 Rulin Wang
Rulin Wang Xiang Guo4†
Xiang Guo4† Yanling Song
Yanling Song Mingtian Wang
Mingtian Wang- 1Water-Saving Agriculture in Southern Hill Area Key Laboratory of Sichuan Province, Chengdu, China
- 2Sichuan Provincial Rural Economic Information Center, Chengdu, China
- 3Institute of Plateau Meteorology, China Meteorological Administration, Chengdu, China
- 4Sichuan Province Agro-meteorological Center, Chengdu, China
- 5Chinese Academy of Meteorological Sciences, Beijing, China
- 6Mianyang Bureau of Meteorology, Mianyang, China
- 7College of Life Science, China West Normal University, Nanchong, China
- 8Sichuan Meteorological Observatory, Chengdu, China
Introduction: Oryza sativa is one of the most important cereal crops globally.
Methods: The aim of this study was to map areas suitable for the growth and conservation of O. sativa under current and future climatic conditions, and to observe the effects of UV variables on the distribution area of O. sativa.
Results: Based on species distribution records, we used the Biomod2 platform to combine climate data, future shared socioeconomic pathways, and elevation data. The ensemble model (EM) was constructed by screening multiple species distribution models (SDMs), including RF, GBM, ANN, and MARS. The ROC value of the joint model is greater than 0.95, indicating that the model has high reliability and accuracy. Mean annual temperature (bio01), temperature seasonality (bio04), minimum temperature in the coldest month (bio06), mean temperature of coldest quarter (bio11), human footprint and human activity impact index (hfv2geo1) and annual average ultraviolet radiation (uvb1annualmeanuv.b) were the most important environmental variables affecting the suitable distribution area of O. sativa. Under the current climate conditions, the suitable habitats of O. sativa are mainly distributed in the south of the Yangtze River. In the future climate scenario, the total suitable habitat area of O. sativa tended to decrease, but the suitable distribution area under the influence of UV was larger than that without UV.
Discussion: Climate change will significantly affect the potential distribution of O. sativa in China and increase its extinction risk. Therefore, it is necessary to provide a reference for the conservation, management, introduction and cultivation of food crops in China.
1 Introduction
Climate change is a persistent and escalating process, undeniably becoming one of the main drivers of changes in species distribution and biodiversity (Pigot et al 2023). It has a profound impact on species’ structure, function, and ecological traits (Sintayehu 2018). This impact has become a major focus in global research on species spatial patterns, particularly in terms of plant distribution. The response of vegetation to climate change and the resulting shifts in plant distribution areas are important areas of study (Chen et al. 2022; O’Connor et al 2020). Climate factors are crucial in determining plant distribution ranges, which can shift with changing climates. The IPCC (Intergovernmental Panel on Climate Change) notes that climate change will affect most natural systems with varying degrees of intensity, threatening food production and increasing the risk of natural disasters (Arias et al 2021). Research indicates that future climate change will alter the suitable climatic regions for plants, posing a serious threat to biodiversity (Tian and Jiang 2015).
Most studies on changes in plant species distribution and their future trends primarily focus on climate factors, with less attention given to the synergistic effects of habitat and climate change (Di Febbraro et al. 2023; Vaccarelli et al. 2023). However, research indicates that both climate change and land use changes jointly impact biodiversity and geographical distribution patterns (Song et al. 2021). As global warming progresses, climate change will not only alter surface temperatures and precipitation patterns but also significantly affect plant geographical distributions, causing plants to adjust their distribution in response to climate shifts. Additionally, habitat loss due to land use changes will exacerbate the impact of climate change on global species and ecosystems (Pant et al 2021). To adapt to climate change, plant communities in terrestrial ecosystems need to migrate to new suitable habitats that offer essential ecological comfort to maintain their lifecycle balance. Some plants have shown a trend of migrating to higher latitudes or elevations to track changes in their ecological niches (Burrows et al. 2014). Conversely, other studies have found that some plants are migrating to lower elevations and latitudes to adapt to changing environments (Lenoir et al. 2010). Climate change introduces high uncertainty in plant cultivation, and blind planting could impact quality and lead to resource allocation imbalances (He et al. 2021). While long-term field trials have traditionally been considered reliable for determining suitable planting areas, they are resource-intensive and require observation over multiple growth cycles (Li et al. 2019). Therefore, predicting potential suitable areas for plants under future climate change and understanding their cultivation potential in new environments is increasingly necessary.
Since the 1990s, human-induced declines in stratospheric ozone concentrations have been accompanied by an increase in ultraviolet (UV) levels compared to the past, which is likely to continue to increase in the future. This phenomenon poses a major potential threat to human health and global agricultural security (Tevini 2004). Plants are always exposed to UV-B radiation as they require light for photosynthesis. However, it is well known that exposure to UV-B radiation directly or indirectly triggers a variety of unfavorable responses that can inhibit plant growth and development (Yadav et al. 2020). Environmental factors and enhanced doses of UV-B radiation have effects on microorganisms, algae, plants, animals and human health (Farman et al. 1985). Studies have shown that UV-B radiation has a significant damaging effect on plants (Tevini and Teramura 1989). Plants, as stationary organisms, have developed a variety of morphological, physiological, biochemical and molecular level responses to various environmental changes, including increased UV radiation, over a long period of evolution. For example, plants adopt multiple protective strategies through specific UV-B signaling pathways, including increased leaf thickness, synthesis of UV-B-reflecting substances, elevated levels of antioxidants, and changes in UV-B uptake secondary metabolites at the cellular level (Yadav et al. 2020). By studying the future distribution range of plants under UV radiation, it is possible to gain a deeper understanding of the mechanisms of plant adaptation to UV-B and provide a theoretical basis for future plant variety improvement.
Rice (Oryza sativa) is one of the most important food crops globally, feeding more than half of the world’s population. It is the second most widely grown cereal crop after wheat (Carrijo et al. 2017; Fukagawa and Ziska 2019). China contributes nearly 30% of the world’s rice production, and approximately 90% of the world’s rice is produced in Asia, making China one of the largest rice producers globally (Samal et al. 2022). Rice plays a crucial role in human nutrition, as it is rich in minerals such as iron, phosphorus, and calcium, as well as vitamins B1 and B2, carbohydrates, and proteins. It is widely used in various dishes to meet human nutritional needs (Muratbek Kyzy et al. 2023). In addition to being a major food source, rice can also be used as a raw material for producing various products, including alcoholic beverages (Ito and Lacerda 2019; Meng et al. 2019) and sugar (Saithong and Lomthong 2019). Rice production is directly linked to economic stability and food security Zhu et al. (2024), having a profound impact on human survival and progress (Cao et al. 2018). However, rice growth is influenced by multiple factors, including genetic information, plant hormones, signaling molecules, nutritional status, and external environmental conditions such as light, temperature, and water (Ali and Baloch 2018; Mittler et al. 2004; Wang H. et al. 2023). Climate change, particularly extreme weather events like typhoons and high temperatures, poses a threat to rice supply (Lenaerts et al. 2019). Additionally, changes in soil nutrients, water quality, and temperature significantly affect rice growth and yield, with these factors collectively determining the growth and development performance of rice.
Species distribution models (SDMs) are essential predictive tools in conservation ecology, effectively studying the ecological relationships between species and their environments, They are capable of predicting species distributions under current and future climate scenarios (Hällfors et al. 2016). SDMs are crucial for assessing the impact of climate change on species distribution. By combining known species distribution data with environmental information, SDMs determine the ecological requirements of species and use statistical or machine learning methods to predict suitable areas under various spatial and temporal conditions. These models are widely used to understand the environmental needs of target species, simulating distribution ranges in multidimensional environments to predict potential suitable habitats under different environmental changes (Elith and Leathwick 2009). SDMs are applied in biodiversity conservation research, species invasion studies, evolutionary biology, and habitat suitability prediction (Barbet-Massin et al. 2018).
There are now dozens of SDMs, each with its own principles, algorithms, and predictive performance (Luo et al. 2017). Due to the different principles and algorithms used by each model, they each have their advantages and limitations. Additionally, the performance of each model can become unstable if the input data changes. Variations in data requirements and algorithms among different SDMs lead to differences in how environmental factors impact species distribution, introducing uncertainty into the simulation results (Guo et al. 2015). Ensemble models (EMs) improve the reliability of modeling results by combining the true data captured by individual models and minimizing their inherent errors (Dormann et al. 2018). In 2003, the integrated modeling platform Biomod, based on R software, was developed and has since been widely recognized and used (Bi et al. 2013). This package, available for free download from CRAN (cran.r-project.org), includes various species distribution modeling algorithms: GLMs (Guisan et al. 2002), generalized boosting models (GBMs) (Elith et al. 2008), generalized additive models (GAMs) (Guisan et al. 2002), classification tree analysis (CTA) (Vayssières et al. 2000), artificial neural networks (ANN) (Lek and Guégan 1999), surface range envelope (SRE) modeling (Busby 1991), flexible discriminant analysis (FDA) (Hastie et al. 1994), multivariate adaptive regression splines (MARS) (Friedman 1990), random forests (RF) (Breiman 2001) and MaxEnt (Lantschner et al. 2019). Biomod2 integrates these model results to provide more accurate species distribution predictions. Compared to single models, Biomod2 enhances prediction accuracy and increases the reliability of research findings by applying different statistical methods (Hao et al. 2019). Due to its ability to compensate for the limitations of individual models, Biomod2 has rapidly become a key tool for species distribution prediction in recent years (Zhao et al. 2021). Although each model has inherent flaws, weighting each model based on TSS (true skill statistic) or ROC (receiver operating characteristic curve) helps achieve optimal simulation performance in ensemble models (Shabani et al. 2016).
Research indicates that climate change and human activities are causing a gradual reduction in the suitable areas and habitats of most plants (Hao et al. 2019), threatening their reproductive capacity. This could also affect rice (O. sativa), potentially leading to food resource shortages. This study integrates the synergistic effects of climate change and land use changes to develop a comprehensive ecological niche model for O. sativa, identifying the main environmental factors limiting its distribution. The study also maps suitable areas for O. sativa and examines the impact of UV radiation on its suitable habitats. By considering climate and land use changes, the aim is to provide more effective references for the conservation and sustainable use of O. sativa resources and offer a framework for managing and planning other staple crops.
2 Material and methods
2.1 Species occurrence data
The distribution data for O. sativa was primarily gathered through resource-sharing platforms, including the Global Biodiversity Information Facility (GBIF, https://www.gbif.org/, accessed on April 5, 2024). Additionally, published literature and related reports were reviewed using keywords such as O. sativa in sources like Flora of China, provincial plant floras, and relevant checklists. A total of 9045 raw records were collected. To reduce the impact of spatial autocorrelation and sampling bias on the final predictions, the SDM toolbox (version 2.5) was used to process the sparse occurrence data, ensuring that only one distribution point was included per 1 km × 1 km grid cell (Zhang et al. 2024). This sparse point method was aligned with the resolution of environmental data to minimize the number of distribution points affected by spatial autocorrelation and to avoid model overfitting (Brown et al. 2017). After excluding records with uncertain information, ambiguous names, non-ground data, and duplicates, 140 valid distribution point coordinates were retained for further data processing and analysis. For ease of modeling, the distribution data file was converted to CSV format.
2.2 Environmental variables
Previous studies have not yet identified the primary environmental variables limiting the distribution of O. sativa. One objective of this research is to analyze how O. sativa adapts to environmental conditions. To achieve this, a broad selection of environmental variables was included, encompassing factors such as climate, soil, topography, and anthropogenic influences. A total of 24 environmental variables were chosen for habitat prediction. These included 19 bioclimatic variables (downloaded from the WorldClim database www.worldclim.org/data/worldclim21.html, accessed on April 18, 2024) and three topographic factors (elevation, aspect, and slope), obtained from the Geospatial Data Cloud (www.gscloud.org, accessed on April 1, 2024). The d1_ph_water data are from the Global Water Quality Database (GLOWAQ) (https://www.glowaq.org), the d1_usda data are from the United States Department of Agriculture (USDA) Soil Survey Geographic Database (SSURGO) (https://websoilsurvey.nrcs.usda.gov), hf_v2geo1 data from the Global Human Footprint Map (https://sedac.ciesin.columbia.edu), and uvb1_annual_mean_uv-b data from NASA’s OMI Satellite Observations Dataset (https://aura.gsfc.nasa.gov). The data were downloaded in raster format from the respective databases, standardized to the same grid size and coordinate system, and then converted to ASCII format using ArcGIS. Climate variables under the Representative Concentration Pathway (SSP2-4.5) scenario were used for future predictions. Future climate data were projected for three periods: the 2050s (average for 2041-2060), the 2070s (average for 2061-2080), and the 2090s (average for 2081-2100).
Correlation analysis and principal component analysis (PCA) were performed on 19 climatic factors using the R software (Figure 1) to address potential multicollinearity issues among environmental variables, which could lead to model overfitting and affect prediction accuracy. Variables with a correlation coefficient exceeding 0.8 were evaluated, and those with higher contributions to O. sativa distribution were prioritized for inclusion. Ultimately, the selected environmental variables included 8 bioclimatic indicators (bio01, bio03, bio04, bio05, bio06, bio09, bio11, and bio17), 1 topographic indicator (elevation), 2 soil indicators (d1_ph_water and d1_usda), and 1 anthropogenic indicator (hf_v2geo1) (Table 1). Additionally, the study separately modeled scenarios with and without UV radiation to analyze its effects.
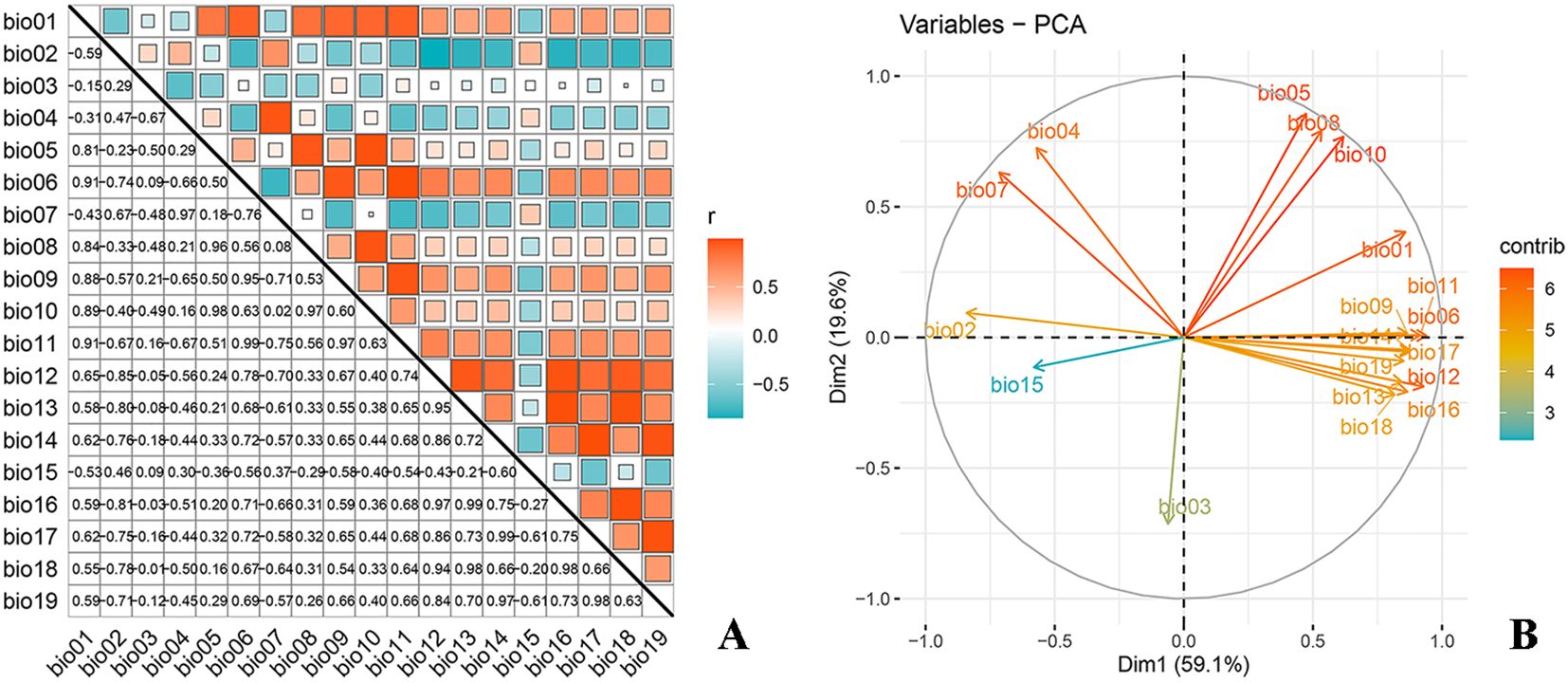
Figure 1. Correlation analysis (A) and PCA analysis (B) of environmental variables.
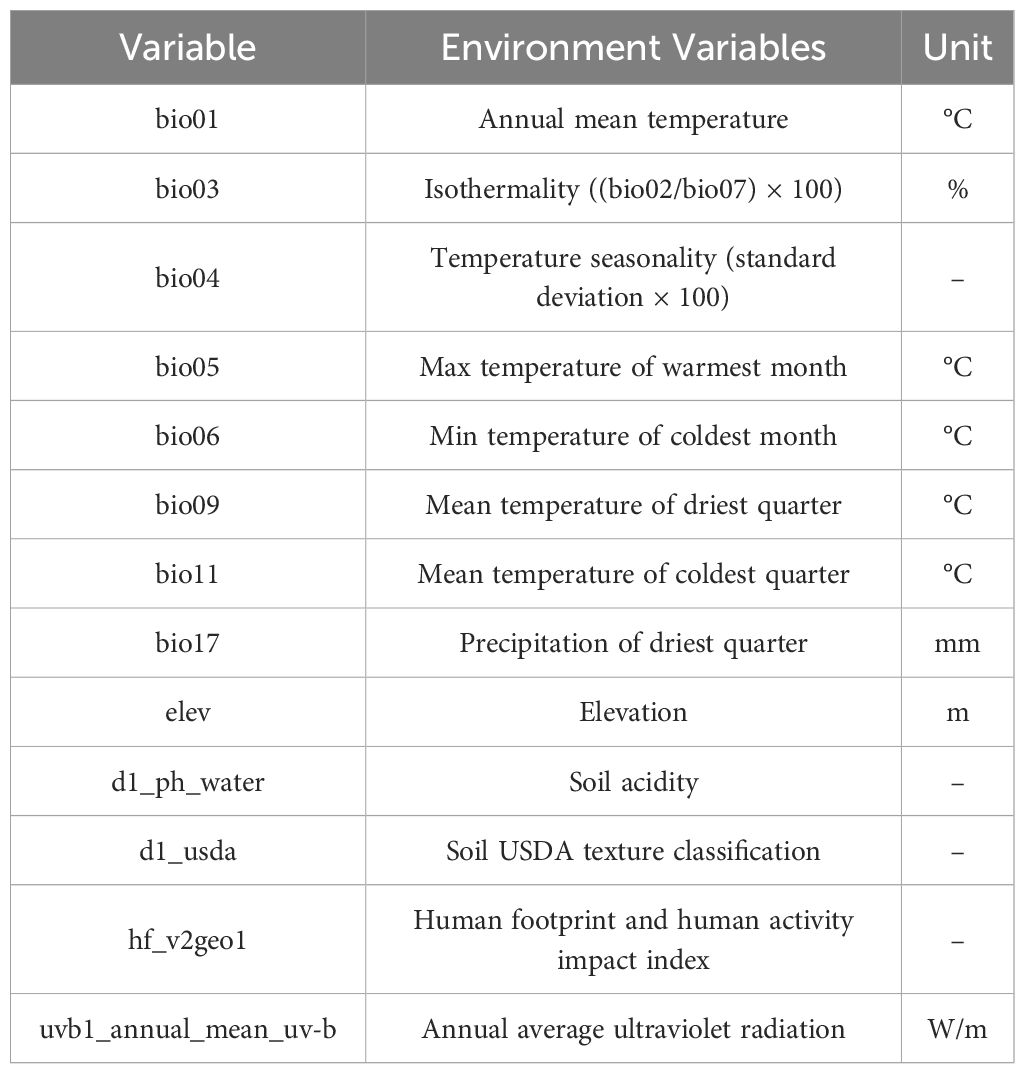
Table 1. Environmental variables retained after screening.
2.3 Construction and evaluation of SDMs
Biomod2 is a comprehensive platform that establishes the relationship between species and environmental variables using statistical and machine learning algorithms. To meet Biomod2’s modeling requirements and better simulate actual distributions, 1000 pseudo-absence points were randomly selected outside the predicted suitable range for modeling (Cengic et al. 2020).
To assess the performance of the species distribution models, the distribution data for O. sativa was randomly divided into two subsets. One subset, containing 75% of the distribution data, was used for model training, while the remaining 25% served as a test dataset to evaluate model performance. To minimize errors from a single modeling approach, this process was repeated 10 times for each model, resulting in 100 individual modeling outcomes (Bi et al. 2022).
Model accuracy was evaluated using ROC curves and the True Skill Statistic (TSS). TSS, introduced by Allouche and Kadmon in 2006, is a widely recognized and effective evaluation metric for SDMs. It combines the advantages of Kappa statistics while addressing its limitations in species occurrence with unimodal response curves (Allouche et al. 2006). TSS values range from 0 to 1, with values greater than 0.7 indicating high predictive accuracy and values below 0.5 suggesting poor accuracy. ROC values range from 0 to 1, with 0 to 0.6 indicating poor predictive performance, 0.6 to 0.8 indicating moderate performance, 0.8 to 0.9 indicating good performance, and 0.9 to 1.0 indicating excellent performance. Models with ROC > 0.90 and TSS > 0.65 were selected. Finally, the EM model was used to determine the area of suitable habitat for O. sativa under current and future climate scenarios.
2.4 Data processing
The selected environmental and distribution data for O. sativa were input into the ensemble model, and the resulting data were visualized using ArcGIS 10.8. The zones were classified based on the suitability index P, normalized by ArcGIS. A threshold of P ≥ 0.05 was used to indicate suitable habitats where the species can survive. Based on this threshold, the suitability was categorized into four classes: unsuitable (0-0.1), low suitability (0.1-0.3), moderate suitability (0.3-0.6), and high suitability (0.6-1). Grid calculations were performed to tabulate the area of each reclassified suitability category, and the weights for each zone were determined. Subsequently, the area of each layer was adjusted according to the actual land area cutout. Using the SDM_Toolbox_v2.5, the loss (areas currently suitable but becoming unsuitable in the future), stable (areas suitable now and in the future), and gain (areas currently unsuitable but becoming suitable in the future) regions for O. sativa were identified for different time periods.
3 Results
3.1 Evaluation of model prediction accuracy
To rigorously assess the accuracy of the models in predicting the distribution of O. sativa, we used data on the distribution of O. sativa in China, combined with environmental variables, and ran the models 10 times to obtain TSS and receiver operating characteristic (ROC) values for the 10 models. The prediction accuracy of individual models was assessed based on ROC and TSS values (Figure 2). The results from the ensemble species distribution models revealed variations in prediction accuracy among the 10 models. Figure 2 indicates that GAM and MAXENT had the poorest prediction accuracy, failing to accurately simulate suitable areas. In contrast, RF had the highest average TSS and ROC values, demonstrating the best accuracy and stability.
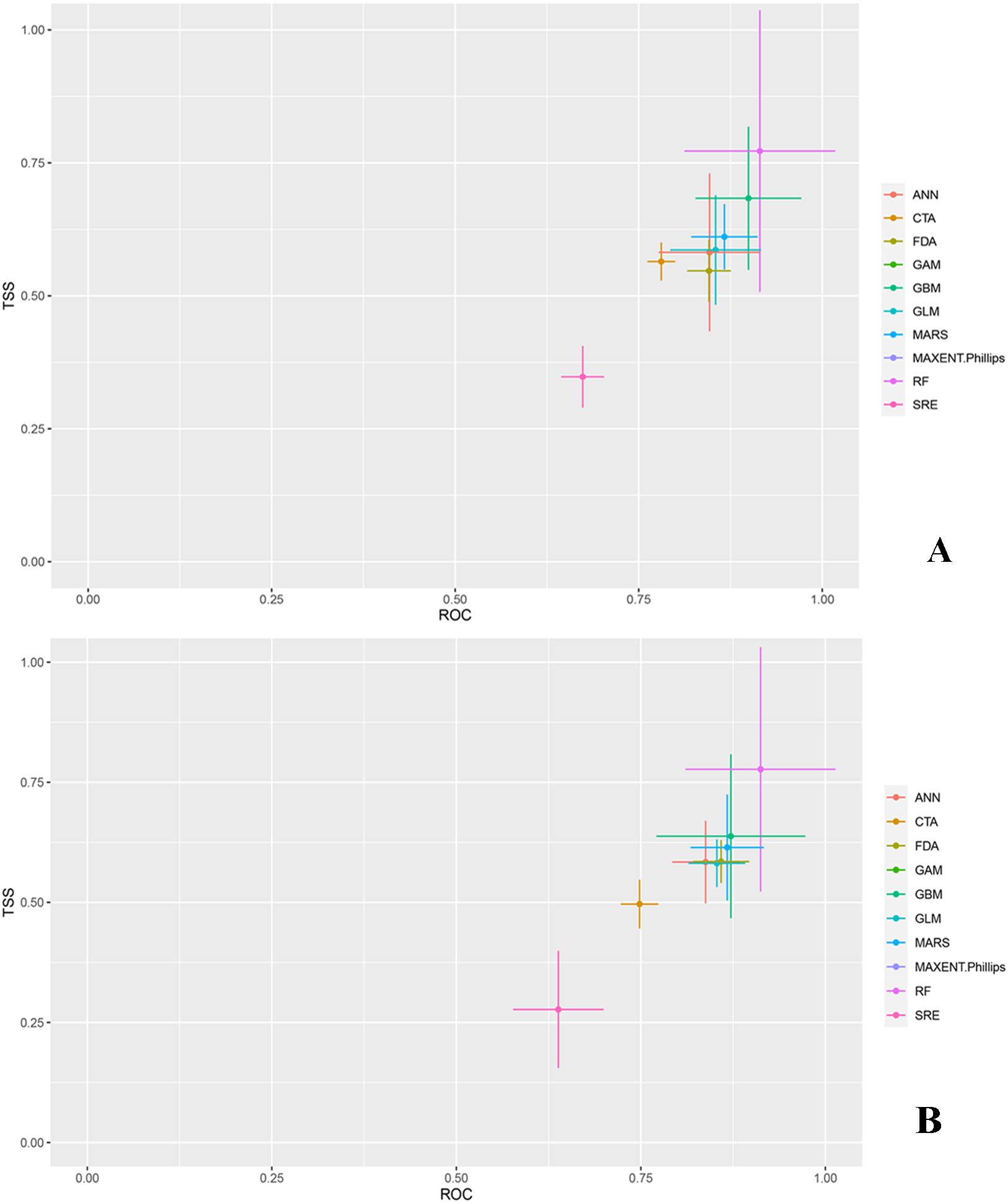
Figure 2. True skill statistics (TSS) and receiver operating characteristic curve (ROC) of 10 SDMs. (A) denotes the TSS and receiver operating characteristic (ROC) values of the model without UV variables, and (B) denotes the TSS and receiver operating characteristic (ROC) values of the model with UV variables. ANN (Artificial Neural Network), CTA (Classification Tree Analysis), FDA (Flexible Discriminant Analysis), GAM (Generalized Additive Model), GBM (Gradient Boosting Machine), GLM (Generalized Linear Model), MARS (Multivariate Adaptive Regression Splines), MAXENT (Maximum Entropy), RF (Random Forest), SRE (Species Distribution Modelling using Rule-based Ensembles).
Models with ROC values > 0.9 and TSS values > 0.65 were selected. For the no UV variable scenario, RF, GBM, ANN, and MARS were chosen, resulting in an ensemble model (EM) with ROC = 0.976 and TSS = 0.825 (Table 2). For the UV variable scenario, RF, GBM, and MARS were selected, leading to an EM with ROC = 0.977 and TSS = 0.836 (Table 2). Except for RF, the ROC and TSS values of the ensemble models were higher than those of individual models, indicating superior prediction accuracy for O. sativa suitable areas. Given its excellent performance, we chose the EM model for further visualization and analysis.
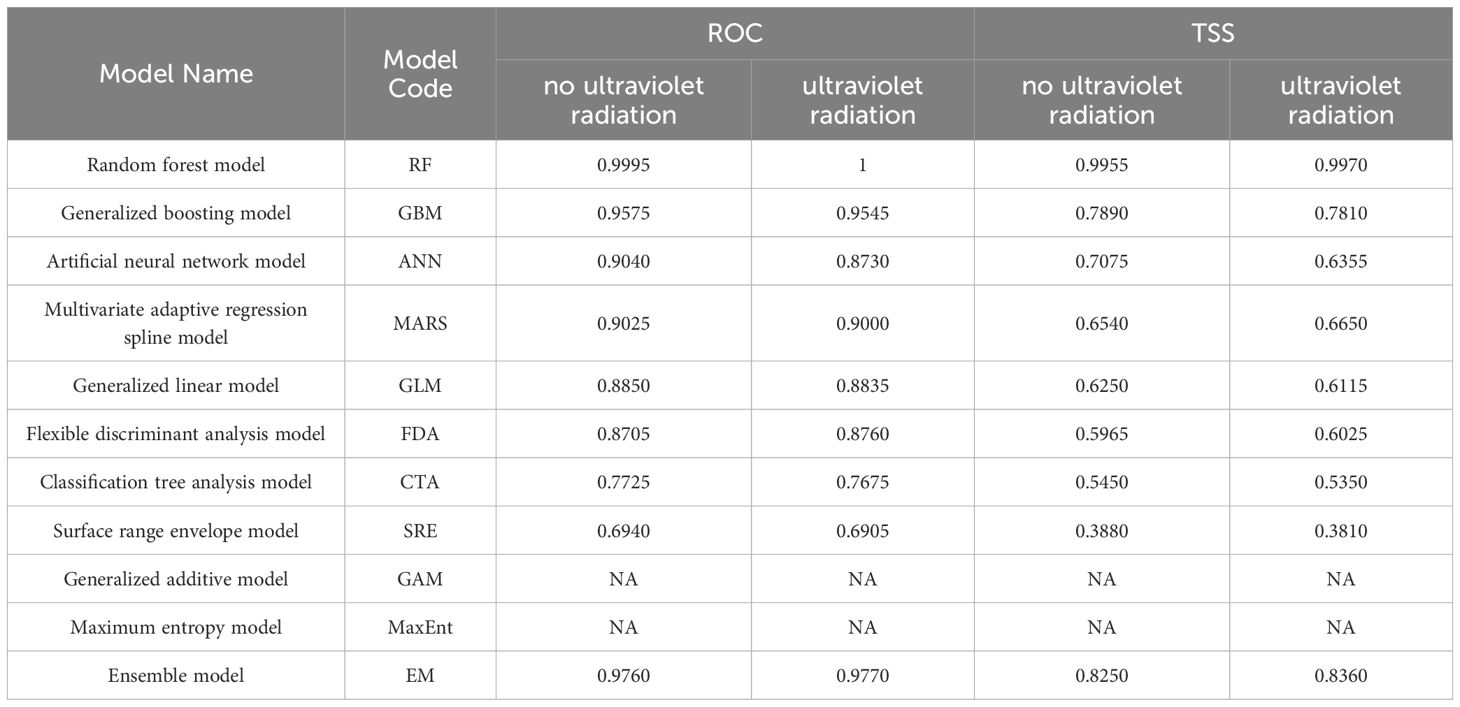
Table 2. The mean value of receiver operating characteristic curve (ROC) and true skill statistic (TSS) of different model algorithms.
3.2 Evaluation of environmental variables
The weights of the nine environmental variables varied (Table 3). In the absence of UV variables, bio01, bio04, bio06, hf_v2geo1, bio11 and altitude were the most important environmental factors affecting O. sativa. In the presence of UV variables, bio01, bio04, bio06, bio11, hf_v2geo1 and uvb1_annual_mean_uv.b were the most important environmental factors affecting O. sativa. Regardless of the UV variables, the biodiversity indices bio01, bio04 and bio06 had the highest weights and were the most important environmental factors affecting O. sativa. Overall, temperature, UV intensity and elevation play a crucial role in determining the habitat range of O. sativa.
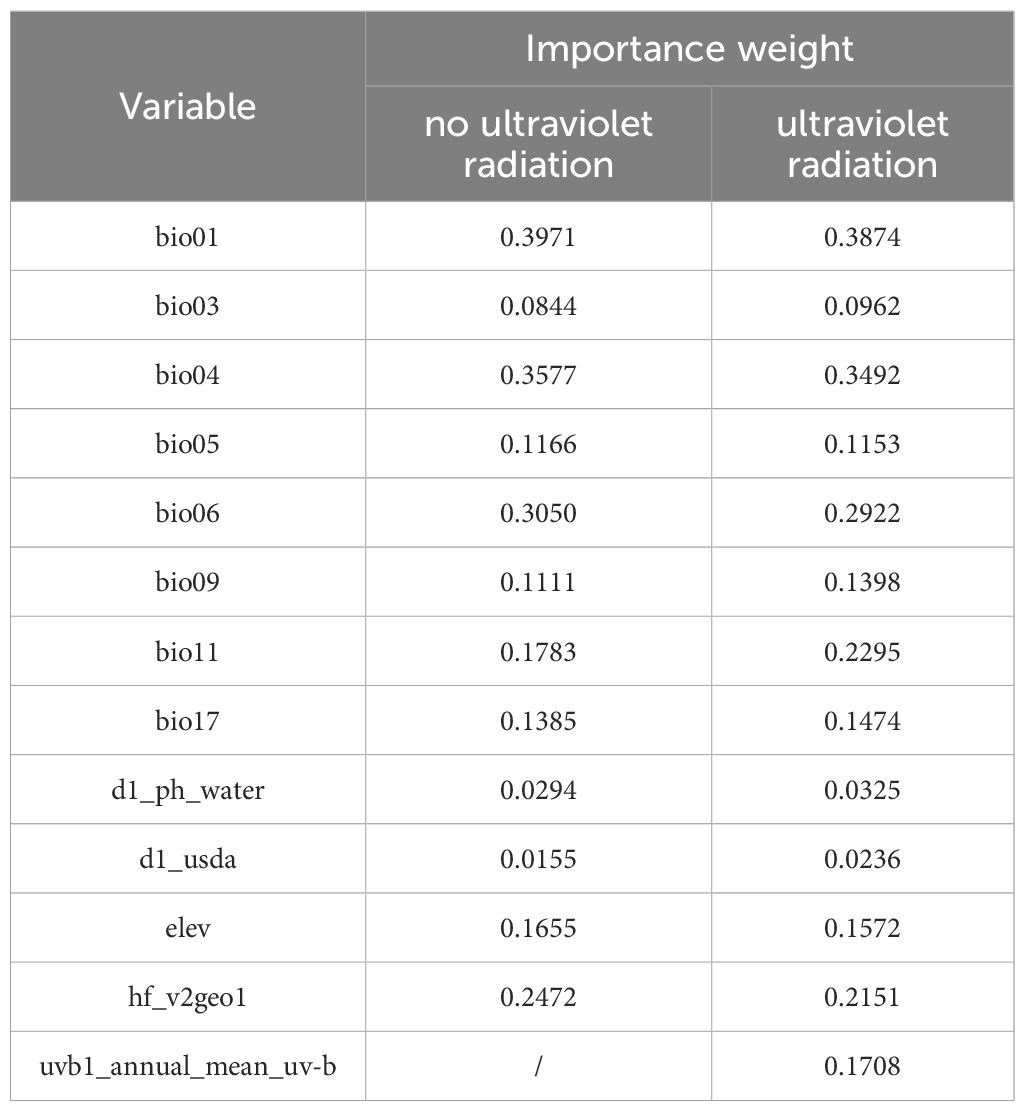
Table 3. Importance weight of environmental variables.
Based on these results, RF shows better performance compared to other models. We used the response curves generated by RF to explore the relationship between current environmental factors and the probability of occurrence. Supplementary Figure S1 illustrates that the habitat suitability of O. sativa exhibits a nonlinear relationship with various environmental variables. Whether or not the UV variable is included, habitat suitability for O. sativa improves as the annual mean temperature (bio01) exceeds 15°C. Similarly, habitat suitability increases as the lowest temperature of the coldest month (bio06) rises above 10°C. The suitability of O. sativa for different habitats first decreases and then increases with seasonal temperature changes, eventually stabilizing. The response curves for d1_ph_water, bio03, and d1_usda show that these variables have relatively stable probabilities and thus have a minor impact on the suitable distribution of O. sativa. The presence or absence of UV variables significantly affects the impact of the precipitation during the driest season (bio17) on the suitable distribution of O. sativa. Higher values of uvb1_annual_mean_uv.b lead to a decrease in habitat suitability for O. sativa.
3.3 Geographical distribution of O. sativa under current and SSP2-4.5 climates
Figure 3 shows that under current climate conditions, the suitable regions for O. sativa are primarily located in the southern part of China. With the inclusion of UV variables, high suitability regions in the current climate have decreased in the northeastern part of Taiwan, as well as in Fujian, Jiangxi, Hunan, Guangxi, Guangdong, Shanxi, Yunnan, Guizhou, and Sichuan. In contrast, high suitability regions have increased in Gansu, Hebei, and Hainan. The medium and low suitability areas in Yunnan, Guizhou, Sichuan, Chongqing, Hunan, and Hubei have decreased, while regions like Shandong, Henan, Hebei, Jiangsu, and Anhui have seen increases.
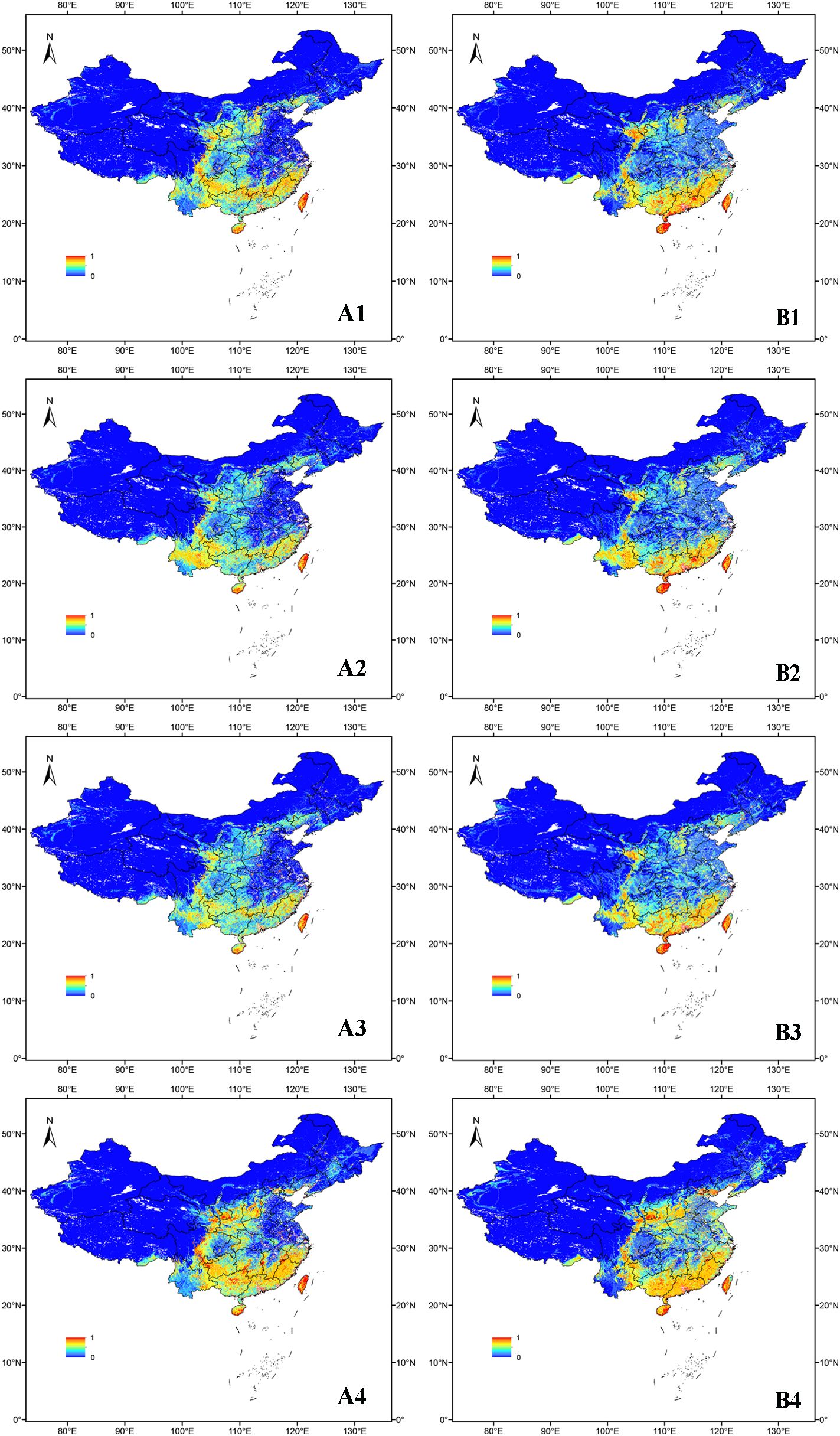
Figure 3. Suitability distribution of O. sativa in the scenarios SSP2-4.5 and current. (A) denotes the distribution without the UV variable and (B) denotes the distribution with the UV variable.1 denotes the distribution in the 2050s under SSP2-4.5, 2 denotes the distribution in the 2070s under SSP2-4.5, 3 denotes the distribution in the 2090s under SSP2-4.5, and 4 denotes the distribution under the current scenario.
In the 2050s SSP2-4.5 scenario, the highly suitable areas for O. sativa with UV variables were mainly located in southeastern China. The area of highly suitable areas was larger than that of the scenario without the UV variable, while the area of unsuitable areas was slightly larger in eastern and central China. By the 1970s, under the SSP2-4.5 scenario, the moderately suitable areas of O. sativa gradually decreased and the highly suitable areas slightly increased.
3.4 Changes of O. sativa distribution under future climatic conditions
Compared to the current climate conditions, the total suitable habitat area for O. sativa shows a decreasing trend under the SSP2-4.5 scenario (Table 4). The contraction of suitable habitats is more significant without UV variables compared to with UV variables, and the expansion of suitable habitats is less under no UV variables, with a notable reduction by half (47,764.8866 km²) by the 2070s. The “No occupancy” areas remain almost unchanged across the three time periods, while the “No change” areas decrease with time, being larger without UV variables compared to with UV variables.
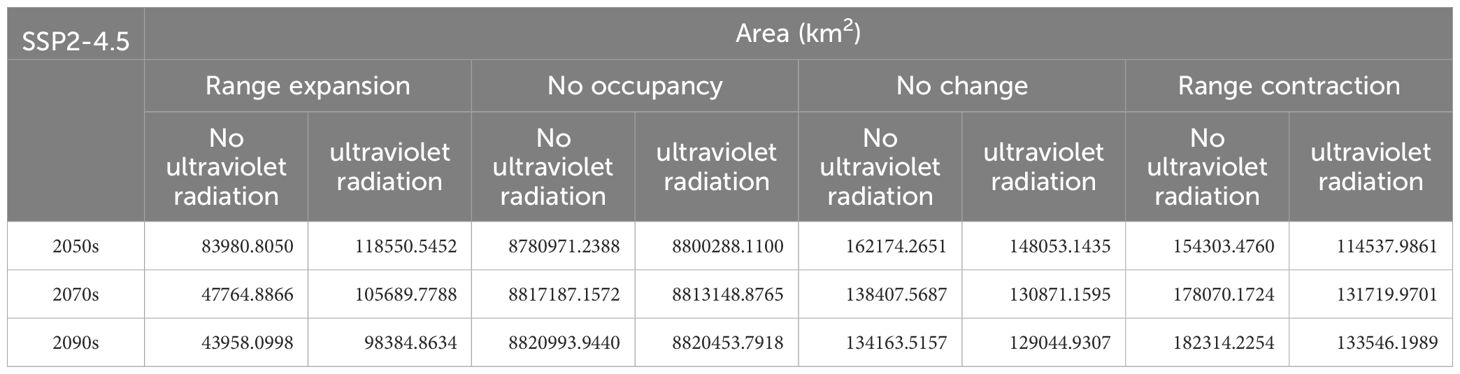
Table 4. Distribution change of O. sativa distribution in 2050s, 2070s and 2090s under SSP2-4.5.
Figure 4 illustrates that in the 2050s, the suitable habitat expansion range for O. sativa is greater with UV variables than without, particularly in Hainan, Shaanxi, and Gansu. The contraction of suitable habitats with UV variables is concentrated in Shaanxi, Shanxi, Hebei, and Beijing, whereas, without UV variables, there is additional contraction in Hunan, Jiangxi, Zhejiang, and Fujian. The “No change” areas are more extensive without UV variables than with UV variables. In the 2070s and 2090s, the expansion range of suitable habitats for O. sativa continues to decrease in both scenarios, and the contraction range continues to increase. The “No change” areas remain more extensive without UV variables compared to with UV variables.
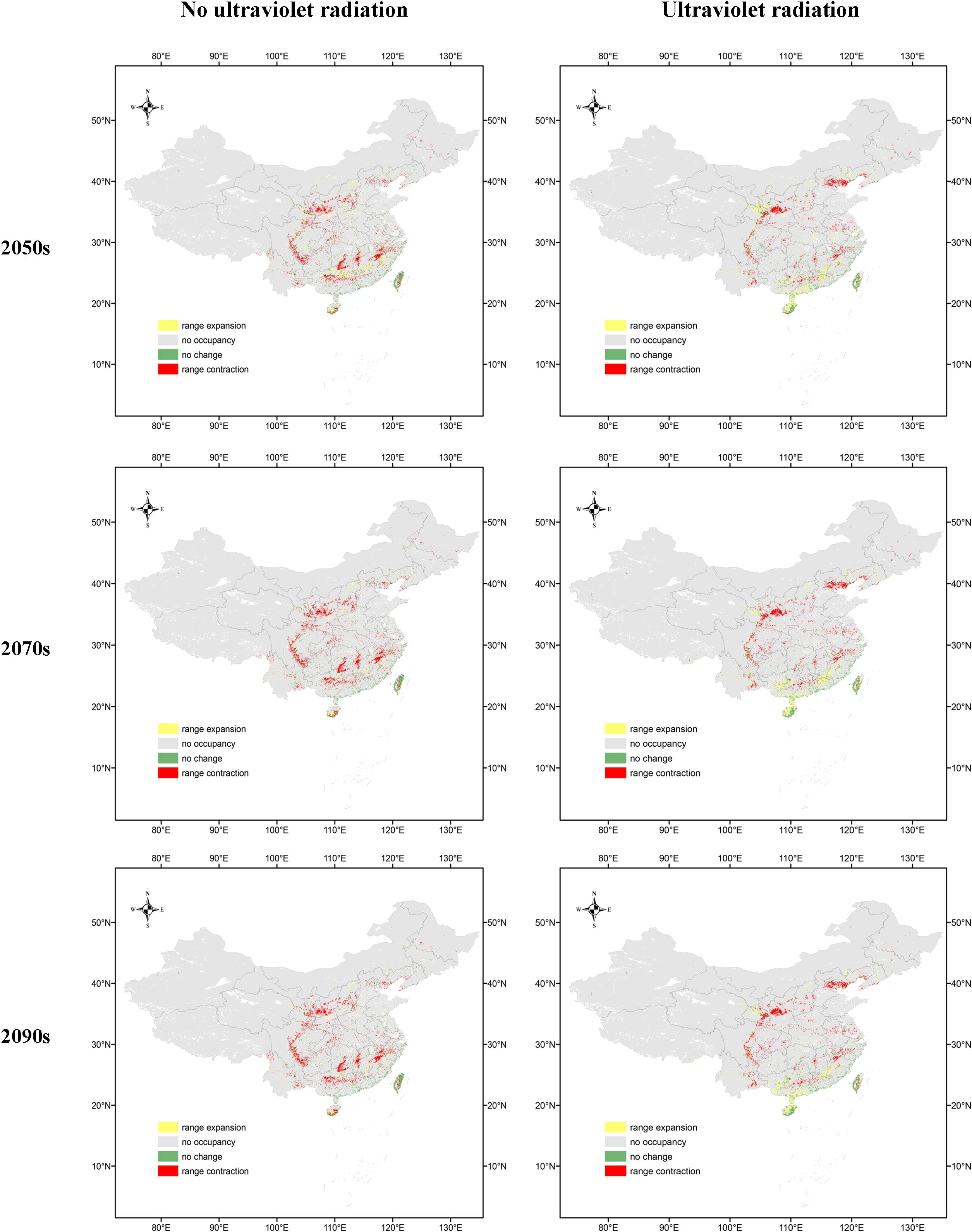
Figure 4. Distribution change of O. sativa in the scenarios SSP2-4.5.
Overall, while the suitable habitat range for O. sativa is expected to decrease in the future, the presence of UV variables results in a greater range of suitable habitats compared to scenarios without UV variables. This indicates that UV radiation is an important environmental factor influencing the distribution of O. sativa.
4 Discussion
Predictions of suitable habitat distribution are influenced by a variety of factors, including the availability of species distribution data, the choice of species distribution models (SDMs), and the type of environmental variable. Models that combine physically present and absent data generally perform better than models that rely only on physically present data when predicting the distribution of suitable habitat (Brotons et al. 2004). In addition to subtle differences in distribution records, differences in the choice of environmental variables, variable screening methods, and parameter settings can affect the bias in the final prediction results. Too many distribution points may lead to model overfitting, while too similar climate data may lead to large covariances in model results (Abdulwahab et al. 2022; Araújo and Peterson 2012). It is important to select the least correlated and biologically plausible predictors for prediction (Weng et al. 2020). The study screened 13 environmental factors for prediction through correlation analysis and principal component analysis (PCA). In order to ensure the accuracy of the prediction of O. sativa’s suitable land distribution, the data collection method was used in the study. Differences in the algorithms of different SDMs, as well as differences in the effects of various environmental factors on the distribution of species, led to some uncertainty in the prediction results of each SDM. Even when using the same dataset, different single models may produce different predictions. A potentially effective approach is to use a hybrid model, such as Biomod2, which combines a set of algorithms that use the same initial dataset and parameterization to make predictions (Di Febbraro et al. 2023; Thuiller 2014). In this study, four different SDMs (RF, GBM, ANN, and MARS) were integrated to improve the accuracy of the integrated model and overcome the limitations of the single-model approach to obtain a more accurate characterization of the geographic distribution of O. sativa. In this study, the accuracy of the models was evaluated using both ROC and TSS methods, and higher ROC values and less variability were obtained using the ensemble model compared to using individual models, which confirms the advantage of using the ensemble model to predict species occurrence (Araújo and New 2007; Breiner et al. 2015; Marmion et al. 2009). This study mapped the potential distribution of O. sativa under current and future climatic conditions. The TSS values of both the combined and RF models were above 0.80, representing excellent predictive ability (Koo et al. 2017).
Climate, topography, and soil play a key role in plant survival. Suitable temperature and moisture are essential for plants to carry out their basic physiological activities. At the same time, topographical factors such as slope, aspect, and elevation indirectly affect plant growth by influencing localized temperature, hydrological conditions, and soil properties. The results of this study showed that environmental variables such as bio01, bio04, bio06, hf_v2geo1, bio11, uvb1_annual_mean_uv.b, and elev had a greater effect on the distribution of O. sativa. O. sativa is a thermophilic plant, but too much elevated temperatures can lead to a reduction in the yield of O. sativa. There were differences in the rate of reduction, and higher temperatures can also lead to a decrease in the quality of rice. The environmental variables bio01 and bio17 have large weights, and they have a significant impact on the amount of water and heat required for O. sativa to expand northward. O. sativa is primarily adapted to grow in warm and humid environments, while dry and cold winters act as a limiting factor in its expansion range. The relatively wide range of bio04 values suggests that O. sativa may be able to adapt to regions with large temperature fluctuations. Larger isotherms imply that O. sativa is able to enhance photosynthetic efficiency at high daytime temperatures, while lower nighttime temperatures help to reduce energy consumption for respiration, thus promoting metabolite accumulation and plant growth (Li et al. 2008). Precipitation has a significant effect on above and below ground organs of plants. studies have shown that insufficient water may lead to reduced plant biomass (Wan et al. 2022), prolonged germination (Thabet et al. 2018), reduced leaf mass (Wan et al. 2022) and reduced root density (Funk et al. 2021), In contrast, adequate water helps to increase root length and leaf area (Song et al. 2022), and promotes plant germination (Plazek et al. 2018). In addition, altitude has an effect on the accumulation of metabolites and nutrients in below- and aboveground organs, which usually interacts with factors such as nutrient availability, water content (Zuo et al. 2022), temperature, UV intensity, CO2 concentration, and UV-B intensity (Zhang et al. 2018).
Solar radiation is the main influence on the growth of most plants (Jiang et al., 2020; Zuo et al., 2022). Solar radiation is one of the driving forces of photosynthesis, through which vegetation converts solar energy into chemical energy to synthesize organic matter needed for plant growth (Xu et al. 2024). Elevated UV intensity leads to decreased soluble protein content in O. sativa leaves, decreased photosynthetic activity, growth inhibition, shorter plants, and decreased yield (Xiaohua 1994). The study showed that the extent of the effect of UV on yield and yield components was greatly influenced by environmental factors such as temperature and sunshine, and that the inhibitory effects of UV on plant height, stem number, and number of spikes varied from year to year, with the inhibitory effects of UV-B being weakened in high-temperature, multi-daylight years, and becoming stronger in low-temperature, low-light years. The results of the study on the effect of UV on the quality of O. sativa indicated that UV exposure during the reproductive growth period and after tasseling had a significant effect on the size of rice grains, resulting in a significant reduction in the proportion of large rice grains (Wen-Hui et al. 2003). O. sativa leaves exposed to higher UV radiation were shorter and thicker, the dry weight of the plant body, i.e. biomass, was reduced and its cells and tissues were damaged. Total intake of root activity, protein and nutrients was reduced and stomata opened less and often closed (Jianping 1992). Enhanced UV radiation can also cause shorter plants, delayed fertility, reduced tiller number, decreased biomass and yield (Dai et al. 1992; Teramura et al. 1991), and smaller grain size of UV-irradiated rice (Kumagai et al. 2001). This is consistent with the study in this paper that future UV enhancement will reduce the suitable distribution range of O. sativa. However, we found in the effects of rice with and without UV that the contraction of the suitable habitat range was greater in the case of the no UV variable than in the case of the UV variable, and the expansion of the suitable habitat range was less in the case of the no UV variable than in the case of the UV variable, These results suggest that there may exist some kind of UV protective mechanism in rice plant to tolerate UV-B exposure (Roy and Roy 2013). Some substances in rice regulate flavonoid accumulation and are involved in UV-B tolerance in rice (Zhang et al. 2022), UV-B radiation induces the synthesis of rice proanthocyanidins B2 and C1 (Li et al. 2023), The enhanced UV-B radiation contributed to suppressing the M. oryzae infection and alleviating its damage to the photosynthesis of rice leaves (Zhang et al. 2024). Possible mechanisms of plant acclimatization to UV-B include changes in their structure, such as anatomical changes in the epidermal layer and epidermal waxes, increased leaf thickness and weight, and increased absorption of UV-B pigments (Bornman et al. 2015).
In this study, the effects of UV radiation on rice were explored in depth for the first time using a combination of models. The effects of UV on the future growth of rice were assessed by combining different modeling approaches. This innovative research approach not only provides more accurate prediction results, but also provides important theoretical support for future strategies to cope with rice cultivation and variety improvement in the context of UV enhancement.
Studies have shown that O. sativa exists in a wide range of high suitability habitats in Hainan, Taiwan and other provinces and regions, suggesting that the species has a high demand for hydrothermal environments. However, due to the complexity of environmental factors affecting plants, it is difficult to comprehensively access all environmental variables affecting the distribution of O. sativa and precisely define their suitability zones. In addition, the application of these predictions to actual planting and restoration requires consideration of factors in land use and its surroundings, such as water quality, vegetation cover, and human activities. Changes in land use, especially due to land use for economic purposes, have also led to a reduction in suitable habitat (Zhang et al., 2018). Reduction in forest cover and fragmentation of highly forested protected areas due to changes in ecological distribution may result in the loss of significant amounts of currently suitable habitat (Jia et al. 2020). In future scenarios, the suitable area for O. sativa will decline significantly. In the future, global temperatures are expected to continue to rise, extreme weather events will be frequent, and human activities will intensify (Wang X. et al. 2023). Therefore, it is expected that the suitable distribution of O. sativa will continue to shrink. Our results are consistent with those of Lonicera oblata (Wu et al. 2021).
5 Conclusions
In this study, we assessed the distribution of O. sativa in China. Potential distribution areas of O. sativa under current and future climatic conditions with and without the influence of UV variables were predicted. An ensemble model (EM) for predicting suitable habitats for O. sativa under current and future climate scenarios (SSP2-4.5) was developed by modeling and predicting four sdms, RF, GBM, ANN and MARS, selected under no UV variables, and three sdms, RF, GBM and MARS, selected under UV variables. Compared with individual models, the ROC of EM was 0.976 and 0.977, and the TSS was 0.825 and 0.836, respectively, and its prediction was more accurate. The results showed that the main environmental factors affecting its distribution were bio01, bio04, bio06, hf_v2geo1, bio11, and uvb1_annual_mean_uv.b. Under the current climatic conditions, the habitats of O. sativa are very suitable mainly distributed south of the Yangtze River. Under future climate scenarios, which will reduce the potential distribution of O. sativa, the contraction range of suitable habitat under the no UV variable is greater than in the case with the UV variable, and the expansion range of suitable habitat under the no UV variable is less than in the case with the UV variable. The prediction was highly reliable and accurately predicted the suitable range and extent of O. sativa, which is important for the future conservation of O. sativa. This study is of great significance for the conservation, management, introduction and cultivation of O. sativa, and it can also provide a reference for the study of other food crops in China.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
RW: Conceptualization, Supervision, Writing – review and editing. XG: Methodology, Software, Writing – review and editing. YS: Investigation, Methodology, Writing – review and editing. YC: Data curation, Formal analysis, Writing – review and editing. YW: Writing – original draft. MW: Software, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by the National Key Research and Development Program of China (2024YFD2301305), the National Natural Science Foundation of China (U20A2022-1), and the Heavy Rain and Drought-Flood Disasters in Plateau and Basin Key Laboratory of Sichuan Province (SCQXKJYJXZD202408; SCQXKJYJXMS202411).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1552770/full#supplementary-material
References
Abdulwahab, U. A., Hammill, E., Hawkins, C. P. (2022). Choice of climate data affects the performance and interpretation of species distribution models. Ecol. Model. 471, 110042. doi: 10.1016/j.ecolmodel.2022.110042
Ali, S., Baloch, A. (2018). Overview of sustainable plant growth and differentiation and the role of hormones in controlling growth and development of plants under various stresses. Recent Patents Food Nutr. Agric. 11 (2), 105–114. doi: 10.2174/2212798410666190619104712
Allouche, O., Tsoar, A., Kadmon, R. (2006). Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 43, 1223–1232. doi: 10.1111/j.1365-2664.2006.01214.x
Araújo, M. B., New, M. (2007). Ensemble forecasting of species distributions. Trends Ecol. Evol. 22, 42–47. doi: 10.1016/j.tree.2006.09.010
Araújo, M. B., Peterson, A. T. (2012). Uses and misuses of bioclimatic envelope modeling. Ecology 93, 1527–1539. doi: 10.1890/11-1930.1
Arias, P., Bellouin, N., Coppola, E., Jones, C., Krinner, G., Marotzke, J., et al. (2021). “Climate change 2021: the physical science basis,” in Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change; technical summary.
Barbet-Massin, M., Rome, Q., Villemant, C., Courchamp, F. (2018). Can species distribution models really predict the expansion of invasive species? PloS One 13, e193085. doi: 10.1371/journal.pone.0193085
Bi, Y., Xu, J., Li, Q., Guisan, A., Thuiller, W., Zimmermann, N. E., et al. (2013). Applying bioMod for model-ensemble in species distributions:a case study for tsuga chinensis in China. Acta Botanica Yunnanica. 35 (5), 647–655.
Bi, Y., Zhang, M., Chen, Y., Wang, A., Li, M. (2022). Applying Biomod2 for modeling of species suitabl e habitats:a case study of Paeonia lactiflora in China. Zhongguo Zhong yao za zhi = Zhongguo zhongyao zazhi = China J. Chin. materia Med. 47 (2), 376–384. doi: 10.19540/j.cnki.cjcmm.20211023.101
Bornman, J. F., Barnes, P. W., Robinson, S. A., Ballaré, C. L., Flint, S. D., Caldwell, M. M. (2015). Solar ultraviolet radiation and ozone depletion-driven climate change: effects on terrestrial ecosystems. Photochemical Photobiological Sci. 14, 88–107. doi: 10.1039/c4pp90034k
Breiner, F. T., Guisan, A., Bergamini, A., Nobis, M. P. (2015). Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol. Evol. 6, 1210–1218. doi: 10.1111/2041-210X.12403
Brotons, L., Thuiller, W., Araújo, M. B., Hirzel, A. H. (2004). Presence-absence versus presence-only modelling methods for predicting bird habitat suitability. Ecography 27, 437–448. doi: 10.1111/j.0906-7590.2004.03764.x
Brown, J. L., Bennett, J. R., French, C. M. (2017). SDMtoolbox 2.0: the next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 5, e4095. doi: 10.7717/peerj.40955
Burrows, M. T., Schoeman, D. S., Richardson, A. J., Molinos, J. G., Hoffmann, A., Buckley, L. B., et al. (2014). Geographical limits to species-range shifts are suggested by climate velocity. Nature 507, 492–495. doi: 10.1038/nature12976
Cao, Z., Qin, M., Lin, X., Zhu, Z., Chen, M. (2018). Sulfur supply reduces cadmium uptake and translocation in rice grains (Oryza sativa L.) by enhancing iron plaque formation, cadmium chelation and vacuolar sequestration. Environ. pollut. 238, 76–84. doi: 10.1016/j.envpol.2018.02.083
Carrijo, D. R., Lundy, M. E., Linquist, B. A. (2017). Rice yields and water use under alternate wetting and drying irrigation: A meta-analysis. Field Crops Res. 203, 173–180. doi: 10.1016/j.fcr.2016.12.002
Cengic, M., Rost, J., Remenska, D., Janse, J. H., Huijbregts, M. A. J., Schipper, A. M. (2020). On the importance of predictor choice, modelling technique, and number of pseudo-absences for bioclimatic envelope model performance. Ecol. Evol. 10 (21), 12307–12317. doi: 10.1002/ece3.6859
Chen, Y., le, X., Chen, Y., Cheng, W., du, J., Zhong, Q., et al. (2022). Identification of the potential distribution area of Cunninghamia lanceolata in China under climate change based on the MaxEnt model. Ying yong sheng tai xue bao = J. Appl. ecology/Zhongguo sheng tai xue xue hui Zhongguo ke xue yuan Shenyang ying yong sheng tai yan jiu suo zhu ban 33, 1207–1214. doi: 10.13287/j.1001-9332.202205.024
Dai, Q., Coronel, V., Vergara, B. S., Barnes, P. W., Quintos, A. T. (1992). Ultraviolet-B radiation effects on growth and physiology of four rice cultivars. Crop Sci. 32, 1269–1274. doi: 10.2135/cropsci1992.0011183X003200050041x
Di Febbraro, M., Bosso, L., Fasola, M., Santicchia, F., Aloise, G., Lioy, S., et al. (2023). Different facets of the same niche: Integrating citizen science and scientific survey data to predict biological invasion risk under multiple global change drivers. Global Change Biol. 29, 5509–5523. doi: 10.1111/gcb.16901
Dormann, C. F., Calabrese, J. M., Guillera-Arroita, G., Matechou, E., Bahn, V., Bartoń, K., et al. (2018). Model averaging in ecology: a review of Bayesian, information-theoretic, and tactical approaches for predictive inference. Ecol. Monogr. 88, 485–504. doi: 10.1002/ecm.1309
Elith, J., Leathwick, J. R. (2009). Species distribution models: ecological explanation and prediction across space and time. Annu. Rev. Ecology Evolution Systematics 40, 677–697. doi: 10.1146/annurev.ecolsys.110308.120159
Elith, J., Leathwick, J. R., Hastie, T. (2008). A working guide to boosted regression trees. J. Anim. Ecol. 77, 802–813. doi: 10.1111/j.1365-2656.2008.01390.x
Farman, J. C., Gardiner, B. G., Shanklin, J. D. (1985). Large losses of total ozone in Antarctica reveal seasonal ClOx/NOx interaction. Nature 315, 207–210. doi: 10.1038/315207a0
Friedman, J. H. (1990). Multivariate adaptive regression splines. Ann. Statist. 19 (1), 1–67. doi: 10.1214/aos/1176347963
Fukagawa, N. K., Ziska, L. H. (2019). Rice: importance for global nutrition. Journal of nutritional science and vitaminology 65, S2–S3. doi: 10.3177/jnsv.65.S2
Funk, J. L., Larson, J. E., Ricks-Oddie, J. L. (2021). Plant traits are differentially linked to performance in a semi-arid ecosystem. Ecology 102 (5), e3318. doi: 10.1002/ecy.v102.5
Guisan, A., Edwards, T. C., Hastie, T. (2002). Generalized linear and generalized additive models in studies of species distributions: setting the scene. Ecol. Model. 157, 89–100. doi: 10.1016/S0304-3800(02)00204-1
Guo, C., Lek, S., Ye, S., Li, W., Liu, J., Li, Z. (2015). Uncertainty in ensemble modelling of large-scale species distribution: Effects from species characteristics and model techniques. Ecol. Model. 306, 67–75. doi: 10.1016/j.ecolmodel.2014.08.002
Hällfors, M. H., Liao, J., Dzurisin, J., Grundel, R., Hyvärinen, M., Towle, K., et al. (2016). Addressing potential local adaptation in species distribution models: implications for conservation under climate change. Ecol. Appl. 26, 1154–1169. doi: 10.1890/15-0926
Hao, T., Elith, J., Guillera-Arroita, G., Lahoz-Monfort, J. J. (2019). A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Diversity Distributions 25, 839–852. doi: 10.1111/ddi.12892
Hastie, T., Tibshirani, R., Buja, A. (1994). Flexible discriminant analysis by optimal scoring. J. Am. Stat. Assoc. 89, 1255–1270. doi: 10.1080/01621459.1994.10476866
He, P., Li, J., Li, Y., Xu, N., Gao, Y., Guo, L., et al. (2021). Habitat protection and planning for three Ephedra using the MaxEnt and Marxan models. Ecol. Indic. 133, 108399. doi: 10.1016/j.ecolind.2021.108399
Ito, V. C., Lacerda, L. G. (2019). Black rice (Oryza sativa L.): A review of its historical aspects, chemical composition, nutritional and functional properties, and applications and processing technologies. Food Chem. 301, 125304. doi: 10.1016/j.foodchem.2019.125304
Jia, Y., Milne, R. I., Zhu, J., Gao, L., Zhu, G., Zhao, G., et al. (2020). Evolutionary legacy of a forest plantation tree species (Pinus armandii): Implications for widespread afforestation. Evolutionary Appl. 13, 2646–2662. doi: 10.1111/eva.13064
Jiang, H., Xu, X., Guan, M., Wang, L., Huang, Y., Jiang, Y. (2020). Determining the contributions of climate change and human activities to vegetation dynamics in agro-pastural transitional zone of northern China from 2000 to 2015. Sci. Total Environ. 718, 134871. doi: 10.1016/j.scitotenv.2019.134871
Koo, K. A., Park, S. U., Seo, C. (2017). Effects of climate change on the climatic niches of warm-adapted evergreen plants: expansion or contraction? Forests 8, 500. doi: 10.3390/f8120500
Kumagai, T., Hidema, J., Kang, H., Sato, T. (2001). Effects of supplemental UV-B radiation on the growth and yield of two cultivars of Japanese lowland rice (Oryza sativa L.) under the field in a cool rice-growing region of Japan. Agriculture Ecosyst. Environ. 83, 201–208. doi: 10.1016/S0167-8809(00)00180-8
Lantschner, M. V., de la Vega, G., Corley, J. C. (2019). Predicting the distribution of harmful species and their natural enemies in agricultural, livestock and forestry systems: an overview. Int. J. Pest Manage. 65, 190–206. doi: 10.1080/09670874.2018.1533664
Lek, S., Guégan, J. F. (1999). Artificial neural networks as a tool in ecological modelling, an introduction. Ecol. Model. 120, 65–73. doi: 10.1016/S0304-3800(99)00092-7
Lenaerts, B., Collard, B. C. Y., de Mey, Y., Demont, M. (2019). Global survey data on rice breeders’ characteristics and willingness to adopt alternative breeding methods. Data Brief 23, 103782. doi: 10.1016/j.dib.2019.103782
Lenoir, J., Gégout, J., Guisan, A., Vittoz, P., Wohlgemuth, T., Zimmermann, N. E., et al. (2010). Going against the flow: potential mechanisms for unexpected downslope range shifts in a warming climate. Ecography 33, 295–303. doi: 10.1111/j.1600-0587.2010.06279.x
Li, F., He, X., Sun, Y., Zhang, X., Tang, X., Li, Y., et al. (2019). Distinct endophytes are used by diverse plants for adaptation to karst regions. Sci. Rep. 9, 5246. doi: 10.1038/s41598-019-41802-0
Li, X., Sheng, J., Li, Z., He, Y., Zu, Y., Li, Y. (2023). Enhanced UV-B radiation induced the proanthocyanidins accumulation in red rice grain of traditional rice cultivars and increased antioxidant capacity in aging mice. Int. J. Mol. Sci. 24 (4), 3397. doi: 10.3390/ijms24043397
Li, K., Wu, W., Zheng, Y., Dai, Y., Mou, L., Liao, K. (2008). Effect of temperature on physiologic indexes, bulb yielding and total alkaloids contents of Fritillaria cirrhosa. China Journal of Chinese Materia Medica. 33 16, 1948–1951.
Luo, M., Wang, H., Lyu, Z. Y. (2017). Evaluating the performance of species distribution models Biomod2 and MaxEnt using the giant panda distribution data. Chinese Journal of Applied Ecology. 28 12, 4001–4006.
Marmion, M., Parviainen, M., Luoto, M., Heikkinen, R. K., Thuiller, W. (2009). Evaluation of consensus methods in predictive species distribution modelling. Diversity Distributions 15, 59–69. doi: 10.1111/j.1472-4642.2008.00491.x
Meng, L., Huang, T., Shi, J., Chen, J., Zhong, F., Wu, L., et al. (2019). Decreasing cadmium uptake of rice (Oryza sativa L.) in the cadmium-contaminated paddy field through different cultivars coupling with appropriate soil amendments. J. Soils Sediments 19, 1788–1798. doi: 10.1007/s11368-018-2186-x
Mittler, R., Vanderauwera, S., Gollery, M., Van Breusegem, F. (2004). Reactive oxygen gene network of plants. Trends Plant Sci. 9, 490–498. doi: 10.1016/j.tplants.2004.08.009
Muratbek Kyzy, A., Yildirim, G. H., Yilmaz, N. (2023). Effects of vermicompost applications on chlorophyll content and flag leaf area in rice (Oryza sativa L.). Cogent Food Agric. 9, 2272460. doi: 10.1080/23311932.2023.2272460
O’Connor, B., Bojinski, S., Röösli, C., Schaepman, M. E. (2020). Monitoring global changes in biodiversity and climate essential as ecological crisis intensifies. Ecol. Inf. 55, 101033. doi: 10.1016/j.ecoinf.2019.101033
Pant, G., Maraseni, T. N., Apan, A. A., Allen, B. L. (2021). Predicted declines in suitabl e habitat for greater one-horned rhinoceros (Rhinoceros unicornis) under future climate and land use change scenarios. Ecol. Evol. 11, 18288–18304. doi: 10.1002/ece3.v11.24
Pigot, A. L., Merow, C., Wilson, A., Trisos, C. H. (2023). Abrupt expansion of climate change risks for species globally. Nat. Ecol. Evol. 7, 1060–1071. doi: 10.1038/s41559-023-02070-4
Plazek, A., Dubert, F., Kopeć, P., Dziurka, M., Kalandyk, A., Pastuszak, J., et al. (2018). Seed hydropriming and smoke water significantly improve low-temperature germination of lupinus angustifolius L. Int. J. Mol. Sci. 19, 992. doi: 10.3390/ijms19040992
Roy, S. C., Roy, T. (2013). Peptide mass fingerprinting of rice (Oryza sativa L.) leaves during UV-B induced stress at seedling stage: A proteom analysis. Indian J. Biotechnol. 12, 504–508.
Saithong, P., Lomthong, T. (2019). Feasibility of Leum Pua glutinous rice substrate for sugar syrup and vinegar production by raw starch degrading enzyme hydrolysis. Int. Food Res. J. 26, 1515–1523.
Samal, P., Babu, S. C., Mondal, B., Mishra, S. N. (2022). The global rice agriculture towards 2050: An inter-continental perspective. Outlook Agric. 51, 164–172. doi: 10.1177/00307270221088338
Shabani, F., Kumar, L., Ahmadi, M. (2016). A comparison of absolute performance of different correlative and mechanistic species distribution models in an independent area. Ecol. Evol. 6, 5973–5986. doi: 10.1002/ece3.2332
Sintayehu, D. W. (2018). Impact of climate change on biodiversity and associated key ecosystem services in Africa: a systematic review. Ecosystem Health Sustainability 4, 225–239. doi: 10.1080/20964129.2018.1530054
Song, S., Chen, A., Zhu, J., Yan, Z., An, Q., Zhou, J., et al. (2022). Structure basis of the caffeic acid O-methyltransferase from Ligusiticum chuanxiong to understand its selective mechanism. Int. J. Biol. Macromolecules 194, 317–330. doi: 10.1016/j.ijbiomac.2021.11.135
Song, H., Ordonez, A., Svenning, J., Qian, H., Yin, X., Mao, L., et al. (2021). Regional disparity in extinction risk: Comparison of disjunct plant genera between eastern Asia and eastern North America. Global Change Biol. 27, 1904–1914. doi: 10.1111/gcb.15525
Teramura, A. H., Ziska, L. H., Sztein, A. E. (1991). Changes in growth and photosynthetic capacity of rice with increased UV-B radiation. Physiologia Plantarum 83, 373–380. doi: 10.1111/j.1399-3054.1991.tb00108.x
Tevini, M. (2004). “Plant responses to ultraviolet radiation stress,” in Chlorophyll a fluorescence: A signature of photosynthesis. Ed. Papageorgiou, G. C. (Springer Netherlands, Dordrecht), 605–621.
Tevini, M., Teramura, A. H. (1989). Uv-B effects on terrestrial plants. Photochem. Photobiol. 50, 479–487. doi: 10.1111/j.1751-1097.1989.tb05552.x
Thabet, S. G., Moursi, Y. S., Karam, M. A., Graner, A., Alqudah, A. M. (2018). Genetic basis of drought tolerance during seed germination in barley. PloS One 13, 11. doi: 10.1371/journal.pone.0206682
Thuiller, W. (2014). Editorial commentary on ‘BIOMOD – optimizing predictions of species distributions and projecting potential future shifts under global change’. Global Change Biol. 20, 3591–3592. doi: 10.1111/gcb.12728
Tian, Z., Jiang, D. (2015). Mid-Holocene and last glacial maximum changes in monsoon area and precipitation over China. Chin. Sci. Bull. 60, 400–410. doi: 10.1360/N972014-00718
Vaccarelli, I., Colado, R., Pallarés, S., Galassi, D. M. P., Sánchez-Fernández, D., Di Cicco, M., et al. (2023). A global meta-analysis reveals multilevel and context-dependent effects of climate change on subterranean ecosystems. One Earth 6, 1510–1522. doi: 10.1016/j.oneear.2023.09.001
Vayssières, M. P., Plant, R. E., Allen-Diaz, B. H. (2000). Classification trees: An alternative non-parametric approach for predicting species distributions. J. Vegetation Sci. 11, 679–694. doi: 10.2307/3236575
Wan, C., Dang, P., Gao, L., Wang, J., Tao, J., Qin, X., et al. (2022). How does the environment affect wheat yield and protein content response to drought? A meta-analysis. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.896985
Wang, X., Duan, Y., Jin, L., Wang, C., Peng, M., Li, Y., et al. (2023). Prediction of historical, present and future distribution of Quercus sect. Heterobalanus based on the optimized MaxEnt model in China. Acta Ecologica Sin. 43, 6590–6604. doi: 10.5846/stxb202205141353
Wang, H., Jiao, X., Zhang, X., Zhang, M., Liu, Y., Chen, X., et al. (2023). Ammonium protects rice against rice stripe virus by activating HDA703/OsBZR1-mediated BR signaling. Plant Sci. 326, 111504. doi: 10.1016/j.plantsci.2022.111504
Weng, Y., Cai, W., Wang, C. (2020). The application and future directions of the Shared Socioeconomic Pathways (SSPs) 16, 215–222. doi: 10.12006/j.issn.1673-1719.2019.078
Wen-Hui, Z., Chuan-Qing, S., Sato, T., Hidama, J., Kumagai, T., Xiang-Kun, W. (2003). Effect of supplement ultraviolet-B radiation on yield and the content of protein in rice (Oryza sativa L.). Acta Agronomica Sin. 29, 908–912.
Wu, Y., Shen, X., Tong, L., Lei, F., Mu, X., Zhang, Z. (2021). Impact of past and future climate change on the potential distribution of an endangered montane shrub lonicera oblata and its conservation implications. Forests 12, 125. doi: 10.3390/f12020125
Xu, L., Fan, Y., Zheng, J., Guan, J., Lin, J., Wu, J., et al. (2024). Impacts of climate change and human activity on the potential distribution of Aconitum leucostomum in China. Sci. Total Environ. 912, 168829. doi: 10.1016/j.scitotenv.2023.168829
Yadav, A., Singh, D., Lingwan, M., Yadukrishnan, P., Masakapalli, S. K., Datta, S. (2020). Light signaling and UV-B-mediated plant growth regulation. J. Integr. Plant Biol. 62, 1270–1292. doi: 10.1111/jipb.12932
Zhang, F., Huang, J., Guo, H., Yang, C., Li, Y., Shen, S., et al. (2022). OsRLCK160 contributes to flavonoid accumulation and UV-B tolerance by regulating OsbZIP48 in rice. Sci. China Life Sci. 65, 1380–1394. doi: 10.1007/s11427-021-2036-5
Zhang, Q., Yang, Y., Wu, J., Li, H., Li, Y., Li, Z., et al. (2024). Enhanced ultraviolet−B radiation suppresses magnaporthe oryzae infection and alleviates its damage to the photosynthesis of rice leaves. Phyton-International J. Exp. Bot. 93, 2613–2628. doi: 10.32604/phyton.2024.056014
Zhang, K., Yao, L., Meng, J., Tao, J. (2018). Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 634, 1326–1334. doi: 10.1016/j.scitotenv.2018.04.112
Zhang, X., Zhou, Y., Xu, T., Zong, S. (2024). Impact of climate change on the habitat suitability of monochamus saltuarius gebler (Coleoptera; cerambycidae) and its natural enemies in China. Forests 15 (1), 33. doi: 10.3390/f15010033
Zhao, G., Cui, X., Sun, J., Li, T., Wang, Q., Ye, X., et al. (2021). Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Indic. 132, 108256. doi: 10.1016/j.ecolind.2021.108256
Zhu, S., Sun, S., Zhao, W., Yang, X., Mao, H., Sheng, L., et al. (2024). Utilizing transcriptomics and proteomics to unravel key genes and proteins of Oryza sativa seedlings mediated by selenium in response to cadmium stress. BMC Plant Biol. 24, 360. doi: 10.1186/s12870-024-05076-7
Keywords: biomod2, climate change, potential geographic distributions, Oryza sativa, ensemble model, climate-suitable region
Citation: Wang R, Guo X, Song Y, Cai Y, Wu Y and Wang M (2025) Effects of ultraviolet radiation as a climate variable on the geographic distribution of Oryza sativa under climate change based on Biomod2. Front. Plant Sci. 16:1552770. doi: 10.3389/fpls.2025.1552770
Received: 29 December 2024; Accepted: 10 March 2025;
Published: 16 April 2025.
Edited by:
Julia Jessica Reimer, University of Applied Sciences Emden Leer, GermanyCopyright © 2025 Wang, Guo, Song, Cai, Wu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mingtian Wang, d2FuZ210MDUxNEAxNjMuY29t
†These authors have contributed equally to this work and share first authorship