 Pedro M. P. Correia
Pedro M. P. Correia Xinyi Dong
Xinyi Dong Mengming Chen
Mengming Chen Anton Frisgaard Nørrevang
Anton Frisgaard Nørrevang Guangbin Luo
Guangbin Luo Michael Palmgren
Michael Palmgren- NovoCrops Center, Department of Plant and Environmental Sciences, University of Copenhagen, Frederiksberg, Denmark
The domestication of grasses has historically favored annual species due to their rapid growth and suitability for crop rotation; however, such crops rely heavily on human input. In contrast, perennial grasses, which live for multiple years, offer significant environmental benefits, such as improved soil health and natural resilience to biotic and abiotic stress, but have not yet been domesticated. Gene editing of yield-related genes presents an opportunity to improve yield stability in perennial cereal crops. However, this process typically requires transformation to introduce gene-editing tools, and many perennial grasses are recalcitrant to traditional in vitro transformation. Alternative in planta transformation methods have recently emerged, offering simpler, faster, and more genotype-independent approaches. These methods bypass the need for tissue culture and could potentially be used to transform recalcitrant plants more efficiently. In this review, we evaluate the potential of in planta transformation methods for developing perennial cereal crops and advocate for exploring the role of such crops in sustainable agriculture.
Perennial grain crops as a sustainable alternative to annual cropping systems
Cereal grain crops, which are members of the grass family (Poaceae), include several widely cultivated species, such as wheat (Triticum aestivum), maize (Zea mays), rice (Oryza sativa), and barley (Hordeum vulgare), that provide the bulk of calories in the human diet (FAO, 2017). Annual grasses were domesticated over thousands of generations, resulting in the selection and retention of gene variants underlying desirable traits. Although the selected traits benefit agricultural production, artificial selection often works against natural selection, resulting in domesticated crops with reduced fitness, an inability to survive outside their cultivation areas, and a reliance on intensive chemical management (Chen et al., 2015; Meyer et al., 2012). Annual cropping systems involve replanting crops each year, which can negatively affect soil health. Frequent tillage contributes to soil degradation, resulting in erosion, greenhouse gas emissions, and decreased soil fertility (Crews et al., 2018; Olsson et al., 2024). The environmental effects of annual cropping underscore the urgent need for a shift toward more sustainable practices.
Perennial agriculture offers numerous advantages over traditional annual cropping systems. Perennial crops have extensive root systems that reduce soil erosion and benefit soil health by improving soil structure and increasing organic matter content. Perennial crops also enhance water use efficiency deep in the soil, making the plants more resilient to drought and reducing the need for irrigation (Figure 1). Additionally, perennial systems sequester more carbon, contributing to the mitigation of climate change. Economically, these practices reduce the need for annual replanting, which lowers labor and input costs while offering more stable income streams for farmers (Chapman et al., 2022; Crews et al., 2018; Olsson et al., 2024; Paul et al., 2024). By improving soil health, water use efficiency, and biodiversity, perennial agriculture presents a sustainable alternative that could help address food security and environmental sustainability issues (Figure 1).
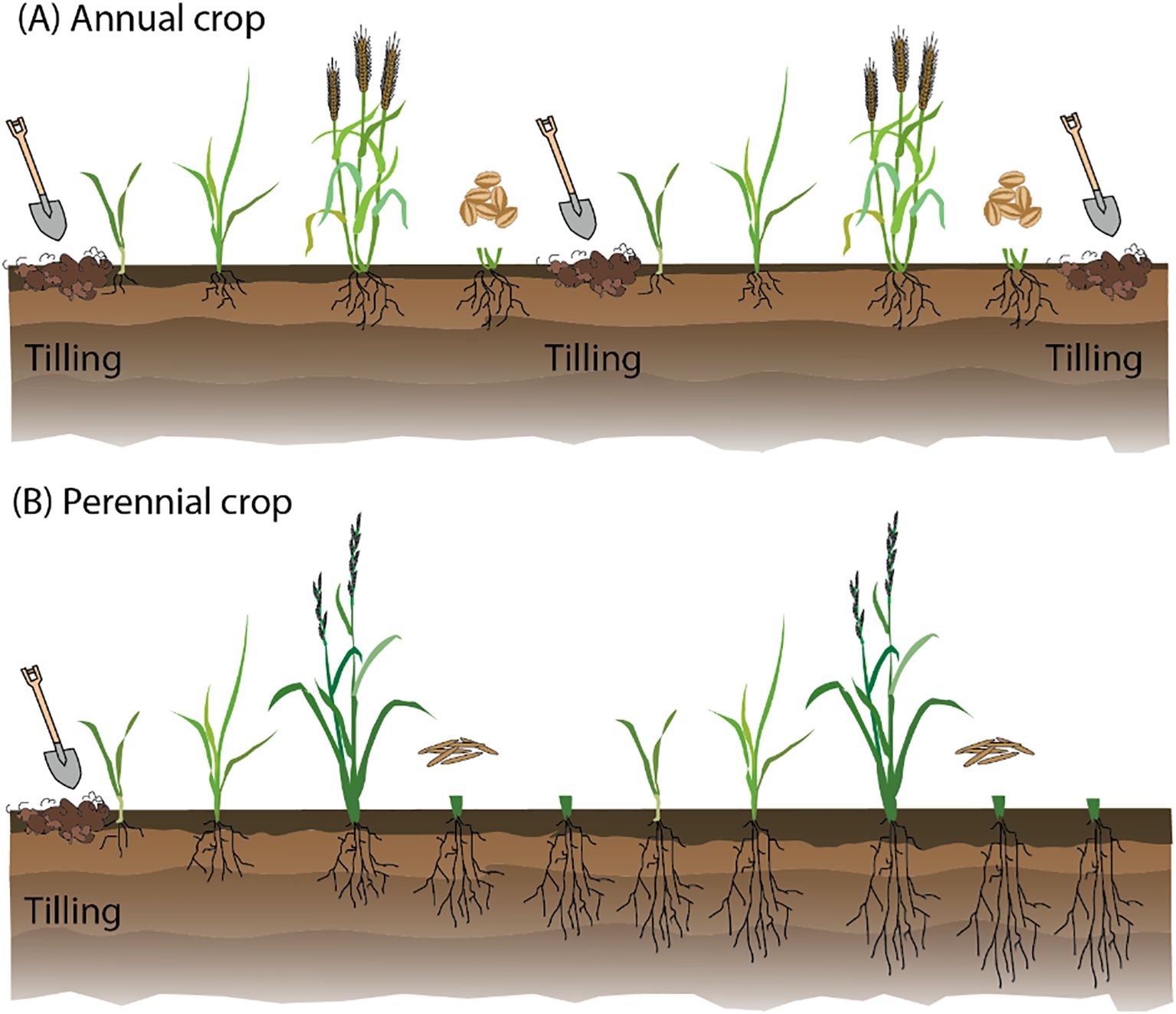
Figure 1. Comparison of annual and perennial cropping systems. (A) Annual crops live for a single season, requiring annual sowing and tilling, which promotes soil erosion, depletion of organic matter, and CO2 loss. These crops are highly dependent on human input for fertilization and protection against abiotic and biotic stress. (B) Perennial crops are sown only in the first year. These plants develop deep and extensive root systems that help preserve soil organic matter, reduce erosion, and improve overall soil health, and offer a sustainable alternative to annual crops.
Accelerating the domestication of perennial grain crops through genome editing
Efforts are underway to develop perennial cereal crops through wide hybridization—i.e., crossing existing annual crops with perennial relatives—and by traditional breeding of wild perennial species. The improvement of perennial intermediate wheatgrass (Thinopyrum intermedium) through traditional breeding (Bajgain et al., 2022) and the development of perennial rice PR23 from the hybridization of Asian cultivated rice (Oryza sativa ssp. indica) with its African wild perennial relative (Oryza longistaminata) (Zhang et al., 2022) are two successful examples of progress toward developing high-yielding perennial cereal crops.
However, high levels of ploidy and heterozygosity in perennial grasses mask alleles that are potentially useful for domestication, particularly in the case of recessive loss-of-function mutations (Østerberg et al., 2017), thereby complicating traditional breeding strategies. Targeted mutagenesis has emerged as a promising alternative tool for accelerating the domestication of new perennial crops. Leveraging an unprecedented understanding of crop domestication processes, the first step in these strategies involves precisely mutating a few genes using novel genome-editing technologies (Østerberg et al., 2017; Chapman et al., 2022; Luo et al., 2022).
There are over 7,000 perennial grass species that remain largely unexplored for domestication purposes (Frawley et al., 2020). Resources such as the Perennial Agriculture Project Global Inventory (PAPGI; http://www.tropicos.org/Project/PAPGI) provide information for assessing the potential utility of undomesticated perennial species (Ciotir et al., 2019). Allied with new targeted mutagenesis methods such as genome editing, these resources could prompt the cultivation of a range of novel perennial alternatives to annual crops. Such practices may become common if, for example, the EU lifts regulations on gene-editing tools such as clustered regularly interspaced palindromic repeats/CRISPR-associated protein9 (CRISPR/Cas9).
Biological challenges in transforming perennial grasses
Genome editing has unlocked the ability to make precise and predictable changes to domestication genes, thereby paving the way for the accelerated domestication of wild perennial grain crops. Yet, the ability to transform and regenerate the target plant remains a prerequisite for successful genome editing. Although the technology for creating transgenic plants is decades old, the transformation of many plant species remains challenging, with high efficiency limited to a few species and even fewer cultivars within these species (Ahmar et al., 2023).
To date, genome-editing components have been delivered into annual cereal crops primarily through Agrobacterium-mediated or biolistic transformation of suitable explants, typically immature embryos. Numerous studies have explored strategies to enhance transformation efficiency and develop alternative tissue culture protocols (Ahmar et al., 2023). Several improved protocols have been published, mainly focusing on optimizing media compositions and refining procedures for Agrobacterium-mediated or biolistic transformation (Matres et al., 2021).
Many perennial grasses require exposure to cold (i.e., vernalization) or accumulated days of warmth for the vegetative to floral transition (Lundgren and Des Marais, 2020), which complicates access to reproductive tissues, such as immature embryos, for transformation. Another hurdle associated with the domestication of perennial grasses is their widespread self-incompatibility, which prevents self-fertilization and demands outcrossing (Baumann et al., 2000). Combined with high levels of ploidy and heterozygosity, this adds variability, complicating the optimization of tissue culture media compositions and transformation procedures. Self-incompatibility can also reduce seed set due to pollen abortion (DeHaan and Van Tassel, 2014), thereby affecting the availability of immature embryos as explants for transformation. Perennials often produce vegetatively propagated organs such as rhizomes and bulbous structures (Chapman et al., 2022); however, whether these organs can serve as substitutes for immature embryos in transformation has not been explored. These biological and technical constraints highlight the need for alternative strategies for transformation that bypass the reliance on immature embryos and tissue culture.
In planta transformation
In planta transformation, also known as in situ transformation, encompasses a diverse array of techniques aimed at directly integrating foreign DNA or editing a plant’s genome, followed by regenerating the cells into a whole plant. These approaches can utilize bacteria (e.g., Agrobacterium strains), viruses, or physical methods (e.g., particle bombardment) to deliver genetic material to cells (e.g., meristem tissues, reproductive tissues, embryos) and integrate it into their genomes (Bélanger et al., 2024). Transformed plants may consequently produce transformed seeds, enabling stable transformation or editing events in the next generation. Here we focus on in planta transformation approaches that have the potential to transform perennial grass species that are recalcitrant to in vitro transformation. We identify the bottlenecks of these methods and assess their potential for genome editing and domestication of perennial cereal crops (Figure 2, Table 1).
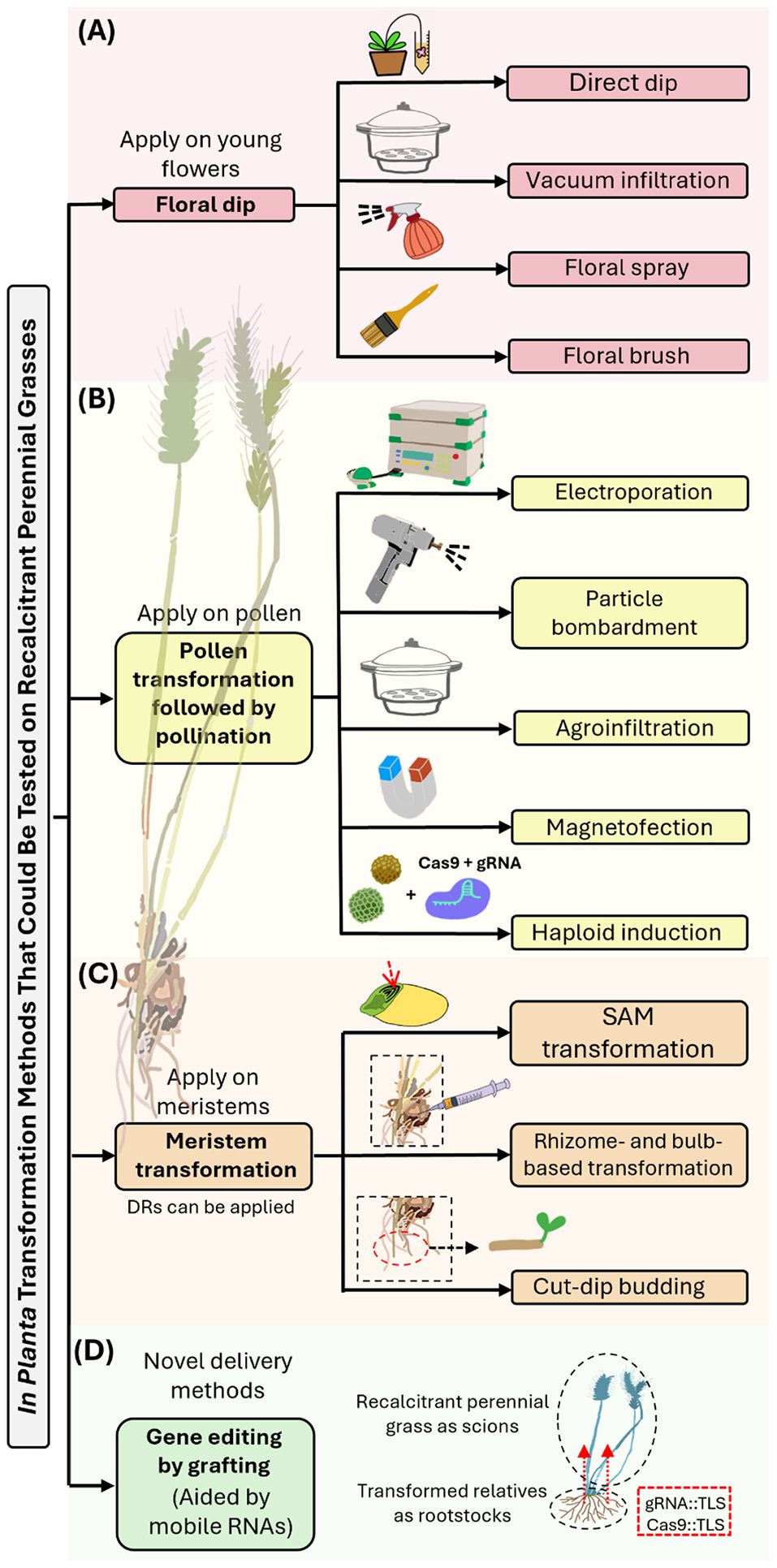
Figure 2. Overview of in planta transformation methods that could be applied to perennial grasses. (A) Floral dip transformation involves dipping young flowers in an Agrobacterium tumefaciens suspension, leading to transgenic seeds via natural fertilization. Methods include direct dip, vacuum infiltration, floral spray, and brush application. While these methods are effective in some annual grasses (Singh and Kumar, 2022), their efficiency in perennial grasses is limited by the unsynchronized anthesis and outcrossing nature of these plants. (B) Pollen transformation is used to deliver gene-editing tools into pollen grains, which are in turn used for pollination. Methods to overcome the challenges of pollen transformation include electroporation, particle bombardment, Agrobacterium infiltration, and magnetofection. Establishing efficient pollen transformation methods and using methods such as haploid induction editing (Kelliher et al., 2019) could facilitate the development of perennial grain crops. (C) Meristem transformation involves direct transformation into shoot meristems, which are composed of embryonic-type cells that divide to form new cells and organs. Methods include Agrobacterium tumefaciens-mediated transformation or bombardment to transform the exposed meristem tissues of embryos, seedlings, or mature plants. Targeting the cell layer that will develop into germ cells from the SAMs of mature embryos using CRISPR/Cas9 can bypass the need for tissue culture and be genotype-independent (Hamada et al., 2018, 2017; Liu et al., 2021; Luo et al., 2023; Tezuka et al., 2024), making it a potential method for perennial grain crops. Methods utilizing Agrobacterium to transform vegetatively propagated organs such as rhizomes and bulbous structures offer new avenues for the transformation of perennial grasses via these organs (Cao et al., 2023; Mei et al., 2024). Direct meristem induction by expressing developmental regulators (DRs) such as Wus2 and Bbm promotes embryo formation and can enhance transformation efficiency and regeneration speed (Lowe et al., 2018; Wang et al., 2023). (D) Novel delivery methods mediated by mobile RNAs, which carry genome-editing tools across plant tissues (Yang et al., 2023), could potentially be employed for targeted heritable gene editing in perennial grass crops.
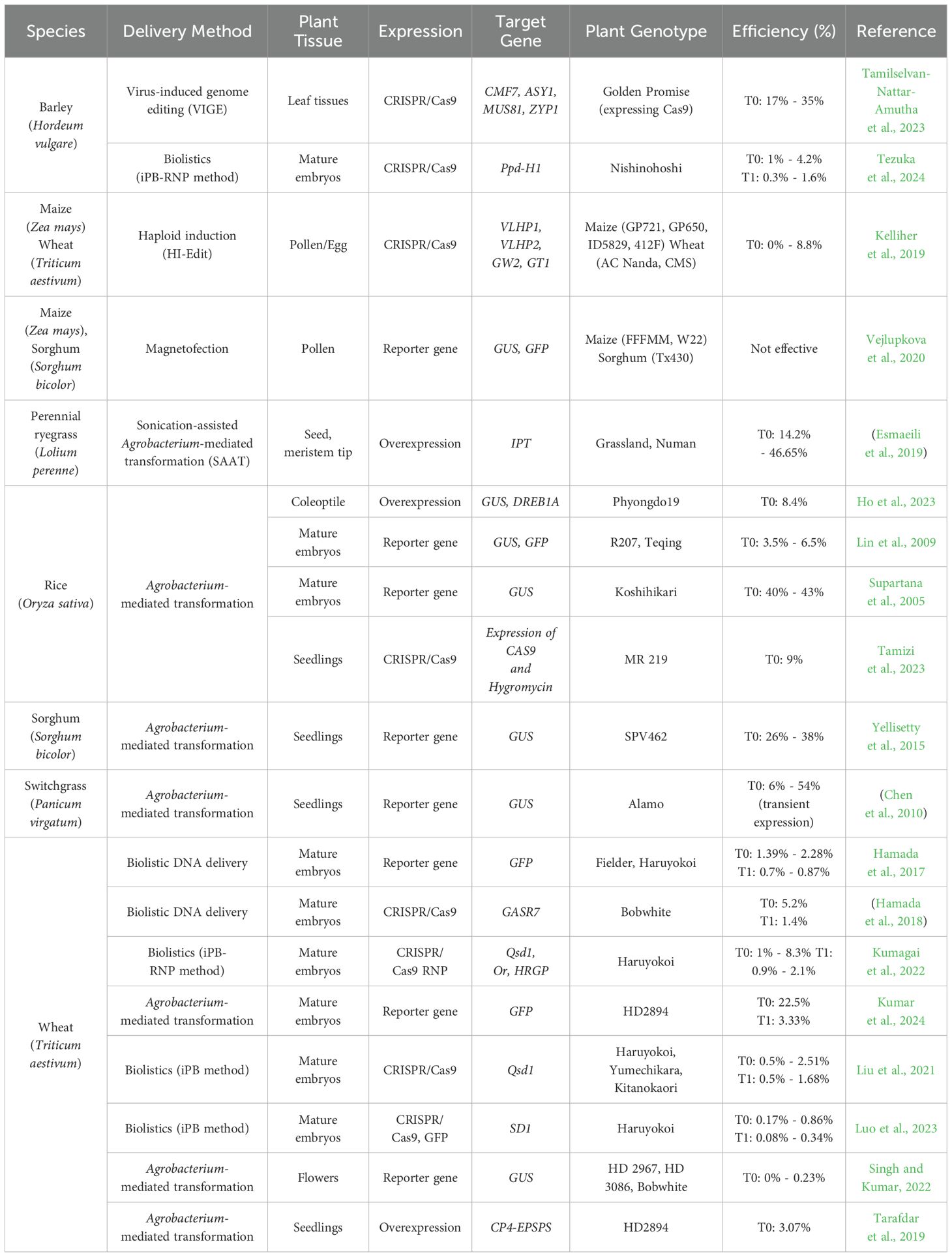
Table 1. Overview of in planta transformation in monocot grass species.
Direct transformation into meristems
A major difference between annuals and perennials is that perennials have growing points (meristems) that remain indeterminate, allowing them to retain the ability to give rise to new tissues or organs after the first growing season. These indeterminate meristems are often located underground in bulbs, rhizomes, tubers, or corms, where they maintain vegetative growth and can undergo a developmental transition to generate a new plant.
In planta transformation of the indeterminate meristems of perennial grain crops may target the cell layer of the mature embryo that will develop into germ cells from the shoot apical meristem (SAM). CRISPR/Cas9 components, such as DNA vectors (Hamada et al., 2018, 2017; Liu et al., 2021, Table 1), ribonucleoprotein (RNP) (Kumagai et al., 2022, Table 1), and double-stranded DNA (dsDNA) donors (Luo et al., 2023, Table 1), can be coated onto gold particles for bombardment and delivery to a specific cell layer. This goal was first achieved in wheat (Hamada et al., 2017) and more recently in barley (Tezuka et al., 2024) (Table 1). However, the small size of seeds and their respective SAMs can make the delivery of CRISPR components challenging. Directly transforming embryonic cells with mobile Cas9 and single-guide RNAs (sgRNAs) capable of intercellular movement may increase the number of target stem cells in the meristem that undergo gene editing. This approach bypasses the need for tissue culture and is genotype-independent, but its potential for transforming perennial grasses remains to be tested (Figure 2).
Recent developments in nanoparticle-based delivery systems offer a promising alternative to particle bombardment (Wang et al., 2019). Nanomaterials such as carbon nanotubes, mesoporous silica nanoparticles, and lipid-based nanocarriers can also be explored for their ability to traverse the plant cell wall and deliver CRISPR/Cas9 components (Lowry et al., 2024; Wang et al., 2019). Key challenges include understanding how nanoparticle physicochemical properties, such as size, shape, surface charge, and aspect ratio can influence their ability to penetrate plant tissues and reach target cells (Hofmann et al., 2020). This approach bypasses the need for tissue culture and is genotype-independent, but its potential for transforming perennial grasses remains to be tested (Figure 2).
Mechanically injured embryos from mature seeds and SAMs from young seedlings have been used for transformation by imbibing the wounded plant material in an Agrobacterium solution. This technique has been used to transform rice (Supartana et al., 2005; Arockiasamy and Ignacimuthu, 2007; Lin et al., 2009; Ho et al., 2023; Tamizi et al., 2023; Sundararajan et al., 2023), sorghum (Sorghum bicolor) (Yellisetty et al., 2015), wheat (Kumar et al., 2024; Tarafdar et al., 2019), and perennial cultivars of ryegrass (Lolium perenne) (Esmaeili et al., 2019), and for transient transformation of perennial switchgrass (Panicum virgatum) (Chen et al., 2010) (Table 1). A modified approach for wheat uses embryos excised from mature seeds, which are centrifuged together with Agrobacterium, resulting in the direct transformation of the SAM (Ye et al., 2022). This method has been used for several plant species, but its potential for transforming perennial grasses has not been tested. One concern is that chimerism, caused by non-uniform transformation, can complicate the transmission of mutations to the progeny (Zlobin et al., 2020; Ye et al., 2022).
Recently, Mei et al. (2024) used the regenerative activity-dependent in planta injection delivery (RAPID) method to successfully transform several dicot species. They directly injected A. tumefaciens into the lower excised ends of stem segments of sweet potato (Ipomoea batatas) and bayhops (Ipomoea pes-caprae), as well as beneath the skin of potato (Solanum tuberosum) tubers. Transgenic progeny were obtained via regeneration and vegetative propagation at transformation frequencies of 12.5% to 40% (Mei et al., 2024). The RAPID system allows for direct gene transfer into regenerative plant tissues such as stem segments, tubers, rhizomes, and bulbs. This approach depends on the strong ability of transformed plants to undergo vegetative propagation, and further optimization of herbicide-based selection is needed to reduce chimerism (Mei et al., 2024; Zhong et al., 2024). The system has been used in sweet potato, potato, bayhops, and Panax notoginseng, providing a robust platform for tissue culture–free transformation. Since perennials often produce vegetatively propagated organs such as rhizomes and bulbous structures, they might also prove to be transformable using this strategy (Figure 2) However, this potential remains to be confirmed in monocot species, which differ significantly in their developmental and physiological responses.
Another method, the “cut-dip budding delivery system,” utilizes Agrobacterium rhizogenes to induce and transform hairy roots from the cut sites of explants (Cao et al., 2023). After generating transformed hairy roots with shoot-forming ability, transformed plants can be regenerated. The method involves infecting root segments by immersing them in an A. rhizogenes suspension (Cao et al., 2023). This technique has been successfully applied in a number of dicot species. Broader application to monocots might be facilitated by the use of disarmed Agrobacterium strains and by excision-based removal of integrated oncogenes (Cao et al., 2023). A similar transformation system remains to be tested in vegetatively propagated organs of perennial grasses (Figure 2).
De novo induction of meristems
Meristem identity is, in part, dictated by developmental regulators (DRs). DRs work in conjunction with plant growth regulators, particularly the plant hormones cytokinin and auxin, to establish and maintain meristem identity. The expression of specific DRs in plant somatic cells can induce other developmental programs. In monocots, Wuschel2 (Wus2) and Baby Boom (Bbm) promote somatic cells to form embryos that develop into whole plants (Lowe et al., 2018; Wang et al., 2023). By expressing Bbm and Wus2, transgenic monocot plants were successfully recovered using genotypes or explant types that were otherwise recalcitrant to genetic transformation (Wang et al., 2023). Ectopic expression of the maize Bbm and Wus2 genes in rice, sugarcane (Saccharum officinarum), sorghum, and perennial switchgrass had a similar effect, suggesting a conserved function among monocot species (Lowe et al., 2016; Xu et al., 2022). Additional DRs, such as GROWTH-REGULATING FACTOR 4 (GRF4), GRF-INTERACTING FACTOR 1 (GIF1) chimera, and WUSCHEL RELATED HOMEOBOX 5 (WOX5) have shown potential for increasing transformation efficiency and the speed of regeneration (Debernardi et al., 2020; Wang et al., 2022; Yang et al., 2024). The use of DRs might expand the application of Agrobacterium infection of meristems to more plant species (Lian et al., 2022; Maher et al., 2020), including perennial grasses (Figure 2).
Mobile RNAs as carriers of genome-editing tools
In plants, messenger RNAs (mRNAs) can move to neighboring cells via plasmodesmata and over long distances through the vascular system to regulate various biological processes in target organs (Kitagawa et al., 2024). The first evidence of RNA mobility emerged from studies on RNA viruses, whose movement proteins enable their intercellular movement from an infected cell to neighboring cells, allowing systemic virus spread (Deom et al., 1987; Wolf et al., 1989). Subsequent investigations revealed a more intricate mechanism that involves transfer RNA-like structures (TLSs) at the 3′-ends of RNAs (Zhang et al., 2009, 2016). Such TLSs are abundant in transcripts found in the phloem sap and serve as mobility signals. In transgenic Arabidopsis (Arabidopsis thaliana) lines, co-transcription of mRNA with TLSs triggers the systemic movement of mRNA between roots and shoots (Heeney and Frank, 2023; Kehr et al., 2022). Remarkably, the transported mRNA components are translated into functional proteins in the receiving cells (Zhang et al., 2016).
Building on this foundational work, the mobility of target RNA has been exploited for genome editing. For example, sgRNAs can be mobilized to the shoot apex, as demonstrated using viral vectors to transform transgenic Arabidopsis plants expressing Cas9. By fusing sgRNAs with mobile Flowering Locus T (FT) transcripts or TLS sequences, heritable gene editing was enhanced (Ellison et al., 2020). In a more recent study, Cas9 RNA and gRNAs were tagged with TLS motifs, allowing both types of RNA to be mobilized across graft junctions. The mobile transcripts efficiently moved from the transgenic rootstock to the wild-type scion, resulting in targeted heritable gene editing in the scion (Yang et al., 2023). The Grafting-Based Gene Editing approach offers a transgene-free alternative for genome editing by delivering mobile gene-editing signals across graft unions. While this technique has been successfully demonstrated in Arabidopsis and Brassica rapa, its broader application is currently constrained by graft incompatibility, particularly in monocot crops (Yang et al., 2023). However, the recent development of a micrografting method for monocots (Reeves et al., 2022) presents an opportunity to achieve transgene-free targeted gene editing in major staple crops and potentially in new perennial crops (Figure 2). To overcome transformation limitations in recalcitrant perennial species, this strategy could use model lines with well-established transformation protocols as rootstocks. For instance, transgenic cereal grain crops such as wheat, transformable wheat cultivar Fielder (Hayta et al., 2019), which have robust transformation systems could serve as donor rootstocks for delivering mobile CRISPR/Cas components to grafted scions. Recent research has shed light on the molecular basis of graft formation, particularly the genetic control of graft attachment and vascular reconnection (Feng et al., 2024; Notaguchi et al., 2008). These discoveries may lead to the emergence of new graftable combinations, and engineering transgenic rootstocks to enhance grafting compatibility could facilitate genetic editing through grafting junctions.
Pollen transformation
Gene-edited plants have been obtained by delivering gene-editing tools into pollen grains (Bélanger et al., 2024; Toda et al., 2023). Here, the pollen grains are transformed and subsequently used to pollinate the recipient egg in vivo, resulting in nonchimeric transformation. Major challenges of this method include the thick cell wall of the pollen grain and the release of nuclease enzymes during pollen germination, as these factors hinder pollen grain transformation and the integration of exogenous DNA, respectively (Eapen, 2011). To overcome these obstacles, various methods could be utilized, including electroporation (Obermeyer and Weisenseel, 1995), particle bombardment (Touraev et al., 1997), Agrobacterium infiltration (Tjokrokusumo et al., 2000), and magnetofection, which uses magnetic force to enhance gene delivery (Zhao et al., 2017) (Figure 2). These techniques have been applied with varying degrees of success across different annual grass species such as maize and sorghum (Eapen, 2011; Toda et al., 2023), but with inconsistent results reported between different laboratories (Vejlupkova et al., 2020; Zhao et al., 2017; Table 1). Some grass species have mechanisms that enhance outcrossing by increasing the probability of pollen dispersal from one plant and its subsequent receipt by an unrelated plant (David and Pham, 1993). Establishing an efficient pollen transformation method for outcrossing perennial grasses could be advantageous (Figure 2).
A method involving haploid induction editing (Hi-Edit) technology has been used to edit elite lines of maize and wheat (Kelliher et al., 2019; Table 1). Here the paternal or maternal plant is a transformable cultivar that has been stably transformed with Cas9 and gRNA. The transformed plant is used as a pollen donor. However, the donor plant contains a mutation in MATRILINEAL (MATL) that eliminates the donor plant’s genome. After fertilization, the Cas9 and gRNA are expressed from the donor plant’s sperm or egg cell, editing the recipient’s chromosome and resulting in the elimination of the donor plant’s chromosome. This results in editing without the need for transformation and the production of a transgene-free edited elite crop (Kelliher et al., 2019). Using this approach, already established transformation systems could be used to edit a recalcitrant species such as a perennial grass (Figure 2). Hi-Edit has been used to deliver genome-editing components into maize and Arabidopsis, reducing chimerism and simplifying the production of edited lines. While efficient, improvements in haploid-doubling protocols are required to enhance their applicability (Kelliher et al., 2019).
Virus-mediated transformation
RNA viruses infect plants and move systemically within the plant. Tobacco rattle virus (TRV), potato virus X (PVX), and barley stripe mosaic virus (BSMV) have been used as vectors to introduce RNA sequences into plants without the need for transformation or regeneration (Awan et al., 2023; Tamilselvan-Nattar-Amutha et al., 2023). However, because Cas9 exceeds the cargo capacity of most viruses, it is usually only feasible to deliver sgRNAs, which requires that the plant host is already a transgenic Cas9-expressing line (Ellison et al., 2020; Liu et al., 2024, 2022, 2023). Endonucleases that are smaller and more compact may serve as alternatives to Cas9. In a recent study, a transposase-associated TnpB endonuclease was packaged with a guide RNA in the RNA virus TRV. Following infection of Arabidopsis with the engineered virus, editing was achieved in a single step (Weiss et al., 2025). This finding suggests the potential of using virus-induced genome editing to transform recalcitrant perennial grasses.
Genotype-independent fast transformation
Zhong et al. (2024) introduced Genotype-independent Fast Transformation (GiFT) as a rapid in planta Agrobacterium-mediated transformation system for soybean (Glycine max). The GiFT method uses wounded germinated seeds as explants, followed by a brief liquid culture phase under sublethal herbicide selection and direct soil transplantation, during which continued in planta selection ensures the preferential regeneration of transgenic shoots. This approach enabled the recovery of healthy, non-chimeric T0 plants within approximately 35 days without extensive tissue culture. GiFT demonstrated high transformation frequencies across a diverse range of elite and recalcitrant soybean varieties and was further validated for CRISPR-Cas12a-mediated genome editing applications. The general applicability of this novel in planta transformation method remains to be tested.
Conclusion
Perennial grasses are typically recalcitrant to transformation using traditional in vitro methods. More direct in planta transformation protocols have recently been shown to be successful for the transformation of several dicots and annual grasses, but whether these methods can be applied to perennial grasses remains to be determined. We hope that this review will inspire researchers focused on developing sustainable agricultural practices to test whether in planta transformation methods can be used to overcome the transformation and genome-editing bottlenecks associated with the improvement of perennial grain crops.
Author contributions
PC: Data curation, Writing – review & editing, Writing – original draft, Investigation. XD: Data curation, Investigation, Writing – review & editing. MC: Writing – review & editing, Data curation, Investigation. AN: Investigation, Data curation, Writing – review & editing. GL: Writing – review & editing, Data curation, Investigation. MP: Writing – review & editing, Supervision, Conceptualization, Funding acquisition, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors are grateful to the Novo Nordisk Foundation (NovoCrops NNF19OC005658; MP), Innovation Fund Denmark (DEEPROOTS #0224-0004313 and PERENNIAL # 079-00008B; MP), and the Villum Foundation (Villum Experiment #57907; PC) for their support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahmar, S., Hensel, G., and Gruszka, D. (2023). CRISPR/Cas9-mediated genome editing techniques and new breeding strategies in cereals – current status, improvements, and perspectives. Biotechnol. Adv. 69, 108248. doi: 10.1016/j.bioteChadv.2023.108248
Arockiasamy, S. and Ignacimuthu, S. (2007). Regeneration of transgenic plants from two indica rice (Oryza sativa L.) cultivars using shoot apex explants. Plant Cell Rep. 26, 1745–1753. doi: 10.1007/s00299-007-0377-9
Awan, M. J. A., Akram, A., Amin, I., and Mansoor, S. (2023). Viral vectors as carriers of genome-editing reagents. Trends Plant Sci. 28, 981–983. doi: 10.1016/j.tplants.2023.05.010
Bajgain, P., Crain, J. L., Cattani, D. J., Larson, S. R., Altendorf, K. R., Anderson, J. A., et al (2022). “Breeding Intermediate Wheatgrass for Grain Production,” in Plant Breeding Reviews. Ed Goldman, I. (Hoboken, NJ: Wiley). doi: 10.1002/9781119874157.ch3
Baumann, U., Bian, X., and Langridge, P. (2000). Self-incompatibility in the grasses. Ann. Bot. 85, 203–209. doi: 10.1006/anbo.1999.1056
Bélanger, J. G., Copley, T. R., Hoyos-Villegas, V., Charron, J.-B., and O’Donoughue, L. (2024). A comprehensive review of in planta stable transformation strategies. Plant Methods 20, 79. doi: 10.1186/s13007-024-01200-8
Cao, X., Xie, H., Song, M., Lu, J., Ma, P., Huang, B., et al. (2023). Cut–dip–budding delivery system enables genetic modifications in plants without tissue culture. Innovation 4, 100345. doi: 10.1016/j.xinn.2022.100345
Chapman, E. A., Thomsen, H. C., Tulloch, S., Correia, P. M. P., Luo, G., Najafi, J., et al. (2022). Perennials as future grain crops: opportunities and challenges. Front. Plant Sci. 13. doi: 10.3389/FPLS.2022.898769/XML/NLM
Chen, X., Equi, R., Baxter, H., Berk, K., Han, J., Agarwal, S., et al. (2010). A high-throughput transient gene expression system for switchgrass (Panicum virgatum L.) seedlings. Biotechnol. Biofuels 3, 9. doi: 10.1186/1754-6834-3-9
Chen, Y. H., Gols, R., and Benrey, B. (2015). Crop domestication and its impact on naturally selected trophic interactions. Annu. Rev. Entomol. 60, 35–58. doi: 10.1146/annurev-ento-010814-020601
Ciotir, C., Applequist, W., Crews, T. E., Cristea, N., DeHaan, L. R., Frawley, E., et al (2019). Building a botanical foundation for perennial agriculture: Global inventory of wild, perennial herbaceous Fabaceae species. Plants People Planet 1, 375–386. doi: 10.1002/ppp3.37
Crews, T. E., Carton, W., and Olsson, L. (2018). Is the future of agriculture perennial? Imperatives and opportunities to reinvent agriculture by shifting from annual monocultures to perennial polycultures. Global Sustainability 1, e11. doi: 10.1017/sus.2018.11
David, J. L. and Pham, J. (1993). Rapid changes in pollen production in experimental outcrossing populations of wheat. J. Evol. Biol. 6, 659–676. doi: 10.1046/j.1420-9101.1993.6050659.x
Debernardi, J. M., Tricoli, D. M., Ercoli, M. F., Hayta, S., Ronald, P., Palatnik, J. F., et al. (2020). A GRF–GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nat. Biotechnol. 38, 1274–1279. doi: 10.1038/s41587-020-0703-0
DeHaan, L. R. and Van Tassel, D. L. (2014). Useful insights from evolutionary biology for developing perennial grain crops 1. Am. J. Bot. 101, 1801–1819. doi: 10.3732/ajb.1400084
Deom, C. M., Oliver, M. J., and Beachy, R. N. (1987). The 30-kilodalton gene product of tobacco mosaic virus potentiates virus movement. Science 237, 389–394. doi: 10.1126/science.237.4813.389
Eapen, S. (2011). Pollen grains as a target for introduction of foreign genes into plants: an assessment. Physiol. Mol. Biol. Plants. 17, 1–8. doi: 10.1007/s12298-010-0042-6
Ellison, E. E., Nagalakshmi, U., Gamo, M. E., Huang, P., Dinesh-Kumar, S., and Voytas, D. F. (2020). Multiplexed heritable gene editing using RNA viruses and mobile single guide RNAs. Nat. Plants 6, 620–624. doi: 10.1038/s41477-020-0670-y
Esmaeili, S., Salehi, H., Khosh-Khui, M., Niazi, A., Tohidfar, M., and Aram, F. (2019). Isopentenyl transferase (IPT) gene transfer to perennial ryegrass through sonication-assisted agrobacterium-mediated transformation (SAAT), vacuum and heat treatment. Mol. Biotechnol. 61, 332–344. doi: 10.1007/s12033-019-00165-7
FAO (2017). The future of food and agriculture – Trends and challenges (Rome: Food and Agriculture Organization of the United Nations).
Feng, M., Augstein, F., Kareem, A., and Melnyk, C. W. (2024). Plant grafting: Molecular mechanisms and applications. Mol. Plant 17, 75–91. doi: 10.1016/j.molp.2023.12.006
Frawley, E. S., Ciotir, C., Micke, B., Rubin, M. J., and Miller, A. J. (2020). An ethnobotanical study of the genus elymus. Econ. Bot. 74, 159–177. doi: 10.1007/s12231-020-09494-0
Hamada, H., Linghu, Q., Nagira, Y., Miki, R., Taoka, N., and Imai, R. (2017). An in planta biolistic method for stable wheat transformation. Sci. Rep. 7, 11443. doi: 10.1038/s41598-017-11936-0
Hamada, H., Liu, Y., Nagira, Y., Miki, R., Taoka, N., and Imai, R. (2018). Biolistic-delivery-based transient CRISPR/Cas9 expression enables in planta genome editing in wheat. Sci. Rep. 8, 14422. doi: 10.1038/s41598-018-32714-6
Hayta, S., Smedley, M. A., Demir, S. U., Blundell, R., Hinchliffe, A., Atkinson, N., et al. (2019). An efficient and reproducible Agrobacterium-mediated transformation method for hexaploid wheat (Triticum aestivum L.). Plant Methods 15, 1–15. doi: 10.1186/S13007-019-0503-Z
Heeney, M. and Frank, M. H. (2023). The mRNA mobileome: challenges and opportunities for deciphering signals from the noise. Plant Cell 35, 1817–1833. doi: 10.1093/plcell/koad063
Ho, U. H., Song, S. R., Choe, Y. I., Pak, M. H., Kim, M. H., Kim, K., et al. (2023). Development of Agrobacterium-mediated in planta transformation protocol through coleoptile in rice. Transgenic Res. 32, 339–349. doi: 10.1007/s11248-023-00355-9
Hofmann, T., Lowry, G. V., Ghoshal, S., Tufenkji, N., Brambilla, D., Dutcher, J. R., et al. (2020). Technology readiness and overcoming barriers to sustainably implement nanotechnology-enabled plant agriculture. Nat. Food 2020 1:7 1, 416–425. doi: 10.1038/s43016-020-0110-1
Kehr, J., Morris, R. J., and Kragler, F.. (2022). Long-Distance Transported RNAs: From Identity to Function. Annu. Rev. Plant Biol. 73, 457–474. doi: 10.1146/annurev-arplant-070121-033601
Kelliher, T., Starr, D., Su, X., Tang, G., Chen, Z., Carter, J., et al. (2019). One-step genome editing of elite crop germplasm during haploid induction. Nat. Biotechnol. 37, 287–292. doi: 10.1038/s41587-019-0038-x
Kitagawa, M., Tran, T. M., and Jackson, D. (2024). Traveling with purpose: cell-to-cell transport of plant mRNAs. Trends Cell Biol. 34, 48–57. doi: 10.1016/j.tcb.2023.05.010
Kumagai, Y., Liu, Y., Hamada, H., Luo, W., Zhu, J., and Kuroki, M. (2022). Introduction of a second “Green Revolution” mutation into wheat via in planta CRISPR/Cas9 delivery. Plant Physiol. 188, 1838–1842. doi: 10.1093/plphys/kiab570
Kumar, S., Vishwakarma, H., Ghosh, G., Singh, J., and Padaria, J. C. (2024). In planta transformation in wheat: an improved protocol to develop wheat transformants. Mol. Biol. Rep. 51, 407. doi: 10.1007/s11033-024-09333-8
Lian, Z., Nguyen, C. D., Liu, L., Wang, G., Chen, J., and Wang, S. (2022). Application of developmental regulators to improve in planta or in vitro transformation in plants. Plant Biotechnol. J. 20, 1622–1635. doi: 10.1111/pbi.13837
Lin, J., Zhou, B., Yang, Y., Mei, J., Zhao, X., Guo, X., et al. (2009). Piercing and vacuum infiltration of the mature embryo: a simplified method for Agrobacterium-mediated transformation of indica rice. Plant Cell Rep. 28, 1065–1074. doi: 10.1007/s00299-009-0706-2
Liu, D., Ellison, E. E., Myers, E. A., Donahue, L. I., Xuan, S., Swanson, R., et al. (2024). Heritable gene editing in tomato through viral delivery of isopentenyl transferase and single-guide RNAs to latent axillary meristematic cells. Proc. Natl. Acad. Sci. 121. doi: 10.1073/pnas.2406486121
Liu, Y., Luo, W., Linghu, Q., Abe, F., Hisano, H., Sato, K., et al. (2021). In planta genome editing in commercial wheat varieties. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.648841
Liu, D., Xuan, S., Prichard, L. E., Donahue, L. I., Pan, C., Nagalakshmi, U., et al. (2022). Heritable base-editing in Arabidopsis using RNA viral vectors. Plant Physiol. 189, 1920–1924. doi: 10.1093/plphys/kiac206
Liu, Q., Zhao, C., Sun, K., Deng, Y., and Li, Z. (2023). Engineered biocontainable RNA virus vectors for non-transgenic genome editing across crop species and genotypes. Mol. Plant 16, 616–631. doi: 10.1016/j.molp.2023.02.003
Lowe, K., La Rota, M., Hoerster, G., Hastings, C., Wang, N., Chamberlin, M., et al. (2018). Rapid genotype “independent” Zea mays L. (maize) transformation via direct somatic embryogenesis. In Vitro Cell Dev. Biol. Plant 54, 240–252. doi: 10.1007/s11627-018-9905-2
Lowe, K., Wu, E., Wang, N., Hoerster, G., Hastings, C., Cho, M.-J., et al. (2016). Morphogenic regulators baby boom and wuschel improve monocot transformation. Plant Cell 28, 1998–2015. doi: 10.1105/tpc.16.00124
Lowry, G. V., Giraldo, J. P., Steinmetz, N. F., Avellan, A., Demirer, G. S., Ristroph, K. D., et al. (2024). Towards realizing nano-enabled precision delivery in plants. Nat. Nanotechnology 19, 1255–1269. doi: 10.1038/s41565-024-01667-5
Lundgren, M. R. and Des Marais, D. L. (2020). Life history variation as a model for understanding trade-offs in plant–environment interactions. Curr. Biol. 30, R180–R189. doi: 10.1016/j.cub.2020.01.003
Luo, G., Najafi, J., Correia, P. M. P., Trinh, M. D. L., Chapman, E. A., Østerberg, J. T., et al. (2022). Accelerated domestication of new crops: yield is key. Plant Cell Physiol. 63, 1624–1640. doi: 10.1093/pcp/pcac065
Luo, W., Suzuki, R., and Imai, R. (2023). Precise in planta genome editing via homology-directed repair in wheat. Plant Biotechnol. J. 21, 668–670. doi: 10.1111/pbi.13984
Maher, M. F., Nasti, R. A., Vollbrecht, M., Starker, C. G., Clark, M. D., and Voytas, D. F. (2020). Plant gene editing through de novo induction of meristems. Nat. Biotechnol. 38, 84–89. doi: 10.1038/s41587-019-0337-2
Matres, J. M., Hilscher, J., Datta, A., Armario-Nájera, V., Baysal, C., He, W., et al. (2021). Genome editing in cereal crops: an overview. Transgenic Res. 30, 461–498. doi: 10.1007/s11248-021-00259-6
Mei, G., Chen, A., Wang, Y., Li, S., Wu, M., Hu, Y., et al. (2024). A simple and efficient in planta transformation method based on the active regeneration capacity of plants. Plant Commun. 5, 100822. doi: 10.1016/j.xplc.2024.100822
Meyer, R. S., DuVal, A. E., and Jensen, H. R. (2012). Patterns and processes in crop domestication: an historical review and quantitative analysis of 203 global food crops. New Phytol. 196, 29–48. doi: 10.1111/j.1469-8137.2012.04253.x
Notaguchi, M., Abe, M., Kimura, T., Daimon, Y., Kobayashi, T., Yamaguchi, et al. (2008). Long-Distance, Graft-Transmissible Action of Arabidopsis FLOWERING LOCUS T Protein to Promote Flowering. Plant Cell Physiol. 49, 1645–1658. doi: 10.1093/pcp/pcn154
Obermeyer, G. and Weisenseel, M. H. (1995). Introduction of impermeable molecules into pollen grains by electroporation. Protoplasma 187, 132–137. doi: 10.1007/BF01280241
Olsson, L., Andersson, E., Ardö, J., Crews, T., David, C., DeHaan, L., et al. (2024). What is the prospect of a perennial grain revolution of agriculture? Global Sustainability 7, e35. doi: 10.1017/sus.2024.27
Østerberg, J. T., Xiang, W., Olsen, L. I., Edenbrandt, A. K., Vedel, S. E., Christiansen, A., et al. (2017). Accelerating the domestication of new crops: feasibility and approaches. Trends Plant Sci. 22, 373–384. doi: 10.1016/j.tplants.2017.01.004
Paul, J. D., Lutsiv, T., and Thompson, H. J. (2024). A Perennial Green Revolution to address 21st-century food insecurity and malnutrition. Food Energy Secur. 13. doi: 10.1002/fes3.568
Reeves, G., Tripathi, A., Singh, P., Jones, M. R. W., Nanda, A. K., Musseau, C., et al. (2022). Monocotyledonous plants graft at the embryonic root–shoot interface. Nature 602, 280–286. doi: 10.1038/s41586-021-04247-y
Singh, P. and Kumar, K. (2022). Agrobacterium-mediated In-planta transformation of bread wheat (Triticum aestivum L.). J. Plant Biochem. Biotechnol. 31, 206–212. doi: 10.1007/s13562-021-00669-x
Sundararajan, S., Nayeem, S., Sivakumar, H. P., and Ramalingam, S. (2023). Agrobacterium-mediated rapid and efficient development of transgenics using shoot apex explants in two elite Indica rice cultivars. Cereal Res. Commun. 51, 917–929. doi: 10.1007/s42976-023-00366-6
Supartana, P., Shimizu, T., Shioiri, H., Nogawa, M., Nozue, M., and Kojima, M. (2005). Development of simple and efficient in planta transformation method for rice (Oryza sativa L.) using Agrobacterium tumefaciens. J. Biosci. Bioeng. 100, 391–397. doi: 10.1263/jbb.100.391
Tamilselvan-Nattar-Amutha, S., Hiekel, S., Hartmann, F., Lorenz, J., Dabhi, R. V., Dreissig, S., et al. (2023). Barley stripe mosaic virus-mediated somatic and heritable gene editing in barley (Hordeum vulgare L.). Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1201446
Tamizi, A. A., Md-Yusof, A. A., Mohd-Zim, N. A., Nazaruddin, N. H., Sekeli, R., Zainuddin, Z., et al. (2023). Agrobacterium-mediated in planta transformation of cut coleoptile: a new, simplified, and tissue culture-independent method to deliver the CRISPR/Cas9 system in rice. Mol. Biol. Rep. 50, 9353–9366. doi: 10.1007/s11033-023-08842-2
Tarafdar, A., Vishwakarma, H., Gothandapani, S., Bhati, M., Biswas, K., Prakash, A., et al. (2019). A quick, easy and cost-effective in planta method to develop direct transformants in wheat. 3 Biotech. 9, 180. doi: 10.1007/s13205-019-1708-6
Tezuka, D., Cho, H., Onodera, H., Linghu, Q., Chijimatsu, T., Hata, M., et al. (2024). Redirecting barley breeding for grass production through genome editing of Photoperiod-H1. Plant Physiol. 195, 287–290. doi: 10.1093/plphys/kiae075
Tjokrokusumo, D., Heinrich, T., Wylie, S., Potter, R., and McComb, J. (2000). Vacuum infiltration of Petunia hybrida pollen with Agrobacterium tumefaciens to achieve plant transformation. Plant Cell Rep. 19, 792–797. doi: 10.1007/s002990050009
Toda, E., Kato, N., Higashiyama, T., and Okamoto, T. (2023). Genome editing approaches using reproductive cells/tissues in flowering plants. Front. Genome. Ed. 4. doi: 10.3389/fgeed.2022.1085023
Touraev, A., Stöger, E., Voronin, V., and Heberle-Bors, E. (1997). Plant male germ line transformation. Plant J. 12, 949–956. doi: 10.1046/j.1365-313X.1997.12040949.x
Vejlupkova, Z., Warman, C., Sharma, R., Scheller, H. V., Mortimer, J. C., and Fowler, J. E. (2020). No evidence for transient transformation via pollen magnetofection in several monocot species. Nat. Plants. 6, 1323–1324. doi: 10.1007/s11248-007-9115-x
Wang, J. W., Grandio, E. G., Newkirk, G. M., Demirer, G. S., Butrus, S., Giraldo, J. P., et al. (2019). Nanoparticle-mediated genetic engineering of plants. Mol. Plant 12, 1037–1040. doi: 10.1016/j.molp.2019.06.010
Wang, N., Ryan, L., Sardesai, N., Wu, E., Lenderts, B., Lowe, K., et al. (2023). Leaf transformation for efficient random integration and targeted genome modification in maize and sorghum. Nat. Plants 9, 255–270. doi: 10.1038/s41477-022-01338-0
Wang, K., Shi, L., Liang, X., Zhao, P., Wang, W., Liu, J., et al. (2022). The gene TaWOX5 overcomes genotype dependency in wheat genetic transformation. Nat. Plants 8, 110–117. doi: 10.1038/s41477-021-01085-8
Weiss, T., Kamalu, M., Shi, H., Li, Z., Amerasekera, J., Zhong, Z., et al. (2025). Viral delivery of an RNA-guided genome editor for transgene-free germline editing in Arabidopsis. Nat. Plants 11, 967–976.. doi: 10.1038/s41477-025-01989-9
Wolf, S., Lucas, W. J., Deom, C. M., and Beachy, R. N. (1989). Movement protein of tobacco mosaic virus modifies plasmodesmatal size exclusion limit. Science 246, 377–379. doi: 10.1126/science.246.4928.377
Xu, N., Kang, M., Zobrist, J. D., Wang, K., and Fei, S. (2022). Genetic transformation of recalcitrant upland switchgrass using morphogenic genes. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.781565
Yang, L., Machin, F., Wang, S., Saplaoura, E., and Kragler, F. (2023). Heritable transgene-free genome editing in plants by grafting of wild-type shoots to transgenic donor rootstocks. Nat. Biotechnol. 41, 958–967. doi: 10.1038/s41587-022-01585-8
Yang, W., Zhai, H., Wu, F., Deng, L., Chao, Y., Meng, X., et al. (2024). Peptide REF1 is a local wound signal promoting plant regeneration. Cell 187, 3024–3038.e14. doi: 10.1016/j.cell.2024.04.040
Ye, X., Shrawat, A., Williams, E., Rivlin, A., Vaghchhipawala, Z., Moeller, L., et al. (2022). Commercial scale genetic transformation of mature seed embryo explants in maize. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.1056190
Yellisetty, V., Reddy, L. A., and Mandapaka, M. (2015). In planta transformation of sorghum (Sorghum bicolor (L.) Moench) using TPS1 gene for enhancing tolerance to abiotic stresses. J. Genet. 94, 425–434. doi: 10.1007/s12041-015-0540-y
Zhang, S., Huang, G., Zhang, Y., Lv, X., Wan, K., Liang, J., et al. (2022). Sustained productivity and agronomic potential of perennial rice. Nat. Sustain. 6, 28–38. doi: 10.1038/s41893-022-00997-3
Zhang, Y., Liang, Z., Zong, Y., Wang, Y., Liu, J., Chen, K., et al. (2016). Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 7, 12617. doi: 10.1038/ncomms12617
Zhang, S., Sun, L., and Kragler, F. (2009). The phloem-delivered RNA pool contains small noncoding RNAs and interferes with translation. Plant Physiol. 150, 378–387. doi: 10.1104/pp.108.134767
Zhang, W., Thieme, C. J., Kollwig, G., Apelt, F., Yang, L., Winter, N., et al. (2016). tRNA-related sequences trigger systemic mRNA transport in plants. Plant Cell 28, 1237–1249. doi: 10.1105/tpc.15.01056
Zhao, X., Meng, Z., Wang, Y., Chen, W., Sun, C., Cui, B., et al. (2017). Pollen magnetofection for genetic modification with magnetic nanoparticles as gene carriers. Nat. Plants 3, 956–964. doi: 10.1038/s41477-017-0063-z
Zhong, H., Li, C., Yu, W., Zhou, H.-p., Lieber, T., Su, X., et al. (2024). A fast and genotype-independent in planta Agrobacterium-mediated transformation method for soybean. Plant Commun. 5, 101063. doi: 10.1016/j.xplc.2024.101063
Keywords: In planta transformation, recalcitrance, perennial agriculture, grain crops, monocots
Citation: Correia PMP, Dong X, Chen M, Nørrevang AF, Luo G and Palmgren M (2025) In planta transformation methods to accelerate the domestication of perennial grain crops. Front. Plant Sci. 16:1638144. doi: 10.3389/fpls.2025.1638144
Received: 30 May 2025; Accepted: 30 June 2025;
Published: 17 July 2025.
Edited by:
Min Chul Kim, Gyeongsang National University, Republic of KoreaReviewed by:
Taras P. Pasternak, Miguel Hernández University of Elche, SpainAlbert Chern Sun Wong, University of Queensland, Australia
Copyright © 2025 Correia, Dong, Chen, Nørrevang, Luo and Palmgren. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael Palmgren, cGFsbWdyZW5AcGxlbi5rdS5kaw==