 Tobias Andermann
Tobias Andermann Alexandre Antonelli
Alexandre Antonelli Russell L. Barrett7,8
Russell L. Barrett7,8 Daniele Silvestro
Daniele Silvestro- 1Department of Biological and Environmental Sciences, University of Gothenburg, Gothenburg, Sweden
- 2Gothenburg Global Biodiversity Centre, University of Gothenburg, Gothenburg, Sweden
- 3Department of Biology, University of Fribourg, Fribourg, Switzerland
- 4Swiss Institute of Bioinformatics, Fribourg, Switzerland
- 5Department of Plant Sciences, University of Oxford, United Kingdom
- 6Royal Botanic Gardens, Kew, Richmond, United Kingdom
- 7Royal Botanic Gardens, Sydney, NSW, Australia
- 8School of Biological Sciences, The University of Western Australia, Crawley, WA, Australia
The reliable mapping of species richness is a crucial step for the identification of areas of high conservation priority, alongside other value and threat considerations. This is commonly done by overlapping range maps of individual species, which requires dense availability of occurrence data or relies on assumptions about the presence of species in unsampled areas deemed suitable by environmental niche models. Here, we present a deep learning approach that directly estimates species richness, skipping the step of estimating individual species ranges. We train a neural network model based on species lists from inventory plots, which provide ground truth data for supervised machine learning. The model learns to predict species richness based on spatially associated variables, including climatic and geographic predictors, as well as counts of available species records from online databases. We assess the empirical utility of our approach by producing independently verifiable maps of alpha, beta, and gamma plant diversity at high spatial resolutions for Australia, a continent with highly heterogeneous diversity patterns. Our deep learning framework provides a powerful and flexible new approach for estimating biodiversity patterns, constituting a step forward toward automated biodiversity assessments.
Introduction
Since the very beginning of biogeographic research, the estimation and extrapolation of species diversity has been of foremost interest (von Humboldt, 1817; Arrhenius, 1921). It is well established that species diversity is distributed unevenly across space, generally following a latitudinal gradient, with increasing diversity from the poles toward the equator (MacArthur, 1965). On a regional level, it has been found that there are substantial differences in species richness among habitats, such as between a forested area and an open grassland (MacArthur, 1965). These observed spatial patterns have led to the formulation of three levels of species diversity: alpha, beta, and gamma diversity (Whittaker, 1960).
Alpha diversity refers to diversity on a local scale, describing the species diversity (richness) within a functional community. For example, alpha diversity describes the observed species diversity within a defined plot or within a defined ecological unit, such as a pond, a field, or a patch of forest. The scale of such ecological units depends on the organism group of interest; while for birds a defined forest or grassland transect of several hundred m2 to several km2 may be appropriate to describe a species community, for insects this could be a single tree. For plants, alpha diversity is often equated to the count of species identified during the inventory of a vegetation plot of defined size (Revermann et al., 2016).
Beta diversity, on the other hand, describes the amount of differentiation between species communities. Unlike the other levels of species diversity, the exact interpretation and quantification of beta diversity varies substantially across studies (see Tuomisto, 2010a,b for a detailed review on this topic). Originally, beta diversity was defined as the ratio between gamma and alpha diversity (, sensu Whittaker, 1972). Today, one commonly used measure of beta diversity is the Sørensen dissimilarity index (see section “Materials and Methods” below for more detail), which captures spatial turnover as well as differences in diversity between sites (Koleff et al., 2003).
Gamma diversity describes the overall species diversity across communities within a larger geographic area. It is often summarized across biogeographic or political units, such as ecoregions or countries (Kier et al., 2005; Brummitt et al., 2021). Alternatively, studies commonly summarize gamma diversity within cells of a spatial grid of fixed cell-size (Goldie et al., 2010; Thornhill et al., 2016). While alpha diversity represents the actual species diversity that can be measured at a given site, gamma diversity more broadly and loosely describes the diversity of species that can be found in the whole area. Gamma diversity is the most communicated level of species diversity when referring to biodiversity hotspots, with tropical regions, in particular the Neotropics, showing the globally highest gamma diversity values (Raven et al., 2020). Alpha diversity, on the other hand, shows different areas of maximum diversity, dependent on the size of the area surveyed, with temperate grasslands showing among the highest species richness on small plots (Wilson et al., 2012).
While species diversity can be directly counted for small plot sizes, for example, during species inventories (alpha diversity), this requires much effort and thus cannot be scaled up to large areas or whole continents (gamma diversity). Therefore, many studies apply some form of modeling and estimation to derive diversity maps for larger areas. For example, gamma diversity is often inferred by modeling individual species distributions and adding these up to obtain the total number of species that occur in a given area (Mutke and Barthlott, 2005; Barthlott et al., 2007). However, this approach has been shown to introduce substantial errors, when cross-checking the diversity predictions with actual species counts in selected grid cells (Aranda and Lobo, 2011). A general shortcoming of these methods is that usually the data available is insufficient to reliably model the ranges for each individual species. This problem intensifies with the number of species in the target group for which to estimate diversity patterns. In some cases, total species diversity is extrapolated for larger groups, based on a selected subset of taxa with good data coverage, under the simplistic assumption that the diversity patterns revealed by these taxa are representative for others (Kier et al., 2005), which is however often not the case (Ritter et al., 2019).
Alternative approaches have been applied to the task of diversity estimation and mapping, which skip the step of modeling individual species ranges. These often involve using occurrence records, floras, and checklists to count the total number of species that has been recorded within large biogeographic regions (Mutke and Barthlott, 2005; Kreft and Jetz, 2007). While such approaches do not require modeling distributions of individual species, they are particularly vulnerable to biases in data collection, as some taxa may be better represented in some checklists and biodiversity repositories than others. This method assumes one single diversity value within each of the regions analyzed, without accounting for diversity differences within these (sometimes large) areas. Although it is possible to interpolate diversity values to a finer resolution using spatial autocorrelation of associated variables such as climatic predictors (Kreft and Jetz, 2007), such gap filling may be difficult to verify and often provides a false sense of confidence for data-poor regions.
With the emergence of continental and global vegetation plot databases (Chytrý et al., 2016; Bruelheide et al., 2019; Sabatini et al., 2021), a new data source with extended spatial coverage has become widely, providing point-estimates of species diversity within clearly delimited areas. Recently, Večeřa et al. (2019) showed the potential of machine learning methods (random forest models) to estimate the expected diversity for fixed size vegetation plots (alpha diversity), based on climatic and other predictors, when trained on alpha diversity data from vegetation plot databases. However, to our knowledge, available machine learning models cannot extrapolate vegetation plot data to larger areas and do not provide estimates of multiple metrics of biodiversity.
Here, we present a deep learning framework that uses neural network models to predict alpha, beta, and gamma diversity. The models are trained to predict plant diversity based on climatic and geographic predictors, measures of human impact, and sampling effort. Our approach requires neither specific distribution information about individual species, nor the manual extrapolation of species richness using methods such as species–area curves (Kier et al., 2005). Instead, our models inherently learn the species–area relationships, allowing prediction of the three diversity metrics at user-defined spatial scales. Our approach is purely data-driven and hypothesis-free, including the selection of the best neural network architecture, to avoid confirmation biases in terms of picking models whose diversity predictions best match previous expectations.
We selected plot-based vegetation survey data from Australia (vascular plants; Tracheophyta) to empirically test the effectiveness of our models in predicting diversity patterns and to validate our methodology. Australia, as an island continent, has the advantage of a clear delimitation of natural boundaries; it has high natural diversity and uneven biological sampling (González-Orozco et al., 2014; Cook et al., 2015; Laffan et al., 2016); high spatial heterogeneity with well-defined and contrasting biomes (Byrne et al., 2008, 2011; Macintyre and Mucina, 2021); a relatively well-documented vascular flora with reliable national databases (Sparrow et al., 2021)1 that feed into the Global Biodiversity Information Facility (GBIF)2; good climatic data3; and a large number of freely available plot-based vegetation records suitable for training deep learning frameworks (Sabatini et al., 2021).
Materials and Methods
Vegetation Plot Data
The values of alpha, beta, and gamma diversity used in this study to train the neural network models were derived from vegetation plot data (species inventories). We downloaded these data from the sPlotOpen database (Sabatini et al., 2021), only using plots where all vascular plants had been assessed. This resulted in a total of 7,896 vegetation plots for Australia (Figure 1). For each vegetation plot, we compiled its area (which ranged from 50 to 10,000 m2) and the list of plant species identified in the plot (ranging from 1 to 115 species). From each of these sites, we compiled measures for alpha, beta, and gamma diversity as described in more detail below (Figure 1), which we used to train our models.
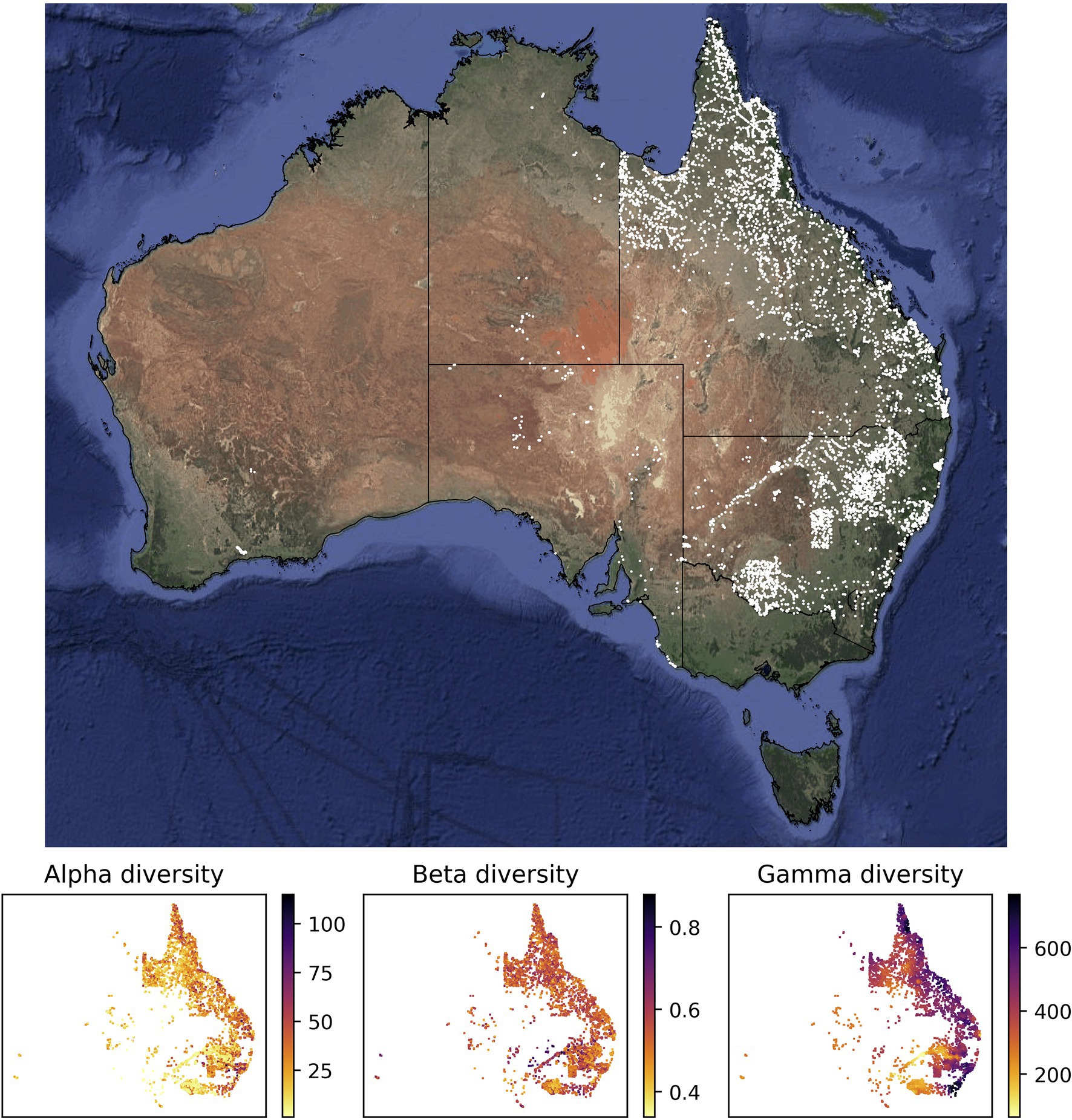
Figure 1. Sites with vegetation plot data used in this study for model training and evaluation. Most of the vegetation plot sites used in this study (white points, 7,896 sites) are located in the easternmost two Australian states Queensland (northeast) and New South Wales (center east). The panels below the map show the compiled measures of alpha, beta, and gamma diversity for all vegetation plot sites. The satellite image of Australia was downloaded via ggmap (Kahle and Wickham, 2013). The spatial scale of the alpha diversity estimates is defined by the plot size of the underlying vegetation plots and differs among sites. Similarly, the gamma diversity values are based on sets of 50 neighboring vegetation plots; depending on the spatial density of vegetation plots, these diversity values are therefore determined across different spatial scales. Both values, the size of the vegetation plot and the spatial scale of each gamma diversity estimate, are used as features in our models.
Calculating gamma diversity required the definition of a surrounding area, preferably containing other vegetation plots, to determine the overall diversity found within the cumulative species lists of several neighboring vegetation plots (Figure 2). To ensure that the same number of vegetation plots was used for calculating the gamma diversity of each site, we defined as the surrounding area a circle around each site encompassing exactly N nearest neighbors (vegetation plots). The gamma diversity for each site was then determined as the number of unique species names extracted from the species lists of the N nearest neighbors within the encompassing circle. After compiling diversity estimated for different values of N (Supplementary Figures S1–S7), we chose an N of 50 for all models in this study, as this value led to the best compromise between a visually discernible spatial structure in the resulting beta and gamma diversity values, while also highlighting regional heterogeneity (Supplementary Figure S3).
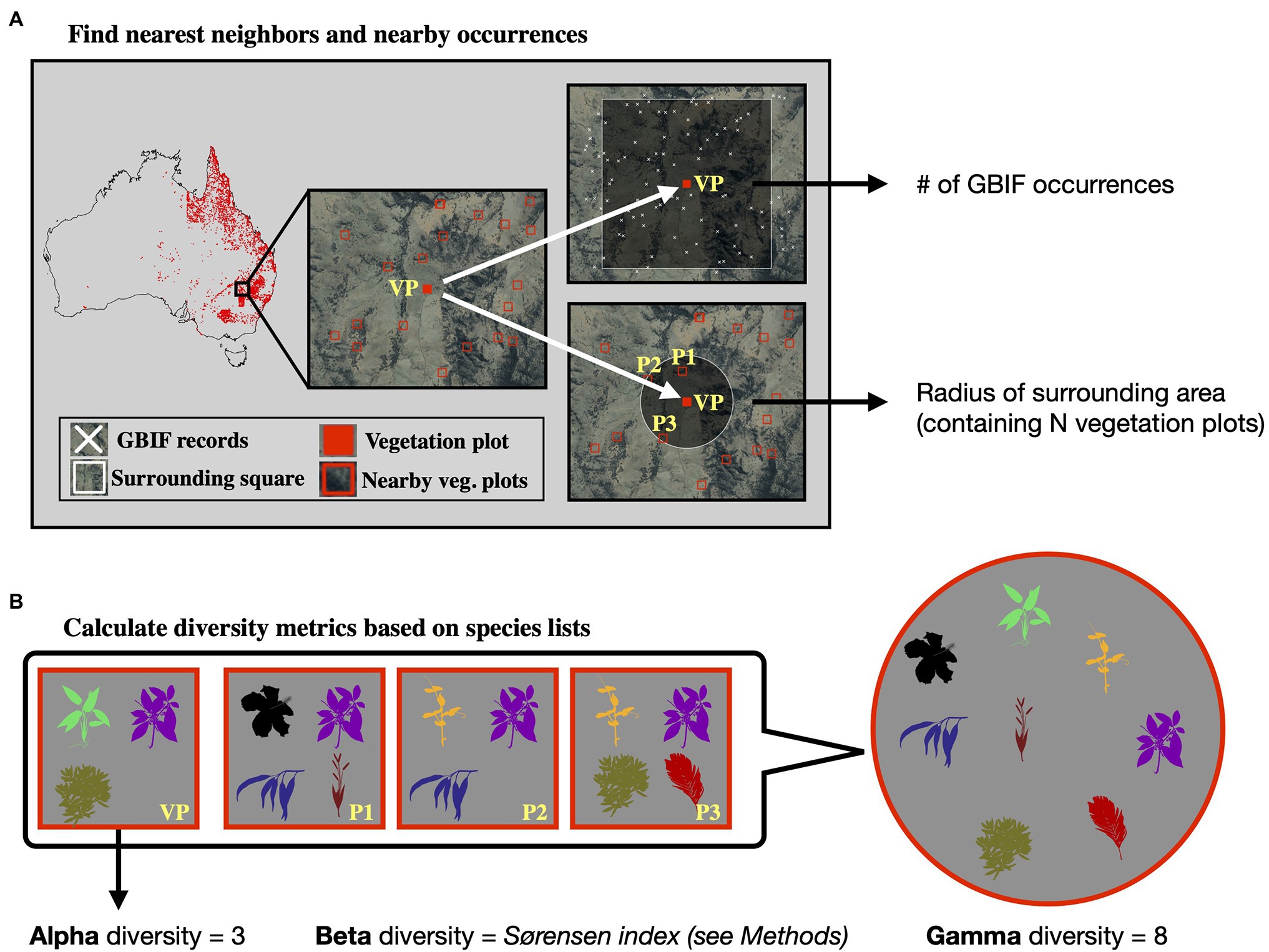
Figure 2. Calculation of diversity measures from vegetation plot data. For a given vegetation plot (VP, solid red square, panel A) we identified the N nearest neighboring vegetation plots in space (N = 3 in this example, represented by plots P1–P3). We exported the radius of the smallest circle encompassing all N neighbors as a feature for model training. Additionally, we exported the number of GBIF occurrences within a square of 10 × 10 km size around the given vegetation plot, as a measure of sampling effort in the general area. Having identified the nearest neighbors (P1–P3), we compared the species lists of these vegetation plots with the focal vegetation plot (VP, panel B). Alpha diversity represents the number of species found in the focal vegetation plot (VP), while gamma diversity represents the total diversity consisting of all species identified among the focal and neighboring vegetation plots. Beta diversity was calculated using the multiple-site Sørensen dissimilarity index (see section “Materials and Methods”), based on the differences in species composition found among the selected vegetation plots.
The radius of this encompassing circle varied between sites, depending on the proximity of other vegetation plots relative to the given site. The extent of the radius itself was used as a feature in our models, allowing the neural network to learn the expected associations between gamma diversity and the size of the area for which it was calculated (the species-area relationship), which we used later when making predictions with this model to adjust the spatial resolution of the predictions.
Finally, beta diversity was calculated using the multiple-site implementation of the Sørensen dissimilarity index (), following the definition in (Baselga, 2010). For a given focal site j with N neighbors, we defined the focal site index as j = N + 1. We iterated through the N neighboring sites (i) and applied the formula:
with
where and are the number of species only present in site i and site j, respectively, is the total number of species in site i (alpha diversity from vegetation plot), and is the total number of species in all sites combined (gamma diversity).
Feature Generation
The alpha, beta, and gamma diversity metrics described above were used as labels to train three models, one for each diversity metric. The predictors (features) used in these models were compiled from different publicly available data sources. To ensure approximately equal size of all grid cells for the raster-based feature data used in this study, we transformed all spatial data into the cylindrical equal-area (CEA) projection, centered at 30° latitude south of the equator.
As a general measure of sampling effort, we compiled the number of recorded species occurrences, available on GBIF, which were found in the vicinity of a given site. We first downloaded all non-fossil vascular plant (Tracheophyta) occurrences for Australia from GBIF that were based on human observations and were not flagged for geospatial issues.4 This includes both native and naturalized species, the latter having uneven spatial distributions related to broad disturbance histories in Australia (Leishman et al., 2017). This resulted in 13,580,191 occurrence records. We then discarded any records with non-binomial species names and cross-checked names of the remaining records against the World Checklist of Vascular Plants, a continuously updated collection of reviewed plant species names (Govaerts et al., 2021). This resulted in 12,622,786 remaining GBIF records. For each site, we defined a 10 × 10 km window centered on the site’s coordinates; we then counted all GBIF occurrences within this window as a measure of sampling effort (Supplementary Figure S8), as well as the number of species found in the GBIF records as a diversity proxy. Both counts were used as individual features in our models.
We also compiled climatic and anthropogenic features for each site. First, we downloaded raster data for 19 bioclimatic variables (BIO1–BIO19) as well as data on elevation from the WorldClim database (Fick and Hijmans, 2017).5 Second, we downloaded raster data on human footprint from wcshumanfootprint.org (Venter et al., 2016), which reflects the magnitude of human disturbance, including information on human population density, agricultural land use, presence of roads and several other data sources. There is a high correlation between population density, agricultural development, and high biodiversity regions in Australia (Keith and Auld, 2017). All data rasters were downloaded at a resolution of 0.5 min of a degree (~1 × 1 km grid). The complete list of features (n = 27) extracted for each site is shown in Table 1. All feature values were rescaled to range between 0 and 1 before being used as input in the neural network.
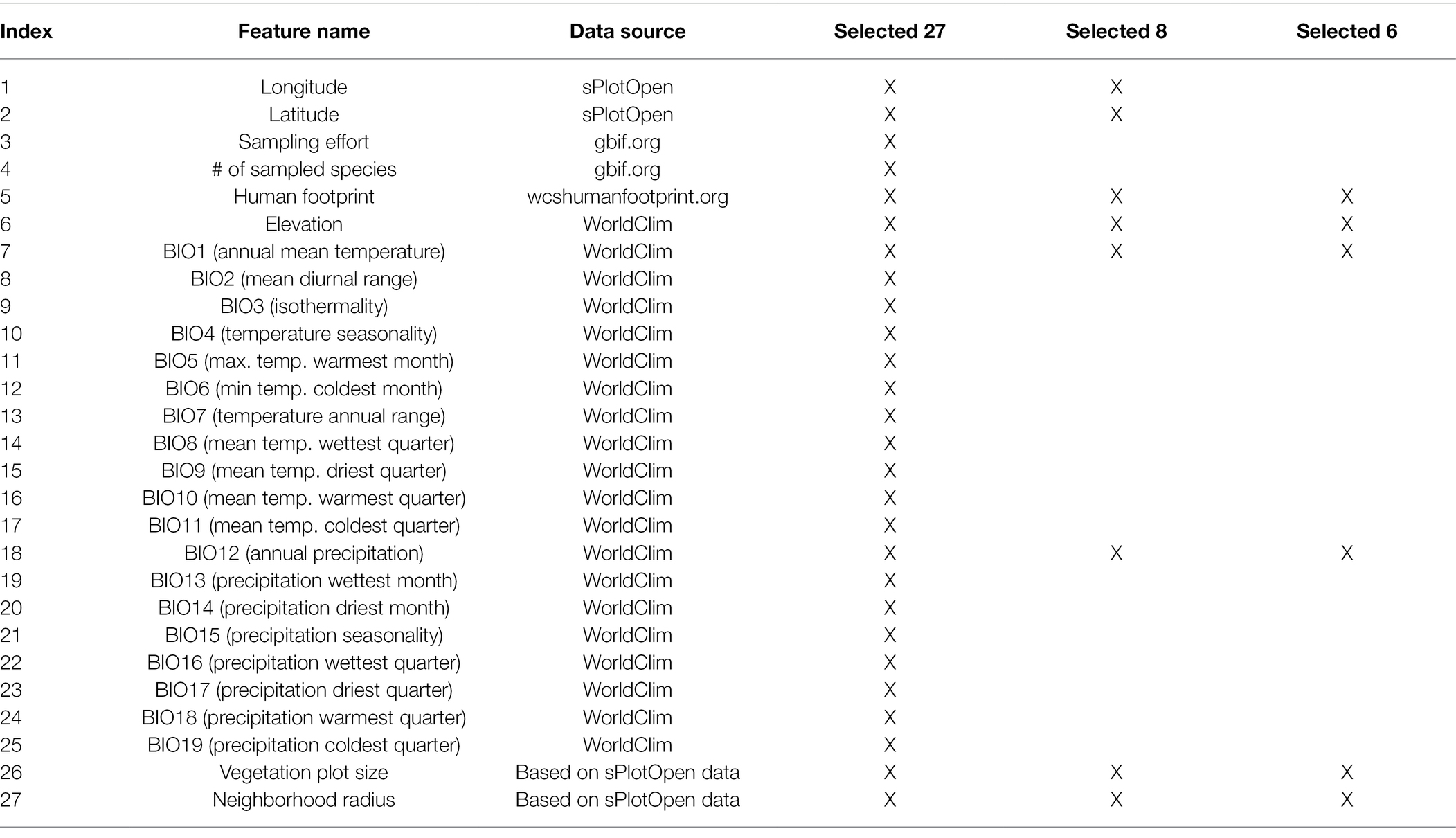
Table 1. Features used in the neural network models.
Neural Network Architecture
We built regression models using fully connected neural networks to learn and then infer species diversity based on the climatic, geographic, human footprint features, as well as general sampling effort reflected by the available GBIF data. While the output values in a neural network regression model can theoretically take any range, rescaling these values to a smaller range generally improves the model convergence and performance. We therefore rescaled our training labels by multiplying the diversity values by the following scaling factors, which were approximated to match the maximum values found in the training data for each diversity metric, thus leading for all values to fall within a range between 0 and 1: alpha scaling factor = 1/100, gamma scaling factor = 1/800 (no rescaling was necessary for beta diversity).
Models differed in the number of hidden layers and number of nodes per layer (see model testing below, Table 2). Further, we applied different fractions of dropout in our models, which leads to randomly removing the specified fraction of nodes in each hidden layer in each training epoch. This has the effect of reducing overfitting toward the training data, as the model is forced to rely less on individual highly optimized weights. We used the rectified linear units function (ReLU) as the activation function within each layer, and a softplus activation function for the output layer. The softplus activation function in the output layer ensures that the output values (diversity estimates) are all within a positive range, while not imposing any restrictions on the possible maximum value.
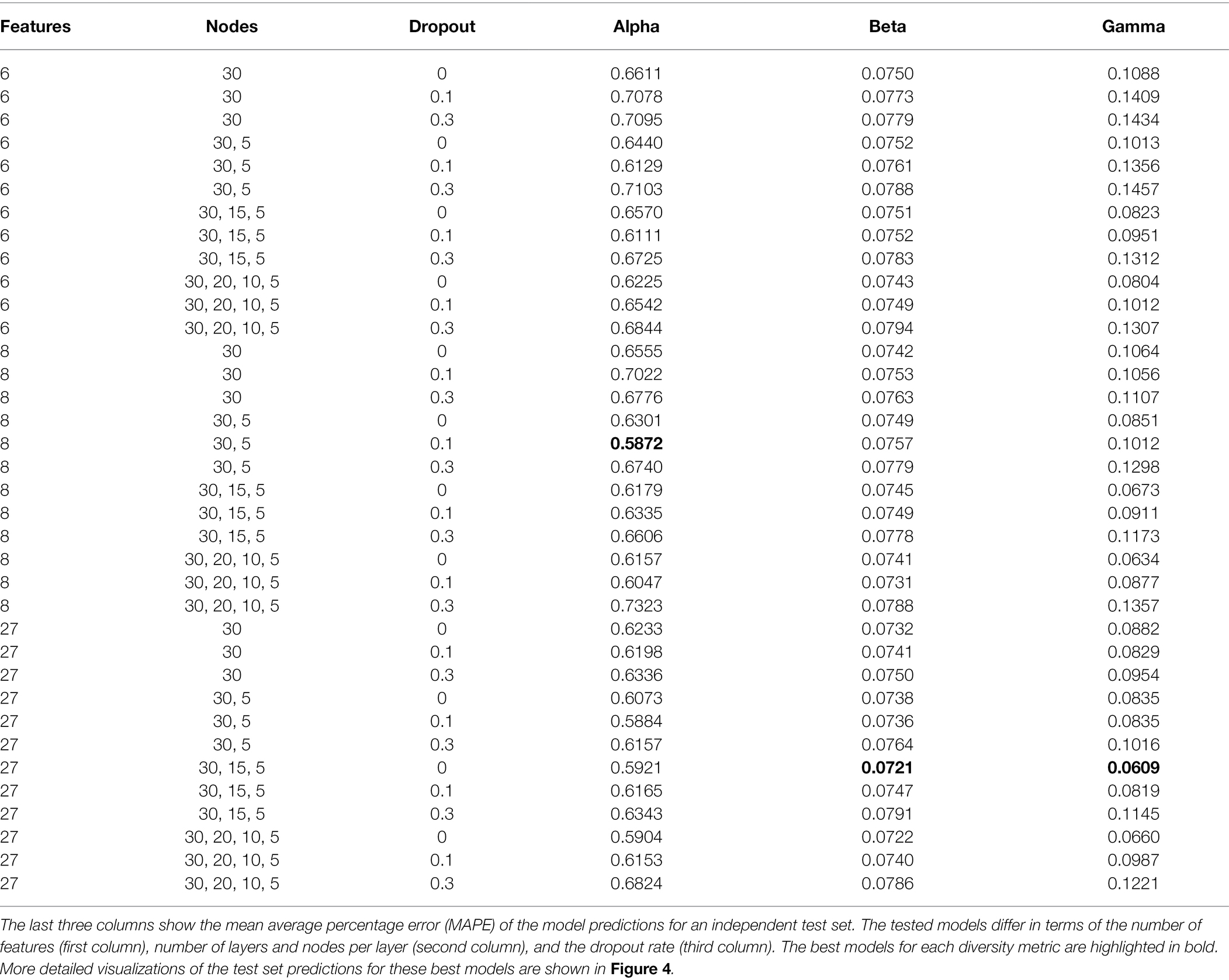
Table 2. Prediction accuracy for test set of all tested models.
For training, we used the mean absolute error (MAE) as the loss function to be minimized. Of the 7,896 training instances (vegetation plot sites), we set aside 20% (1,579 instances) as an independent test set. We assigned another 20% (1,579 instances) of the data as a validation set, which we used to determine the optimal number of training epochs that minimizes the validation set MAE, while preventing overfitting toward the training data. All models were trained with the remaining 60% of the data (4,738 instances), using a batch size of 40 instances.
Model Testing and Evaluation
We tested a range of different training configurations for each diversity metric, specifically testing different combinations of input features, different numbers of hidden layers and nodes per layer, and different dropout fractions (Table 2). Based on the diversity predictions for our independent test set, we calculated the mean absolute percentage error (MAPE) for each model, which differs from the MAE in being a relative error, scaled by the absolute values of the predictions. For each diversity metric we determined the best model configuration by picking the model with the lowest MAPE score.
After identifying the most suitable settings through model testing, we retrained this best model for each diversity metric, using all 7,896 training instances. To avoid overfitting towards the training data, we trained these production models only until the optimal epoch determined during model testing. For each diversity metric we trained an ensemble of 50 models with different random starting seeds, using the best model settings. We averaged the predictions across all these 50 models for each diversity metric, and also calculated the coefficient of variation (the standard deviation divided by the mean) as a measure of variation of the predicted diversity values, representing uncertainty.
Prediction Data
To produce the predictions of alpha, beta, and gamma diversity across Australia, we defined a grid with a cell size of 10 × 10 km and extracted the 27 features for each of the cell centroids. We set the plotsize feature for all points to 500 m2 (the most common vegetation plot size in training data, Supplementary Figure S9). Therefore, the predicted alpha diversity values reflect the expected number of plant species to be found in a plot of size 500 m2. The radius feature, describing the size of the surrounding area around a point for which gamma diversity is estimated, was set to 5 km, to approximately match the size of the grid cells (10 km× 10 km square).
By adjusting the radius feature, our trained models can be used to predict beta and gamma diversity at user-defined spatial resolutions, as it can be adapted to match the given cell size. Similarly, adjusting the plot size feature allows us to predict alpha diversity for any given plot size. This enables flexibility in predicting species diversity at different spatial resolutions of the prediction grid, while inherently accounting for species-area relationships, as these are learned by the model. For both the radius feature and the plot size feature, the selected values for prediction should be chosen to be within the range of values present in the training data (Supplementary Figure S9).
Results
An overview of all tested models is shown in Table 2. For alpha diversity, we identified as the best model the following configuration: eight features (see Table 1), two layers with 30, and five nodes, and a dropout rate of 0.1. For beta and gamma diversity, the following configuration was identified as the best model: all 27 features, three layers with 30, 15, and 5 nodes, respectively, and no dropout (dropout rate = 0). We identified the following training epochs as the stopping points for model training, as they constituted the best compromise between minimizing the training loss while avoiding overfitting (rounded to the nearest 50): 1500 epochs (alpha), 750 epochs (beta), and 1700 epochs (gamma, see Supplementary Figure S10). We used these numbers of training epochs to train the ensemble of 50 productions models for each diversity metric.
Alpha Diversity Predictions
The best alpha diversity model predicted the test set, consisting of approximately 1,600 vegetation plots, with a mean absolute percentage error (MAPE) of 58.72% (Figure 3). This means that the predicted diversity for the average test set instance was within an approximately 60% range of the true diversity value. This comparably high prediction error is likely caused by the fact that the alpha diversity training instances show a complex spatial pattern, with no easily discernible areas of high or low diversity (Figure 1). The fact that most of the training features are spatially autocorrelated (such as the BioClim climatic layers) makes it difficult for the model to infer a strong signal from these features during training for predicting alpha diversity. The predictions made by an ensemble of 50 trained alpha models show comparably large uncertainties in some areas (Figure 4), with a median coefficient of variation across all cells of 0.30. The areas of highest uncertainty—exceeding the median value—are located mostly in the western half of Australia (grey areas in Figure 4), presumably due to the limited training data from those regions (Figure 1).
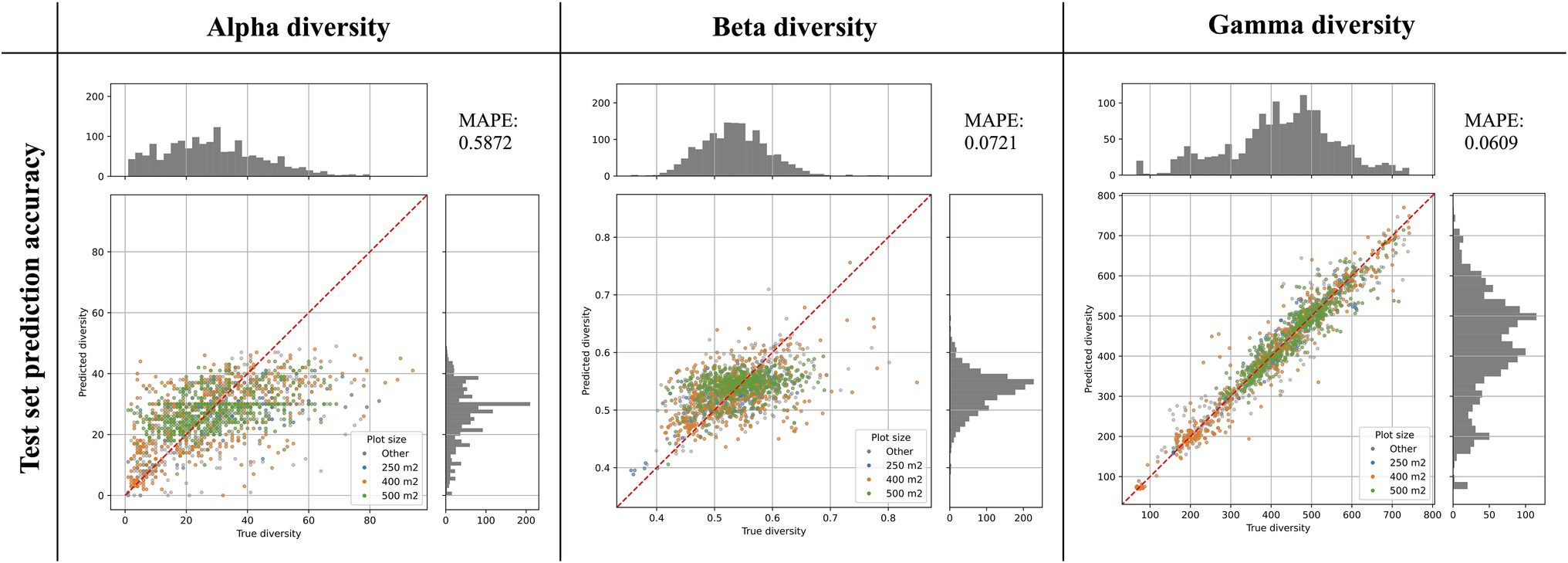
Figure 3. Prediction accuracy of best models as determined on an independent test set. The scatter plots show the predicted diversity (y-axes) plotted against the true diversity (x-axes) for the best alpha, beta, and gamma diversity models. These estimates were made for a randomly selected and independent test set (N = 1,579 instances), exclusively consisting of instances that were not used during model training. The points are colored by the vegetation plot-size associated with each data point (see legend). The red diagonal line shows for reference the best-case scenario, if all labels were predicted 100% accurately. Histograms show the total distribution of values for the true diversity values (top) and the predicted diversity values (right). For each model we calculated the Mean Absolute Percentage Error (MAPE), shown in the top-right corner of each plot.
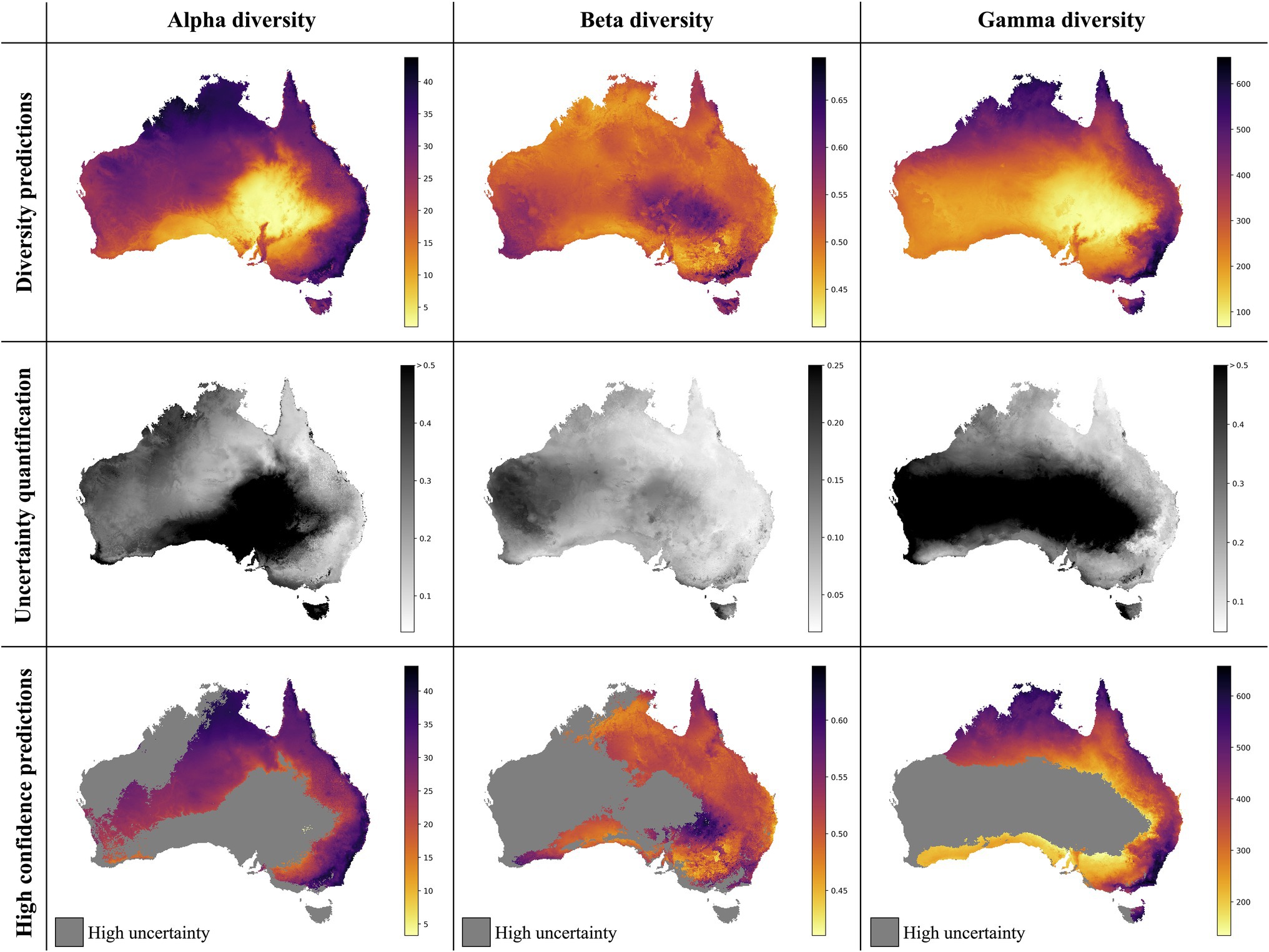
Figure 4. Neural Network predictions for alpha, beta, and gamma diversity of vascular plants. The neural network models were trained separately on alpha, beta, or gamma diversity estimates, which we compiled from available vegetation plot data (Figure 1). The alpha diversity maps (left column) show the number of vascular plant species expected to be found in a 500-m2 plot (most common plot-size found in the vegetation plot data; Supplementary Figure S2). The beta diversity maps (center column) quantifies the spatial turnover and differences in species compositions (Sørensen dissimilarity index, relative to the total diversity) between such 500 m2 plots within each grid cell (10 × 10 km). The gamma diversity maps (right columns) show the total species richness within each grid cell. The top row shows the predictions averaged across an ensemble of 50 independently trained models, using different starting seeds. The center row shows the coefficient of variation for each grid cell, as a measure of prediction uncertainty. High values (dark grey/black) correspond to grid cells with less consistent diversity predictions. The bottom row shows the average diversity predictions for only those grid cells with the most consistent diversity predictions (coefficient of variation smaller than median across all grid cells), while high-uncertainty grid cells are marked in grey.
The overall highest alpha diversity predictions are found along the eastern coast of Australia, from the northernmost tip of Queensland to the most southwestern part of Victoria (Figure 4). A potential drop in alpha diversity is visible in the area around Cairns, extending about 100 km south from the city area, perhaps corresponding with the Burdekin-Lynd gap, an area that has been shown to constitute a range gap for several species (Edwards et al., 2017). However, these grid cells are predicted with comparably high uncertainty, giving only weak support for this observed pattern. Other areas of medium to high alpha diversity inferred by our model are the top end of the Northern Territory, as well as the north Kimberley in northern Western Australia.
Beta Diversity Predictions
The best beta diversity model resulted in a MAPE of 7.21%, thus yielding a substantially higher accuracy compared to the alpha diversity model. Similarly, the median coefficient of variation across all prediction grid cells was low with (0.09), indicating high consistency in the predicted diversity pattern. The high-uncertainty cells, identified as having a coefficient of variation above the median, largely overlap with those identified for the alpha diversity model, covering the majority of Western Australia (Figure 4). Perhaps being the least intuitive of the three diversity metrics, areas with a high predicted beta diversity within our framework represent sites that are expected to show large differences in species composition between vegetation plots within the defined area (a given grid cell).
Differently to alpha diversity, the majority of the eastern coastal areas show medium to low beta diversity values. Higher beta diversity is inferred for the southeastern part of Australia, particularly in higher elevations between Canberra and Melbourne. High species turnover is also inferred for the arid eastern desert of central Australia, as well as for south-western Australia.
Gamma Diversity Predictions
With a MAPE score of 6.09%, our gamma model yielded the most accurate predictions among the three diversity metrics. The median coefficient of variation of gamma predictions across all of Australia was 0.37. As for the other two models, this variation was largely driven by high uncertainty grid cells in the western half of the continent (Figure 4). Our model predictions of gamma diversity across Australia identify several vascular plant biodiversity hotspots, such as the tropical and subtropical forests in northeastern Queensland, the tropical and subtropical grasslands across northern Australia, as well as the temperate forests and the montane grasslands and shrublands of south-eastern Australia (Figure 5). Below we discuss the specific spatial diversity patterns that were predicted by our models in more detail (see section “Discussion”).
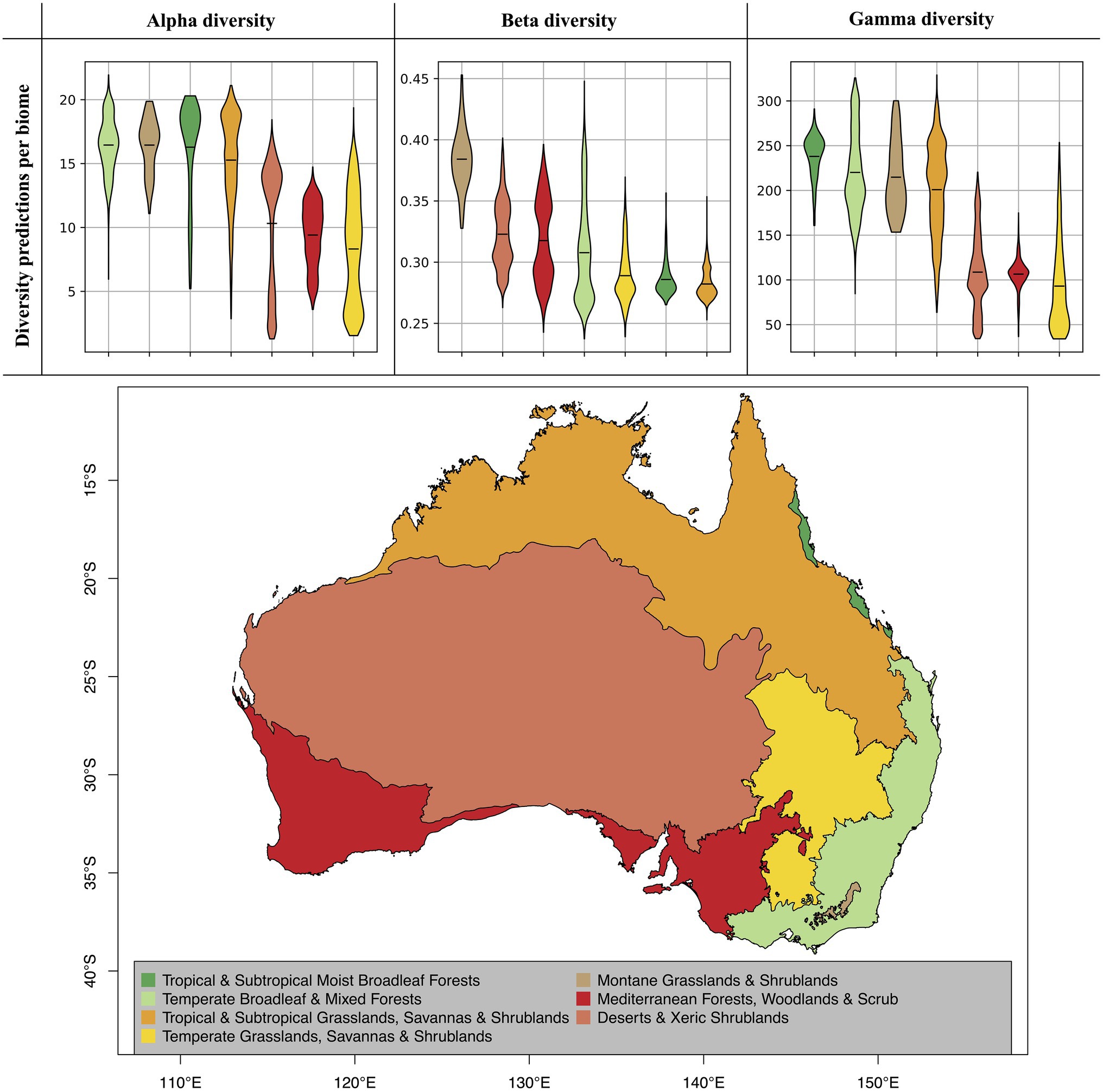
Figure 5. Diversity predictions by biome. The violine plots show the range of diversity predictions across all grid cells within a given biome, excluding high uncertainty predictions (see Figure 4). The horizontal black lines inside the violine plots mark the mean estimate for each biome. The biomes, which are displayed on the map, were compiled from the Terrestrial Ecoregions of the World (TEOW) data (Olson et al., 2001).
When evaluating our model predictions on a per-biome basis, excluding high uncertainty predictions as identified in Figure 4, we identify differences in predicted diversity between biome types (Figure 5). For alpha and gamma diversity, we find the highest average diversity predictions for tropical forests, temperate forests, montane shrublands and grasslands, and tropical and subtropical grasslands and savannas. Our beta diversity estimates, on the other hand, show a rather uniform pattern across biomes, with the exception of montane grasslands and shrublands, which show the highest species turnover. The high beta diversity identified for the montane biome may be driven by the increased elevational gradients in this area, as species turnover has been found to be higher along elevational gradients (Venn et al., 2017; Albrecht et al., 2021).
Discussion
Using Neural Networks for Diversity Predictions
Here we developed and applied a novel approach of estimating species diversity, using neural networks. We showcased our model, using vegetation plot data that are openly available through the sPlotOpen database for Australia, and showed that it can be used to accurately predict diversity on different scales (alpha, beta, and gamma). This enables us to produce maps of species diversity at a wide range of spatial resolutions. The main advantages of our approach, as compared to previous approaches of modeling species diversity, are that (i) it does not require the modeling of distribution ranges for individual species (e.g., Mutke and Barthlott, 2005; Barthlott et al., 2007), (ii) it does not require an a priori definition of species-area relationships (e.g., Kier et al., 2005), (iii) it does not require the assumption of monotonic and usually oversimplifying relationships (e.g., linear or exponential) between predictors and response variable (e.g., Cingolani et al., 2010), and (iv) it allows the direct quantification of uncertainty in the predictions.
Given these advantages, and the easy combination of different features (predictors) of continuous or categorical nature, our deep learning model, represents a promising new tool for the task of predicting diversity. This study and other recent work (e.g., Večeřa et al., 2019) demonstrate how such models can be trained on readily available data from public databases. Further, the versatility in terms of data input into these models allows for new ways of accounting for the effects of sampling effort (GBIF sampling density feature) and of human disturbances (human footprint feature) on diversity estimates. The advantage of these features, as well as the additional climatic features used in our models, is that these data are available on a global, spatially detailed scale (<1 km2). Previous studies have shown the utility of these data for modeling biotic properties of the landscape, such as phylogenetic diversity (Park et al., 2020). Predictors with even higher spatial resolution, such as remote sensing data (e.g., satellite images or 3D point clouds from airborne laser scanning), could help to improve the accuracy of the models presented in this study even further.
Remote sensing data are a promising and potentially highly informative data source for the task of biodiversity estimation (Gholizadeh et al., 2020; Moat et al., 2021). These data have been successfully applied in several recent studies for modeling vegetation attributes such as biomass (Breidenbach et al., 2021), growing stock volume (Lindgren et al., 2021), and plant size (Söderberg et al., 2021), and can be applied for global inventory of habitats and for estimating the trait diversity within these habitats (Cavender-Bares et al., 2020). These data sources, which are already successfully applied for many biodiversity-related purposes (see overview in Cavender-Bares et al., 2020), will likely play a key role for future developments in the field of automated biodiversity assessments, and can be readily added as additional features to neural network models as the ones presented in this study.
The neural network models trained in this study do not allow investigating direct causal relationships between predictors (features) and the response variable (species diversity). As neural networks are very complex models with many parameters, direct relationships cannot be inferred in the same way as with classic mechanistic models (e.g., linear regression models). However, different methods have been developed to increase the interpretability of neural networks (Lundberg and Lee, 2017), which can for example be used to investigate the importance of individual predictors on the overall test accuracy (permutation feature importance, sensu Breiman, 2001).
The gamma diversity predictions reached the overall highest accuracy (Figure 3), likely because they carry the strongest spatial signal and can therefore be predicted more easily with the used features, many of which are themselves spatially autocorrelated. This spatial signal in the gamma diversity values is noticeable when evaluating the training data (Figure 1), where there are spatially coherent areas of overall low and overall high gamma diversity, whereas for alpha and beta diversity the spatial patterns in the data are more disjunct. The reason that alpha and beta diversity cannot be estimated with equally good accuracy is that the model is unable to learn the small spatial differences in alpha or beta diversity shown even among neighboring points (Figure 1) based on the available features.
Here, we focus on the taxonomic aspect of diversity, i.e., species richness. Besides taxonomic diversity, there are other types of biodiversity, such as phylogenetic diversity and functional diversity (e.g., Swenson, 2011). The approach presented in this study can theoretically be applied equally to these other types of biodiversity. However, this would require additional information on the identified species in each vegetation plot. In the case of modeling phylogenetic diversity, information on the phylogenetic relationships and distances of all identified species would be required. In the case of functional diversity, information about species’ ecology and functional traits would be needed, which can be compiled for many species from large databases, such as the TRY database (Kattge et al., 2011). Such data could be further complemented by automated machine learning methods for functional trait compilation, for example from digitized herbarium specimen (Davis et al., 2020). Alternatively, similar deep learning models could be designed that rely on training data other than vegetation plots, such as functional diversity estimates informed by sites for which different functional attributes have been compiled (e.g., Bagousse-Pinguet et al., 2019). Particularly for functional diversity, several suitable predictors are readily available that could be used as features in such models, such as soil data (Commonwealth Scientific and Industrial Research Organisation, 2014), hydrological data (Australian Government, 2021), as well as the climate predictors used in our models (Fick and Hijmans, 2017).5
Correlation Between Diversity Metrics
Previous studies have found all three diversity metrics to be correlated (Cingolani et al., 2010). Here, we find that the maps produced for alpha and for gamma diversity overall show similar diversity hotspots, while beta diversity shows a different spatial pattern (Figures 4, 5). There is a wide variety of definitions of beta diversity, some which are directly correlated to alpha and gamma diversity (e.g., Whittaker’s original definition of , sensu Whittaker, 1960). However, the Sørensen dissimilarity index used in this study does not display such a direct correlation to either alpha or gamma diversity, leading to the distinctly different spatial pattern observed in our predictions (Figure 1).
While the patterns of alpha and gamma diversity inferred by our models are strongly correlated, they do differ in some areas. There is potential for areas with low gamma diversity to exhibit relatively high densities of species, leading to high alpha diversity estimates within smaller defined areas, such as the 500 m2 vegetation plots used in our predictions. This is particularly the case for vegetation types consisting of species with relatively small individual plant sizes (such as grasslands and shrublands), which in comparison with forests allow for a potentially denser accumulation of individuals. These differences in average plant size often lead to open habitat grasslands displaying comparatively high alpha diversity values, particularly on small plot sizes (Wilson et al., 2012).
The difference between alpha and gamma diversity is a matter of spatial scale. While alpha diversity describes the number of species in a specific species community (vegetation plot, ~500 m2), gamma diversity describes the number of species in a larger geographic area (grid cell in spatial raster, ~100 km2). In our approach, as in most regression tasks, we expect the predictions of alpha or gamma diversity to be reliable only within the spatial scales that are well represented in the training data, i.e., for alpha diversity within a range between 50 and 10,000 m2, and for gamma diversity between ~100 and 40,000 km2 (Supplementary Figure S9). Since these ranges do not overlap with each other, these are considered to be separate diversity metrics in our model. However, if it were feasible to manually count all species occurring in a vegetation plot the size of a grid cell in our prediction raster, this alpha diversity estimate would be expected to match the predicted gamma diversity of the same, equally sized grid cell.
Biases in Training Data
Sampling biases pose a serious challenge for biodiversity reconstruction in countries of uneven spatial sampling, such as Australia (Piccolo et al., 2020). In our approach, we account for geographic bias in the training data by quantifying the uncertainty in the diversity predictions. Areas of high prediciton accuracy identified by our models, largely reflect those areas with little or no training instances. Additionally, we add the count of GBIF occurrence records in the vicinity of any given training instance as a measure of general sampling effort. Recent studies have addressed the issue of differences in sampling effort in more detail for defined regions and have pointed a way forward in addressing and accounting for this issue, using strategically sampled empirical data (Gioia and Hopper, 2017). However, such efforts are labor- and time-intensive and may not be feasible on continental scales. Alternatively, computational tools that can readily quantify spatial biases based on public database data are a promising way forward towards better accounting for the issue of spatial sampling biases (Zizka et al., 2021).
The ground truth diversity data derived from vegetation plots, which were used in this study for model training, are subject to several potential biases. Previous studies have found an effect of the number of observers conducting the inventory, plot-size, and vegetation type on how reliably and consistently species are being identified, particularly effecting the detection of rare and cryptic species (Vittoz and Guisan, 2007). While we did control for plot-size by adding it as a feature to our models, the potential effects of the number of observers and the vegetation type could not be as easily modeled in this framework. Beyond the issue of data availability, these predictors cannot be added as features to the model, as their values cannot be assumed or compiled for un-surveyed areas for which we want to make predictions with the trained model. However, these biases could be addressed in future applications of these models by apply additional data filtering and bias correction steps, that go beyond the cleaning steps already implemented in the sPlotOpen database (Sabatini et al., 2021).
Predicted Diversity Patterns for Australia
Our model predictions of alpha and gamma diversity identify several vascular plant biodiversity hotspots for Australia, such as (i) the tropical and subtropical forests in northeastern Queensland, (ii) the temperate forests and the montane grasslands and shrublands of southeastern Australia, (iii) the tropical savanna dominated ecosystems of the Northern Territory, and (iv) northern Western Australia (Figures 4, 5). These areas of high vascular plant diversity largely correlate with findings of previous studies, (e.g., Steffen, 2009; Goldie et al., 2010; Yeates et al., 2014; Thornhill et al., 2016) and are highly correlated with broader climatic patterns (Ooi et al., 2017).
One notable difference of our model predictions compared to previous work is the south-west of Western Australia, which is often inferred as a plant diversity hotspot (e.g., Myers et al., 2000; Steffen, 2009), but was predicted with comparably low alpha and gamma diversity by our models. This south-west Australian biodiversity hotspot may not have been predicted accurately—as also indicated by the large prediction uncertainty identified by our model—due to alternate evolutionary patterns in the region that have led to higher diversity than might otherwise be predicted in this very old and climatically buffered, infertile landscape (an OCBIL; see Hopper et al., 2016). It is also interesting to note that the models predict similar alpha diversity between the Kimberley region of Western Australia and the top end of the Northern Territory, as recent surveys demonstrate that this is indeed the case (Barrett and Barrett, unpublished data).
Interestingly, our beta diversity model inferred high species turnover for the arid eastern desert of central Australia. While this region has the lowest estimates for alpha and gamma diversity, the species turnover (relative to the total diversity) is inferred to be among the highest on the continent, likely reflecting a complex mosaic of Mediterranean, temperate, and arid vegetation types in this region (Fox, 2007).
Data Availability Statement
Publicly available datasets were analyzed in this study. These data can be found in the project’s Zenodo repository (https://doi.org/10.5281/zenodo.6394915). All code used in this project can be found on the project’s GitHub repository, v1.0.0 (https://github.com/tobiashofmann88/NN_diversity_prediction/tree/v1.0.0).
Author Contributions
TA, AA, and DS contributed to conception and design of the study. TA compiled the data, wrote the code, ran all analyses, and wrote the first draft of the manuscript with the contributions of sections written by AA, RB, and DS. All authors contributed to the article and approved the submitted version.
Funding
TA and DS acknowledge the funding from the Swedish Research Council (2019-04739). AA acknowledges the financial support from the Swedish Research Council (2019-05191), the Swedish Foundation for Strategic Research (FFL15-0196) and the Royal Botanic Gardens, Kew. DS also received funding from the Swiss National Science Foundation (PCEFP3_187012). All computations were carried out on the Kebnekaise computing cluster, as part of the High Performance Computing Center North (HPC2N), which is funded by the Swedish National Infrastructure for Computing (SNIC), as well as the Kempe Foundations and the Knut and Alice Wallenberg Foundation.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor declared a past co-authorship with the author AA.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank two reviewers and the editor Charles Davis for valuable input on earlier versions of this manuscript. Further, the authors thank the members of the Gothenburg Global Biodiversity Centre (GGBC) for feedback during the early stages of this project, in particular Søren Faurby.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.839407/full#supplementary-material
Footnotes
References
Albrecht, J., Peters, M. K., Becker, J. N., Behler, C., Classen, A., Ensslin, A., et al. (2021). Species richness is more important for ecosystem functioning than species turnover along an elevational gradient. Nat. Ecol. Evol. 5, 1582–1593. doi: 10.1038/s41559-021-01550-9
Aranda, S. C., and Lobo, J. M. (2011). How well does presence-only-based species distribution modelling predict assemblage diversity? A case study of the Tenerife flora. Ecography 34, 31–38. doi: 10.1111/j.1600-0587.2010.06134.x
Australian Government (2021). National Surface Water Information. Available at: https://www.ga.gov.au/scientific-topics/national-location-information/national-surface-water-information (Accessed February 5, 2022).
Bagousse-Pinguet, Y. L., Soliveres, S., Gross, N., Torices, R., Berdugo, M., and Maestre, F. T. (2019). Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality. Proc. Natl. Acad. Sci. U. S. A. 116, 8419–8424. doi: 10.1073/pnas.1815727116
Barthlott, W., Hostert, A., Kier, G., Küper, W., Kreft, H., Mutke, J., et al. (2007). Geographic patterns of vascular plant diversity at continental to global scales (Geographische muster der Gefäßpflanzenvielfalt im kontinentalen und globalen Maßstab). Erdkunde 61, 305–315. doi: 10.3112/erdkunde.2007.04.01
Baselga, A. (2010). Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 19, 134–143. doi: 10.1111/j.1466-8238.2009.00490.x
Breidenbach, J., Ivanovs, J., Kangas, A., Nord-Larsen, T., Nilsson, M., and Astrup, R. (2021). Improving living biomass C-stock loss estimates by combining optical satellite, airborne laser scanning, and NFI data. Can. J. For. Res. 51, 1472–1485. doi: 10.1139/cjfr-2020-0518
Bruelheide, H., Dengler, J., Jiménez-Alfaro, B., Purschke, O., Hennekens, S. M., Chytrý, M., et al. (2019). sPlot – A new tool for global vegetation analyses. J. Veg. Sci. 30, 161–186. doi: 10.1111/jvs.12710
Brummitt, N., Araújo, A. C., and Harris, T. (2021). Areas of plant diversity—what do we know? Plants People Planet 3, 33–44. doi: 10.1002/ppp3.10110
Byrne, M., Steane, D. A., Joseph, L., Yeates, D. K., Jordan, G. J., Crayn, D., et al. (2011). Decline of a biome: evolution, contraction, fragmentation, extinction and invasion of the Australian Mesic zone biota. J. Biogeogr. 38, 1635–1656. doi: 10.1111/j.1365-2699.2011.02535.x
Byrne, M., Yeates, D. K., Joseph, L., Kearney, M., Bowler, J., Williams, M. A. J., et al. (2008). Birth of a biome: insights into the assembly and maintenance of the Australian arid zone biota. Mol. Ecol. 17, 4398–4417. doi: 10.1111/j.1365-294X.2008.03899.x
Cavender-Bares, J., Gamon, J. A., and Townsend, P. A. (Eds.) (2020). Remote Sensing of Plant Biodiversity. Cham, Switzerland: Springer Nature.
Chytrý, M., Hennekens, S. M., Jiménez-Alfaro, B., Knollová, I., Dengler, J., Jansen, F., et al. (2016). European vegetation archive (EVA): an integrated database of European vegetation plots. Appl. Veg. Sci. 19, 173–180. doi: 10.1111/avsc.12191
Cingolani, A. M., Vaieretti, M. V., Gurvich, D. E., Giorgis, M. A., and Cabido, M. (2010). Predicting alpha, beta and gamma plant diversity from physiognomic and physical indicators as a tool for ecosystem monitoring. Biol. Conserv. 143, 2570–2577. doi: 10.1016/j.biocon.2010.06.026
Commonwealth Scientific and Industrial Research Organisation (2014). ASRIS—Australian Soil Resource Information System. Available at: https://www.asris.csiro.au/index.html (Accessed February 5, 2022).
Cook, L. G., Hardy, N. B., and Crisp, M. D. (2015). Three explanations for biodiversity hotspots: small range size, geographical overlap and time for species accumulation. An Australian case study. New Phytol. 207, 390–400. doi: 10.1111/nph.13199
Davis, C. C., Champ, J., Park, D. S., Breckheimer, I., Lyra, G. M., Xie, J., et al. (2020). A new method for counting reproductive structures in digitized herbarium Specimens using mask R-CNN. Front. Plant Sci. 11:1129. doi: 10.3389/fpls.2020.01129
Edwards, R. D., Crisp, M. D., Cook, D. H., and Cook, L. G. (2017). Congruent biogeographical disjunctions at a continent-wide scale: quantifying and clarifying the role of biogeographic barriers in the Australian tropics. PLoS One 12:e0174812. doi: 10.1371/journal.pone.0174812
Fick, S. E., and Hijmans, R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Fox, A. (2007). Wild Habitats: A Natural History of Australian Ecosystems. Syndey, New South Wales: ABC books.
Gholizadeh, H., Gamon, J. A., Helzer, C. J., and Cavender-Bares, J. (2020). Multi-temporal assessment of grassland α- and β-diversity using hyperspectral imaging. Ecol. Appl. 30:e02145. doi: 10.1002/eap.2145
Gioia, P., and Hopper, S. D. (2017). A new phytogeographic map for the southwest Australian floristic region after an exceptional decade of collection and discovery. Bot. J. Linn. Soc. 184, 1–15. doi: 10.1093/botlinnean/box010
Goldie, X., Gillman, L., Crisp, M., and Wright, S. (2010). Evolutionary speed limited by water in arid Australia. Proc. R. Soc. B 277, 2645–2653. doi: 10.1098/rspb.2010.0439
González-Orozco, C. E., Ebach, M. C., Laffan, S., Thornhill, A. H., Knerr, N. J., Schmidt-Lebuhn, A. N., et al. (2014). Quantifying Phytogeographical regions of Australia using geospatial turnover in species composition. PLoS One 9:e92558. doi: 10.1371/journal.pone.0092558
Govaerts, R., Nic Lughadha, E., Black, N., Turner, R., and Paton, A. (2021). The world checklist of vascular plants, a continuously updated resource for exploring global plant diversity. Sci. Data 8:215. doi: 10.1038/s41597-021-00997-6
Hopper, S. D., Silveira, F. A. O., and Fiedler, P. L. (2016). Biodiversity hotspots and Ocbil theory. Plant and Soil 403, 167–216. doi: 10.1007/s11104-015-2764-2
Kahle, D., and Wickham, H. (2013). Ggmap: spatial visualization with ggplot2. R J. 5:144. doi: 10.32614/RJ-2013-014
Kattge, J., Díaz, S., Lavorel, S., Prentice, I. C., Leadley, P., Bönisch, G., et al. (2011). TRY—a global database of plant traits. Glob. Chang. Biol. 17, 2905–2935. doi: 10.1111/j.1365-2486.2011.02451.x
Keith, D. A., and Auld, T. D. (2017). “Conservation of Australian vegetation,” in Australian Vegetation (Cambridge, England: Cambridge University Press), 677–710.
Kier, G., Mutke, J., Dinerstein, E., Ricketts, T. H., Küper, W., Kreft, H., et al. (2005). Global patterns of plant diversity and floristic knowledge. J. Biogeogr. 32, 1107–1116. doi: 10.1111/j.1365-2699.2005.01272.x
Koleff, P., Gaston, K. J., and Lennon, J. J. (2003). Measuring beta diversity for presence–absence data. J. Anim. Ecol. 72, 367–382. doi: 10.1046/j.1365-2656.2003.00710.x
Kreft, H., and Jetz, W. (2007). Global patterns and determinants of vascular plant diversity. PNAS 104, 5925–5930. doi: 10.1073/pnas.0608361104
Laffan, S. W., Rosauer, D. F., Di Virgilio, G., Miller, J. T., González-Orozco, C. E., Knerr, N., et al. (2016). Range-weighted metrics of species and phylogenetic turnover can better resolve biogeographic transition zones. Methods Ecol. Evol. 7, 580–588. doi: 10.1111/2041-210X.12513
Leishman, M. R., Gallagher, R. V., Catford, J. A., Morgan, J. W., Grice, A. C., and Setterfield, S. A. (2017). “Invasive plants and pathogens in Australia,” in Australian Vegetation (Cambridge, England: Cambridge University Press), 207–229.
Lindgren, N., Wästlund, A., Bohlin, I., Nyström, K., Nilsson, M., and Olsson, H. (2021). Updating of forest stand data by using recent digital photogrammetry in combination with older airborne laser scanning data. Scand. J. For. Res. 36, 401–407. doi: 10.1080/02827581.2021.1936153
Lundberg, S., and Lee, S.-I. (2017). A Unified Approach to Interpreting Model Predictions. Available at: https://arxiv.org/abs/1705.07874v2 (Accessed February 5, 2022).
MacArthur, R. H. (1965). Patterns of species diversity. Biol. Rev. 40, 510–533. doi: 10.1111/j.1469-185X.1965.tb00815.x
Macintyre, P. D., and Mucina, L. (2021). The biomes of Western Australia: a vegetation-based approach using the zonality/azonality conceptual framework. N. Z. J. Bot. 0, 1–23. doi: 10.1080/0028825X.2021.1890154
Moat, J., Orellana-Garcia, A., Tovar, C., Arakaki, M., Arana, C., Cano, A., et al. (2021). Seeing through the clouds—mapping desert fog oasis ecosystems using 20 years of MODIS imagery over Peru and Chile. Int. J. Appl. Earth Obs. Geoinf. 103:102468. doi: 10.1016/j.jag.2021.102468
Mutke, J., and Barthlott, W. (2005). Patterns of vascular plant diversity at continental to global scale. Biol. Skr. 55, 521–537.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial Ecoregions of the world: A new map of life on earth. Bioscience 51, 933–938. doi: 10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Ooi, M. K. J., Auld, T., Beaumont, L., and Bradstock, R. (2017). “Climatic influence over vegetation pattern and process,” in Australian Vegetation (Cambridge, England: Cambridge University Press), 182–206.
Park, D. S., Willis, C. G., Xi, Z., Kartesz, J. T., Davis, C. C., and Worthington, S. (2020). Machine learning predicts large scale declines in native plant phylogenetic diversity. New Phytol. 227, 1544–1556. doi: 10.1111/nph.16621
Piccolo, R. L., Warnken, J., Chauvenet, A. L. M., and Castley, J. G. (2020). Location biases in ecological research on Australian terrestrial reptiles. Sci. Rep. 10:9691. doi: 10.1038/s41598-020-66719-x
Raven, P. H., Gereau, R. E., Phillipson, P. B., Chatelain, C., Jenkins, C. N., and Ulloa Ulloa, C. (2020). The distribution of biodiversity richness in the tropics. Sci. Adv. 6:eabc6228. doi: 10.1126/sciadv.abc6228
Revermann, R., Finckh, M., Stellmes, M., Strohbach, B. J., Frantz, D., and Oldeland, J. (2016). Linking land surface phenology and vegetation-plot databases to model terrestrial plant α-diversity of the Okavango Basin. Remote Sens. 8:370. doi: 10.3390/rs8050370
Ritter, C. D., Häggqvist, S., Karlsson, D., Sääksjärvi, I. E., Muasya, A. M., Nilsson, R. H., et al. (2019). Biodiversity assessments in the 21st century: the potential of insect traps to complement environmental samples for estimating eukaryotic and prokaryotic diversity using high-throughput DNA metabarcoding. Genome 62, 147–159. doi: 10.1139/gen-2018-0096
Sabatini, F. M., Lenoir, J., Hattab, T., Arnst, E. A., Chytrý, M., Dengler, J., et al. (2021). sPlotOpen – An environmentally balanced, open-access, global dataset of vegetation plots. Glob. Ecol. Biogeogr. 30, 1740–1764. doi: 10.1111/geb.13346
Söderberg, J., Wallerman, J., Almäng, A., Möller, J. J., and Willén, E. (2021). Operational prediction of forest attributes using standardised harvester data and airborne laser scanning data in Sweden. Scand. J. For. Res. 36, 306–314. doi: 10.1080/02827581.2021.1919751
Sparrow, B., Tokmakoff, A., Leitch, E., Guerin, G., O’Neill, S., Macdonald, C., et al. (2021). TERN surveillance monitoring program: plant occurrence. Version 1.0.0. Terrestrial ecosystem research Network (TERN). (Dataset). Available at: https://portal.tern.org.au/tern-surveillance-monitoring-plant-occurrence/23235 (Accessed December 19, 2021).
Steffen, W. (2009). Australia’s Biodiversity and Climate Change. Clayton, Victoria: Csiro Publishing.
Swenson, N. G. (2011). The role of evolutionary processes in producing biodiversity patterns, and the interrelationships between taxonomic, functional and phylogenetic biodiversity. Am. J. Bot. 98, 472–480. doi: 10.3732/ajb.1000289
Thornhill, A. H., Mishler, B. D., Knerr, N. J., González-Orozco, C. E., Costion, C. M., Crayn, D. M., et al. (2016). Continental-scale spatial phylogenetics of Australian angiosperms provides insights into ecology, evolution and conservation. J. Biogeogr. 43, 2085–2098. doi: 10.1111/jbi.12797
Tuomisto, H. (2010a). A diversity of beta diversities: straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 33, 2–22. doi: 10.1111/j.1600-0587.2009.05880.x
Tuomisto, H. (2010b). A diversity of beta diversities: straightening up a concept gone awry. Part 2. Quantifying beta diversity and related phenomena. Ecography 33, 23–45. doi: 10.1111/j.1600-0587.2009.06148.x
Večeřa, M., Divíšek, J., Lenoir, J., Jiménez-Alfaro, B., Biurrun, I., Knollová, I., et al. (2019). Alpha diversity of vascular plants in European forests. J. Biogeogr. 46, 1919–1935. doi: 10.1111/jbi.13624
Venn, S., Kirkpatrick, J., McDougall, K., Walsh, N., Whinam, J., and Williams, R. J. (2017). “Alpine, sub-alpine and sub-Antarctic vegetation of Australia,” in Australian Vegetation (Cambridge, England: Cambridge University Press), 461–489.
Venter, O., Sanderson, E. W., Magrach, A., Allan, J. R., Beher, J., Jones, K. R., et al. (2016). Global terrestrial human footprint maps for 1993 and 2009. Sci. Data 3:160067. doi: 10.1038/sdata.2016.67
Vittoz, P., and Guisan, A. (2007). How reliable is the monitoring of permanent vegetation plots? A test with multiple observers. J. Veg. Sci. 18, 413–422. doi: 10.1111/j.1654-1103.2007.tb02553.x
von Humboldt, A. (1817). De distributione geographica plantarum secundum coeli temperiem et altitudinem montium prolegomena. Edinburgh, Scotland: libraria graeco-latino-germanica.
Whittaker, R. H. (1960). Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 30, 279–338. doi: 10.2307/1943563
Whittaker, R. H. (1972). Evolution and measurement of species diversity. Taxon 21, 213–251. doi: 10.2307/1218190
Wilson, J. B., Peet, R. K., Dengler, J., and Pärtel, M. (2012). Plant species richness: the world records. J. Veg. Sci. 23, 796–802. doi: 10.1111/j.1654-1103.2012.01400.x
Yeates, D. K., Metcalf, D. J., Westcott, D. A., and Butler, A. (2014). “Australia’s biodiversity: status and trends,” in Biodiversity: Science and Solutions for Australia (Melbourne, Victoria: CSIRO Publishing).
Keywords: neural network, machine learning, species richness, biodiversity, plant, Australia, diversity pattern, deep learning
Citation: Andermann T, Antonelli A, Barrett RL and Silvestro D (2022) Estimating Alpha, Beta, and Gamma Diversity Through Deep Learning. Front. Plant Sci. 13:839407. doi: 10.3389/fpls.2022.839407
Edited by:
Charles Davis, Harvard University, United StatesReviewed by:
John Arthur Gamon, University of Nebraska-Lincoln, United StatesVinod Kumar, Government Degree College, Ramban, India
Copyright © 2022 Andermann, Antonelli, Barrett and Silvestro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tobias Andermann, dG9iaWFzYW5kZXJtYW5uODhAZ21haWwuY29t