidase (PPO) (Si et al., 2012), lipoxygenase (LOX or LPX) (Verlotta et al., 2010), and peroxidase (PER) (Asins and Perez de la Vega, 1985; Fraignier et al., 2000; Ficco et al., 2014).
As previously noted, the gene with major effect on YPC and YI traits is the PSY, which is essential for the carotenoid accumulation in the kernels (Gallagher et al., 2004). Phylogenetic approaches have identified three different PSY isoforms: PSY1, PSY2, and PSY3 mapped on homoeologous chromosome groups 7, 5, and 3, respectively (Dibari et al., 2012). Atienza et al. (2007) was the first study to show that PSY1 was located on chromosomes 7A and 7B of durum wheat. The gene coding for PSY1 on chromosome 7B was found to be co-segregant with a carotenoid QTL, while the PSY1 on chromosome 7A behaved as a co-dominant marker explaining 20–28% of the phenotypic variability (Pozniak et al., 2007). The effect of the PSY1 loci on the endosperm yellowness of wheat is variable depending on the genetic background. Therefore, the percentage of explained variability of endosperm yellowness for PSY1-A1 has been found ranging from medium (10–30%), to high (30–50%) and very high (> 50%) (Blanco et al., 2011; Colasuonno et al., 2014), while the reported values for PSY1-B1 are from low (< 10%) to medium (10–30%) (Zhang and Dubcovsky, 2008; Roncallo et al., 2012) in durum wheat. Overall, the effect of alternative alleles of PSY-A1 appears to be the most important in the variation of semolina yellowness (SY) using different durum wheat populations (Campos et al., 2016; Vargas et al., 2016). In these studies, PSY1-Al was associated to higher SY due to the allelic variant l of this gene. The use of molecular markers linked to PSY1-A1 (i.e., Psy1-A1_STS and YP7A-2 studied jointly) in MAS was suitable to enhance grain yellow pigmentation (Campos et al., 2016). In addition, at 35 days after anthesis, PSY1-A1 was 21-fold higher expressed in the high-yellowness compared to the low-yellowness genotypes evaluated, corroborating the major role of PSY1-A1 in the genotypes associated to high SY (Vargas et al., 2016).
Additional evidence highlights how other genes are involved in the control of grain amber color. The full-length DNA sequence of a ZDS gene on wheat chromosome 2A, designated Zds-A1, was cloned, and a co-dominant functional marker, YP2A-1, was designed based on the polymorphisms of two alleles (Dong et al., 2012). The functional marker, explaining 11.3% of the phenotypic variance for YP content, was co-segregating with a QTL for YP content detected on chromosome 2A in a DH population.
The lycopene ε-cyclase gene (LCYE) associated to a QTL on chromosome 3A, playing an important role in the regulation of lutein content in wheat grain (Howitt et al., 2009). An SNP marker in LCYE was detected between two Australian wheat genotypes, and a highly significant (P < 0.01) association with a QTL on chromosome 3A in two mapping populations showed that LCYE is involved in color differences at a functional level (Crawford and Francki, 2013).
Zeng et al. (2015) were able to clone the lycopene β-cyclase gene (LCYB) and describe its function and connection with β-carotene biosynthesis in wheat grains. Their results suggest that LCYB has a key function in β-carotene biosynthesis in wheat and that LCYB may be useful to breed new wheat cultivars with high provitamin A content using RNA interference (RNAi) to hinder specific carotenogenic genes in the wheat endosperm.
Qin et al. (2012) characterized two genes, HYD1 and HYD2, encoding β-hydroxylases in wheat. They observed different expression patterns for different HYD genes and homeologs in vegetative tissues and developing grains of tetraploid and hexaploid wheats, indicating their distinct regulatory control in tissue, grain development, and ploidy-specific fashions. The expression of HYD-B1 achieved highest levels at the last stage of grain filling, showing how carotenoids were still synthesized in mature grains, raising the nutritional value of kernels.
In a recent investigation by Colasuonno et al. (2017), 24 candidate genes encompassed in the biosynthesis and catabolism of carotenoid compounds have been reported using wheat comparative genomics. SNPs identified in the coding sequences of 19 candidate genes enabled their chromosomal location and precise map positions on the two bread and durum reference consensus maps studied (Maccaferri et al., 2014; Wang et al., 2014). Six candidate genes (PSY1, PSY2, CYP97A3, VDE, ABA2, and AAO3) showing between one to five SNPs were significantly associated to YI by genome-wide association mapping in a collection of 233 accessions of tetraploid wheat, suggesting their involvement in the yellow pigment biosynthesis or catabolism. PSY1, BCH1, CYP97A3, VDE, and ABA2 were also associated to YPC. The phenotypic variation (R2) explained by each of these markers ranged between 5.9% and 16.3% for YI and from 7.4% to 14.8% for YPC.
Candidate Genes for Carotenoid Degradation
Lipoxygenase (LPX) genes, involved in the catabolic pathway, are the most characterized. LPX enzymes are non-heme iron-containing and dioxygenases are found in all kingdoms (Zhang et al., 2015), catalyzing the addition of oxygen in polyunsaturated fatty acids that possesses a cis, cis-1,4 pentadiene system (Garbus et al., 2009). In plants, lipoxygenases are found in leaves, seedlings, and seeds. LPX activity produces ROS that can induce oxidation and degradation of carotenoids (Borrelli et al., 2003). In durum wheat, there are different lipoxygenase genes and alleles contributing to the variation of pasta yellowness (Verlotta et al., 2010; Borrelli and Trono, 2016).
Holtman et al. (1996) reported that lipoxygenase-1 is responsible for LPX activity in barley seeds. Hessler et al. (2002) sequenced several wheat fragments, which were assigned to the Lpx-1 locus based on their similarity to barley genes. De Simone et al. (2010) reported different levels of Lpx-1 and Lpx-3 transcripts at maturity between cultivars with contrasting LPX activities, whereas Lpx-2 transcripts were absent at this stage. Lpx-B1 locus was located on the short arm of chromosome 4B (Hessler et al., 2002; Garbus et al., 2009; Verlotta et al., 2010), and five related genes and allele sequences have been reported: Lpx-B1.1a (Genbank HM126466), Lpx-B1.1b (Genbank HM126468), and Lpx-B1.1c (Genbank HM126470) for the Lpx-B1.1 locus, and Lpx-B1.2 (Genbank HM126467) and Lpx-B1.3 (Genbank HM126469) (Hessler et al., 2002; Carrera et al., 2007; Verlotta et al., 2010).
QTL analyses in durum wheat showed that 36–54% of the variation in LPX activity is explained by Lpx-B1 (Carrera et al., 2007; Verlotta et al., 2010). Carrera et al. (2007) reported a deletion in the Lpx-B1.1 gene, later named Lpx-B1.1c allele, which possesses a deletion covering from the second intron up to the last exon. This allele correlates with higher levels of pasta yellowness, due to the large deletion on its sequence, but it is not correlated with semolina or flour color, suggesting that the role of lipoxygenase on carotenoid degradation occurs in the pasta-making process rather than in the development of the grains (Carrera et al., 2007; Verlotta et al., 2010).
Verlotta et al. (2010) genotyped the presence of the Lpx-B1.1 alleles in combination with either Lpx-B1.2 or Lpx-B1.3 in a diverse modern/old durum wheat population, and found three haplotypes: haplotype I (Lpx-B1.3 and Lpx-B1.1b), haplotype II (Lpx-B1.2 and Lpx-B1.1a), and haplotype III (Lpx-B1.2 and Lpx-B1.1c), exhibiting high, intermediate, and low levels of functional Lpx-B1 transcripts and enzymatic activity in mature grains, respectively, which is also correlated with β-carotene bleaching. Carrera et al. (2007) reported sequences corresponding to the Lpx-2 and Lpx-3 on chromosome group 5 and 4, respectively, and Lpx-A3 showed significant effects on semolina color, but not on LPX mature grain activity, proving that the LPX activity given by Lpx-A3 acts earlier on grain development.
PPO (EC 1.14.18.1) catalyzes the oxidation of phenolic acids, producing short-chain polymers related to undesirable browning or darkening of pasta products, reducing its apparent quality (Watanabe et al., 2006). There are two non-linked genes controlling PPO activity on durum wheat that have been identified on the homoeologous chromosome 2A and 2B (Jimenez and Dubcovsky, 1999; Nair and Tomar, 2001; Simeone et al., 2002; Watanabe et al., 2004). Watanabe et al. (2006) reported that the locus on chromosome 2A was linked to PPO activity explaining 49.1% of the trait variation in an RIL population made from the crosses between Jennah Khetifa and Cham 1. Two Ppo paralogous families were mapped on the homoeologous group 2, named Ppo-1 (Ppo-A1 and Ppo-B1) and Ppo-2 (Ppo-A2 and Ppo-B2) (Beecher et al., 2012). Ppo-A1 was found to have a major role in PPO activity (Simeone et al., 2002; Sun et al., 2005; Anderson et al., 2006; He et al., 2007). Using a marker developed for common wheat (PPO18) (Sun et al., 2005), four Ppo-A1 alleles, named Ppo-A1b, Ppo-A1f, Ppo-A1e, and Ppo-A1g, were detected on durum wheat (He et al., 2009). Taranto et al. (2012) linked the different Ppo-A1 alleles to distinct levels of PPO activity in 113 accessions of tetraploid wheat. Ppo-A1f was associated to high PPO activity, whereas Ppo-A1b and Ppo-A1g were related to low PPO activity, although they argued there was a consistent variability on the PPO activities associated with each allele. Using the reverse primer of marker PPO18 and a new forward primer specific for the Ppo-A1 allele, Taranto et al. (2012) developed a new marker (MG18) able to detect the same alleles than PPO18 in a collection of 228 accessions of old, intermediate, and modern tetraploid wheats, but in a more efficient manner, reducing the variability on PPO activity of each allele-related group. Further decrease on PPO activity can be achieved by selecting also alleles for low PPO activity of the Ppo-B1 and Ppo-B2 paralogous genes, using markers MG08 and MG33, respectively, developed by Taranto et al. (2015). Ppo-B1 and Ppo-B2 were located 11 cM apart on chromosome arm 2BL. Marker MG08 identified four Ppo-B1 alleles, related to high (Ppo-B1c) and low (Ppo-B1a, Ppo-B1b, and Ppo-B1d) PPO activity, whereas marker MG33 recognized two Ppo-B2 alleles, associated to high (Ppo-B2d) and low (Ppo-B2a) PPO activity levels. The use of these markers in MAS breeding programs has the potential to further improve the color of pasta and durum wheat derived end products.
The aldehyde oxidase 3 (AO-A3) gene, located on chromosome 7AL, has been significantly associated to YI and to a QTL linked for YPC (Colasuonno et al., 2017b). Aldehyde oxidase enzymes (AO; EC 1.2.3.1) play roles in the final catalytic steps from carotenoids to ABA (Seo and Koshiba, 2002). qRT-PCR experiments revealed higher levels of AO-A3 expression in the low YPC cultivar Ciccio in comparison to the high YPC cultivar Svevo. This gene also showed higher expression levels in the later stages of seed formation than AO-A1 and AO-A2, suggesting a major role in the final stages of seed development (Colasuonno et al., 2017b). Colasuonno et al. (2017b) developed a marker for AO-A3 for DHPLC, which could be useful for MAS programs.
Peroxidase (PER) genes have received less attention than the other carotenoid degradation genes on durum wheat, with most of the studies being conducted on common wheat, and no durum wheat specific markers for low peroxidase activity are available to date. Peroxidases (EC 1.11.1.7.) are oxidoreductases that oxidize a vast array of compounds present in plants, using hydrogen peroxide as substrate. They are related to pasta brownness due to the oxidation of phenolic substances. Studies showed that peroxidase genes are located on the homoeologous chromosome groups 1, 2, 3, 4, and 7 (Kobrehel and Feillet, 1975; Benito and de la Vega, 1979; Bosch et al., 1987; Liu et al., 1990; Wei et al., 2015). Up to 12 peroxidase isoforms are present in the durum grain, varying in quantity during kernel development, maturation, and germination, with some isoforms having specific locations between milling fractions (bran, semolina, and embryo), in which isoform P-5 is of importance because of its endosperm specific location, contributing to the darkening of pasta products (Feillet et al., 2000; Fraignier et al., 2000). Fortunately, PER do not show activity during pasta processing, likely due to the unavailability of hydrogen peroxide, its main substrate, whereas it is abundant in semolina (Feillet et al., 2000; Ficco et al., 2014).
Novel Mutations in Carotenoid Genes
New advances in wheat genomics resources and in molecular technologies contributed to increase the knowledge of carotenoid genes. This included the screening of mutant resource containing chemically induced point mutation variation in candidate genes through TILLING strategy (Targeting Induced Local Lesions in Genomes) (Uauy et al., 2009; Colasuonno et al., 2016; Richaud et al., 2018).
Among all genes involved in the carotenoid pathway, LCYE and LCYB were the only genes extensively studied with this approach, highlighting the complexity of the trait and the difficulty of its modification.
Colasuonno et al. (2016) screened 1,140 mutant lines (0.70–0.85% ethyl methane sulfonate, EMS) focusing on these two target genes. The denaturing high-performance liquid chromatography (DHPLC) allowed to identify a total of 38 and 21 mutations for LCYE and LCYB genes, respectively, with a mutation density of 1/77 kb. Similarly, the analysis of 1,370 DNA individuals from the durum wheat Kronos TILLING mutant population (Uauy et al., 2009) allowed to identify 39 mutants for the LCYE homologues (Richaud et al., 2018) through CelI/agarose method.
Although in both studies, premature stop codons or deleterious missense mutations had been identified, no significant differences in the increment of β-carotene and total carotenoid content among the lines and the relative control were detected. This could be attributed to a high number of effective genes underlying this complex trait and their influence on the final phenotype.
Additional availability of an in silico TILLING population (Krasileva et al., 2017) allowed to identify knockout alleles in these target genes and give information about their mutation rate. For instance, of over 1,500 EMS mutagenized lines from the Kronos cultivar, 76 and 128 mutations had been detected in the LCYE and LCYB protein coding regions, respectively. Among all these mutations, only for LCYE gene 7 premature stop codons or deleterious missense mutations resulted to have significant effect on the change of protein composition. The low rate of deleterious SNPs for the target genes marked their main role into the biosynthesis and how some unknown mechanism prevented mutations in these key carotenoid enzymes.
Extensive studies specific for other carotenoid genes are needed to understand the available mutations and their potential effects if combined in double mutants on the final phenotype.
Transfer of QTL or Genes Linked to High Ypc
Backcross breeding has been used to transfer gene(s) or QTL of interest from a certain genetic background into an elite cultivar lacking for carotenoids. Subsequently, MAS technology validated the additive effect of the locus/candidate gene and assessed its impact on the new genetic background (Hospital, 2005).
Even though there is a great number of works of QTL linked to high YPC identification (Table 1), direct validation on durum wheat and use through introgression is limited. Patil et al. (2018) developed a marker, PSY-1SSR, based on the microsatellite variations in the promoter region of Psy-1, allowing the identification of eight alleles of Psy-A1 and seven alleles of Psy-B1 simultaneously, linked to Qyp.macs-7A, a major QTL for YPC on the long arm of chromosome 7A identified in a PDW 233/Bhalegaon 4 RIL population (Patil et al., 2018). They used this marker to improve YPC through MAS of two different low YPC Indian cultivars, MACS 3125 and HI 8498, and they were able to follow the introgression of the allele Psy-A1SSRe (linked to high yellowness) from PDW 23, using backcross breeding. MACS 3125 backcrossed lines showed a significant increase in YPC (6.16–7.7 ppm) over the recurrent parent MACS 3125 (3.57 ppm). HI 8498 introgressed lines also showed a significant YPC increase (5.0–7.46 ppm) in comparison to their recurrent parent (3.26 ppm).
MAS is currently being used by CIMMYT and by the Canadian durum wheat breeding programs (Randhawa et al., 2013; N’Diaye et al., 2018) by selecting materials with low LOX activity, with the implementation of the LOXA marker (Carrera et al., 2007), targeting the Lpx-B1.1c allele (Verlotta et al., 2010) for the generation of breeding lines (Randhawa et al., 2013; Dreisigacker et al., 2016).
Even though there is a great number of works of QTL linked to high YPC identification (Table 1), direct validation on durum wheat and use through introgression is limited. Patil et al. (2018) developed a marker, PSY-1SSR, based on the microsatellite variations in the promoter region of Psy-1, allowing the identification of eight alleles of Psy-A1 and seven alleles of Psy-B1 simultaneously, linked to Qyp.macs-7A, a major QTL for YPC on the long arm of chromosome 7A identified in a PDW 233/Bhalegaon 4 RIL population (Patil et al., 2018). They used this marker to improve YPC through MAS of two different low YPC Indian cultivars, MACS 3125 and HI 8498, and they were able to follow the introgression of the allele Psy-A1SSRe (linked to high yellowness) from PDW 23, using backcross breeding. MACS 3125 backcrossed lines showed a significant increase in YPC (6.16–7.7 ppm) over the recurrent parent MACS 3125 (3.57 ppm). HI 8498 introgressed lines also showed a significant YPC increase (5.0–7.46 ppm) in comparison to their recurrent parent (3.26 ppm).
MAS is currently being used by CIMMYT and by the Canadian durum wheat breeding programs (Randhawa et al., 2013; N’Diaye et al., 2018) by selecting materials with low LOX activity, with the implementation of the LOXA marker (Carrera et al., 2007), targeting the Lpx-B1.1c allele (Verlotta et al., 2010) for the generation of breeding lines (Randhawa et al., 2013; Dreisigacker et al., 2016).
Future Perspectives and Conclusions
Understanding the biosynthetic pathway for carotenoid pigment accumulation requires many efforts due to the durum wheat polyploidy and its quantitative nature. The information examined in this article explains the significant goals that have been reached in the last two decades in understanding the genetic and the molecular mechanisms underlying the metabolism and regulation of wheat carotenoids. Furthermore, the characterization of specific plant materials and the release of the durum wheat genome sequences, together with the development of more accurate classes of DNA-based markers and consensus maps, have allowed the identification of important genes involved in the control of carotenoid biosynthesis and catabolism.
Clearly, the most studied and repeatable QTL are those located on chromosomes 3AS (linked to the LCYE gene), 7AL, and 7BL (both tightly linked to the PSY1 genes). Diagnostic markers are available in all these regions for MAS application. Hopefully, other carotenoid QTL regions will likely be further characterized in the future, taking advantage of the recent results and tools for identifying the candidate genes involved in the accumulation/degradation of the carotenoid compounds. This will certainly increase the speed of the genetic gains of carotenoid accumulation, which will benefit the breeding programs and the pasta industry. According to these new resources, we can anticipate an implementation in genotypic selection flanking the traditional phenotypic selection in the durum wheat breeding programs. At the same time, the additive effects of the genes involved in yellowness will generate improved plants through several breeding cycles able to incorporate the beneficial alleles introgressed. Future developments on MAS breeding will focus on selecting many genes alleles at once in order to reach such purpose. Despite all the research in this subject, efforts should be taken on the transfer of knowledge between the bench and the field, because of the current use of the markers described in this review that could potentially benefit the durum wheat breeding programs globally.
Finally, further emphasis of future activities will encompass the analysis of the genetic variability present in the durum wheat germplasm collections (i.e., pre-breeding work), and the TILLING populations, to better understand the functions of the genes involved in the structural and the regulatory system responsible for the YPC trait. Advanced techniques (i.e., CRISPR-Cas9–based genome editing method), will be useful if combined and used to understand the homoeologous silenced gene acting additively and imposing effects on both the total gene expression and the resulting phenotype. Taking these strategies together, the characterization of each gene could provide opportunities for diversifying the genetic architecture of carotenoid pigments and expand the existing allelic variation available for wheat improvement.
Author Contributions
PC, AG, RP, and AS designed the review. MM, GC, RT, AB, IM, JA-F, and AC prepared the manuscript. All authors read and approved the final manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The article processing charge was funded by Conicyt (Fondecyt Regular n° 1161298). This work was partially supported by Conicyt [grants Fondecyt Regular n° 1161298 (AS) and Fondecyt Postdoctorado n° 3180432 (AC)]. This research was also partially funded by a research grant from ISEA AGROSERVICE SpA (S. Severino Marche, MC, Italy).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2019.01347/full#supplementary-material
References
AACC International. (2013). “AACC International Official Method 14-50.01”, In: Approved Methods of the American Association of Cereal Chemists, tenth ed. (MN, U.S.A: St. Paul).
Al-Babili, S., Bouwmeester, H. J. (2015). Strigolactones, a Novel Carotenoid-Derived Plant Hormone. Annu. Rev. Plant Biol. 66 (1), 161–186. doi: 10.1146/annurev-arplant-043014-114759
Anderson, J. V., Patrick Fuerst, E., Hurkman, W. J., Vensel, W. H., Morris, C. F. (2006). Biochemical and genetic characterization of wheat (Triticum spp.) kernel polyphenol oxidases. J. Cereal Sci. 44, 353–367. doi: 10.1016/j.jcs.2006.06.008
Asins, M. J., Perez de la Vega, M. (1985). The inheritance of tetraploid wheat seed peroxidases. Theor. Appl. Genet. 71, 61–67. doi: 10.1007/BF00278255
Atienza, S. G., Ballesteros, J., Martin, A., Hornero-Mendes, D. (2007). Genetic variability of carotenoid concentration and degree of esterification among Tritordeum (xTritordeum Ascherson et Graebner) and durum wheat accessions. Agric Food Chem. doi: 10.1021/jf070342p
Beecher, B. S., Carter, A. H., See, D. R. (2012). Genetic mapping of new seed-expressed polyphenol oxidase genes in wheat (Triticum aestivum L.). Theor. Appl. Genet. 124, 1463–1473. doi: 10.1007/s00122-012-1801-2
Beleggia, R., Platani, C., Nigro, F., Papa, R. (2011). Yellow pigment determination for single kernels of durum wheat (Triticum durum Desf.). Cereal Chem. 88, 504–508. doi: 10.1094/cchem-02-11-0013
Bendich, A., Olson, J. A. (1989). Biological actions of carotenoids. FASEB J. 3, 1927–1932. doi: 10.1096/fasebj.3.8.2656356
Benito, C., de la Vega, M. P. (1979). The chromosomal location of peroxidase isozymes of the wheat kernel. Theor. Appl. Genet. 55, 73–76. doi: 10.1007/bf00285193
Blanco, A., Colasuonno, P., Gadaleta, A., Mangini, G., Schiavulli, A., Simeone, R., et al. (2011). Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds in durum wheat. J. Cereal Sci. 54, 255–264. doi: 10.1016/j.jcs.2011.07.002
Borrelli, G. M., De Leonardis, A. M., Fares, C., Platani, C., Di Fonzo, N. (2003). Effects of modified processing conditions on oxidative properties of semolina dough and pasta. Cereal Chem. 80, 225–231. doi: 10.1094/cchem.2003.80.2.225
Borrelli, G. M., De Leonardis, A. M., Platani, C., Troccoli, A. (2008). Distribution along durum wheat kernel of the components involved in semolina colour. J. Cereal Sci. 48, 494–502. doi: 10.1016/j.jcs.2007.11.007
Borrelli, G. M., Trono, D. (2016). Molecular approaches to genetically improve the accumulation of health-promoting secondary metabolites in staple crops-A case study: the Lipoxygenase-B1 genes and regulation of the carotenoid content in pasta products. Int. J. Mol. Sci. 17, 1177. doi: 10.3390/ijms17071177
Bosch, A., Vega, C., Benito, C. (1987). The peroxidase isozymes of the wheat kernel: tissue and substrate specificity and their chromosomal location. Theor. Appl. Genet. 73, 701–706. doi: 10.1007/bf00260779
Brandolini, A., Hidalgo, A., Moscaritolo, S. (2008). Chemical composition and pasting properties of einkorn (Triticum monococcum L. subsp. monococcum) whole meal flour. J. Cereal Sci. 47, 599–609. doi: 10.1016/j.jcs.2007.07.005
Brandolini, A., Hidalgo, A., Gabriele, S., Heun, M. (2015). Chemical composition of wild and feral diploid wheats and their bearing on domesticated wheats. J. Cereal Sci. 63, 122–127. doi: 10.1016/j.jcs.2015.03.005
Breseghello, F., Sorrells, M. E. (2006). Association mapping of kernel size and milling quality in wheat (Triticum aestivum L.) cultivars. Genetics. 172(2), 1165–1177.
Britton, G. (1995). Structure and properties of carotenoids in relation to function. FASEB J. 9, 1551–1558. doi: 10.1096/fasebj.9.15.8529834
Britton, G. (1998). “Overview of carotenoid biosynthesis,” in Carotenoids: Biosynthesis and Metabolism. Eds. Britton, G., Liaaen-Jenen, S., Pfander, H. (Basel, Switzerland: Birkhauser Verlag), 13–147.
Britton, G., Khachik, F., (2009). “Carotenoids in Food,” in Carotenoids: Nutrition and Health, vol. 5 . Eds. Britton, G., Pfander, H., Liaaen-Jensen, S. (Basel: Birkhäuser Basel), 45–66.
Burkhardt, S., Böhm, V. (2007). Development of a new method for the complete extraction of carotenoids from cereals with special reference to durum wheat (Triticum durum Desf.). J. Agric. Food Chem. 55, 8295–8301. doi: 10.1021/jf0712853
Bush, W. S., Moore, J. H. (2012). Chapter 11: genome-wide association studies. PloS Comput. Biol. 8, e1002822. doi: 10.1371/journal.pcbi.1002822
Campos, K. M., Royo, C., Schulthess, A., Villegas, D., Matus, I., Ammar, K., et al (2016). Association of phytoene synthase Psy1-A1 and Psy1-B1 allelic variants with semolina yellowness in durum wheat (Triticum turgidum L. var. durum). Euphytica 207, 109–117. doi: 10.1007/s10681-015-1541-x
Carrera, A., Echenique, V., Zhang, W., Helguera, M., Manthey, F., Schrager, A., et al. (2007). A deletion at the Lpx-B1 locus is associated with low lipoxygenase activity and improved pasta color in durum wheat (Triticum turgidum ssp. durum). J. Cereal Sci. 45, 67–77. doi: 10.1016/j.jcs.2006.07.001
Cazzonelli, C. I., Pogson, B. J. (2010). Source to sink: regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 15, 266–274. doi: 10.1016/j.tplants.2010.02.003
Colasuonno, P., Gadaleta, A., Giancaspro, A., Nigro, D., Giove, S., Incerti, O., et al. (2014). Development of a high-density SNP-based linkage map and detection of yellow pigment content QTLs in durum wheat. Mol. Breed. 34, 1563–1578. doi: 10.1007/s11032-014-0183-3
Colasuonno, P., Incerti, O., Lozito, M. L., Simeone, R., Gadaleta, A., Blanco, A. (2016). DHPLC technology for high-throughput detection of mutations in a durum wheat TILLING population. BMC Genet. 17, 17:43. doi: 10.1186/s12863-016-0350-0
Colasuonno, P., Lozito, M. L., Marcotuli, I., Nigro, D., Giancaspro, A., Mangini, G., et al. (2017a). The carotenoid biosynthetic and catabolic genes in wheat and their association with yellow pigments. BMC Genomics 18, 122. doi: 10.1186/s12864-016-3395-6
Colasuonno, P., Marcotuli, I., Lozito, M. L., Simeone, R., Blanco, A., Gadaleta, A. (2017b). Characterization of aldehyde oxidase (AO) genes involved in the accumulation of carotenoid pigments in wheat grain. Front. Plant Sci. 8, 863. doi: 10.3389/fpls.2017.00863
Cong, L., Wang, C., Li, Z., Chen, L., Yang, G., Wang, Y., et al. (2010). cDNA cloning and expression analysis of wheat (Triticum aestivum L.) phytoene and ζ-carotene desaturase genes. Mol. Biol. Rep. 37, 3351–3361. doi: 10.1007/s11033-009-9922-7
Crawford, A. C., Francki, M. G. (2013). Chromosomal location of wheat genes of the carotenoid biosynthetic pathway and evidence for a catalase gene on chromosome 7A functionally associated with flour b* colour variation. Mol. Genet. Genomics 288, 483–493. doi: 10.1007/s00438-013-0767-3
Choe, E., Min, D. B. (2005). Chemistry and reactions of reactive oxygen species in foods. J. Food Sci. 70, R142–R159. doi: 10.1111/j.1365-2621.2005.tb08329.x
De Simone, V., Menzo, V., De Leonardis, A. M., Ficco, D. B. M., Trono, D., Cattivelli, L., et al. (2010). Different mechanisms control lipoxygenase activity in durum wheat kernels. J. Cereal Sci. 52, 121–128. doi: 10.1016/j.jcs.2010.04.003
DellaPenna, D., Pogson, B. J. (2006). Vitamin synthesis in plants: tocopherols and carotenoids. Annu. Rev. Plant Biol. 57, 711–738. doi: 10.1146/annurev.arplant.56.032604.144301
Dibari, B., Murat, F., Chosson, A., Gautier, V., Poncet, C., Lecomte, P., et al. (2012). Deciphering the genomic structure, function and evolution of carotenogenesis related phytoene synthases in grasses. BMC Genomics 13, 221. doi: 10.1186/1471-2164-13-221
Digesù, A. M., Platani, C., Cattivelli, L., Mangini, G., Blanco, A. (2009). Genetic variability in yellow pigment components in cultivated and wild tetraploid wheats. J. Cereal Sci. 50, 210–218. doi: 10.1016/j.jcs.2009.05.002
Dong, C. H., Ma, Z. Y., Xia, X. C., Zhang, L. P., He, Z. H. (2012). Allelic variation at the TaZds-A1 locus on wheat chromosome 2A and development of a functional marker in common wheat. J. Integr. Agric. 11, 1067–1074. doi: 10.1016/S2095-3119(12)60099-9
Dreisigacker, S., Sehgal, D., Reyes Jaimez, A., Luna Garrido, B., Muñoz Zavala, S., Núñez Ríos, C., et al., (2016). “CIMMYT Wheat Molecular Genetics,” in Laboratory Protocols and Applications to Wheat Breeding (Mexico: CIMMYT).
Elouafi, I., Nachit, M. M., Martin, L. M. (2001). Identification of a microsatellite on chromosome 7B showing a strong linkage with yellow pigment in durum wheat (Triticum turgidum L. var. durum). Hereditas 135, 255–261. doi: 10.1111/j.1601-5223.2001.t01-1-00255.x
Feillet, P., Autran, J.-C., Icard-Vernière, C. (2000). MINI REVIEW Pasta brownness: an Assessment. J. Cereal Sci. 32, 215–233. doi: 10.1006/jcrs.2000.0326
Ficco, D. B. M., Mastrangelo, A. M., Trono, D., Borrelli, G. M., De Vita, P., Fares, C., et al. (2014). The colours of durum wheat: a review. Crop Pasture Sci. 65, 1–15. doi: 10.1071/cp13293
Fiedler, J. D., Salsman, E., Liu, Y., Michalak de Jiménez, M., Hegstad, J. B., Chen, B., et al. (2017). Genome-wide association and prediction of grain and semolina quality traits in durum wheat breeding populations. Plant Genome 10 (3), 1–12. doi: 10.3835/plantgenome2017.05.0038.
Fraignier, M.-P., Michaux-Ferrière, N., Kobrehel, K. (2000). Distribution of peroxidases in durum wheat (Triticum durum). Cereal Chem. 77, 11–17. doi: 10.1094/cchem.2000.77.1.11
Fu, B. X., Chiremba, C., Pozniak, C. J., Wang, K., Nam, S. (2017). Total phenolic and yellow pigment contents and antioxidant activities of durum wheat milling fractions. Antioxidants 6, 78. doi: 10.3390/antiox6040078
Gallagher, C. E., Matthews, P. D., Li, F., Wurtzel, E. T. (2004). Gene duplication in the carotenoid biosynthetic pathway preceded evolution of the grasses. Plant Physiol. 135, 1776–1783. doi: 10.1104/pp.104.039818
Garbus, I., Carrera, A. D., Dubcovsky, J., Echenique, V. (2009). Physical mapping of durum wheat lipoxygenase genes. J. Cereal Sci. 50, 67–73. doi: 10.1016/j.jcs.2009.02.008
Garbus, I., Soresi, D., Romero, J., Echenique, V. (2013). Identification, mapping and evolutionary course of wheat lipoxygenase-1 genes located on the A genome. J. Cereal Sci. 58, 298–304. doi: 10.1016/j.jcs.2013.05.012
Guaadaoui, A., Benaicha, S., Elmajdoub, N., Bellaoui, M., Hamal, A. (2014). What is a bioactive compound? A combined definition for a preliminary consensus. Int. J. Nutr. Food Sci. 3, 174–179. doi: 10.11648/j.ijnfs.20140303.16
Gupta, P. K., Rustgi, S., Kulwal, P. L. (2005). Linkage disequilibrium and association studies in higher plants: Present status and future prospects. Plant Mol. Biol. 57, 461–485. doi: 10.1007/s11103-005-0257-z
Halliwell, B. (2011). Free radicals and antioxidants - quo vadis? Trends Pharmacol. Sci. 32, 125–130. doi: 10.1016/j.tips.2010.12.002
He, X. Y., He, Z. H., Morris, C. F., Xia, X. C. (2009). Cloning and phylogenetic analysis of polyphenol oxidase genes in common wheat and related species. Genet. Resour. Crop Evol. 56, 311. doi: 10.1007/s10722-008-9365-3
He, X. Y., He, Z. H., Zhang, L. P., Sun, D. J., Morris, C. F., Fuerst, E. P., et al. (2007). Allelic variation of polyphenol oxidase (PPO) genes located on chromosomes 2A and 2D and development of functional markers for the PPO genes in common wheat. Theor. Appl. Genet. 115, 47–58. doi: 10.1007/s00122-007-0539-8
He, X. Y., Zhang, Y. L., He, Z. H., Wu, Y. P., Xiao, Y. G., Ma, C. X., et al. (2008). Characterization of phytoene synthase 1 gene (Psy1) located on common wheat chromosome 7A and development of a functional marker. Theor. Appl. Genet. 116, 213–221. doi: 10.1007/s00122-007-0660-8
Hentschel, V., Kranl, K., Hollmann, J., Lindhaeur, M. G., Bohm, V., Bitsch, R. (2002). Spectrophotometric determination of yellow pigment content and evaluation of carotenoids by high-performance liquid chromatography in durum wheat grain. J. Agric. Food Chem. 50, 6663–6668. doi: 10.1021/jf025701p
Hessler, T. G., Thomson, M. J., Benscher, D., Nachit, M., Sorrells, M., (2002). Association of a lipoxygenase locus, Lpx-B1, with variation in lipoxygenase activity in durum wheat seeds. Crop Sci. 42, 1695–1700. doi: 10.2135/cropsci2002.1695.
Holtman, W., Gert van, D., Norbert, J. A. S., Anneke, C. D. (1996). Differential expression of lipoxygenase isoenzymes in embryos of germinating barley. Plant Physiol. 111, 569–576. doi: 10.1104/pp.111.2.569
Hospital, F. (2005). Selection in backcross programmes. Philos. Trans. R. Soc Lond. B. Biol. Sci. 360, 1503–1511. doi: 10.1098/rstb.2005.1670
Howitt, C. A., Cavanagh, C. R., Bowerman, A. F., Cazzonelli, C., Rampling, L., Mimica, J. L., et al. (2009). Alternative splicing, activation of cryptic exons and amino acid substitutions in carotenoid biosynthetic genes are associated with lutein accumulation in wheat endosperm. Funct. Integr. Genomics 9, 363–376. doi: 10.1007/s10142-009-0121-3
ICC. (1990). “ICC Method 152,” in Standard Methodsof the International Association for Cereal Science and Technology (Detmold, Germany: (Verlag Moritz SchäferICC)).
Irakli, M. N., Samanidou, V. F., Papadoyannis, I. N. (2011). Development and validation of an HPLC method for the simultaneous determination of tocopherols, tocotrienols and carotenoids in cereals after solid-phase extraction. J. Sep. Sci. 34, 1375–1382. doi: 10.1002/jssc.201100077
Jahns, P., Holzwarth, A. R. (2012). The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 1817, 182–193. doi: 10.1016/j.bbabio.2011.04.012
Jimenez, M., Dubcovsky, J. (1999). Chromosome location of genes affecting polyphenol oxidase activity in seeds of common and durum wheat. Plant Breed. 118, 395–398. doi: 10.1046/j.1439-0523.1999.00393.x
Kabbaj, H., Sall, A. T., Al-Abdallat, A., Geleta, M., Amri, A., Filali-Maltouf, A., et al. (2017). Genetic diversity within a Global panel of durum wheat (Triticum durum) landraces and modern germplasm reveals the history of alleles exchange. Front. Plant Sci. 8, 1277. doi: 10.3389/fpls.2017.01277
Ke, Q., Kang, L., Kim, H. S., Xie, T., Liu, C., Ji, C. Y., et al. (2019). Down-regulation of lycopene ε-cyclase expression in transgenic sweetpotato plants increases the carotenoid content and tolerance to abiotic stress. Plant Sci. 281, 52–60. doi: 10.1016/j.plantsci.2019.01.002
Kean, E. G., Bordenave, N., Ejeta, G., Hamaker, B. R., Ferruzzi, M. G. (2011). Carotenoid bioaccessibility from whole grain and decorticated yellow endosperm sorghum porridge. J. Cereal Sci. 54, 450–459. doi: 10.1016/j.jcs.2011.08.010
Kelly, M. E., Ramkumar, S., Sun, W., Colon Ortiz, C., Kiser, P. D., Golczak, M., et al. (2018). The biochemical basis of vitamin A production from the asymmetric carotenoid β-cryptoxanthin. ACS Chem. Biol. 13, 2121–2129. doi: 10.1021/acschembio.8b00290
Khoo, H. E., Prasad, K. N., Kong, K. W., Jiang, Y., Ismail, A. (2011). Carotenoids and their isomers: color pigments in fruits and vegetables. Molecules 16, 1710–1738. doi: 10.3390/molecules16021710
Kobrehel, K., Feillet, P. (1975). Identification of genomes and chromosomes involved in peroxidase synthesis of wheat seeds. Can. J. Bot. 53, 2336–2344. doi: 10.1139/b75-259
Konopka, I., Kozirok, W., Rotkiewicz, D. (2004). Lipids and carotenoids of wheat grain and flour and attempt of correlating them with digital image analysis of kernel surface and cross-sections. Food Res. Int. 37, 429–438. doi: 10.1016/j.foodres.2003.12.009
Krasileva, K. V., Vasquez-Gross, H. A., Howell, T., Bailey, P., Paraiso, F., Clissold, L., et al. (2017). Uncovering hidden variation in polyploidy wheat. Proceed Nat. Acad. Sci. 114, E913. doi: 10.1073/pnas.1619268114
Lachman, J., Martinek, P., Kotikova, Z., Orsák, M., Šulc, M. (2017). Genetics and chemistry of pigments in wheat grain -A review. J. Cereral Sci. 74, 145–154. doi: 10.1016/j.jcs.2017.02.007
Landrum, J. T. (2009). “Carotenoids: physical, chemical, and biological functions and properties,”. Ed. Landrum, J. T. (Boca Raton, FL: CRC Press). doi: 10.1201/9781420052312
Leenhardt, F., Lyan, B., Rock, E., Boussard, A., Potus, J., Chanliaud, E., et al. (2006). Genetic variability of carotenoid concentration, and lipoxygenase and peroxidase activities among cultivated wheat species and bread wheat varieties. Eur. J. Agron. 25, 170–176. doi: 10.1016/j.eja.2006.04.010
Leong, H. Y., Show, P. L., Lim, M. H., Ooi, C. W., Ling, T. C. (2018). Natural red pigments from plants and their health benefits: a review. Food Rev. Int. 34, 463–482. doi: 10.1080/87559129.2017.1326935
Lipka, A. E., Buckler, E. S., Tian, F., Peiffer, J., Li, M., Gore, M. A., et al. (2012). GAPIT: genome association and prediction integrated tool. Bioinformatics 28, 2397–2399. doi: 10.1093/bioinformatics/bts444
Liu, C. J., Chao, S., Gale, M. D. (1990). The genetical control of tissue-specific peroxidases, Per-1, Per-2, Per-3, Per-4, and Per-5 in wheat. Theor. Appl. Genet. 79 (3), 305–313. doi: 10.1007/bf01186072
Maccaferri, M., Sanguineti, M. C., Natoli, V., Ortega, J. L. A., Salem, M. B., Bort, J., et al. (2006). A panel of elite accessions of durum wheat (Triticum durum Desf.) suitable for association mapping studies. Plant Gen. Res. 4 (1), 79–85.
Maccaferri, M., Cane’, M. A., Sanguineti, M. C., Salvi, S., Colalongo, M. C., Massi, A., et al. (2014). A consensus framework map of durum wheat (Triticum durum Desf.) suitable for linkage disequilibrium analysis and genome-wide association mapping. BMC Genomics 15, 873. doi: 10.1186/1471-2164-15-873
Maccaferri, M., Harris, N. S., Twardziok, S. O., Pasam, R. K., Gundlach, H., Spannagl, M., et al. (2019). Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 51(5), 885–895. doi: 10.1038/s41588-019-0381-3
Maccaferri, M., Sanguineti, M. C., Demontis, A., El-Ahmed, A., Garcia del Moral, L., Maalouf, F., et al. (2011). Association mapping in durum wheat grown across a broad range of water regimes. J. Exp. Bot. 62(2), 409–438.
Mazzeo, M. F., Di Stasio, L., D’Ambrosio, C., Arena, S., Scaloni, A., Corneti, S., et al. (2017). Identification of early represented gluten proteins during durum wheat grain development. J. Agric. Food Chem. 65, 3242–3250. doi: 10.1021/acs.jafc.7b00571
McCaig, T. N., McLeod, J. G., Clarke, J. M., DePauw, R. M. (1992). Measurement of durum pigment with an NIR instrument operating in the visible range. Cereal Chem. 69, 671–672.
Mezzomo, N., Ferreira, S. R. S. (2016). Carotenoids functionality, sources, and processing by supercritical technology: a review. J. Chem. 1–16. doi: 10.1155/2016/3164312
Mezzomo, N., Tenfen, L., Farias, M. S., Friedrich, M. T., Pedrosa, R. C., Ferreira, S. R. S. (2015). Evidence of anti-obesity and mixed hypolipidemic effects of extracts from pink shrimp (Penaeus brasiliensis and Penaeus paulensis) processing residue. J. Supercrit. Fluids 96, 252–261. doi: 10.1016/j.supflu.2014.09.021
Morris, C. F. (2018). Determinants of wheat noodle color. J. Sci. Food Agric. 98, 5171–5180. doi: 10.1002/jsfa.9134
N’Diaye, A., Haile, J. K., Cory, A. T., Clarke, F. R., Clarke, J. M., Knox, R. E., et al. (2017). Single marker and haplotype-based association analysis of semolina and pasta colour in elite durum wheat breeding lines using a high-density consensus map. PloS One 12, e0170941. doi: 10.1371/journal.pone.0170941
N’Diaye, A., Haile, J. K., Nilsen, K. T., Walkowiak, S., Ruan, Y., Singh, A. K., et al. (2018). Haplotype loci under selection in Canadian durum wheat germplasm over 60 years of breeding: Association with grain yield, quality traits, protein loss, and plant height. Front. Plant Sci. 9 (1589), 1–19. doi: 10.3389/fpls.2018.01589
Nair, S. K., Tomar, S. M. S. (2001). Genetics of phenol colour reaction of grains and glumes in tetraploid and hexaploid wheats. J. Genet. Breed. 55, 369–373.
Nazco, R., Villegas, D., Ammar, K., Peña, R. J., Moragues, M., Royo, C. (2012). Can Mediterranean durum wheat landraces contribute to improved grain quality attributes in modern cultivars? Euphytica 185, 1–17. doi: 10.1007/s10681-011-0588-6
Nisar, N., Li, L., Lu, S., Khin, Nay, C., Pogson, Barry, J. (2015). Carotenoid metabolism in plants. Mol. Plant 8, 68–82. doi: 10.1016/j.molp.2014.12.007
Oliver, J., Palou, A. (2000). Chromatographic determination of carotenoids in foods. J. Chromatogr. A. 881, 543–555. doi: 10.1016/S0021-9673(00)00329-0
Olson, J. A., Krinsky, N. I. (1995). Introduction: the colorful, fascinating world of the carotenoids: important physiologic modulators. FASEB J. 9, 1547–1550. doi: 10.1096/fasebj.9.15.8529833
Panfili, G., Fratianni, A., Irano, M. (2004). Improved normal-phase high-performance liquid chromatography procedure for the determination of carotenoids in cereals. J. Agric. Food Chem. 52, 6373–6377. doi: 10.1021/jf0402025
Patil, R., Oak, M., Deshpande, A., Tamhankar, S. (2018). Development of a robust marker for Psy-1 homoeologs and its application in improvement of yellow pigment content in durum wheat. Mol. Breed. 38, 136. doi: 10.1007/s11032-018-0895-x
Patil, R. M., Oak, M. D., Tamhankar, S. A., Sourdille, P., Rao, V. S. (2008). Mapping and validation of a major QTL for yellow pigment content on 7AL in durum wheat (Triticum turgidum L. ssp durum). Mol. Breed. 21, 485–496. doi: 10.1007/s11032-007-9147-1
Paznocht, L., Kotikova, Z., Orsak, M., Lachman, J., Martinek, P. (2019). Carotenoid changes of colored-grain wheat flours during bun-making. Food Chem. 277, 725–734. doi: 10.1016/j.foodchem.2018.11.019
Pozniak, C. J., Knox, R. E., Clarke, F. R., Clarke, J. M. (2007). Identification of QTL and association of a phytoene synthase gene with endosperm colour in durum wheat. Theor. Appl. Genet. 114, 525–537. doi: 10.1007/s00122-006-0453-5
Qin, X., Zhang, W., Dubcovsky, J., Tian, L. (2012). Cloning and comparative analysis of carotenoid beta-hydroxylase genes provides new insights into carotenoid metabolism in tetraploid (Triticum turgidum ssp. durum) and hexaploid (Triticum aestivum) wheat grains. Plant Mol. Biol. 80, 631–646. doi: 10.1007/s11103-012-9972-4
Randhawa, H. S., Asif, M., Pozniak, C., Clarke, J. M., Graf, R. J., Fox, S. L., et al. (2013). Application of molecular markers to wheat breeding in Canada. Plant Breed. 132, 458–471. doi: 10.1111/pbr.12057
Reimer, S., Pozniak, C. J., Clarke, F. R., Clarke, J. M., Somers, D. J., Knox, R. E., et al. (2008). Association mapping of yellow pigment in an elite collection of durum wheat cultivars and breeding lines. Genome 51, 1016–1025. doi: 10.1139/g08-083
Richaud, D., Stange, C., Gadaleta, A., Colasuonno, P., Parada, R., Schwember, A. R. (2018). Identification of Lycopene epsilon cyclase (LCYE) gene mutants to potentially increase β-carotene content in durum wheat (Triticum turgidum L. ssp. durum) through TILLING. PloS One 13, e0208948. doi: 10.1371/journal.pone.0208948
Rodriguez-Amaya, D., Kimura, M., (2004). “HarvestPlus Handbook for Carotenoids Analysis,” in HarvestPlus Technical Monographs Series, vol. 2. (Washington DC, USA and Cali, Colombia). IFPRI CIAT,.
Rodriguez-Concepcion, M., Avalos, J., Bonet, M. L., Boronat, A., Gomez-Gomez, L., Hornero-Mendez, D., et al. (2018). A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 70, 62–93. doi: 10.1016/j.plipres.2018.04.004
Roncallo, P. F., Cervigni, G. L., Jensen, C., Miranda, R., Carrera, A. D., Helguera, M., et al. (2012). QTL analysis of main and epistatic effects for flour color traits in durum wheat. Euphytica 185, 77–92. doi: 10.1007/s10681-012-0628-x
Rosello, M., Royo, C., Alvaro, F., Villegas, D., Nazco, R., Soriano, J. M. (2018). Pasta-making quality QTLome from Mediterranean durum wheat landraces. Front. Plant Sci. 9, 1512. doi: 10.3389/fpls.2018.01512
Sandmann, G. (2001). Genetic manipulation of carotenoid biosynthesis: strategies, problems and achievements. Trends Plant Sci. 6, 14–17. doi: 10.1016/S1360-1385(00)01817-3
Schroeder, W. A., Johnson, E. A. (1995). Singlet oxygen and peroxyl radicals regulate carotenoid biosynthesis in Phaffia rhodozyma. J. Biol. Chem. 270, 18374–18379. doi: 10.1074/jbc.270.31.18374
Schulthess, A., Matus, I., Schwember, A. (2013). Genotypic and environmental factors and their interactions determine semolina color of elite genotypes of durum wheat (Triticum turgidum L. var. durum) grown in different environments of Chile. Field Crops Res. 149, 234–244. doi: 10.1016/j.fcr.2013.05.001
Schulthess, A., Schwember, A. R. (2013). Improving durum wheat (Triticum turgidum L. var durum) grain yellow pigment content through plant breeding. Cienc. Invest. Agr. 40, 475–490. doi: http://dx.doi.org/10.7764/rcia.v40i3.1157
Seo, M., Koshiba, T. (2002). Complex regulation of ABA biosynthesis in plants. Trends Plant Sci. 7, 41–48.
Shen, R., Yang, S., Zhao, G., Shen, Q., Diao, X. (2015). Identification of carotenoids in foxtail millet (Setaria italica) and the effects of cooking methods on carotenoid content. J. Cereal Sci. 61, 86–93. doi: 10.1016/j.jcs.2014.10.009
Si, H., Ma, C., Wang, X., He, X. (2012). Variability of polyphenol oxidase (PPO) alleles located on chromosomes 2A and 2D can change the wheat kernel PPO activity. Aust. J. Crop Sci. 6, 444–449.
Simeone, R., Pasqualone, A., Clodoveo, M., Blanco, A. (2002). Genetic mapping of polyphenol oxidase in tetraploid wheat. Cell. Mol. Biol. Lett. 7, 763–769.
Singh, A., Reimer, S., Pozniak, C. J., Clarke, F. R., Clarke, J. M., Knox, R. E., et al. (2009). Allelic variation at Psy1-A1 and association with yellow pigment in durum wheat grain. Theor. Appl. Genet. 118, 1539–1548. doi: 10.1007/s00122-009-1001-x
Sissons, M. (2008). Role of durum wheat composition on the quality of pasta and bread. Food 2, 75–90.
Sommer, A., Davidson, F. R. (2002). Assessment and control of vitamin A deficiency: The annecy accords. J. Nutr. 132, 2845S–2850S. doi: 10.1093/jn/132.9.2845S
Subira, J., Peña, R. J., Álvaro, F., Ammar, K., Ramdani, A., Royo, C. (2014). Breeding progress in the pasta-making quality of durum wheat cultivars released in Italy and Spain during the 20th Century. Crop Pasture Sci. 65, 16–26. doi: 10.1071/CP13238
Sun, D. J., He, Z. H., Xia, X. C., Zhang, L. P., Morris, C. F., Appels, R., et al. (2005). A novel STS marker for polyphenol oxidase activity in bread wheat. Mol. Breed. 16, 209–218. doi: 10.1007/s11032-005-6618-0
Sun, T., Yuan, H., Cao, H., Yazdani, M., Tadmor, Y., Li, L. (2018). Carotenoid metabolism in plants: The role of plastids. Mol. Plant 11, 58–74. doi: 10.1016/j.molp.2017.09.010
Taranto, F., Delvecchio, L. N., Mangini, G., Del Faro, L., Blanco, A., Pasqualone, A. (2012). Molecular and physico-chemical evaluation of enzymatic browning of whole meal and dough in a collection of tetraploid wheats. J. Cereal Sci. 55, 405–414. doi: 10.1016/j.jcs.2012.02.005
Taranto, F., Mangini, G., Pasqualone, A., Gadaleta, A., Blanco, A. (2015). Mapping and allelic variations of Ppo-B1 and Ppo-B2 gene-related polyphenol oxidase activity in durum wheat. Mol. Breed. 35, 80. doi: 10.1007/s11032-015-0272-y
Tsilo, T., Hareland, A., Chao, S., Anderson, J. A. (2011). Genetic mapping and QTL analysis of flour color and milling yield related traits using recombinant inbred lines in hard red spring wheat. Crop Sci. 51, 237–246. doi: 10.2135/cropsci2009.12.0711
Uauy, C., Paraiso, F., Colasuonno, P., Tran, R. K., Tsai, H., Berardi, S., et al. (2009). A modified TILLING approach to detect induced mutations in tetraploid and hexaploid wheat. BMC Plant Biol. 9 (115), 1–14. doi: 10.1186/1471-2229-9-115
van den Berg, H., Faulks, R., Granado, H. F., Hirschberg, J., Olmedilla, B., Sandmann, G., et al. (2000). The potential for the improvement of carotenoid levels in foods and the likely systemic effects. J. Sci. Food Agric. 80, 880–912.
Vargas, V. H., Schulthess, A., Royo, C., Matus, I., Schwember, A. R. (2016). Transcripts levels of Phytoene synthase 1 (Psy-1) are associated to semolina yellowness variation in durum wheat (Triticum turgidum L. ssp. J. Cereal Sci. 68, 155–163. doi: 10.1016/j.jcs.2016.01.011
Verlotta, A., De Simone, V., Mastrangelo, A. M., Cattivelli, L., Papa, R., Trono, D. (2010). Insight into durum wheat Lpx-B1: a small gene family coding for the lipoxygenase responsible for carotenoid bleaching in mature grains. BMC Plant Biol. 10, 263. doi: 10.1186/1471-2229-10-263
Wang, S., Wong, D., Forrest, K., Allen, A., Chao, S., Huang, B. E., et al. (2014). Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 12, 787–796. doi: 10.1111/pbi.12183
Watanabe, N., Akond, A. S. M. G. M., Nachit, M. M. (2006). Genetic mapping of the gene affecting polyphenol oxidase activity in tetraploid durum wheat. J. Appl. Genet. 47, 201–205. doi: 10.1007/BF03194624
Watanabe, N., Takeuchi, A., Nakayama, A. (2004). Inheritance and chromosomal location of the homoeologous genes affecting phenol colour reaction of kernels in durum wheat. Euphytica 139, 87–93. doi: 10.1007/s10681-004-2255-7
Wei, J., Geng, H., Zhang, Y., Liu, J., Wen, W., Zhang, Y., et al. (2015). Mapping quantitative trait loci for peroxidase activity and developing gene-specific markers for TaPod-A1 on wheat chromosome 3AL. Theor. Appl. Genet. 128, 2067–2076. doi: 10.1007/s00122-015-2567-0
Young, A., Lowe, G. (2018). Carotenoids-Antioxidant properties. Antioxidants 7, 28. doi: doi 10.3390/antiox7020028
Zeng, J., Wang, X., Miao, Y., Wang, C., Zang, M., Chen, X., et al. (2015). Metabolic engineering of wheat provitamin A by simultaneously overexpressing CrtB and silencing carotenoid hydroxylase (TaHYD). J. Agric. Food Chem. 63, 9083–9092. doi: 10.1021/acs.jafc.5b04279
Zhang, B., Liu, J., Cheng, L., Zhang, Y., Hou, S., Sun, Z., et al. (2019). Carotenoid composition and expression of biosynthetic genes in yellow and white foxtail millet [Setaria italica (L.) Beauv]. J. Cereal Sci. 85, 84–90. doi: 10.1016/j.jcs.2018.11.005
Zhang, F., Chen, F., Wu, P., Zhang, N., Cui, D. (2015). Molecular characterization of lipoxygenase genes on chromosome 4BS in Chinese bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 128, 1467–1479. doi: 10.1007/s00122-015-2518-9
Zhang, W., Chao, S., Manthey, F., Chicaiza, O., Brevis, J. C., Echenique, V., et al. (2008). QTL analysis of pasta quality using a composite microsatellite and SNP map of durum wheat. Theor. Appl. Genet. 117, 1361–1377. doi: 10.1007/s00122-008-0869-1
Zhang, W., Dubcovsky, J. (2008). Association between allelic variation at the Phytoene synthase 1 gene and yellow pigment content in the wheat grain. Theor. Appl. Genet. 116, 635–645. doi: 10.1007/s00122-007-0697-8
Keywords: durum wheat, grain yellow pigment content, carotenoids, yellow index, marker-assisted selection
Citation: Colasuonno P, Marcotuli I, Blanco A, Maccaferri M, Condorelli GE, Tuberosa R, Parada R, de Camargo AC, Schwember AR and Gadaleta A (2019) Carotenoid Pigment Content in Durum Wheat (Triticum turgidum L. var durum): An Overview of Quantitative Trait Loci and Candidate Genes. Front. Plant Sci. 10:1347. doi: 10.3389/fpls.2019.01347
Received: 12 June 2019; Accepted: 27 September 2019;
Published: 07 November 2019.
Edited by:
Jose Luis Gonzalez Hernandez, South Dakota State University, United StatesReviewed by:
Amidou N’Diaye, University of Saskatchewan, CanadaSergio G. Atienza, Spanish National Research Council (CSIC), Spain
Ajay Kumar, North Dakota State University, United States
Copyright © 2019 Colasuonno, Marcotuli, Blanco, Maccaferri, Condorelli, Tuberosa, Parada, de Camargo, Schwember and Gadaleta. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrés R. Schwember, YXNjaHdlbWJlckB1Yy5jbA==
†These authors have contributed equally to this work