 Xuexi Ma1,2,3,4
Xuexi Ma1,2,3,4 Lianlian Fan1,3,4
Lianlian Fan1,3,4 Meiniu Yang1,3
Meiniu Yang1,3 Jiangyue Li1,3Meng Yan2Ziyue Yang1,2
Jiangyue Li1,3Meng Yan2Ziyue Yang1,2 Xi Chen1,3Bo Zhang1
Xi Chen1,3Bo Zhang1 Yaoming Li1,3,4*
Yaoming Li1,3,4* Yingzhi Gao2,5*
Yingzhi Gao2,5*- 1Key Laboratory of Ecological Safety and Sustainable Development in Arid Lands, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi, China
- 2College of Grassland Science, Xinjiang Agricultural University, Key Laboratory of Grassland Resources and Ecology of Western Arid Desert Area of the Ministry of Education, Urumqi, China
- 3Chinese Academy of Sciences (CAS) Research Center for Ecology and Environment of Central Asia, Urumqi, China
- 4Bayinbuluk Alpine Grassland Observation and Research Station of Xinjiang, Bayinbuluk, Xinjiang, China
- 5Institute of Grassland Science, Northeast Normal University, Key Laboratory of Vegetation Ecology of the Ministry of Education, State Environmental Protection Key Laboratory of Wetland Ecology and Vegetation Restoration, Changchun, China
Global climate change and overgrazing have exacerbated shrub encroachment in arid and semi-arid grasslands and risking the stability and multifunctionality of grassland ecosystems. This shift between shrub and grassland raises a great concern to explicitly analyze the distribution of shrubs into arid and semi-arid grasslands and related environmental adaptation by using their nonstructural carbohydrates (NSCs) concentrations, and assess their allocation strategies and key determinants of these strategies. This research study was conducted in the Altai mountains of northwest of China and specifically focused on Spiraea, a dominant shrub in this region, and analyzed the NSCs component concentrations and their allocation in Spiraea leaves across different grassland types. The results showed significant variations in the NSCs concentrations, components, and allocation in Spiraea leaves across various grassland types, demonstrating higher overall values in meadows rather than grasslands (p < 0.05). Moreover, the total NSCs concentrations(the sum of soluble sugar and starch concentrations, and the ratio of soluble sugar to starch) exhibited consistent responses to environmental changes, and indicated increasing trends with elevation, soil water content (SWC), and soil total nitrogen concentrations (TN), while decreasing with mean annual temperature (MAT). Notably, MAT and SWC were emerged as the factors with the highest influence on NSCs concentrations and allocation in Spiraea leaves. These findings indicate that Spiraea in the Altai mountains can rapidly adapt to environmental changes across different grassland types by calibrating their NSCs concentrations and composition. These findings also present an insightful theoretical foundation for managing shrub-grassland encroachments in the Altai mountains region and other arid and semi-arid regions.
1 Introduction
Nonstructural carbohydrates (NSCs) are crucial energy sources and osmotic adjustment substances in plants that sustain plant growth and facilitate survival under stressful conditions (Hartmann and Trumbore, 2016; Yin et al., 2021)., NSCs include starch (energy storage) and soluble sugars (direct energy and osmotic adjustment) These two components can interconvert under specific environmental conditions to help plants adapt to diverse environments (McDowell, 2011; Zhang et al., 2023). For instance, under similar growth conditions, plants with higher NSCs concentrations tend to have higher chances of survival and better ability to do so (Blumstein et al., 2023; O’Brien et al., 2014). Therefore, NSCs storage can shield plants against carbon supply deficiencies, allowing them to cope with abiotic and biotic stresses. Consequently, NSCs storage is an important physiological indicator for assessing plant responses to environmental changes (Barker Plotkin et al., 2021; Martínez-Vilalta et al., 2016). Thus, explaining not only the concentrations of starch and soluble sugars within plants but also their allocation strategies is necessary to understand how plants adapt to diverse environmental conditions.
The factors influencing NSCs concentrations and allocation strategy include precipitation, temperature, and soil physicochemical properties (Guo et al., 2020; O’Brien et al., 2014; Zhang et al., 2024a). Increased drought intensity and duration correspond with a decline in the starch content in most woody plant organs and a rise in the concentration of soluble sugars, causing an increase in the ratio of soluble sugar to starch. Consequently, plants convert more starch into soluble sugars for osmotic adjustment, thereby maintaining cell osmotic pressure and ensuring plant growth and survival under drought conditions (He et al., 2020; Liu et al., 2019). Similarly, plants demonstrate analogous responsive characteristics in response to temperature stress, where high or low temperatures trigger starch hydrolase activity, prompting the conversion of starch into soluble sugars (Thalmann and Santelia, 2017; Yin and Zhang, 2016). In their study on alpine coniferous forests in the Qinghai–Tibet Plateau, Zhang et al. (2024a) argued that plant NSCs, especially those found in aboveground leaves, are mainly influenced by the combined effects of temperature and precipitation. Along with hydrothermal conditions, soil nutrient supply is a key factor affecting NSCs concentrations and allocation in plants. As a result of continuous climate changes and shifts in regional precipitation patterns, northwestern China is transitioning toward a warmer and wetter climate. In this context, soil nitrogen and phosphorus supply can promote plant growth and stressor resistance, thereby compensating for any adverse conditions such as drought and, subsequently, influencing NSCs concentrations and dynamics in plants (Zhang et al., 2024b; Zhou et al., 2011). Therefore, the concentration and allocation dynamics of plant NSCs demonstrate strong species- and region-specific characteristics.
The Altai mountains are a transitional zone between the Taiga biogeographical community and the arid desert biogeographical community. This adds to the significance of their grasslands as far as northern China is concerned. However, human activities such as overgrazing have contributed to severe degradation of the grassland ecosystem in the Altai mountains (Li et al., 2024). Moreover, the coverage and diversity of herbaceous plants in the grassland have steadily decreased. In contrast, the coverage of shrubs has rapidly increased, gradually replacing herbaceous plants as the dominant species in the region, causing grassland shrub encroachment (Ding and Eldridge, 2023; Moore et al., 2020; Ma and Gao, 2025). This has reduced the grassland ecosystem’s productivity, carbon sequestration capacity, nutrient cycling, and water-holding capacity, threatening its multifunctionality (Eldridge et al., 2011; Pierce et al., 2019). The shrub species of the genus Spiraea, with their developed root systems and better drought and nutrient-poor tolerance than those of herbaceous plants, are widely distributed in the Altai mountains grassland. They have become the dominant species in the region’s grassland shrub encroachment. However, the exact mechanisms underlying the adaptation of Spiraea to the Altai mountains’ diverse climatic conditions in terms of NSCs composition and allocation remain shrouded in uncertainty shrouded in concentration and their response to environmental gradients are poorly understood. Hence, the present study proposed two scientific hypotheses. First, a decrease in precipitation and temperature will cause a corresponding decrease in plant starch concentration and a corresponding increase in the ratio of soluble sugar to starch. Second, how do climatic and edaphic variables across grassland types influence the NSC allocation strategy of Spiraea, and which factors most strongly drive variation? To validate these hypotheses, this study selected the Altai mountains as the research area and focused on the dominant shrub species, Spiraea, as the subject of investigation. By systematically collecting samples of Spiraea from various grassland types and thoroughly analyzing the NSCs concentrations and proportions in their leaves, the study aimed to explore the NSCs concentrations and allocation patterns within Spiraea foliage under different environmental conditions. Further, it attempted to identify the key factors influencing NSCs in Spiraea leaves, thus presenting crucial scientific evidence for the stability and informed management of grassland ecosystems amid global climate change. This study is the first to quantify leaf NSC concentrations in Spiraea across Altai mountain grasslands, revealing how shrub encroachment influences ecosystem responses to climate change.
2 Materials and methods
2.1 Study area
The Altai mountains, which stretch nearly 1,200 km in a northwest direction along the borders of Russia, China, and Mongolia, reach a maximum elevation of 4,374 m. The Chinese segment of the Altai mountains (44°11’~46°20’N, 84°31’~90°00’E) is situated in the heart of the Eurasian continent. It occupies the middle southern slope of the mountain range, running in a northwest–southeast direction and gradually descending. This region is a significant climatic and natural boundary influenced by the Pacific and Atlantic Oceans that is marked by a continental temperate climate, boasting cold winters and abundant snowfall. The annual precipitation averages around 300 mm, demonstrating a vertical zonation pattern ranging from 100 mm in the lower mountainous regions to 600 mm in the higher alpine areas. Additionally, the annual mean temperature varies between -3.6°C and 1.8°C (Huang et al., 2015).
The Altai mountains region demonstrates complete vertical vegetation zonation, with the grassland zone primarily condensed at elevations between 800 and 1,800 m. Recently, the grasslands in this region have revealed a marked tendency toward shrub encroachment in response to global climate change and overgrazing. In this context, the dominant shrubs belong to the genus Spiraea (Spiraea salicifolia L.), including species such as Spiraea hypericifolia L., Spiraea media Schmidt, and Lonicera japonica Thunb (Ma et al., 2025).
2.2 Experimental design
In July 2023, we collected leaves from five healthy Spiraea plants across different grassland types in the Altai mountains (Figure 1). These grassland types included temperate steppe desert (TSD), constituting one site; temperate desert steppe (TDS), constituting three sites; temperate steppe (TS), constituting 10 sites; temperate meadow steppe (TMS), constituting five sites; and mountain meadow (MM), constituting four sites (Figure 1). The mean annual precipitation (MAP) gradient across the sampling sites ranged from 155 mm to 354 mm, whereas the mean annual temperature (MAT) gradient ranged from -2.92°C to 5.31°C. For each sampling site, a large plot of 100 m × 100 m was established, within which five small plots of 10 m × 10 m were set up following a five-point sampling method. Further, within each small plot, leaves from five healthy Spiraea plants were randomly collected between 11:00 and 16:00 to minimize diurnal variation in NSC concentrations and mixed to form one sample, with five replicate samples in total. We transported samples to the laboratory and fixed them at 105°C for 10 minutes. Subsequently, they were dried at 65°C to constant weight before being pulverized in a ball mill for future use.
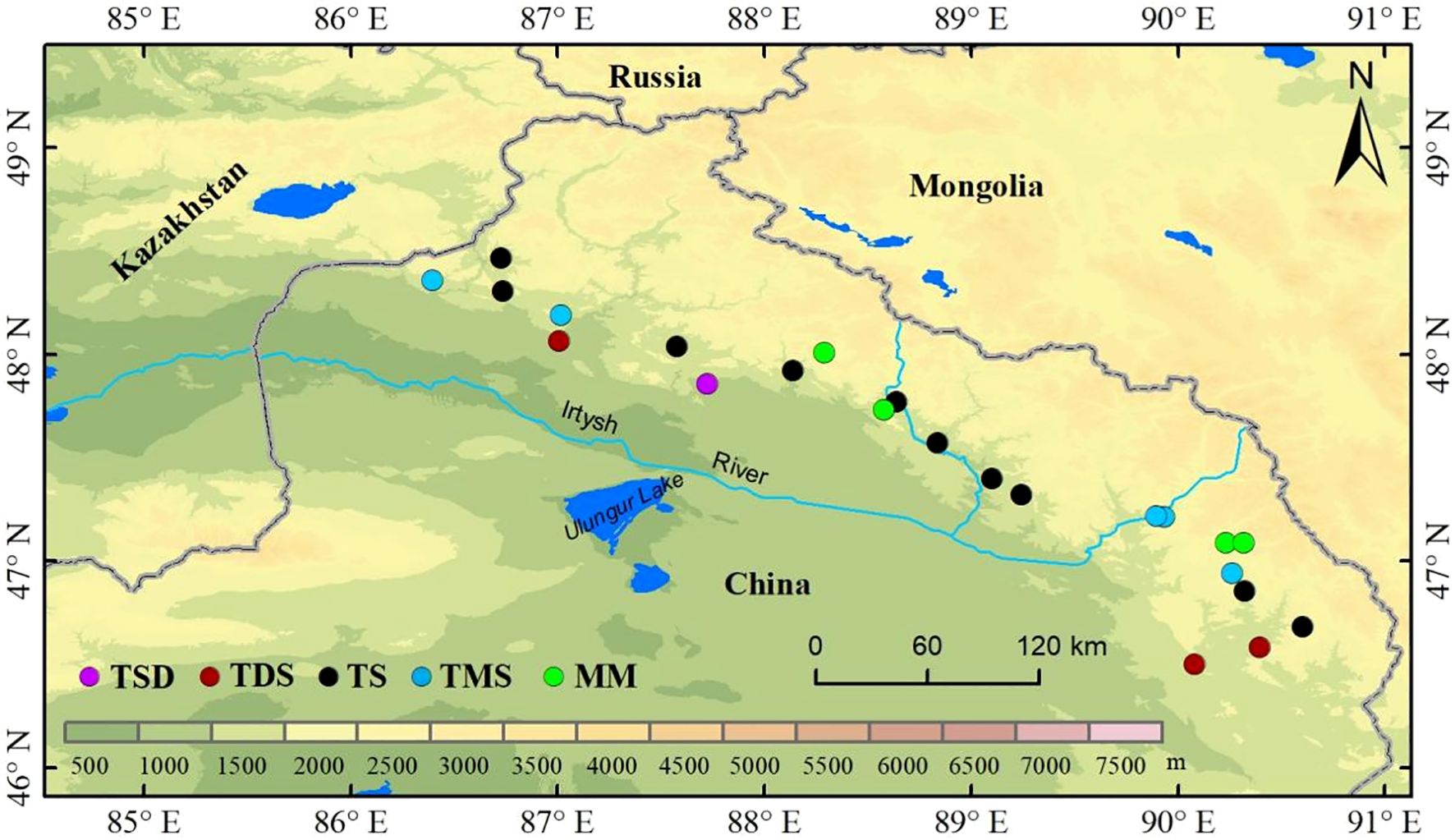
Figure 1. Distribution map of sample points.
Concurrently, soil samples were collected from the 0–10 cm layer using a soil auger within each small plot at each site, with five replicates per site. The collected soil samples were sieved through a 2 mm mesh to remove large debris or roots. Then, they were divided into two portions. While one was air-dried to determine soil organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), available phosphorus (AP), pH, and electrical conductivity (EC), the other was refrigerated at 4°C and analyzed for soil water content (SWC) and available nitrogen (AN) within one week.
2.3 Determination of NSCs concentrations
Primarily, NSCs comprise starch and total soluble sugars, as these two components account for over 90% of the total NSCs concentrations. Therefore, in the present study, the total NSCs concentrations were calculated as the sum of soluble sugars and starch—both of which were measured through the traditional anthrone–sulfuric acid method (Zhang et al., 2023). Using this method, 0.15 g of dried and powdered plant leaf samples were accurately weighed and placed in 10 mL of 80% absolute ethanol. Afterward, the mixture was extracted in a boiling water bath for 10 minutes. This mixture was then centrifuged at 4,000 r·min-1 for 10 minutes to collect the supernatant as the extract of soluble sugars. Subsequently, 10 mL of 30% (v/v) perchloric acid was added to the centrifuged precipitate, and the mixture was allowed to stand overnight. After accurate extraction in a water bath at 80°C for 10 minutes to ensure complete hydrolysis of starch, the mixture was cooled and centrifuged again at 4,000 r·-1 for 10 minutes. Following this, the supernatant was used to determine the starch concentration.
2.4 Collection and measurement of environmental factors
The gravimetric method was used to determine SWC. Under this method, fresh soil samples were dried in an oven at 105°C for 48 hours until a constant weight was achieved, followed by weighing. The pH value and EC were measured using the potentiometric method, with soil-to-water ratios of 1:2.5 and 1:5, respectively. Moreover, SOC was determined using the HCl titration-combustion method on a carbon-nitrogen analyzer (Multi 3100C/N, Analytik Jena AG, Germany). Furthermore, using a fully automated flow analyzer, the TN and TP concentrations in the soil were measured after digestion with concentrated sulfuric acid, perchloric acid, and hydrofluoric acid, and AP concentration was extracted with 0.5 mol L-1 NaHCO3 solution (Bran Luebbe, AA3, Germany). The Kjeldahl method was employed for TN determination, while the molybdenum-antimony anti-colorimetry method was used for both TP and AP. Additionally, the AN concentration in the soil was determined using the alkaline hydrolysis diffusion method (Yin et al., 2024).
2.5 Statistical analysis
To begin with, the obtained data were subjected to normality and homogeneity of variance tests. The differences in soluble sugars, starch, soluble sugar/starch ratio, and total NSCs among different grassland types were compared using a one-way analysis of variance (ANOVA), followed by the least significant difference (LSD) test. Additionally, the graphics were created using the ggplot2 package in R 4.2.1; Spearman correlation analysis and random forest modeling were conducted using the randomForest and linkET packages in R 4.1.3 to assess the impact of abiotic factors on soluble sugars, starch, soluble sugar/starch ratio, and total NSCs. Furthermore, regression analyses were performed between key abiotic factors and these NSCs components to identify potential abiotic factors influencing their concentrations. To explain the complex relationships between environmental factors and NSCs in Spiraea, a piecewise structural equation model (SEM) was constructed using the “piecewiseSEM” package in R 4.2.1, which was used to determine the direct and indirect effects of climatic factors and soil physicochemical properties on NSCs. Model evaluation was based on Fisher’s C significance (0.05 < p < 1) and the AIC, with stepwise refinement of the model as per the significance of path coefficients (Tian et al., 2022). Throughout the analysis and visualization process, various packages were utilized, including “dplyr,” “ggplot2,” “patchwork,” “tidyverse,” and “vegan.”
3 Results
3.1 Effects of different grassland types on physical and chemical properties of soil
Notable differences were observed in the soil physicochemical properties across grassland types. The SWC, SOC, TN, and AN levels were higher in TMS and MM soils than in TDS and TS soils (Figure 2, p < 0.05). Conversely, the soil pH and AP concentrations were significantly lower in TMS and MM soils than in TDS and TS soils (p < 0.05).
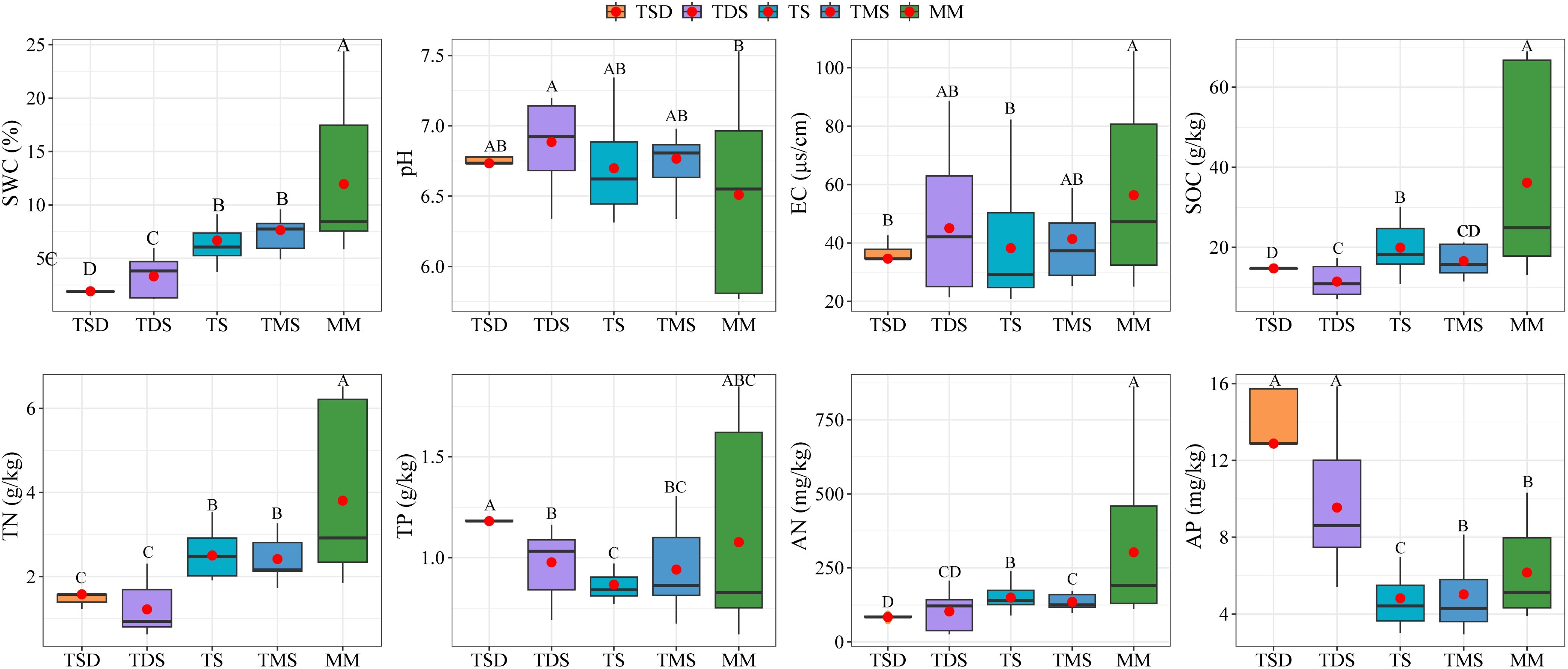
Figure 2. Effects of different grassland types on soil physicochemical properties; the different letters indicate significant differences between treatments at p < 0.05.
3.2 Characteristics of differences in NSCs concentrations of Spiraea across different grassland types
Significant differences were observed in the total soluble sugars, starch, total NSCs concentrations, and the ratio of soluble sugar to starch of Spiraea across different grassland types. The soluble sugar, starch, NSCs concentrations, and soluble sugar/starch ratio in Spiraea from TMS and MM soils were significantly higher than those from TDS and TS soils (Figure 3, p < 0.05). Further, the soluble sugar concentration in Spiraea from TSD soils was significantly lower than that from TMS and MM soils (p < 0.05), while no significant differences were found in starch and NSCs concentrations (p > 0.05).
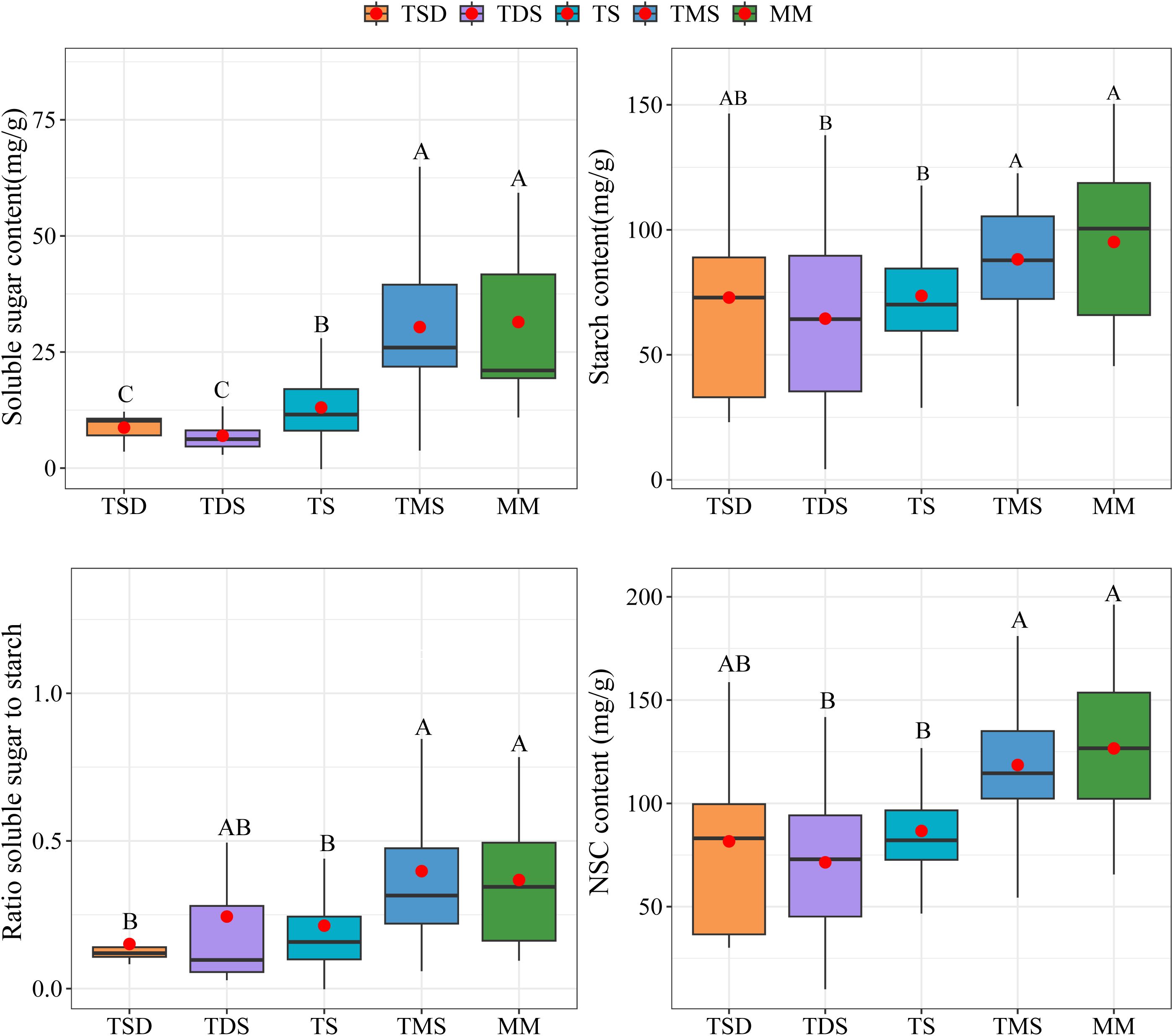
Figure 3. Differences in NSCs and their components of Spiraea across various grassland types; different letters indicate significant differences between treatments at p < 0.05.
3.3 Relationship between NSCs concentrations in Spiraea and environmental factors
The random forest analysis found that climate and soil environmental factors explained a significantly higher proportion of the variation in total soluble sugar concentration (37.95%) and total NSCs concentrations (27.99%) than starch concentration (2.84%) and the ratio of soluble sugar to starch (5.82%). As seen in Figure 4A, elevation, MAT, SWC, and TN were identified as the most important predictors influencing the variation in NSCs and their component concentrations in Spiraea leaves (Figure 4A). Moreover, as Figure 4B illustrates, NSCs and their individual component concentrations in Spiraea leaves significantly increased with increasing elevation, SWC, and TN while significantly decreasing with increasing MAT.

Figure 4. (A) Random forest analysis of key factors influencing the concentrations of NSCs and their components in Spiraea leaves; (B) Relationship between the concentrations of NSCs and their components in Spiraea leaves and environmental variables; here, SS represents total soluble sugars, ST represents the starch concentration in Spiraea leaves, and ELE represents elevation.
3.4 Key determinants influencing the NSCs concentrations in Spiraea
As Figure 5 illustrates, we employed piecewise SEM to analyze how climate, soil particle size, and physicochemical properties directly and indirectly influence the spatial variation of total NSCs in Spiraea. The results suggest that MAT, soil particle size, and physicochemical properties collectively explain 33% of the spatial variation in NSCs in Spiraea. Notably, MAT emerged as the primary factor influencing the spatial variation of NSCs, exerting a negative effect primarily through its indirect impacts on SWC, soil particle size, and physicochemical properties. Further, SWC had a secondary but significant impact on its spatial variation. In contrast, soil particle size exhibited the least influence.
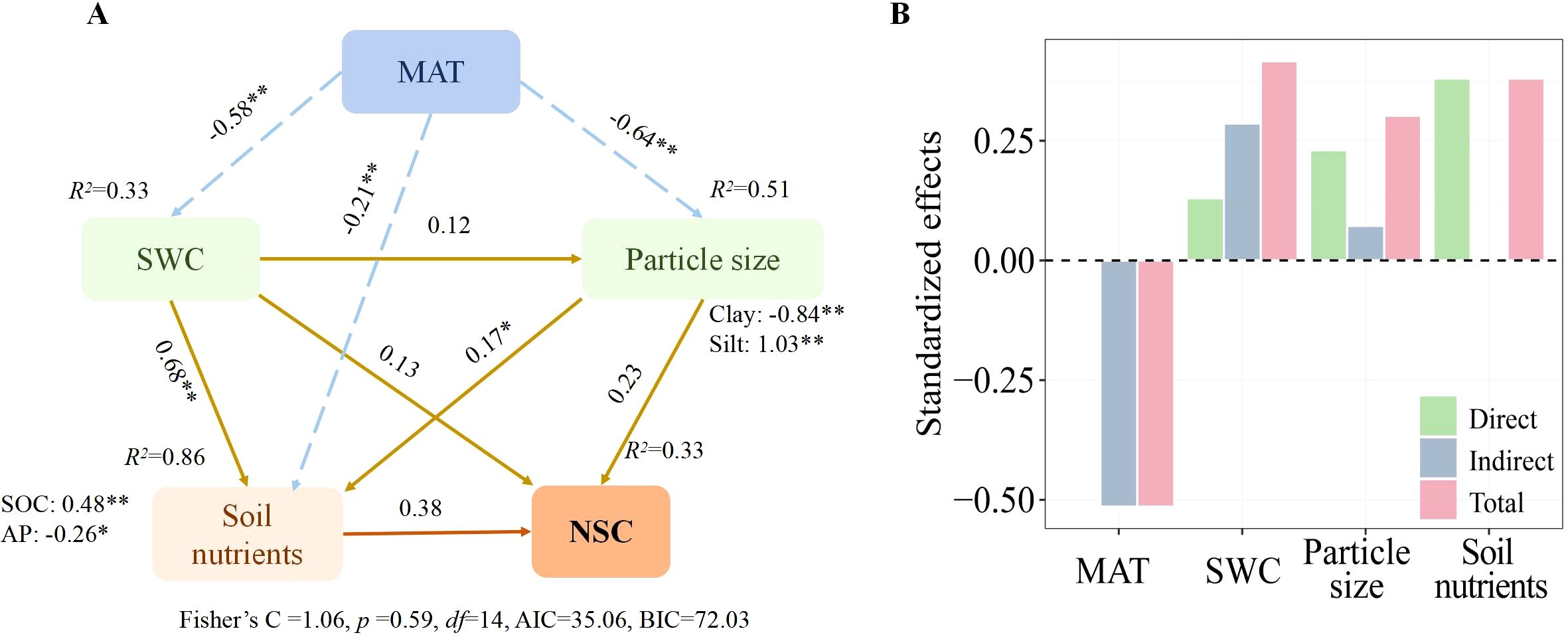
Figure 5. (A) A piecewise SEM calculating the direct and indirect effects of climatic factors, soil nutrients, SWC, and soil particle size on the NSCs in Spiraea; (B) The overall effects of all predictors on NSCs concentrations in Spiraea; here, the dashed lines indicate negative effects, while solid lines indicate positive effects; ** and * denote significance levels of p < 0.01 and p < 0.05, respectively.
4 Discussion
4.1 Influence of hydrothermal conditions on the NSCs concentrations of Spiraea
Hydrothermal conditions limit plant productivity and growth in various ecosystem types within arid regions, exerting direct and indirect influences on plant physiological metabolism (Dickman et al., 2019; Zhao et al., 2022). As organs highly sensitive to environmental changes, plant leaves can reflect adaptive responses through timely changes in NSC concentrations. This study revealed that MAT and SWC significantly impact the total NSCs, their individual component concentrations, and the ratio of soluble sugar to starch in different grassland types of the Altai Mountains, which aligns with our hypothesis. Under the higher SWC conditions, Spiraea can enhance its metabolic activities and carbon reserve synthesis by acquiring more water, thereby increasing the concentrations of soluble sugars and starch (O’Brien et al., 2014; Zhang et al., 2024a). Moreover, a meta-analysis indicates that NSCs accumulate under moderate water stress, whereas severe or prolonged drought may cause net carbon reserve losses (He et al., 2020). This suggests that along with altitudinal gradients, plants balance temperature and water availability. This trade-off in resource utilization determines the distribution and concentration variations of carbohydrates in Spiraea.
As the factor limiting plant growth in arid regions the most, water directly influences photosynthesis, as well as the synthesis and allocation of NSCs in plants (Adams et al., 2017; Sevanto et al., 2014). Precipitation in mountainous areas is more unevenly distributed than in plain ecosystems, leaving plants more susceptible to water limitations during their growth process. Subsequently, this affects photosynthetic carbon fixation. Generally, plants growing in arid environments retain high NSCs reserves and allocate a larger proportion of these reserves as soluble sugars, thus reducing water potential during turgor loss and promoting their survival during droughts (Wiley and Helliker, 2012). Our results indicate that the NSCs concentrations in Spiraea are influenced more by SWC than by precipitation, which contradicts both our hypothesis and previous findings. This discrepancy is caused by the region’s soil texture and topographical environment, where precipitation in mountainous areas is more vulnerable to runoff and less likely to infiltrate the soil. Notably, SWC is a crucial factor here as it is more accessible to plants. An increase in SWC corresponds with an increase in the total NSCs concentrations and its components in Spiraea plants. Under adequate water conditions, Spiraea efficiently fixes soluble sugars and starch through photosynthesis, likely as a strategy to endure harsh winter periods (Jang and Sheen, 1994; Yin et al., 2021).
Temperature exerts a more significant influence than SWC on the composition of NSCs in different grassland types of the Altai Mountains. This may be because of the regional climate, where the Altai Mountains, located at a higher latitude and in a mountainous area, receive more precipitation (approximately 300 mm) than other arid grasslands at similar latitudes. Further, the lower temperatures lead to slower water evaporation and higher water availability. Several studies have found that plant carbon allocation efficiency and growth rate are temperature-dependent, especially in high-latitude regions (García-Carreras et al., 2018). Moreover, soluble sugars, as important energy sources and osmotic adjustment substances in plants, amass in response to environmental stresses such as low temperatures, thereby enhancing the plant’s resistance (Rodríguez-Calcerrada et al., 2011; Klopotek and Kläring, 2014). Therefore, as temperatures decrease, Spiraea, under low-temperature stress, increases its NSCs concentrations, particularly soluble sugars, to elevate leaf osmotic pressure. Additionally, they secrete starch-hydrolyzing enzymes to hydrolyze part of the starch into soluble sugars, thus maintaining normal metabolic activities (Yin and Zhang, 2016; Michaletz, 2018; He et al., 2023).
4.2 Impact of grassland type on the NSCs concentrations of Spiraea
This study’s findings validate our hypothesis that different grassland types significantly impact the NSCs and their component concentrations in Spiraea. We found that the concentrations of soluble sugars and total NSCs in Spiraea from high-altitude MMs and TMSs are significantly higher than those in other grassland types. This variation may be because of the low-temperature conditions at higher altitudes, which slow down plant metabolic rates (Körner, 2007), reducing the rate of carbon reserve decomposition. Simultaneously, the higher SWC and nutrient supply (such as SOC and TN) enhance plant photosynthetic efficiency and promote carbohydrate accumulation (Hoch and Körner, 2009; Blumstein et al., 2023). Conversely, in low-altitude TDSs and TSDs, plants experience greater environmental stress due to higher temperatures and the scarcity of SWC and nutrients. Moreover, the more frequent utilization of their carbohydrates for maintaining basic metabolism and survival adaptations results in significantly lower NSCs concentrations, with the lowest soluble sugar/starch ratio (Moore et al., 2021; Ryhti et al., 2022). This suggests that changes in altitude, hydrothermal conditions, and soil nutrients jointly drive significant differences in the resource allocation strategies of Spiraea, gradually transitioning from a rapid growth mode to an adaptation mode focused on storage.
Differences in grassland types significantly alter soil physicochemical properties, indirectly influencing the variation in plant NSCs and their component concentrations (Zhang et al., 2023), with SOC and TN being especially important in this process. While SOC functions as a carbon source for soil microbial activity and plant metabolism that enhances the plant’s carbon fixation and storage capacity (Huang et al., 2024), nitrogen is a key limiting factor for plant growth in arid and semi-arid regions that promotes NSCs accumulation by enhancing plant nitrogen uptake and protein synthesis capabilities, which, in turn, increases enzyme activity related to photosynthesis and carbon metabolism. This promotes water utilization and transport, enhancing carbon utilization strategies (Schimel et al., 2004; Garnett et al., 2009; Wang et al., 2022). This was further confirmed in the present study, where SEM revealed that soil nitrogen significantly influences the NSCs concentrations in Spiraea that grows in the Altai Mountains. As soil TN content increases, there is a corresponding increase in plant soluble sugars, total NSCs, and the ratio of soluble sugar to starch. This trend reflects nitrogen supply, which alleviates limitation in plants, enhancing photosynthetic activity and promoting the accumulation of soluble sugars and other substances (Zhou et al., 2011; Xie et al., 2018). Furthermore, the availability of these nutrients is not only related to their concentration but also regulated by soil particle composition, such as the proportion of clay and sand particles. A high proportion of clay particles aids in nutrient retention, whereas a high concentration of sand particles may lead to nutrient loss, particularly in mountainous regions (Wang et al., 2020). Therefore, soil nutrients directly influence plant physiological metabolic processes and resource allocation strategies, driving the dynamic changes in NSCs and their components in Spiraea.
5 Conclusion
Spiraea in the Altai Mountains can adapt to diverse grassland environments by regulating the concentrations and composition of NSCs. This study found that SWC, MAT, soil TNC, and altitude can directly or indirectly affect NSCs concentrations and distribution. Among these factors, MAT and SWC most significantly influence the NSCs concentrations and components of Spiraea in different grassland types. As MAT decreases and SWC and soil TNC increase, the soluble sugars, starch, and total NSCs concentrations in Spiraea leaves show an upward trend. Thus, these findings offer physiological and ecological insights into why Spiraea has become a dominant species in the grassland shrub encroachment of this region, offering theoretical support for scientifically managing grassland shrub encroachment in this area. However, further research is necessary to investigate the NSCs allocation strategies of Spiraea above and below ground, as well as their variation patterns during different growth stages.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Author contributions
XM: Formal Analysis, Investigation, Methodology, Software, Validation, Writing – original draft, Writing – review & editing. LF: Conceptualization, Investigation, Writing – review & editing. MY: Investigation, Software, Visualization, Writing – original draft. JL: Formal Analysis, Visualization, Writing – original draft. MY: Data curation, Visualization, Writing – original draft. ZY: Investigation, Methodology, Writing – original draft. XC: Conceptualization, Funding acquisition, Writing – review & editing. BZ: Conceptualization, Writing – review & editing, Resources. YL: Conceptualization, Methodology, Supervision, Writing – review & editing. YG: Conceptualization, Formal Analysis, Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by the National Natural Science Foundation of China (W2412123), the Tianshan Talent Project of Xinjiang Uygur Autonomous Region, China (2022TSYCLJ0056), the Third Xinjiang Scientific Expedition Program (2021xjkk0603) and the Xinjiang Agricultural University Graduate School-level Scientific Research and Innovation Program (XJAUGRI2024001).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adams, H. D., Zeppel, M. J., Anderegg, W. R., Hartmann, H., Landhäusser, S. M., Tissue, D. T., et al. (2017). A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 1, 1285–1291. doi: 10.1038/s41559-017-0248-x
Barker Plotkin, A., Blumstein, M., Laflower, D., Pasquarella, V. J., Chandler, J. L., Elkinton, J. S., et al. (2021). Defoliated trees die below a critical threshold of stored carbon. Funct. Ecol. 35, 2156–2167. doi: 10.1111/1365-2435.13891
Blumstein, M., Gersony, J., Martínez-Vilalta, J., and Sala, A. (2023). Global variation in nonstructural carbohydrate stores in response to climate. Global Change Biol. 29, 1854–1869. doi: 10.1111/gcb.16573
Dickman, L. T., McDowell, N. G., Grossiord, C., Collins, A. D., Wolfe, B. T., Detto, M., et al. (2019). Homoeostatic maintenance of nonstructural carbohydrates during the 2015–2016 El Niño drought across a tropical forest precipitation gradient. Plant Cell Environ. 42, 1705–1714. doi: 10.1111/pce.13501
Ding, J. and Eldridge, D. (2023). The success of woody plant removal depends on encroachment stage and plant traits. Nat. Plants 9, 58–67. doi: 10.1038/s41477-022-01307-7
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Roger, E., Reynolds, J. F., and Whitford, W. G. (2011). Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol. Lett. 14, 709–722. doi: 10.1111/j.1461-0248.2011.01630.x
García-Carreras, B., Sal, S., Padfield, D., Kontopoulos, D. G., Bestion, E., Schaum, C. E., et al. (2018). Role of carbon allocation efficiency in the temperature dependence of autotroph growth rates. Proc. Natl. Acad. Sci. 115, E7361–E7368. doi: 10.1073/pnas.1800222115
Garnett, T., Conn, V., and Kaiser Brent, N. (2009). Root based approaches to improving nitrogen use efficiency in plants. Plant Cell Environ. 32, 1272–1283. doi: 10.1111/j.1365-3040.2009.02011.x
Guo, J. S., Gear, L., Hultine, K. R., Koch, G. W., and Ogle, K. (2020). Non-structural carbohydrate dynamics associated with antecedent stem water potential and air temperature in a dominant desert shrub. Plant Cell Environ. 43, 1467–1483. doi: 10.1111/pce.13749
Hartmann, H. and Trumbore, S. (2016). Understanding the roles of nonstructural carbohydrates in forest trees–from what we can measure to what we want to know. New Phytol. 211, 386–403. doi: 10.1111/nph.2016.211.issue-2
He, R., Shi, H., Hu, M., Zhou, Q., Zhang, Q., and Dang, H. (2023). Divergent effects of warming on nonstructural carbohydrates in woody plants: a meta-analysis. Physiologia Plantarum 175, e14117. doi: 10.1111/ppl.v175.6
He, W., Liu, H., Qi, Y., Liu, F., and Zhu, X. (2020). Patterns in nonstructural carbohydrate contents at the tree organ level in response to drought duration. Global Change Biol. 26, 3627–3638. doi: 10.1111/gcb.15078
Hoch, G. and Körner, C. (2009). Growth and carbon relations of tree line forming conifers at constant vs. variable low temperatures. J. Ecol. 97, 57–66. doi: 10.1111/j.1365-2745.2008.01447.x
Huang, L., Gao, Y., Li, Y., Zhang, T., Hu, D., and Wang, L. (2015). Growth of Siberia larch in the middle east of Altay Mountains and its response to climate change. Arid Land Geography 38, 1169–1178. doi: 10.13826/j.cnki.cn65-1103/x.2015.06.010
Huang, Q., Wang, B., Shen, J., Xu, F., Li, N., Jia, P., et al. (2024). Shifts in C-degradation genes and microbial metabolic activity with vegetation types affected the surface soil organic carbon pool. Soil Biol. Biochem. 192, 109371. doi: 10.1016/j.soilbio.2024.109371
Jang, J. C. and Sheen, J. (1994). Sugar sensing in higher plants. Plant Cell 6, 1665–1679. doi: 10.1016/S1360-1385(97)89545-3
Klopotek, Y. and Kläring, H. P. (2014). Accumulation and remobilisation of sugar and starch in the leaves of young tomato plants in response to temperature. Scientia Hortic. 180, 262–267. doi: 10.1016/j.scienta.2014.10.036
Körner, C. (2007). The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 22, 569–574. doi: 10.1016/j.tree.2007.09.006
Li, Y., Zhang, Z., Zhou, X., Gao, M., Duan, J., Xue, Y., et al. (2024). Transformation and mechanisms of climate wet/dry change on the northern Tibetan Plateau under global warming: A perspective from paleoclimatology. Sci. China Earth Sci. 67, 1932–1951. doi: 10.1007/s11430-023-1260-6
Liu, H., Shangguan, H., Zhou, M., Airebule, P., Zhao, P., He, W., et al. (2019). Differentiated responses of nonstructural carbohydrate allocation to climatic dryness and drought events in the Inner Asian arid timberline. Agric. For. Meteorology 271, 355–361. doi: 10.1016/j.agrformet.2019.03.008
Ma, X., Fan, L., Fakher, A., Li, Y., Mao, J., Yang, M., et al. (2025). Shrub encroachment: A catalyst for enhanced soil nutrients storage in the Altai Mountains. Plants 14, 623. doi: 10.3390/plants14040623
Ma, X. and Gao, Y. (2025). Impact of shrub encroachment on soil hydrological processes in grassland. Acta Prataculturae Sin. 34, 212–222. doi: 10.11686/cyxb2024201
Martínez-Vilalta, J., Sala, A., Asensio, D., Galiano, L., Hoch, G., Palacio, S., et al. (2016). Dynamics of non-structural carbohydrates in terrestrial plants: a global synthesis. Ecol. Monogr. 86, 495–516. doi: 10.1002/ecm.1231
McDowell, N. G. (2011). Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant physiol. 155, 1051–1059. doi: 10.1104/pp.110.170704
Michaletz, S. T. (2018). Evaluating the kinetic basis of plant growth from organs to ecosystems. New Phytol. 219, 37–44. doi: 10.1111/nph.2018.219.issue-1
Moore, T. E., Jones, C. S., Chong, C., and Schlichting, C. D. (2020). Impact of rainfall seasonality on intraspecific trait variation in a shrub from a Mediterranean climate. Funct. Ecol. 34, 865–876. doi: 10.1111/1365-2435.13533
Moore, C. E., Meacham-Hensold, K., Lemonnier, P., Slattery, R. A., Benjamin, C., Bernacchi, C. J., et al. (2021). The effect of increasing temperature on crop photosynthesis: from enzymes to ecosystems. J. Exp. Bot. 72, 2822–2844. doi: 10.1093/jxb/erab090
O’Brien, M., Leuzinger, S., Philipson, C., Tay, J., and Hector, A. (2014). Drought survival of tropical tree seedlings enhanced by nonstructural carbohydrate levels. Nat. Climate Change 4, 710–714. doi: 10.1038/nclimate2281
Pierce, N. A., Archer, S. R., and Bestelmeyer, B. T. (2019). Competition suppresses shrubs during early, but not late, stages of arid grassland-shrubland state transition. Funct. Ecol. 33, 1480–1490. doi: 10.1111/1365-2435.13336
Rodríguez-Calcerrada, J., Shahin, O., Del Rey, M. D. C., and Rambal, S. (2011). Opposite changes in leaf dark respiration and soluble sugars with drought in two Mediterranean oaks. Funct. Plant Biol. 38, 1004–1015. doi: 10.1071/FP11135
Ryhti, K., Schiestl-Aalto, P., Tang, Y., Rinne-Garmston, K. T., Ding, Y., Pumpanen, J., et al. (2022). Effects of variable temperature and moisture conditions on respiration and nonstructural carbohydrate dynamics of tree roots. Agric. For. Meteorology 323, 109040. doi: 10.1016/j.agrformet.2022.109040
Schimel, J. P., Bilbrough, C., and Welker, J. A. (2004). Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities. Soil Biol. Biochem. 36, 217–227. doi: 10.1016/j.soilbio.2003.09.008
Sevanto, S., McDowell, N. G., Dickman, L. T., Pangle, R., and Pockman, W. T. (2014). How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell Environ. 37, 153–161. doi: 10.1111/pce.2014.37.issue-1
Thalmann, M. and Santelia, D. (2017). Starch as a determinant of plant fitness under abiotic stress. New Phytol. 214, 943–951. doi: 10.1111/nph.2017.214.issue-3
Tian, P., Zhao, X., Liu, S., Sun, Z., Jing, Y., and Wang, Q. (2022). Soil microbial respiration in forest ecosystems along a north-south transect of eastern China: Evidence from laboratory experiments. Catena 211, 105980. doi: 10.1016/j.catena.2021.105980
Wang, J., Qu, F., Liang, J., Yang, M., and Hu, X. (2022). Bacillus velezensis SX13 promoted cucumber growth and production by accelerating the absorption of nutrients and increasing plant photosynthetic metabolism. Scientia Hortic. 301, 111151. doi: 10.1016/j.scienta.2022.111151
Wang, R., Wu, H., Sardans, J., Li, T., Liu, H., Peñuelas, J., et al. (2020). Carbon storage and plant-soil linkages among soil aggregates as affected by nitrogen enrichment and mowing management in a meadow grassland. Plant Soil 457, 407–420. doi: 10.1007/s11104-020-04749-0
Wiley, E. and Helliker, B. (2012). A re-evaluation of carbon storage in trees lends greater support for carbon limitation to growth. New Phytol. 195, 285–289. doi: 10.1111/j.1469-8137.2012.04180.x
Xie, H., Yu, M., and Cheng, X. (2018). Leaf non-structural carbohydrate allocation and C: N: P stoichiometry in response to light acclimation in seedlings of two subtropical shade-tolerant tree species. Plant Physiol. Biochem. 124, 146–154. doi: 10.1016/j.plaphy.2018.01.013
Yin, B., Li, J., Zhang, Q., Wu, N., Zhang, J., Rong, X., et al. (2021). Freeze-thaw cycles change the physiological sensitivity of Syntrichia caninervis to snow cover. J. Plant Physiol. 266, 153528. doi: 10.1016/j.jplph.2021.153528
Yin, B., Li, J., Zhang, Q., Zhang, S., Liu, Z., Zhou, X., et al. (2024). Snow depth has greater influence on moss biocrusts’ soil multifunctionality than the number of freeze-thaw cycles. Appl. Soil Ecol. 199, 105420. doi: 10.1016/j.apsoil.2024.105420
Yin, B. F. and Zhang, Y. M. (2016). Physiological regulation of Syntrichia caninervis Mitt. in different microhabitats during periods of snow in the Gurbantünggüt Desert, northwestern China. J. Plant Physiol. 194, 13–22. doi: 10.1016/j.jplph.2016.01.015
Zhang, P., Ding, J., Wang, Q., McDowell, N. G., Kong, D., Tong, Y., et al. (2024a). Contrasting coordination of non-structural carbohydrates with leaf and root economic strategies of alpine coniferous forests. New Phytol. 243, 580–590. doi: 10.1111/nph.v243.2
Zhang, S., Yang, A., Zang, Y., Guo, K., Zhou, X., Rong, X., et al. (2024b). Slope position affects nonstructural carbohydrate allocation strategies in different types of biological soil crusts in the Gurbantunggut Desert. Plant Soil, 1–14. doi: 10.1007/s11104-024-06951-w
Zhang, S., Zhang, Q., Li, Y., Lu, Y., Zhou, X., Yin, B., et al. (2023). Shrubs have a greater influence on the nonstructural carbohydrates of desert mosses along precipitation decreased. Environ. Exp. Bot. 216, 105530. doi: 10.1016/j.envexpbot.2023.105530
Zhao, L., Chen, H., Chen, B., Wang, Y., and Sun, H. (2022). Drought shapes photosynthetic production traits and water use traits along with their relationships with leaves of typical desert shrubs in Qaidam. Forests 13, 1652. doi: 10.3390/f13101652
Keywords: grassland shrub encroachment, Altai Mountains, nonstructural carbohydrates, soluble sugars, starch
Citation: Ma X, Fan L, Yang M, Li J, Yan M, Yang Z, Chen X, Zhang B, Li Y and Gao Y (2025) Allocation strategy of nonstructural carbohydrates in Spiraea L. across different grassland types in the Altai Mountains. Front. Plant Sci. 16:1562363. doi: 10.3389/fpls.2025.1562363
Received: 17 January 2025; Accepted: 15 May 2025;
Published: 06 June 2025.
Edited by:
Xiali Guo, Guangxi University, ChinaReviewed by:
Guilin Wu, Chinese Academy of Sciences (CAS), ChinaShujun Zhang, Xinjiang University, China
Copyright © 2025 Ma, Fan, Yang, Li, Yan, Yang, Chen, Zhang, Li and Gao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yaoming Li, bHltQG1zLnhqYi5hYy5jbg==; Yingzhi Gao, Z2FveXoxMDhAbmVudS5lZHUuY24=