 Zunji Jian1
Zunji Jian1 Lixiong Zeng1,2
Lixiong Zeng1,2 Lei Lei1,2*
Lei Lei1,2* Changfu Liu1,2
Changfu Liu1,2 Yafei Shen1,2
Yafei Shen1,2 Jiajia Zhang1
Jiajia Zhang1 Wenfa Xiao1,2
Wenfa Xiao1,2 Mai-He Li3,4,5
Mai-He Li3,4,5- 1Key Laboratory of Forest Ecology and Environment of National Forestry and Grassland Administration, Ecology and Nature Conservation Institute, Chinese Academy of Forestry, Beijing, China
- 2Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing, China
- 3Forest Dynamics, Swiss Federal Institute for Forest, Snow and Landscape Research WSL, Birmensdorf, Switzerland
- 4Key Laboratory of Geographical Processes and Ecological Security in Changbai Mountains, Ministry of Education, School of Geographical Sciences, Northeast Normal University, Changchun, China
- 5School of Life Science, Hebei University, Baoding, China
Forest management changes the physical environments and nutrient dynamics and then regulates the forest productivity. Soil phosphorus (P) availability is critical for productivity in tropical and subtropical forests. However, it was still poorly understood how soil P content and fraction respond to various forest management practices in these regions. Here, we measured the soil total P, available P, and Hedley’s P fractions, including inorganic and organic P (Pi and Po), in subtropical pine plantations treated with understory removal (UR), non-dominant species thinning (NDST) and dominant species thinning (DST) after nine years. Compared to plantations without management (CK), treatments such as UR, NDST, and DST decreased soil total P at 0–10 cm and soil available P at 0–10 cm and 10–20 cm. Increases in resin-Pi, NaOH-Pi, and C.HCl-Pi resulted in a higher total Pi in 0–10 cm (p < 0.05) in treated plots (UR, NDST, and DST) than in CK plots. UR, NDST, and DST treatments increased NaHCO3-Po and NaOH-Po (p < 0.05) but decreased C.HCl-Po at a depth of 0–10 cm. Regardless of management treatments, soil total P, available P, and P fractions in 0–10 cm showed higher contents than those in 10–20 cm. There were positive relationships between total P and total Po (p < 0.01) and between available P and total Pi. There were also positive relationships between total P, available P, NaHCO3-Pi, and NaOH-Pi (p < 0.05). In conclusion, forest management such as UR, NDST, and DST decreased soil total P and available P, and transforming soil P fractions to available P will meet the P demand following management in the pine plantations of subtropical China.
Introduction
Due to phosphorus (P) leaching losses and recalcitrant P fraction formation (Walker and Syers, 1976), low soil P availability often limits productivity in forest ecosystems, especially in tropical and subtropical regions (Vitousek et al., 2010; Reichert et al., 2022). Stand-level management practices, such as understory removal (UR) and selective thinning, improve plant growth by changing the physical environments (Trentini et al., 2017; Giuggiola et al., 2018) and nutrient cycles (Rocha et al., 2019; Qiu et al., 2020). Therefore, clarifying the responses of soil P pools to UR and thinning practices is essential in developing management strategies to improve forest productivity (Noormets et al., 2015).
Although the thinning effects on soil P pools differed in a specific forest, the global meta-analysis found that selective thinning generally increases soil total P and available P contents (Zhou et al., 2021; Zhang et al., 2023). In tropical rain forests, thinning can decrease soil total P and organic P (Imai et al., 2012; Lontsi et al., 2019). However, thinning was found not to affect soil total P in the subtropical spruce forests (Hu et al., 2016) or to decrease soil available P in subtropical coniferous mixed forests (Zhou et al., 2015). Thinning treatment increased soil available P in temperate spruce and larch plantations (Zhou et al., 2019; Zhao et al., 2023) but not in a temperate pine forest (Kaye et al., 2005). Moreover, a meta-analysis pointed out that UR decreases soil total P but increases soil available P (Zhang et al., 2022a). Usually, due to differences in plant growth and their P demand for various species (Reichert et al., 2022), soil total and available P dynamics under forest management may differ in various forests, and thus, further studies are needed.
Soil P includes inorganic (Pi) and organic (Po) forms, which are divided into multiple fractions (Hedley et al., 1982; Tiessen and Moir, 1993). Although soil total P stock is always larger than vegetation P stock (Imai et al., 2010), it is challenging to meet the P demand for plant growth due to differences in the availability of multiple P forms (Hedley et al., 1982). The P fractions extracted by Hedley’s method have been used to explore the environmental and management effects on soil P pools (Negassa and Leinweber, 2009) and provide crucial information on soil P dynamics (Liu et al., 2021). Previous studies found that variations in soil P fractions varied with thinning intensity, forest type, soil type, and geographic location (Ye et al., 2018; Liu et al., 2018b). UR treatment only significantly reduced residual-P concentration (Zhang et al., 2022b). To our knowledge, however, which soil P fractions are available as potentially usable P for plants after forest management is unknown.
Here, we collected soil samples in pine plantations treated with UR and selective thinning after nine years in subtropical China. We measured the total P, available P, and Hedley’s P fractions, including Pi and Po, to examine the differences between management treatments. Our previous study observed a significant positive effect of soil P availability on stand productivity in pine plantations (Jian et al., 2022a). We therefore hypothesize that forest management decreases soil total P and available P contents (Hypothesis 1) due to the increased plant growth after forest management (Lei et al., 2021). Because soil microbes often accelerate the transformation of Po to Pi (Fan et al., 2018; Liu et al., 2021), we expect that soil Pi rather than Po fractions will increase after forest management (Hypothesis 2). To meet increased P demand for plant growth following forest management (Table 1), we predict that soil available P increases with decreasing soil Po and residual-P fractions (Hypothesis 3). A previous study showed that ectomycorrhizal (ECM) and arbuscular mycorrhizal (AM) trees responded differently to Pi addition in tropical and subtropical forests (Liu et al., 2018a). Therefore, in the selected pine (an ECM tree) plantation, our results will provide new insights into understanding soil P dynamics under forest management.
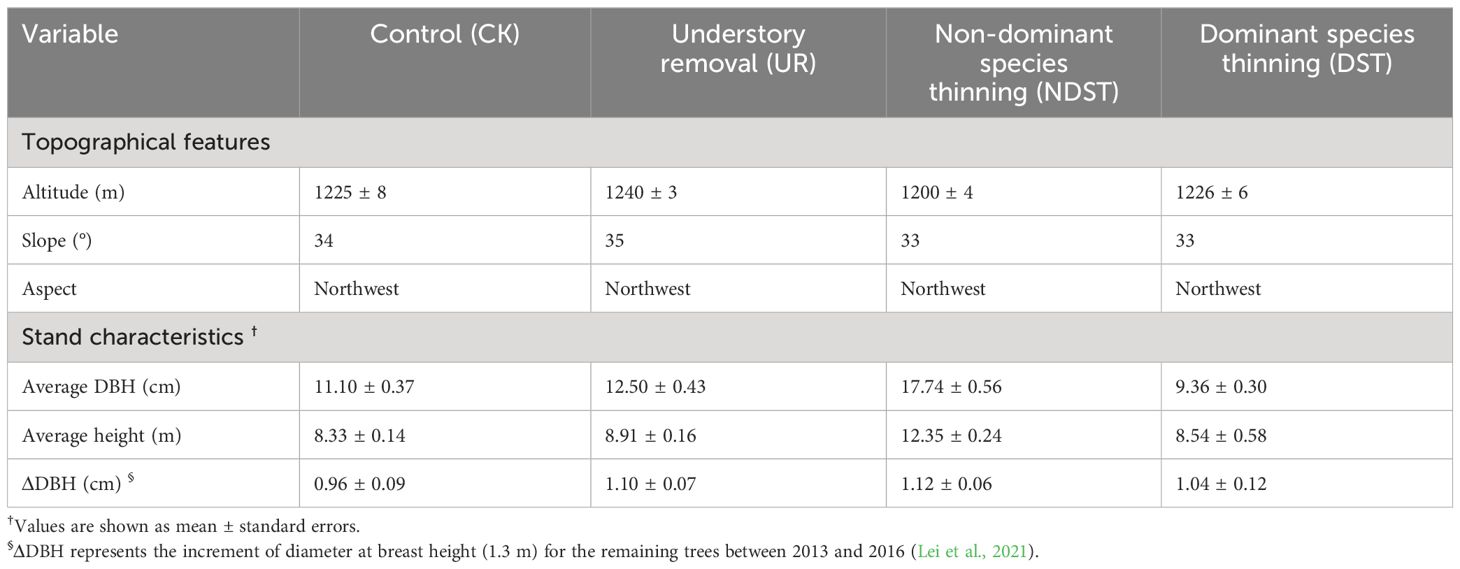
Table 1 Information on the selected plantations after treatments in September 2013.
Materials and methods
Site description
The study was conducted at Jiulingtou Forest Farm (30°59′N, 110°47′E) in subtropical China. The site is characterized by a typical humid monsoon climate, with a mean annual temperature of 16.9°C and a mean annual precipitation between 1000 mm and 1250 mm (Lei et al., 2018). The zonal soil type is yellow-brown soil (Cambisols, Shi et al., 2010) with a 1.0–1.2 m depth. Reforestation in the study region was widely implemented to alleviate land degradation resulting from deforestation of climax vegetation.
The forest management experiment was performed in an aerially seeded Pinus massoniana forest established in the 1970s. At the beginning of the forest management experiment, the stand density was approximately 1700 stems per hectare (Lei et al., 2018; Shen et al., 2018), and the species composition mainly included coexisting trees (e.g., Toxicodendron vernicifluum, Betula luminifera, and Cunninghamia lanceolataand) and understory shrubs (e.g., Pyracantha fortuneana, Litsea pungens, and Lespedeza bicolour). In the early stage of forest management, soil total P is between 0.20 g·kg-1 and 0.24 g·kg-1 in this pine plantation (Lei et al., 2018). These soil total P contents had been categorized as very low in tropical and subtropical regions (Reichert et al., 2022). In other words, the pine plantations investigated in this study represent a P-deficient ecosystem to some extent.
Experimental design
In September 2013, four experiment plots (20 m × 20 m) were set within three large pine plantations (Lei et al., 2018). One treatment for one plot in each plantation randomly put the following four practices. (1) Control (CK): the forest remains in its original state, except for slight disturbances (e.g., sampling) during the investigation. (2) Understory removal (UR): understory shrubs were removed yearly to reduce competition with overstory vegetation. (3) Non-dominant species thinning (NDST): the non-dominant species whose DBH is more than 5 cm were reduced by 15% (the basal area calculated) to reduce competition with dominant species. (4) Dominant species thinning (DST): the pine trees with a DBH of more than 17.9 cm were reduced by 75% (the basal area calculated) to reduce competition with other trees. All forest management practices have not treated any roots, living herbs, and floor litters (Lei et al., 2018) and significantly enhanced the annual increment of DBH for the remaining trees (Table 1).
Soil sampling and analyses
Mineral soils at 0–10 cm and 10–20 cm were randomly sampled from nine points using a soil auger in July 2022. The soil samples in each plot were manually mixed and air-dried indoors to pass through a 2-mm sieve. Total P and available P contents were measured by plasma emission spectroscopy (IRIS Intrepid II XSP, Thermo Fisher Scientific, USA) following digestion with NHO3-HClO4-HF solution and using a continuous flow analyzer (Analytical AA3 Auto Analyser, SEAL, Germany) after extraction with HCl-H2SO4 solution, respectively (Jian et al., 2022b). The P fractions, including resin-Pi, NaHCO3-Pi, NaHCO3-Po, NaOH-Pi, NaOH-Po, HCl-Pi, C.HCl-Pi, C.HCl-Po, and residual-P, were measured by Hedley’s method (Hedley et al., 1982) and its modification (Tiessen and Moir, 1993).
Statistical analysis
Data were tested to meet the normality requirements (Kolmogorow-Smirnow test) and homogeneity (Bartlett test) and were logarithmically transformed when necessary for subsequent analysis. One-way analyses of variance (ANOVA) and the Tukey HSD test were used to determine the differences in soil P pools among forest management practices and soil layers. The correlations among soil P pools were examined using Pearson’s correlation analysis and general linear regression models. All analyses were implemented in R 4.3.0 (R Development Core Team, 2023).
Results
Soil total P and available P contents
Forest management treatments and their interactions with soil layers did not significantly affect soil total P and available P (Supplementary Table S1). Compared to CK, treatments such as UR, NDST, and DST declined soil total P (0–10 cm; Figure 1A) and soil available P (0–10 cm and 10–20 cm; Figure 1B) with no significant level. Regardless of management treatments, soil total P (0.28 ± 0.02 g·kg-1) in-depth 0–10 cm was greater (p < 0.001) than that in-depth 10–20cm (0.25 ± 0.02 g·kg-1) (Figure 1A).
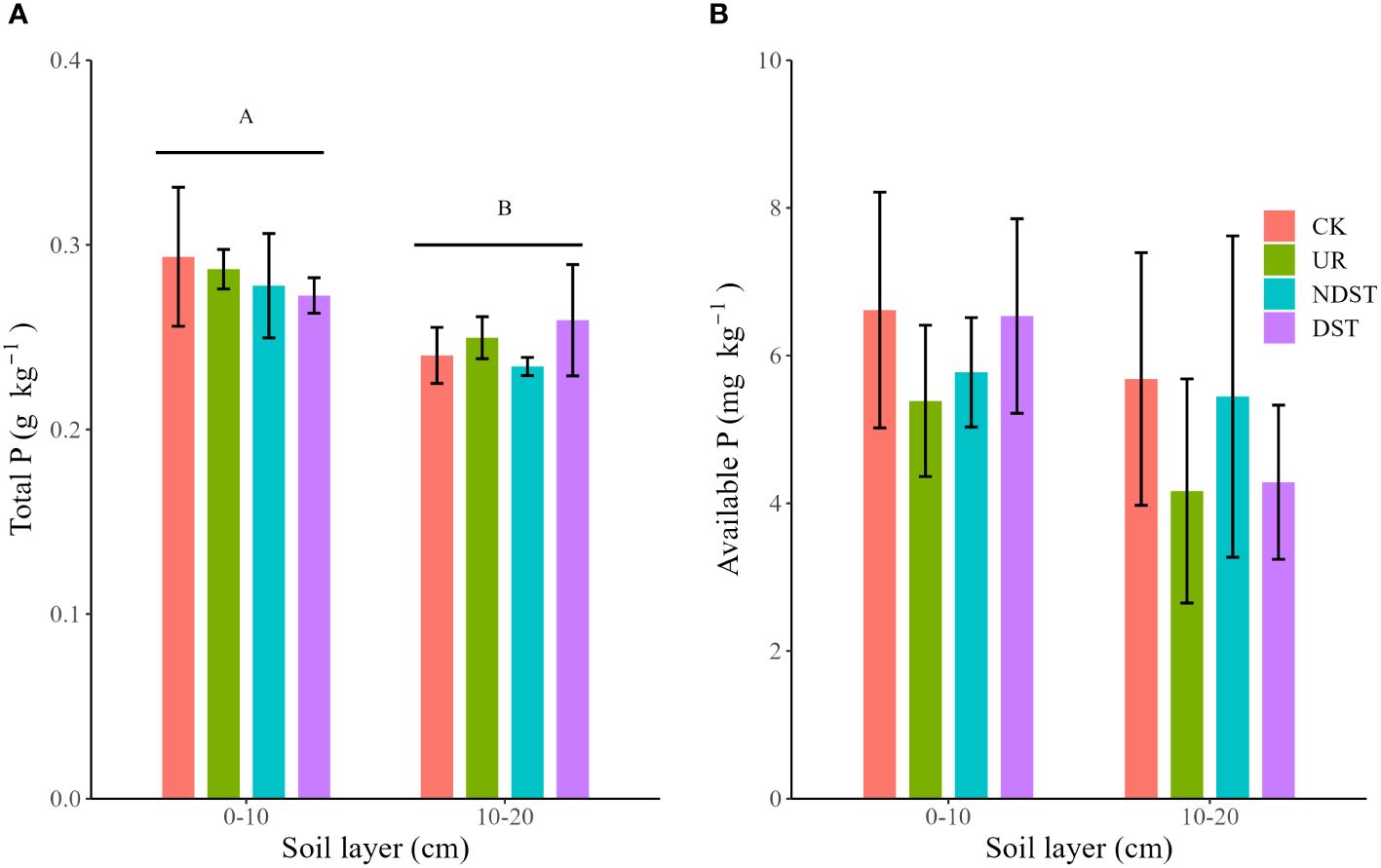
Figure 1 Comparison of total P (A) and available P (B) among forest management treatments. Values are means ± 1 standard deviation (n = 3). Different capital letters represent significant differences between soil layers (p < 0.05). CK, control; UR, understory removal; NDST, non-dominant species thinning; and DST, dominant species thinning.
Soil P fractions
Forest management treatments, soil layers, and their interactions differently affected soil P fractions (p < 0.05; Supplementary Table S1). Soil C.HCl-Pi in 0–10 cm and C.HCl-Po in 10–20 cm were higher in NDST and DST plots than in CK plots (p < 0.05; Table 2). Overall, increased resin-Pi, NaOH-Pi and C.HCl-Pi in treatment plots resulted in higher total Pi in 0–10 cm in UR (51.53 ± 2.88 mg·kg-1), NDST (60.79 ± 6.49 mg kg-1) and DST (58.26 ± 4.31 mg·kg-1) than in CK plots (46.87 ± 2.44 mg·kg-1) (p < 0.05; Table 2). Compared to CK, treatments, including UR, NDST, and DST, increased NaHCO3-Po and NaOH-Po but decreased C.HCl-Po at 0–10 cm. Moreover, differences in P fractions between 0–10 cm and 10–20 cm varied in different treatments (Table 2).
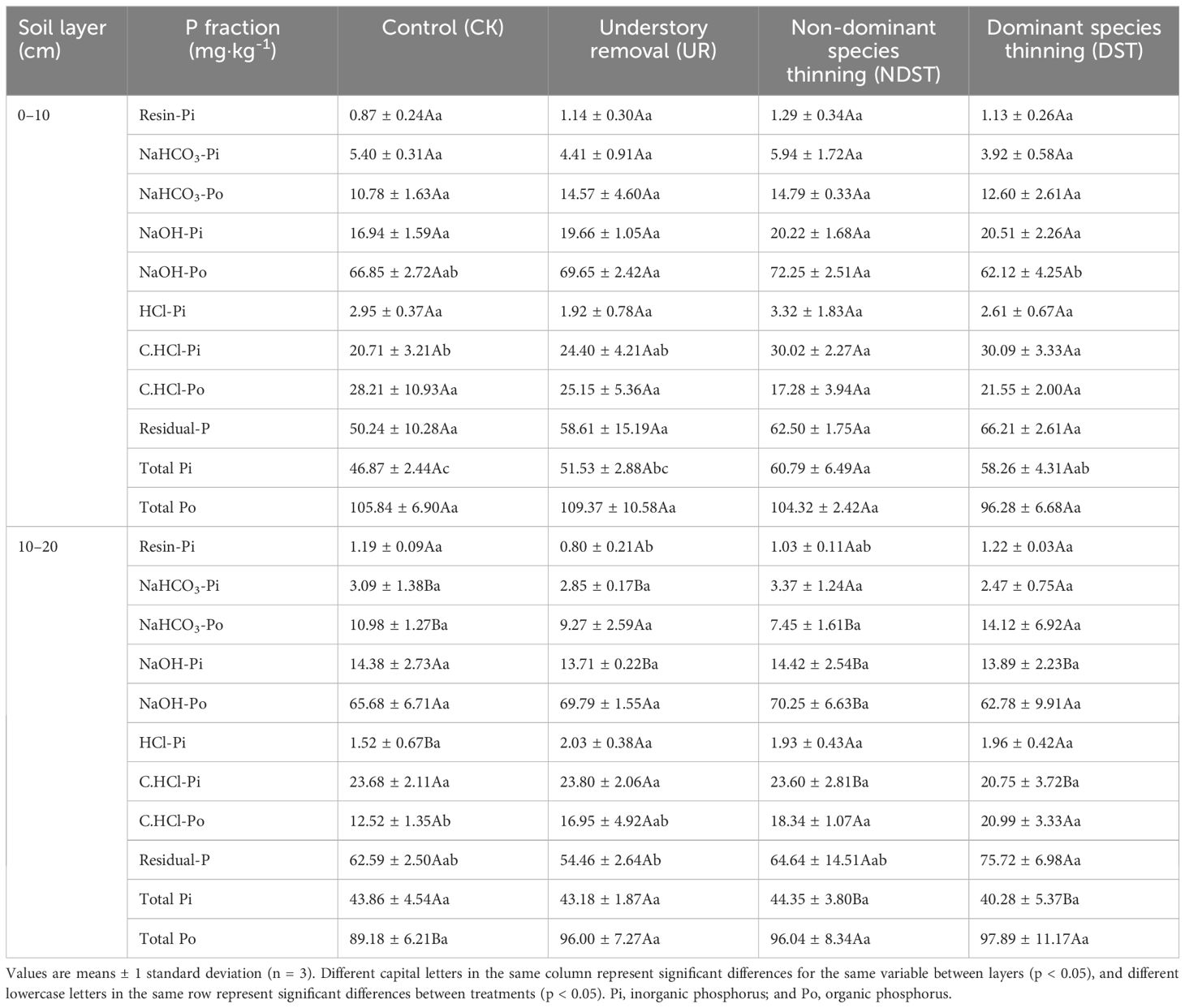
Table 2 Comparison of soil P fractions among forest management treatments.
Relationships between soil P pools
Soil total P was positively related to soil NaHCO3-Pi (R2 = 0.29, p < 0.01; Figure 2B), NaHCO3-Po (R2 = 0.32, p < 0.01; Figure 2C) and NaOH-Pi (R2 = 0.24, p < 0.05; Figure 2D). Soil total P was not significantly related to soil resin-Pi, NaOH-Po, HCl-Pi, C.CHCl-Pi, C.CHCl-Po and residual-P (Figures 2A, E–I). Soil available P was positively correlated with NaHCO3-Pi (R2 = 0.15, with a marginally significant level of 0.059; Figure 3B) and soil NaOH-Pi (R2 = 0.19, p < 0.05; Figure 3D) but was negatively correlated with soil residual-P (R2 = 0.25, p < 0.05; Figure 3I). Soil available P was not significantly correlated with soil resin-Pi, NaHCO3-Po, NaOH-Po, HCl-Pi, C.CHCl-Pi and C.CHCl-Po (Figures 3A, C, E–H). Also, there were positive relationships between total P and total Po (R2 = 0.31, p < 0.01; Figure 4B) and between available P and total Pi (R2 = 0.13, with a marginally significant level of 0.089; Figure 4C). There were not significant relationships between total P and total Pi (Figure 4A) and between available P and total Po (Figure 4D). Significantly positive relationships among NaHCO3-Pi, NaOH-Pi, and HCl-Pi, as well as between resin-Pi and residual-P, were observed (Supplementary Figure S1).
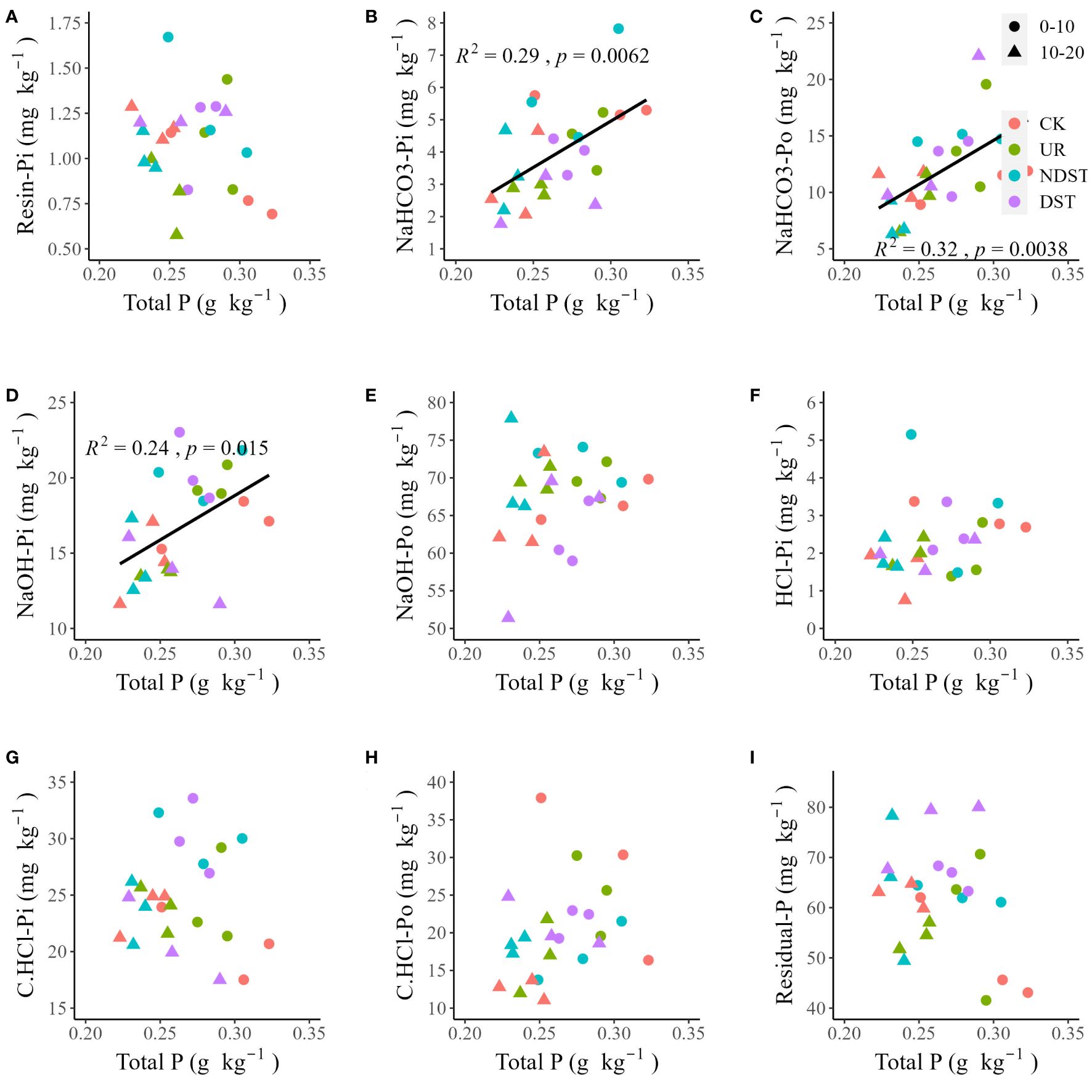
Figure 2 Correlations of soil total P with resin-Pi (A), NaHCO3-Pi (B), NaHCO3-Po (C), NaOH-Pi (D), NaOH-Po (E), HCl-Pi (F), C.HCl-Pi (G), C.HCl-Po (H) and residual-P (I). CK, control; UR, understory removal; NDST, non-dominant species thinning; DST, dominant species thinning; 0–10, 0–10 cm soil layer; 10–20, 10–20 cm soil layer; Pi, inorganic phosphorus; and Po, organic phosphorus.
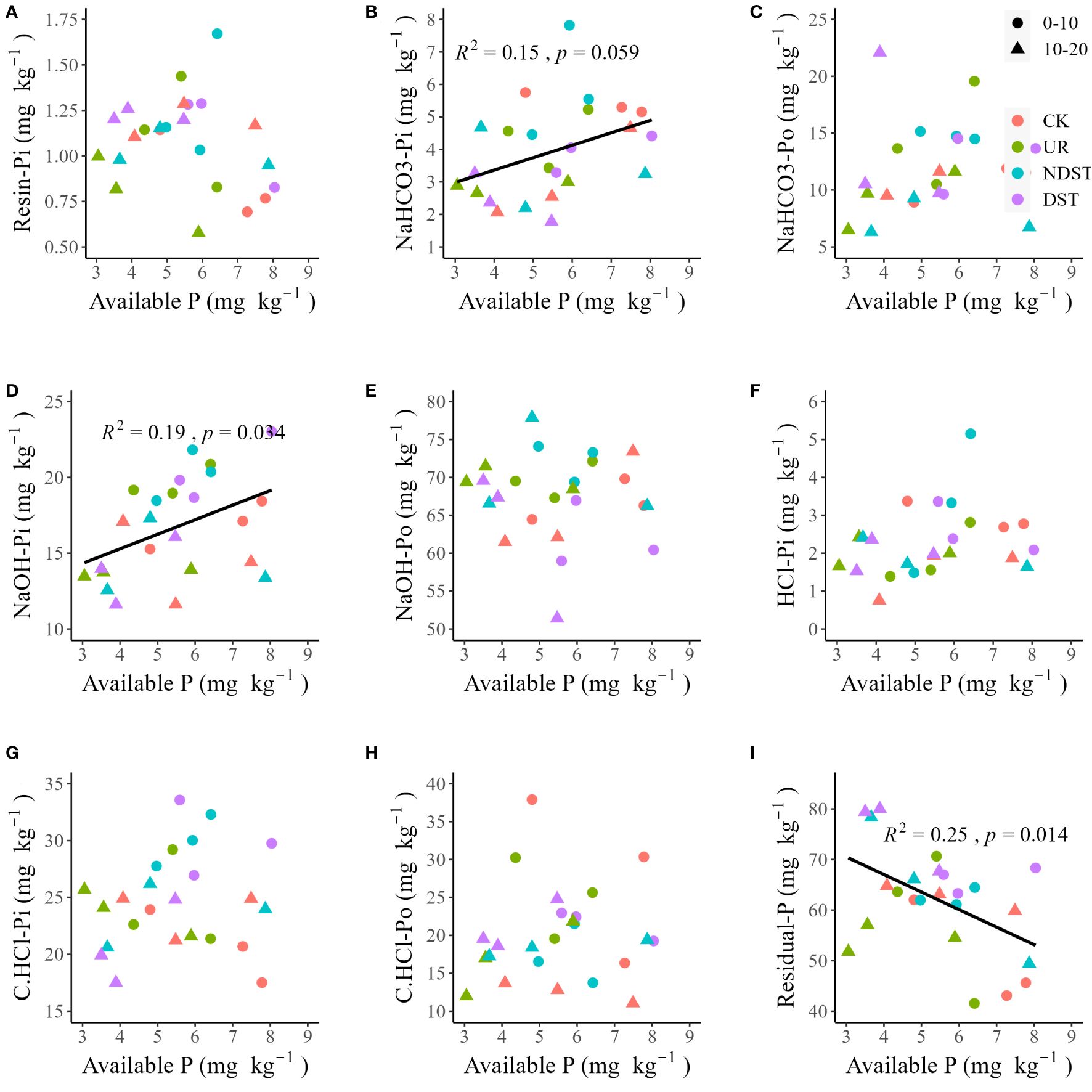
Figure 3 Correlations of soil available P with resin-Pi (A), NaHCO3-Pi (B), NaHCO3-Po (C), NaOH-Pi (D), NaOH-Po (E), HCl-Pi (F), C.HCl-Pi (G), C.HCl-Po (H) and residual-P (I). CK, control; UR, understory removal; NDST, non-dominant species thinning; DST, dominant species thinning; 0–10, 0–10 cm soil layer; 10–20, 10–20 cm soil layer; Pi, inorganic phosphorus; and Po, organic phosphorus.
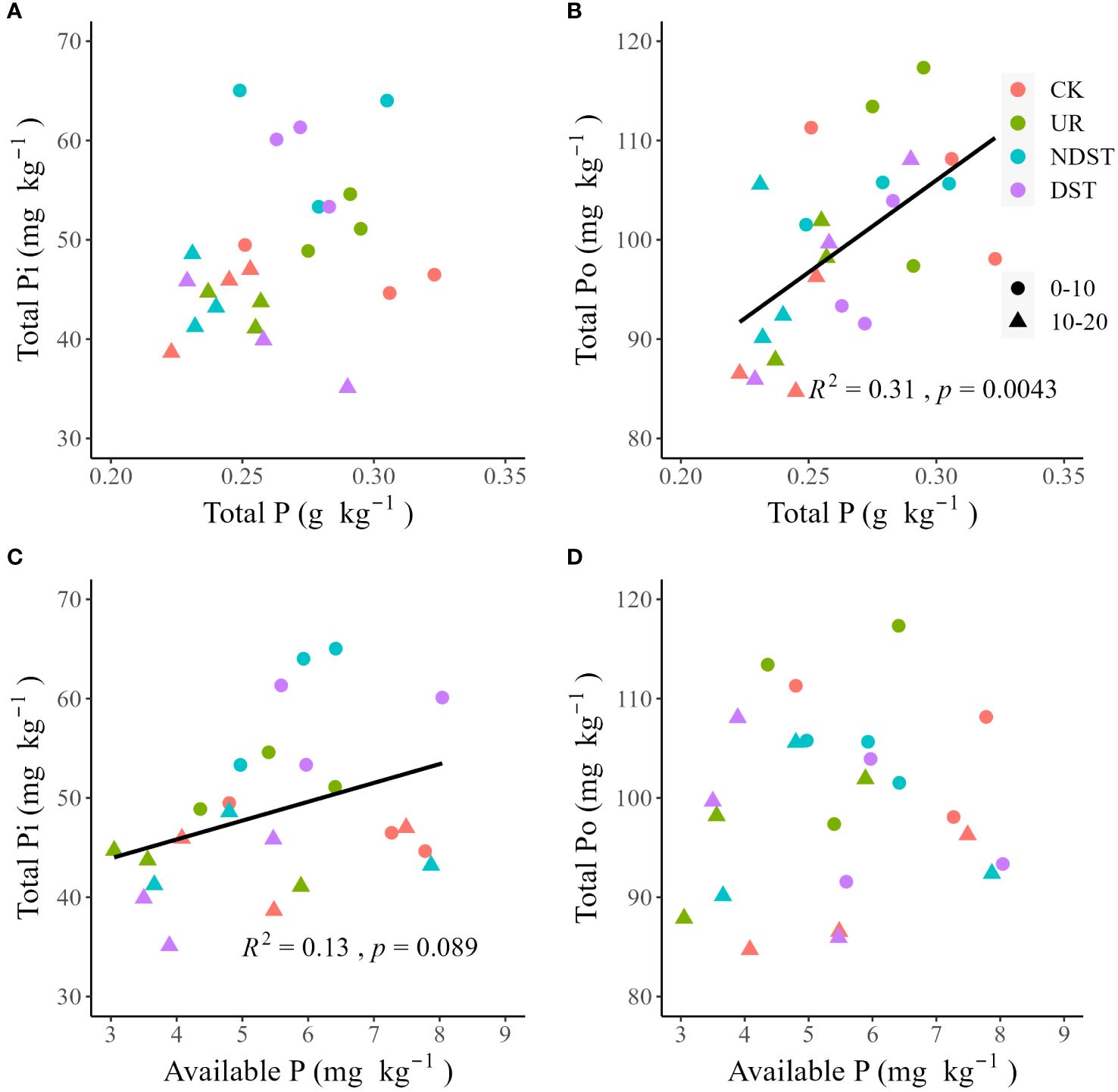
Figure 4 Correlations of total P with total Pi (A) and total Po (B), as well as correlations of available P with total Pi (C) and total Po (D). CK, control; UR, understory removal; NDST, non-dominant species thinning; DST, dominant species thinning; 0–10, 0–10 cm soil layer; 10–20, 10–20 cm soil layer; Pi, inorganic phosphorus; and Po, organic phosphorus.
Discussions
Forest management decreased soil P content
Our results identified that both total P and available P in UR, NDST, and DST plantations were lower than those in CK plots (Figure 1), which agreed with the first hypothesis but did not support the results of the previous meta-analysis (Zhou et al., 2021; Zhang et al., 2022a, 2023). In the P-deficient pine plantations (Reichert et al., 2022; Jian et al., 2022b), the rapid growth of the remaining trees after forest management (Table 1) accelerated the plant P uptake from soils (Rocha et al., 2019) and then decreased soil P. Moreover, high total P and available P contents in topsoils were likely related to the P biogeochemical cycle: movement of soil P from subsoil (Zhou et al., 2021) and return of P element in plant organs to the topsoil via litter, detritus, and roots decomposition (Hu et al., 2016). Unfortunately, this study did not measure the relevant data, and further confirmation is needed for these mechanisms.
Interestingly, total P (0.27–0.29 g·kg-1) and available P (5.39–6.62 mg·kg-1) of 0–10 cm after nine years were higher than those in the initial stage of forest management (0.22–0.24 g·kg-1 and 0.86–2.15 mg·kg-1, respectively; Shen et al., 2017; Lei et al., 2021). Forest management increased soil temperature (Zeng et al., 2023) and understory diversity (Wang et al., 2019), which accelerates litter decomposition (Hu et al., 2016) and promotes soil microbial biomass (Lei et al., 2021) and activity (Rocha et al., 2019), and then improved P availability in the pine plantations.
Diverse effects of forest management on soil P fractions
Plants can absorb directly resin-P, NaHCO3-Pi, and NaHCO3-Po from soils (Hedley et al., 1982). Compared to CK plots, these soil P fractions in UR, NDST, and DST plots increased in 0–10 cm but decreased in 10–20 cm (Table 2), consistent with previous results (Liu et al., 2018b; Ye et al., 2018). The high affinity of humic acid for Fe and Al ions can weaken the adsorption of Fe and Al ions at mineral surfaces to Pi (Gérard, 2016), thus preventing P deposition and increasing soil P availability. Also, root exudates can decompose the moderately labile P and occluded P (Broeckling et al., 2008), and microorganisms can hydrolyze Po (Zhang et al., 2016), thereby promoting the transformation of NaOH-Po and C.HCl-Po to the active P (Fan et al., 2018; Liu et al., 2021).
As a potential P source for plant absorption, soil NaOH-Pi was higher, but soil NaOH-Po was lower in DST plots than in CK plots (Table 2). These results differed from previous studies where slight and moderate logging significantly reduced NaOH-Po but increased NaOH-Pi (Liu et al., 2018b; Ye et al., 2018). Likely, the understory shrubs and herbs with arbuscular mycorrhizal symbiosis in DST plots (Wang et al., 2019) promoted the secretion of phosphatase and root exudates, thereby mineralization of Po. Moreover, several studies did not observe the thinning effects on soil occluded P (Hu et al., 2016; Liu et al., 2018b; Ye et al., 2018). However, NDST and DST treatments significantly increased soil C.HCl-Pi in 0–10 cm and C.HCl-Po in 10–20 cm in the pine plantations (Table 2). Differences in climatic factors and soil types in various study areas are likely related to the inconsistent findings, which need further confirmation.
The potential contribution of residual-P to available P after forest management
Forest management increased total Pi but decreased total Po of 0–10 cm (Table 2), supporting previous findings (Liu et al., 2018b; Ye et al., 2018) and our second hypothesis. These results suggested that forest management treatments exacerbated the P demand of plants due to high growth (Table 1). On the one hand, Pi is the main form of plant absorption and utilization (Hedley et al., 1982), and the soil Pi content is mainly dominated by the balancing process between different P fractions (Walker and Syers, 1976), leading to an increase in soil total Pi. On the other hand, the selected pine species are ECM, and mycorrhizal symbiotes can absorb Po fractions (Liu et al., 2018a; Mei et al., 2024), leading to a decrease in soil total Po. Residual-P accounted for approximately 24.6%-35.6% of soil P fractions (Table 2). It was negatively related to available P (Figure 3I) and positively associated with resin-Pi (Supplementary Figure S1). These results pointed out the potential contribution of residual-P to available P after forest management, which partially supports our third hypothesis. We also acknowledge that this finding is from correlation analysis and theoretical inference, and further confirmation of the transformation mechanism between soil P fractions is needed to determine whether plants can absorb various P fractions.
Conclusions
Compared to plantations without management, total P and available P declined while total Pi increased in topsoils in subtropical pine plantations treated by understory removal and thinning after nine years. Selective thinning treatments promoted the accumulation of occluded P, including C.HCl-Pi and C.HCl-Po. The negative relationship between residual-P and available P and the positive relationship between residual-P and resin-Pi suggest that transforming residual-P into available P may significantly contribute to the high plant P demands due to the high growth rate after forest management.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
ZJ: Formal analysis, Methodology, Writing – original draft. LZ: Conceptualization, Data curation, Writing – review & editing. LL: Conceptualization, Funding acquisition, Writing – review & editing. CL: Funding acquisition, Supervision, Writing – review & editing. YS: Investigation, Project administration, Writing – review & editing. JZ: Investigation, Writing – review & editing. WX: Conceptualization, Supervision, Writing – review & editing. M-HL: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the National Natural Science Foundation of China (32101501; 32192434).
Acknowledgments
We thank the National Forest Ecosystem Station of Three Gorges Reservoir Area in Zigui County for its assistance in field survey and Laboratory work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer QZ declared a shared affiliation with the authors ZJ, LZ, LL, CL, YS, JZ, WX to the handling editor at the time of review.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2024.1416852/full#supplementary-material
References
Broeckling, C. D., Broz, A. K., Bergelson, J., Manter, D. K., Vivanco, J. M. (2008). Root exudates regulate soil fungal community composition and diversity. Appl. Environ. Microb. 74, 738–744. doi: 10.1128/AEM.02188-07
Fan, Y., Lin, F., Yang, L., Zhong, X., Wang, M., Zhou, J., et al. (2018). Decreased soil organic P fraction associated with ectomycorrhizal fungal activity to meet increased P demand under N application in a subtropical forest ecosystem. Biol. Fert. Soils 54, 149–161. doi: 10.1007/s00374-017-1251-8
Gérard, F. (2016). Clay minerals, iron/aluminum oxides, and their contribution to phosphate sorption in soils - a myth revisited. Geoderma 262, 213–226. doi: 10.1016/j.geoderma.2015.08.036
Giuggiola, A., Zweifel, R., Feichtinger, L. M., Vollenweider, P., Bugmann, H., Haeni, M., et al. (2018). Competition for water in a xeric forest ecosystem - effects of understory removal on soil micro-climate, growth and physiology of dominant Scots pine trees. For. Ecol. Manage. 409, 241–249. doi: 10.1016/j.foreco.2017.11.002
Hedley, M. J., Stewart, J. W. B., Chauhan, B. S. (1982). Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations. Soil Sci. Soc Am. J. 46, 970–976. doi: 10.2136/sssaj1982.03615995004600050017x
Hu, B., Yang, B., Pang, X., Bao, W., Tian, G. (2016). Responses of soil phosphorus fractions to gap size in a reforested spruce forest. Geoderma 279, 61–69. doi: 10.1016/j.geoderma.2016.05.023
Imai, N., Kitayama, K., Titin, J. (2010). Distribution of phosphorus in an above-to-below-ground profile in a Bornean tropical rain forest. J. Trop. Ecol. 26, 627–636. doi: 10.1017/S0266467410000350
Imai, N., Kitayama, K., Titin, J. (2012). Effects of logging on phosphorus pools in a tropical rainforest of Boreneo. J. Trop. For. Sci. 24, 5–17. doi: 10.2307/23616947
Jian, Z., Lei, L., Ni, Y., Xu, J., Xiao, W., Zeng., L. (2022b). Soil clay is a key factor affecting soil phosphorus availability in the distribution area of Masson pine plantations across subtropical China. Ecol. Indic. 144, 109482. doi: 10.1016/j.ecolind.2022.109482
Jian, Z., Ni, Y., Lei, L., Xu, J., Xiao, W., Zeng, L. (2022a). Phosphorus is the key soil indicator controlling productivity in planted Masson pine forests across subtropical China. Sci. Total Environ. 822, 153525. doi: 10.1016/j.scitotenv.2022.153525
Kaye, J. P., Hart, S. C., Fule, P. Z., Covington, W. W., Moore, M. M., Kaye, M. W. (2005). Initial carbon, nitrogen, and phosphorus fluxes following ponderosa pine restoration treatments. Ecol. Appl. 15, 1581–1593. doi: 10.1890/04-0868
Lei, L., Xiao, W., Zeng, L., Frey, B., Huang, Z., Zhu, J., et al. (2021). Effects of thinning intensity and understory removal on soil microbial community in Pinus massoniana plantations of subtropical China. Appl. Soil Ecol. 167, 104055. doi: 10.1016/j.apsoil.2021.104055
Lei, L., Xiao, W., Zhu, J., Huang, Z., Cheng, R., Gao, S., et al. (2018). Thinning but not understory removal increased heterotrophic respiration and total soil respiration in Pinus massoniana stands. Sci. Total Environ. 621, 1360–1369. doi: 10.1016/j.scitotenv.2017.10.092
Liu, X., Burslem, D. F. R. P., Taylor, J. D., Taylor, A. F. S., Khoo, E., Majalap-Lee, N., et al. (2018a). Partitioning of soil phosphorus among arbuscular and ectomycorrhizal trees in tropical and subtropical forests. Ecol. Lett. 21, 713–723. doi: 10.1111/ele.12939
Liu, X., Cheng, X., Tian, H., Jia, X., Han, H. (2018b). Characteristics of soil phosphorus fractions under different thinning intensities in Larix principis-rupprechtii plantation and the affecting factors. Chin. J. Appl. Ecol. 29, 3941–3948. doi: 10.13287/j.1001–9332.201812.029
Liu, Y., Zhang, G., Luo, X., Hou, E., Zheng, M., Zhang, L., et al. (2021). Mycorrhizal fungi and phosphatase involvement in rhizosphere phosphorus transformations improves plant nutrition during subtropical forest succession. Soil Biol. Biochem. 153, 108099. doi: 10.1016/j.soilbio.2020.108099
Lontsi, R. T., Corre, M. D., van Straaten, O., Veldkamp, E. (2019). Changes in soil organic carbon and nutrient stocks in conventional selective logging versus reduced-impact logging in rainforests on highly weathered soils in Southern Cameroon. For. Ecol. Manage. 451, 117522. doi: 10.1016/j.foreco.2019.117522
Mei, Y., Zhang, M., Cao, G., Zhu, J., Zhang, A., Bai, H., et al. (2024). Endofungal bacteria and ectomycorrhizal fungi synergistically promote the absorption of organic phosphorus in Pinus massoniana. Plant Cell Environ. 47, 600–610. doi: 10.1111/pce.14742
Negassa, W., Leinweber, P. (2009). How does the Hedley sequential phosphorus fractionation reflect impacts of land use and management on soil phosphorus: a review. J. Plant Nutr. Soil Sc. 172, 305–325. doi: 10.1002/jpln.200800223
Noormets, A., Epron, D., Domec, J. C., McNulty, S. G., Fox, T., Sun, G., et al. (2015). Effects of forest management on productivity and carbon sequestration: a review and hypothesis. For. Ecol. Manage. 355, 124–140. doi: 10.1016/j.foreco.2015.05.019
Qiu, X., Wang, H., Peng, D., Liu, X., Yang, F., Li, Z., et al. (2020). Thinning drives C:N:P stoichiometry and nutrient resorption in Larix principis-rupprechtii plantations in North China. For. Ecol. Manage. 462, 117984. doi: 10.1016/j.foreco.2020.117984
R Development Core Team (2023). R: a language and environment for statistical computing (version 4.3.0) (Vienna, Austria: R Foundation for Statistical Computing). Available at: http://www.r-project.org/.
Reichert, T., Ramming, A., Fuchslueger, L., Lugli, L. F., Quesada, C. A., Fleischer, K. (2022). Plant phosphorus-use and -acquisition strategies in Amazonia. New Phytol. 234, 1126–1143. doi: 10.1111/nph.17985
Rocha, J. H. T., Menegale, M. L. C., Rodrigues, M., Goncalves, J. L. M., Pavinato, P. S., Foltran, E. C., et al. (2019). Impacts of timber harvest intensity and P fertilizer application on soil P fractions. For. Ecol. Manage. 437, 295–303. doi: 10.1016/j.foreco.2019.01.051
Shen, Y., Cheng, R., Xiao, W., Yang, S., Guo, Y., Wang, N., et al. (2018). Labile organic carbon pools and enzyme activities of Pinus massoniana plantation soil as affected by understory vegetation removal and thinning. Sci. Rep. 8, 573. doi: 10.1038/s41598–017-18812-x
Shen, Y., Wang, N., Cheng, R., Xiao, W., Yang, S., Guo, Y. (2017). Short-term effects of low intensity thinning on the fine root dynamics of Pinus massoniana plantations in the Three Gorges Reservoir Area, China. Forests 8, 428. doi: 10.3390/f8110428
Shi, X., Yu, D., Xu, S., Warner, E. D., Wang, H., Sun, W., et al. (2010). Cross-reference for relating Genetic Soil Classification of China with WRB at different scales. Geoderma 155, 344–350. doi: 10.1016/j.geoderma.2009.12.017
Tiessen, H., Moir, J. O. (1993). “Characterization of available P by sequential extraction,” in Soil sampling and methods of analysis. Ed. Carter, M. R. (Lewis Publishers, New York), 75–86.
Trentini, C. P., Campanello, P. I., Villagra, M., Ritter, L., Ares, A., Goldstein, G. (2017). Thinning of loblolly pine plantations in subtropical Argentina: impact on microclimate and understory vegetation. For. Ecol. Manage. 384, 236–247. doi: 10.1016/j.foreco.2016.10.040
Vitousek, P. M., Porder, S., Houlton, B. Z., Chadwick, O. A. (2010). Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 20, 5–15. doi: 10.1890/08-0127.1
Walker, T. W., Syers, J. K. (1976). The fate of phosphorus during pedogenesis. Geoderma 15, 1–19. doi: 10.1016/0016-7061(76)90066-5
Wang, X., Hu, W., Lei, L., Xiao, W., Pan, L., Zeng, L. (2019). Short-term effects of different forest management practices and spatial structure of Pinus massoniana stand. J. Southwest Forestry Univ. 39, 9–19. doi: 10.11929/j.swfu.201809021
Ye, Y., Zhao, J., Liu, C., Guan, Q. (2018). Effects of thinning on phosphorus fractions of rhizosphere soil in Pinus massoniana plantations. Chin. J. Ecol. 37, 1364–1370. doi: 10.13292/j.1000–4890.201805.035
Zeng, L., Xiao, W., Liu, C., Lei, L., Jian, Z., Shen, Y., et al. (2023). Effects of thinning and understorey removal on soil extracellular enzyme activity vary over time during forest recovery after treatment. Plant Soil 492, 457–469. doi: 10.1007/s11104-023-06187-0
Zhang, X., Chen, L., Wang, Y., Jiang, P., Hu, Y., Ouyang, S., et al. (2023). Plantations thinning: a meta-analysis of consequences for soil properties and microbial functions. Sci. Total Environ. 877, 162894. doi: 10.1016/j.scitotenv.2023.162894
Zhang, L., Xu, M., Liu, Y., Zhang, F., Hodge, A., Feng, G. (2016). Carbon and phosphorus exchange may enable cooperation between an arbuscular mycorrhizal fungus and a phosphate-solubilizing bacterium. New Phytol. 210, 1022–1032. doi: 10.1111/nph.13838
Zhang, S., Yang, X., Li, D., Li, S., Chen, Z., Wu., J. (2022a). A meta-analysis of understory plant removal impacts on soil properties in forest ecosystems. Geoderma 426, 116116. doi: 10.1016/j.geoderma.2022.116116
Zhang, X., Zhao, Q., Sun., Q., Mao, B., Zeng, D. (2022b). Understory vegetation interacts with nitrogen addition to affect soil phosphorus transformations in a nutrient-poor Pinus sylvestris var. Mongolica plantation. For. Ecol. Manage. 507, 120026. doi: 10.1016/j.foreco.2022.120026
Zhao, C., Su, N., Wang, H., Xing, H., Shen, H., Yang, L. (2023). Effects of thinning on soil nutrient availability and fungal community composition in a plantation medium-aged pure forest of Picea koraiensis. Sci. Rep. 13, 2492. doi: 10.1038/s41598–023-29498–9
Zhou, Z., Wang, C., Jin, Y., Sun, Z. (2019). Impacts of thinning on soil carbon and nutrients and related extracellular enzymes in a larch plantation. For. Ecol. Manage. 450, 117523. doi: 10.1016/j.foreco.2019.117523
Zhou, T., Wang, C., Zhou, Z. (2021). Thinning promotes the nitrogen and phosphorus cycling in forest soils. Agr. For. Meteorol. 311, 108665. doi: 10.1016/j.agrformet.2021.108665
Keywords: understory removal, selective logging, Hedley’s P fraction, soil P dynamic, P-deficient plantation, Pinus massoniana
Citation: Jian Z, Zeng L, Lei L, Liu C, Shen Y, Zhang J, Xiao W and Li M-H (2024) Effects of thinning and understory removal on soil phosphorus fractions in subtropical pine plantations. Front. Plant Sci. 15:1416852. doi: 10.3389/fpls.2024.1416852
Received: 13 April 2024; Accepted: 10 June 2024;
Published: 25 June 2024.
Edited by:
Maribela Pestana, University of Algarve, PortugalReviewed by:
Zhenfeng Xu, Sichuan Agricultural University, ChinaQian Zhang, Chinese Academy of Forestry, China
Carlos A. Alexandre, University of Evora, Portugal
Copyright © 2024 Jian, Zeng, Lei, Liu, Shen, Zhang, Xiao and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lei Lei, Y2FmbGVpbGVpQDE2My5jb20=