 Sofija Nešić
Sofija Nešić Dragana Bosnić
Dragana Bosnić Jelena Samardžić
Jelena Samardžić Ivana Nikolić
Ivana Nikolić Aziz Aziz
Aziz Aziz Dragana Nikolić
Dragana Nikolić- 1Group for Plant Molecular Biology, Institute of Molecular Genetics and Genetic Engineering, Department of Microbiology and Plant Biology, University of Belgrade, Belgrade, Serbia
- 2Induced Resistance and Plant Bioprotection (RIBP), University of Reims Champagne Ardenne, UFR Sciences, Reims, France
Fungal pathogens are one of the main causes of yield losses in many crops, severely affecting agricultural production worldwide. Among the various approaches to alleviate this problem, beneficial microorganisms emerge as an environmentally friendly and sustainable alternative. In addition to direct biocontrol action against pathogens, certain plant growth-promoting bacteria (PGPB) enhance the plant immune defense to control diseases through induced systemic resistance (ISR). Paraburkholderia phytofirmans PsJN has been shown as an efficient biocontrol agent against diseases. However, the specific mechanisms underlying these beneficial effects at both local and systemic level remain largely unknown. In this study, we investigated the transcriptional response of Arabidopsis thaliana at above- and below-ground levels upon interaction with P. phytofirmans PsJN, and after Botrytis cinerea infection. Our data clearly support the protective effect of P. phytofirmans PsJN through ISR against B. cinerea in plants grown in both soil and hydroponic conditions. The comparative transcriptome analysis of the mRNA and miRNA sequences revealed that PsJN modulates the expression of genes involved in abiotic stress responses, microbe-plant interactions and ISR, with ethylene signaling pathway genes standing out. In roots, PsJN predominantly downregulated the expression of genes related to microbe perception, signaling and immune response, indicating that PsJN locally provoked attenuation of defense responses to facilitate and support colonization and the maintenance of mutualistic relationship. In leaves, the increased expression of defense-related genes prior to infection in combination with the protective effect of PsJN observed in later stages of infection suggests that bacterial inoculation primes plants for enhanced systemic immune response after subsequent pathogen attack.
1 Introduction
Harnessing plant-associated microbes and their role in plant health represents a promising environment friendly strategy for a sustainable agriculture. Most of beneficial bacteria including the plant growth promoting bacteria (PGPB) associated with plants in the close vicinity of their roots (rhizosphere) or within the plant tissues (as endophytes) play a crucial role in plant health and productivity. These bacteria promote plant growth and prime plants for enhanced resistance against abiotic and biotic stress (Lugtenberg and Kamilova, 2009). The beneficial bacteria-mediated resistance relies mainly on the priming process of plant immune system (Pieterse et al., 2014; Conrath et al., 2015; Gruau et al., 2015). However, little headway has been made in identifying the molecular mechanisms involved in the primed mechanism linked to PGPB-induced resistance.
Plant immunity is a complex system that is a result of a co-evolution between plants and pathogens. It has been shown that interaction with beneficial bacteria can help plants to better react to a subsequent pathogen attack (Pieterse et al., 2014). This defense strategy triggered by PGPB, to prime plants against pathogens, is called induced systemic resistance (ISR). As a result, plants develop an earlier and stronger defense reaction to the pathogen attack, delaying symptom development, reducing disease severity and inhibiting pathogen growth (van Loon, 2007; Cartieaux et al., 2008).
Phytopathogenic fungi are a dominant cause of plant diseases. Botrytis cinerea is a phytopathogenic fungus affecting a wide range of hosts. As an aggressive necrotroph, it is responsible for major crop loss every year (Bi et al., 2023). Preventing this pathogen, particularly at an early stage, is of great importance. PGPB like Bacillus cereus AR156, Burkholderia sp. AU4i, Pseudomonas fluorescens PTA-CT2 and Paraburkholderia phytofirmans PsJN have been shown to increase plant tolerance to this pathogen in Arabidopsis and grapevine (Miotto-Vilanova et al., 2016; Nie et al., 2017; Nguyen et al., 2020; Colavolpe et al., 2021).
P. phytofirmans PsJN (previously known as Burkholderia phytofirmans PsJN) has been shown to colonize a wide variety of plant species including grapevine, rice, and tomato, as well as Arabidopsis thaliana (Poupin et al., 2013; Issa et al., 2018; Timmermann et al., 2019; King et al., 2023). P. phytofirmans PsJN also activates plant defense system in different crop species and in A. thaliana, thereby reducing their susceptibility to various pathogens (Timmermann et al., 2017; Colavolpe et al., 2021). This beneficial bacterial strain dwells in the rhizosphere as well as within plant tissues as an endophyte, supporting its beneficial affect in promoting plant growth and their resistance to various abiotic and biotic stresses. Poupin et al. (2013) have shown that a single inoculation of Arabidopsis with PsJN influences the whole life cycle of the plant, accelerating growth rate and shortening vegetative period, both of which are important for crop production. PsJN enhances plant growth and health through various mechanisms, including the production of growth-promoting phytohormones like auxins, the enzyme 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase that reduces stress hormone ethylene levels, and the secretion of siderophores in the rhizosphere that increase iron availability (Esmaeel et al., 2018).
Exploring the molecular mechanisms of plant-microbiota interactions is essential for understanding how plants respond to beneficial microbes. Global transcriptome analyses on PsJN effects in plants are scarce, leaving a gap in understanding the mechanisms involved in beneficial effect of PsJN. In the study investigating PsJN-Arabidopsis interaction, before pathogen challenge, 405 genes (corresponding to 1.8% of the analyzed Arabidopsis genome) were regulated by the bacterium. A principal component analysis showed that PsJN-induced plant responses to the pathogen could be differentiated from those induced by the pathogen itself. A network comprising four gene clusters was changed by PsJN, including genes related to jasmonate, ethylene, salicylic acid and ROS pathways (Timmermann et al., 2019). Another research highlights the distinct effects of PsJN and native Burkholderia endophytes found in rice (King et al., 2023). The colonization of the rice endosphere by PsJN elicits a unique transcriptomic response compared to native strains. This response encompasses alterations in genes associated with secondary metabolism, immunity, and phytohormones (King et al., 2023). The molecular and biochemical responses of A. thaliana plants inoculated with the PsJN strain were examined under both short-term and long-term salt stress throughout their entire life cycle. The findings revealed transcriptional responses linked to salt tolerance, including a rapid activation of general abiotic stress-responsive genes, and the transcriptional regulation of genes related to nutrient uptake (Pinedo et al., 2015). Future research should explore whether these insights can be applied to other plant models and beneficial microbes, thereby enhancing the potential of microbiome-based strategies for improving crop production and resilience.
One of the key regulators of plant development, growth and stress response are small RNAs (Borges and Martienssen, 2015; Waheed et al., 2021). They are known to modulate their target genes, adjusting their expression, and thus the response of plants to their environment (Axtell and Bowman, 2008). Micro RNAs are a class of small non-coding RNAs that regulate expression of the target genes through induced mRNA degradation and translational repression (Iwakawa and Tomari, 2013; Li et al., 2013b; Middleton et al., 2021). sRNAs have been shown to play a positive role in plant immune defense against various pathogenic microbes, including bacteria, fungi and oomycetes (Kong et al., 2022).
In this study we evaluated the P. phytofirmans PsJN activity in enhancing both A. thaliana resistance to the necrotrophic fungus B. cinerea and plant growth, and explored mechanisms underlying PsJN-mediated resistance by analyzing transcriptional alterations occurring in A. thaliana after challenge with B. cinerea. We more especially focused on transcriptional response using mRNA and sRNA sequencing in both roots and leaves upon interactions with P. phytofirmans PsJN and after pathogen challenge.
2 Materials and methods
2.1 Plant material and growth conditions
Seeds of Arabidopsis thaliana Col-0 were surface-sterilized with 70% (v/v) ethanol for 5 min, followed by 20 min incubation in a sterilizing solution (10% commercial bleach, 0.05% (v/v) Tween 20, dH2O) (Nikolić et al., 2023). The seeds were washed 3 times with sterile distilled water (dH2O). After 5 days of stratification in the dark at +4°C, seeds were placed on soil and covered with cling foil for two weeks for optimal humidity during germination. Plants were grown under short-day regime (10h day/ 14h night), light intensity of 150 μmol m−2s−1, relative humidity of 55-65%, day/night temperatures of 21°C/19°C. Arabidopsis plants were also grown hydroponically following protocol Conn et al. (2013). In hydroponics, plants were grown under the same conditions (light intensity, day length and humidity) as plants grown in soil.
2.2 Bacterial growth conditions and plant inoculation
Paraburkholderia phytofirmans PsJN (obtained from Leibniz Institute DSMZ - German Collection of Microorganisms and Cell Cultures) was grown in LB or minimal media (Coleman et al., 2002) for 18 h, in an orbital shaker (150 rpm) at 30°C. For soil drench inoculation, bacteria were pelleted at 2500 g/4°C for 15 min, washed and resuspended in 10 mM MgSO4. Three-week-old plants were soil-drenched with PsJN to reach a final concentration of 1 × 108 CFU/g soil. The same volume of 10 mM MgSO4 was used as a mock treatment (control). For hydroponic inoculation, bacteria were grown in minimal media pelleted at 1500 g/4°C for 15 min and washed with hydroponic medium. PsJN suspension was added to the germination medium during the first week of seedling growth. Adult plants were inoculated two weeks prior to fungal infection, by adding PsJN (1 × 106 CFU/ml hydroponic medium) at each medium exchange.
2.3 Fungal growth conditions, infection and disease analysis
Botrytis cinerea strain 630 was grown on potato dextrose agar (PDA) plates at 22°C under indirect light. Conidia were collected from plates with sterile potato dextrose broth (PDB 12 g/l) and filtered to remove hyphae. Conidial concentration was adjusted to 1 × 106 conidia/ml (Aziz et al., 2003). Collected conidia were used immediately or kept frozen at -80°C until use. For plant infection, germinated spores incubated at 22°C for 18–22 h were used. Two weeks after bacterial inoculation, plants were infected with 20 µl of germinated conidia per developed leaf. To aid in infection, light intensity was lowered to 60-80 μmol m−2s−1 and humidity increased to 80-100%. To provide high humidity necessary for B. cinerea infection, plants were held in closed transparent boxes one day before, and another day after the pathogen challenge. Also, aeration of roots in hydroponics was withheld during the two days of treatments. To assess the progression of pathogen attack, the average size of disease lesions developed four days post infection was measured and analyzed using image analysis software ImageJ (Schneider et al., 2012). For the analysis of gene expression, to obtain a homogeneous application, germinated conidia suspension was applied to the whole leaf areas. PDB was used in the same manner as a mock treatment.
2.4 Sampling
Roots and leaves of plants grown hydroponically were sampled at 24 hours post fungal infection (hpi), weighed, and immediately frozen in liquid nitrogen and stored at -80C until RNA extraction. Roots were also washed 2–3 times with sterile distilled H2O before freezing. Three replicates were collected for each treatment, each replicate comprising three individual plants. Treatments were: plants mock inoculated and mock infected - CNI, plants mock inoculated and infected with B. cinerea - CBc, plants inoculated with PsJN and mock infected - PNI, plants inoculated with PsJN and infected with B. cinerea - PBc.
2.5 RNA extraction
Leaf or root tissue samples collected at 24hpi were ground using mortar and pestle. 50mg of powder was used for RNA extraction. RNA was extracted with microRNA Purification Kit (Norgen BioTek) following protocol for plants. Both small and long RNA isolates were treated with the Ambion® DNA-free™ DNase Treatment and Removal DNA kit. As total RNA was required as an input for RNA sequencing, small and long RNA fractions were combined. Three samples per treatment group per tissue were used for RNAseq (24 samples) as well as sRNAseq (24 samples).
2.6 mRNA library construction and sequencing
Twenty-four samples were sent for RNA sequencing performed by Novogene Bioinformatics Technology Co., Ltd. (Cambridge, UK). Integrity and purity of RNA was checked with Agilent 2100 Bioanalyzer, samples passed quality control were used for mRNA library construction. Library preparation included poly A enrichment and cDNA synthesis. Prepared libraries were sequenced on NovaSeq X Plus Series platform (Illumina, Inc., San Diego, CA, USA) using paired-end 150 bp sequencing strategy.
2.7 Bioinformatics analysis
The sequencing results were analyzed with standard Novogene pipeline. Clean data (clean reads) was obtained from raw data using fastp software. Paired-end cleaned reads were aligned to the reference genome using Hisat2 (v 2.0.5) (Mortazavi et al., 2008). Arabidopsis thaliana Genome assembly TAIR10.1 (ncbi_arabidopsis_thaliana_gcf_000001735_4_tair10_1) was used as the reference genome. The mapped reads of each sample were assembled by StringTie (v1.3.3b) (Pertea et al., 2015). Quantification of gene expression level was performed using FeatureCounts v1.5.0-p3 (Liao et al., 2014). DESeq2 (1.20.0) was used to analyze differentially expressed genes (DEGs) (Love et al., 2014). Genes with an adjusted P-value ≤ 0.05 found by DESeq2 were assigned as differentially expressed. Gene Ontology (GO) enrichment analysis (Young et al., 2010) of differentially expressed genes was implemented by the clusterProfiler R package (v 3.8.1), in which gene length bias was corrected. GO terms with corrected P-value less than 0.05 were considered significantly enriched by differential expressed genes.
Additionally, MapMan software was used to provide a general overview and graphic representation of DEGs involved in metabolic pathways and other biological processes (Thimm et al., 2004).
2.8 Small RNA library construction and sequencing
Twenty-four samples (twelve from leaves and twelve from roots) were sent to Novogene Bioinformatics Technology Co., Ltd. (Cambridge, UK) for small RNA sequencing. After adapter ligation to both ends of small RNA, first strand cDNA was synthesized during hybridization with reverse transcription primer. The double-stranded cDNA library was generated through PCR enrichment. After purification and size selection, libraries with insertions between 18~40 bp were ready for sequencing. The library was checked with Qubit and real-time PCR for quantification and Agilent 2100 Bioanalyzer for size distribution detection. Single-end 50-bp sequencing of small RNA libraries was performed on Illumina NovaSeq 6000 platform.
2.9 Small RNA bioinformatics analysis
The sequencing results were analyzed with standard Novogene pipeline for small RNAs. Clean reads were used to map small RNA to reference genome using bowtie (Langmead et al., 2009). To analyze small RNA expression and distribution on the reference sequence mapping was done without mismatch or with only one mismatch. Mapped small RNA tags were used to look for known miRNA. miRBase22.0 was used as reference, modified software mirdeep2 (Friedländer et al., 2012) and srna-tools-cli were used to obtain the potential miRNA and draw the secondary structures. Predicting the target gene of miRNA was performed by psRobot_tar in psRobot (Wu et al., 2012). Quantification of miRNA expression levels were estimated by TPM (transcript per million) through the following criteria (Zhou et al., 2011): Normalization formula: Normalized expression = mapped readcount/Total reads * 1000000. Differential expression analysis of two conditions/groups was performed using the DESeq2. The P-values were adjusted using the Benjamini & Hochberg method. Corrected P-value of 0.05 was set as the threshold for significantly differential expression by default. GO enrichment analysis was used on the target gene candidates of differentially expressed miRNAs.
The miRNA-mRNA network was constructed by using Cytoscape 3.10.1 software (Shannon et al., 2003).
3 Results
3.1 Paraburkholderia phytofirmans PsJN protects Arabidopsis thaliana against Botrytis cinerea
To test whether the PGPB P. phytofirmans PsJN induces a protective effect in A. thaliana challenged with B. cinerea, mature plants, previously inoculated with P. phytofirmans PsJN at the root level or mock-inoculated were infected with germinated spores of B. cinerea. Four days after infection, disease lesions were analyzed. Control plants (mock-inoculated) showed typical symptoms of necrosis, while B. cinerea-induced necrotic lesions in PsJN-inoculated plants were significantly smaller (P<0.0001). These results support the idea that PsJN triggered a systemic resistance against B. cinerea. This PsJN-induced resistance was observed in A. thaliana grown in both soil and hydroponics (Figure 1).
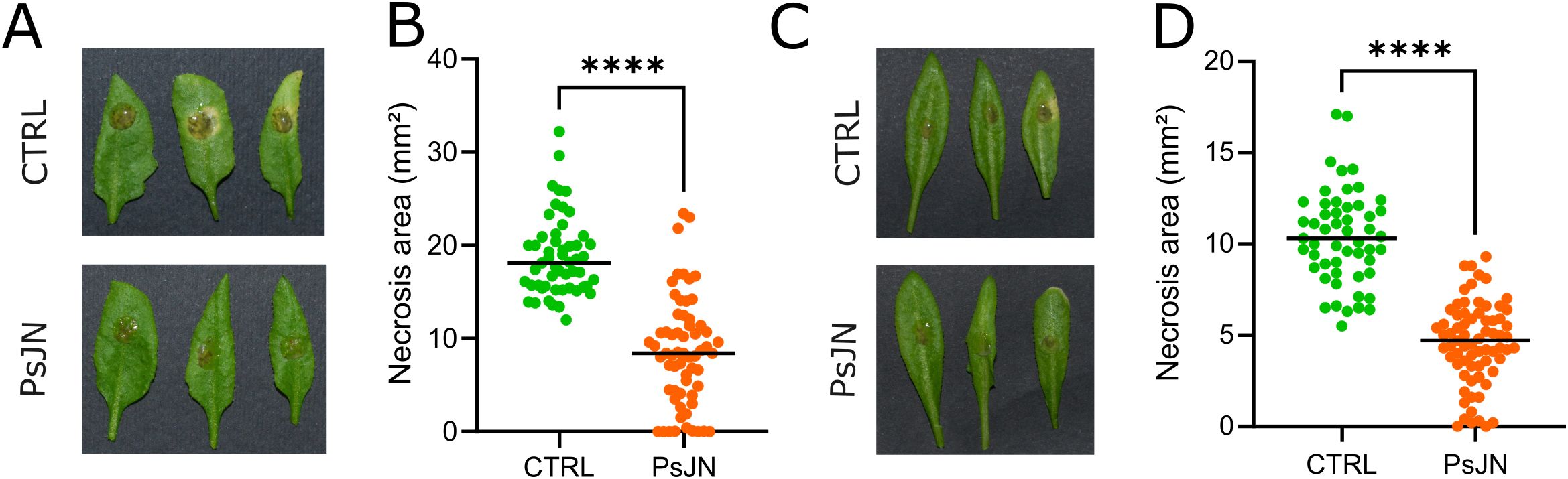
Figure 1. Paraburkholderia phytofirmans PsJN protects Arabidopsis thaliana against Botrytis cinerea and promotes plant growth. Representative photographs of Arabidopsis leaves infected with B. cinerea previously mock-inoculated or inoculated with PsJN; plants grown in soil (A) and hydroponics (C). Necrosis area caused by B. cinerea in plants grown in soil (B) and hydroponics (D). **** indicate significant difference at P < 0.0001 as determined by Mann-Whitney test analysis.
Aside from increasing A. thaliana resistance to B. cinerea, PsJN promoted growth of inoculated plants by increasing both leaf and root biomass (Supplementary Figure S1).
3.2 Transcriptome profiling of the PsJN-inoculated Arabidopsis plants
Recent advancements in next-generation sequencing have resulted in a surge of transcriptomic research examining alterations in host gene expression during interactions between plants and plant growth-promoting bacteria (PGPB). We performed RNA-Seq for the RNA isolated from roots and leaves of both PsJN-inoculated and non-inoculated Arabidopsis. Plants were grown in hydroponics to facilitate analysis of root transcriptome, alongside with the transcriptome of leaves.
Before B. cinerea infection, transcriptomic analysis showed a total of 3388 genes upregulated and 3346 genes downregulated in Arabidopsis roots inoculated with PsJN, compared to the non-inoculated control (comparison PNI vs CNI, found by DESeq2 with adjusted p-value ≤ 0.05). The number of differentially expressed genes (DEGs) showing upregulation of FC > 1.5 was 1746, and the number showing downregulation of FC < 0.6 was 2468. In leaves of PsJN-inoculated plants, 2573 were upregulated and 2438 downregulated compared to the non-inoculated control. The number of DEGs showing upregulation of FC > 1.5 was 598, and the number showing downregulation of FC < 0.6 was 514 (Figure 2; Supplementary Table S1).
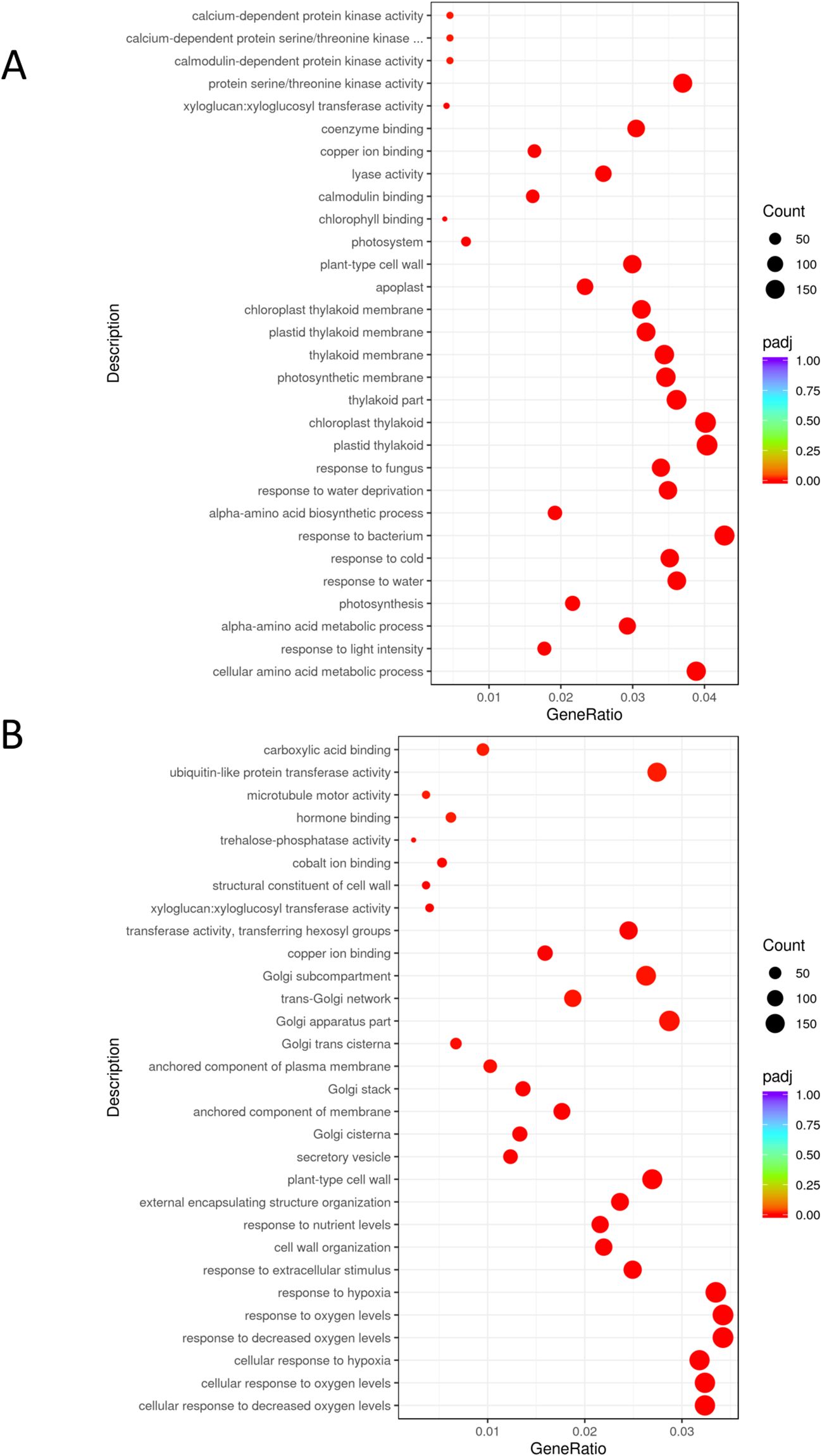
Figure 2. Gene Ontology (GO) enrichment analysis of DEGs in Arabidopsis induced by PsJN inoculation (comparison PNI vs CNI). GO enrichment chart displays the top 30 most significant pathways comprising genes with significantly changed expression in (A) leaves and (B) roots. The colors represent different levels of significance, with red indicating highly significant terms. padj: adjusted p-value.
At 24h post-infection with B. cinerea, 507 genes were upregulated and 474 genes were downregulated in Arabidopsis roots inoculated with PsJN, compared to the non-inoculated plants (comparison PBc vs CBc). The number of DEGs showing upregulation of FC > 1.5 was 81, and the number showing downregulation of FC < 0.6 was 172. In B. cinerea-infected leaves of PsJN-treated plants, 150 genes were upregulated and 224 were downregulated compared to the non-inoculated plants. The number of DEGs showing upregulation of FC > 1.5 was 55, and the number showing downregulation of FC < 0.6 was 115 (Supplementary Table S1).
Transcriptome analysis of plant roots and leaves before B. cinerea infection (PNI vs CNI) revealed several DEGs involved in responses to both abiotic and biotic stresses (Figure 2; Supplementary Table S1). Data indicated that PsJN modulates the expression of a number of genes involved in ethylene synthesis and signaling pathway in both leaves (Figure 3A) and roots (Figure 3B). PsJN induces a significant upregulation of ACO1 and ACO3 genes, encoding 1-aminocyclopropane-1-carboxylic acid oxidase in the roots (Figure 3B). Two other genes involved in ethylene perception like Ethylene Response ETR2 and Ethylene Response Sensor ERS2 were also upregulated in both leaves and roots of plants inoculated with PsJN. The transcriptome analysis also revealed that PsJN differently affects the expression of numerous ethylene response factors ERFs, depending on their target genes and the plant organ (Figure 3). The most of ERF genes are upregulated in leaves, while in roots both up and downregulation of different ERF genes is detected as a response to PsJN inoculation.
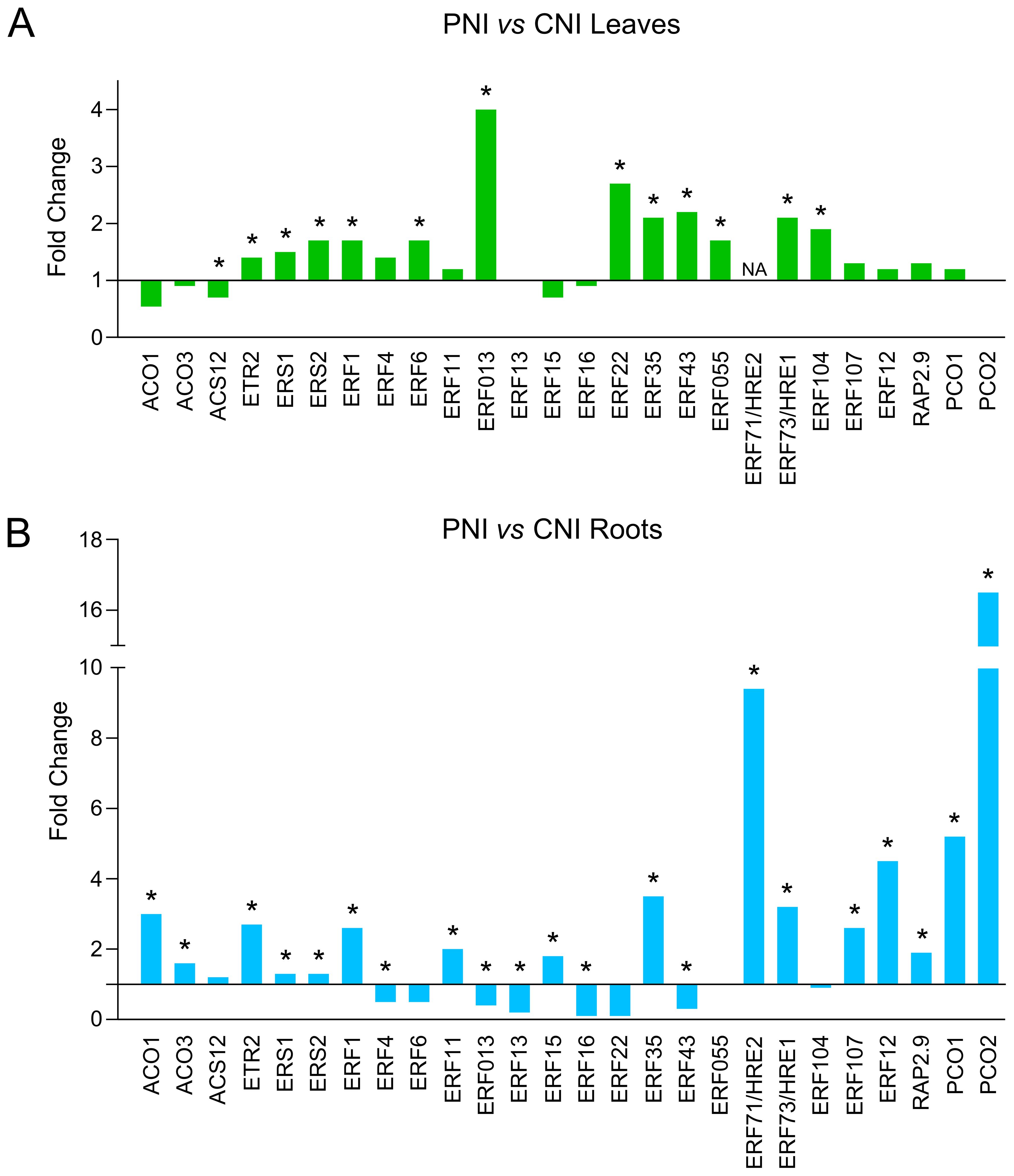
Figure 3. Expression levels of genes involved in ethylene synthesis and signaling pathway in roots and leaves of PsJN-treated plants before B cinerea inoculation. (A) Leaves of PsJN-treated plants vs non-inoculated control. (B) Roots of PsJN-treated plants vs non-inoculated control. * denotes values with adjusted P-value < 0.05.
3.3 PsJN modulates the expression of genes related to abiotic stress responses
Gene Ontology of the PsJN-upregulated genes indicated functional categories of biological processes including pathways related to responses to abiotic stimuli, decreased oxygen levels, nutrient levels and absence of light (Figure 2). We identified a significant number of genes related to hypoxia sensing and response in both roots and leaves (Figures 3, 4). This could be related to hydroponic plant growth conditions without aeration of roots during 48h of treatment or to the pre-exposure of the plants to high humidity before B. cinerea infection. The key oxygen sensors are members of the ERFVIIs family: Hypoxia-Responsive ERFs (HRE) and Related to Apetala2 (RAP2) (Licausi et al., 2010; Gibbs et al., 2011). The expression of HRE1 and HRE2 increased significantly in the roots of Arabidopsis grown with PsJN, while HRE1 was upregulated in the leaves. PsJN did not significantly affect the expression of key RAP transcription factors, with only RAP2.9 being upregulated twofold in the roots. PCO1 and PCO2 (encoding plant cysteine oxidases), which also act as oxygen sensors and are involved in the N-end rule pathway, were also upregulated in roots inoculated with PsJN.
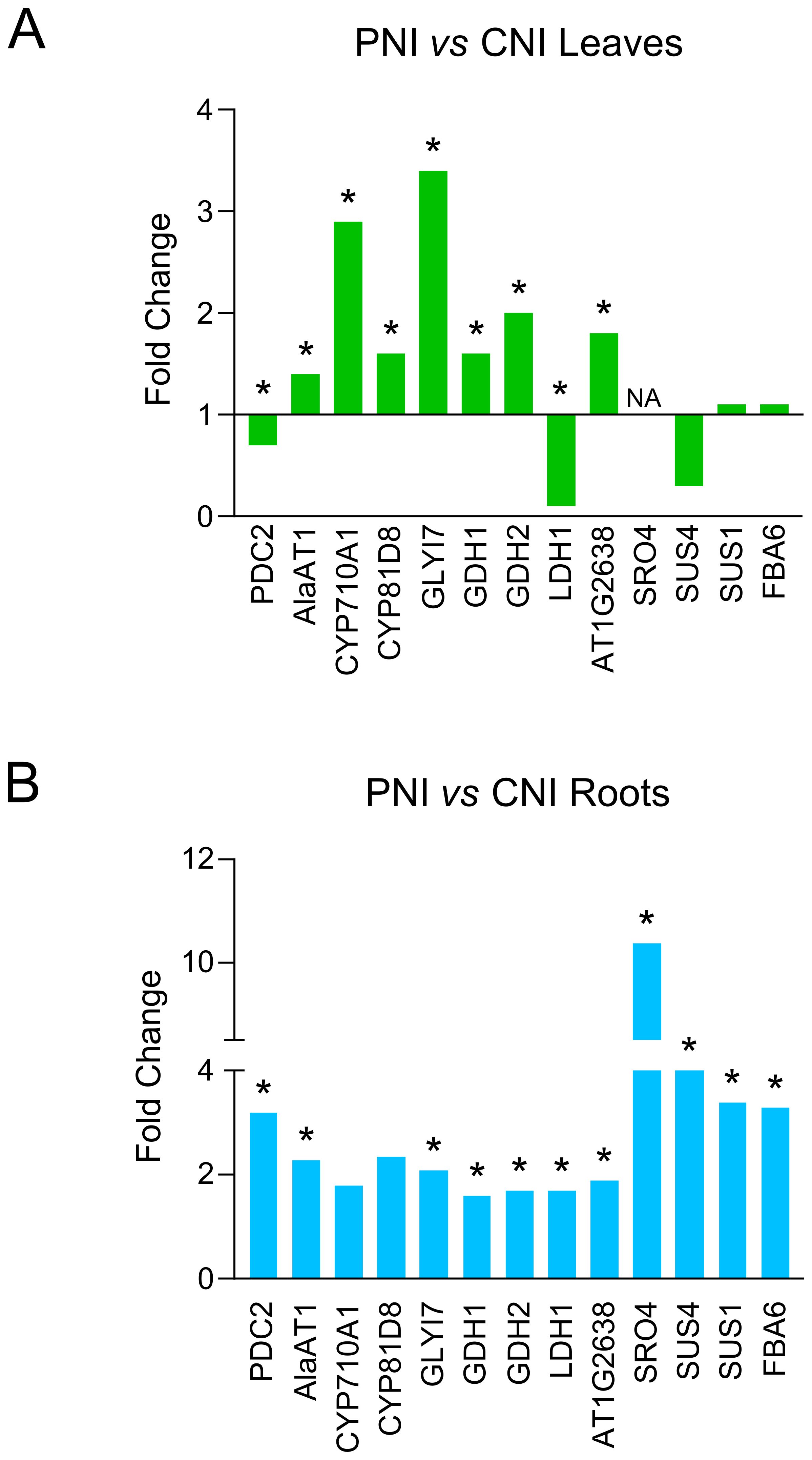
Figure 4. Expression levels of genes involved in hypoxia response in roots and leaves of PsJN-treated plants before B. cinerea inoculation. (A) Leaves of PsJN-treated plants vs non-inoculated control. (B) Roots of PsJN-treated plants vs non-inoculated control. * denotes values with adjusted P-value < 0.05.
Analyzing transcriptome sequencing in the roots, we also found that PsJN significantly upregulated transcription of some genes involved in fermentation processes using pyruvate as a starting substrate, including PDC2 (pyruvate decarboxylase2), LDH1 (lactate dehydrogenase), and AlaAT1 (alanine aminotransferase). We also noticed a significant increase of other genes important for plant survival under low-oxygen conditions, such as GDH1 and GDH2 (glutamate dehydrogenase), SUS1 and SUS4 (encoding sucrose synthases), FBA6 (fructose-1,6-bisphosphate aldolase 6), an enzyme involved in glycolysis and gluconeogenesis and GLI17 (glyoxalase I) (Figure 4).
3.4 PsJN modulates the expression of genes associated with biotic stress responses
Using the MapMan software (Thimm et al., 2004), we obtained an overview of the PsJN-induced changes in pathways involved in biotic stress. As shown in Figures 5, 6, the PsJN-induced change in A. thaliana transcriptome was much more pronounced prior to B. cinerea infection than in plants 24 hours after B. cinerea infection (24hpi). Before infection (Figure 5; Supplementary Tables S2, S3), the most noticeable changes induced by PsJN were in genes related to signaling, proteolysis and cell wall. While in roots the most of the signaling-associated genes (like E3 ubiquitin-protein ligases and proteases of different types) were downregulated, in leaves PsJN inoculation led to an opposite effect. PsJN altered the expression level of transcription factors involved in biotic stress responses belonging to ERF, bZIP, WRKY, MYB and DOF families. Genes related to phytohormone signaling pathways were also affected by bacterial inoculation. There is a noticeable tendency for upregulation of genes involved in ethylene pathway in leaves in similar way as for ERF transcription factors, while in roots the expression of different genes involved in ethylene regulation were differently altered. Interestingly, while both up and downregulation of cell wall-related genes was observed in leaves, in roots PsJN predominantly led to a decreased level of their expression. A prominent influence of PsJN was noticed on the expression of genes involved in proteolysis in both roots and leaves, more conspicuously, though, in roots. Pathogenesis related (PR) genes were mostly downregulated in PsJN inoculated roots, while more upregulated PR genes were found in leaves of the same plants.
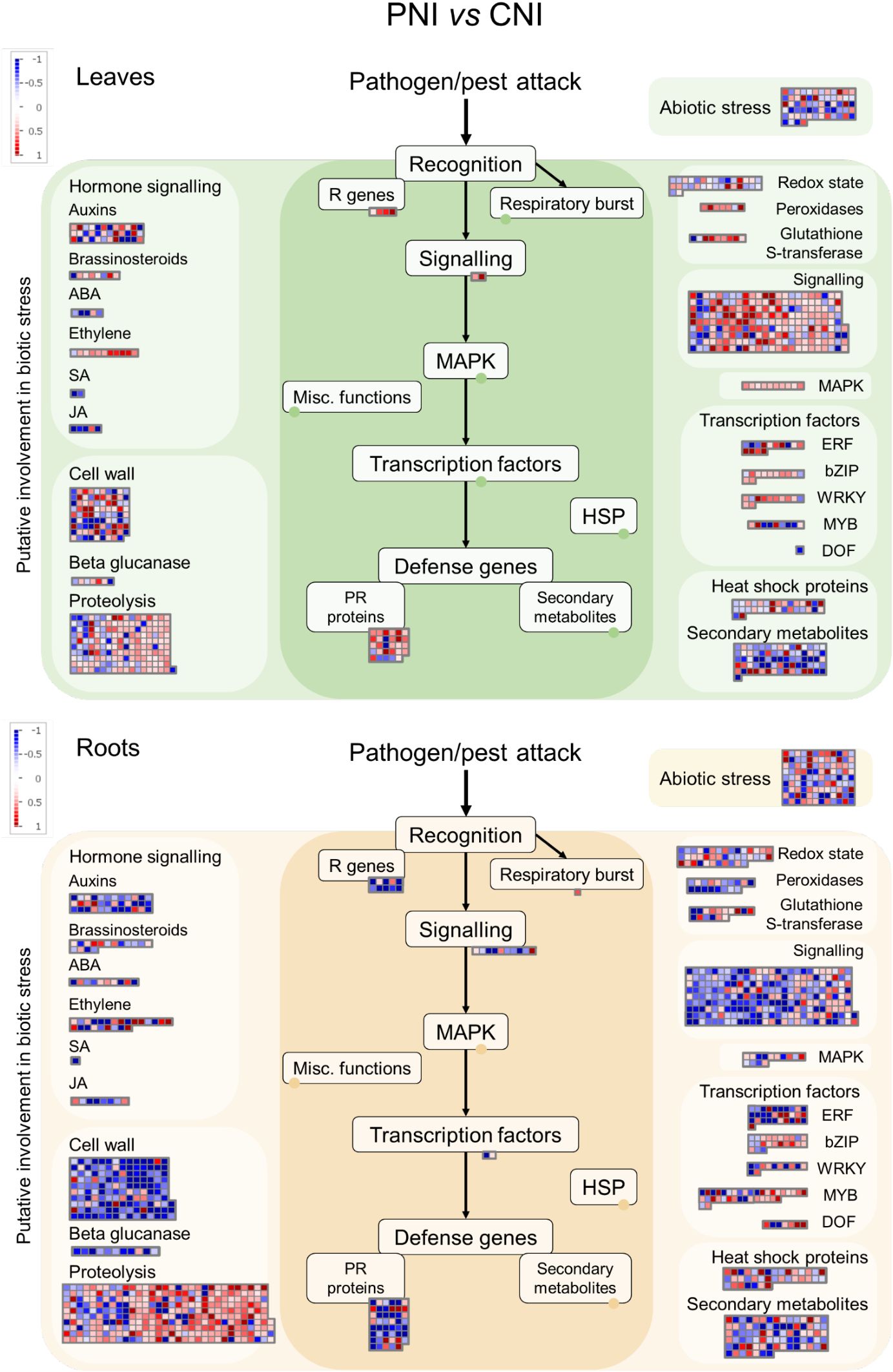
Figure 5. MapMan analysis of genes differentially expressed in roots and leaves of Arabidopsis in response to PsJN before B. cinerea infection. The green panels indicate leaves, while the beige panels represent roots.
Among the much smaller number of PsJN-influenced genes in plants 24 hours after B. cinerea infection, we still notice stimulation of ERF transcription factors in leaves (Figure 6; Supplementary Tables S4, S5). Also, although fewer cell wall-related genes were affected they are predominantly decreased in roots, while some of the genes were upregulated in leaves.
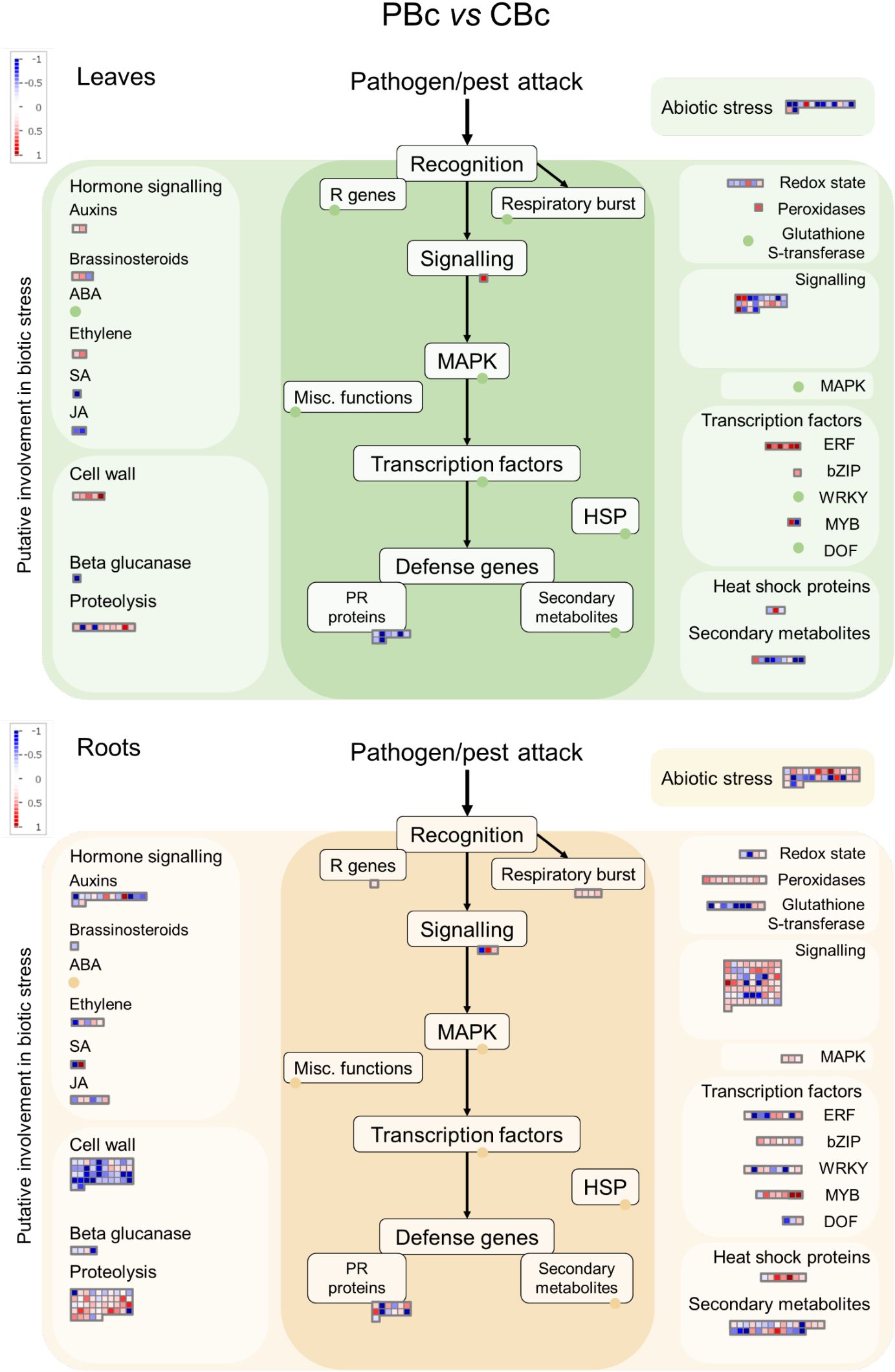
Figure 6. MapMan analysis of genes differentially expressed in in roots and leaves of Arabidopsis response to PsJN 24 hours after B. cinerea infection. The green panels indicate leaves, while the beige panels represent roots.
3.5 Small RNA analysis
To explore whether PsJN inoculation influences miRNA expression in A. thaliana under hydroponic conditions and during B. cinerea infection, small RNA from leaves and roots, in the four different treatments, were sequenced. An average of 30 million clean reads was generated for each sRNA library. Of that, 19.2 million reads were 18–30 nt in length. Small RNA identified as micro-RNA were further analyzed.
3.6 Identification of known and potentially novel miRNAs
In leaves, a total of 307 miRNAs were detected belonging to 109 miRNA families, while 28 of detected miRNAs were novel miRNAs. In roots, a total of 228 miRNAs belonging to 96 miRNA families were detected during analysis, 15 of which were novel miRNA. Families of miRNAs with most miRNAs detected were MIR156 and MIR159 in both leaves and roots.
3.7 miRNA expression patterns
Micro RNA expression was barely affected by PsJN inoculation, in both non-infected and B. cinerea-infected plants (Supplementary Table S6).
In non-infected leaves only one miRNA (ath-miR8183) was significantly upregulated (more than four-fold). Predicted targets of this miRNA are related to protein degradation and DNA synthesis. In roots, there were no miRNAs with significantly changed expression.
During B. cinerea infection, PsJN inoculation led to a significant change of only two miRNAs in leaves. Ath-miR163 and ath-miR845a were downregulated; ath-miR845a was lowered more than two-fold, while ath-miR163 was affected to a lesser extent. Putative targets of these miRNAs are related to phytohormone metabolism (especially for salicylic acid, SA), DNA synthesis, posttranslational modification of proteins and cell division. In roots, only one miRNA (ath-miR161.1) was upregulated and two miRNAs (ath-miR158a-3p and ath-miR159c) were downregulated. Genes involved in transcription regulation were recognized as the potential targets of these miRNAs.
To get an insight into the changes in the miRNA expression pattern induced by B. cinerea infection, regardless of PsJN inoculation, we analyzed miRNAs differentially expressed in plants infected with B. cinerea (CBc) or mock infected (CNI). In leaves, twenty miRNAs were significantly (corrected P-value < 0.05) differentially expressed, half of them upregulated and the other half downregulated upon infection (Supplementary Table S6). Five of the upregulated miRNAs (ath-miR157a-3p, ath-miR161.2, ath-miR168b-3p, ath-miR169f-3p and ath-miR858b) and five of the downregulated (ath-miR161.1, ath-miR1888a, ath-miR5020b, ath-miR5634 and ath-miR863-5p) were more than two-fold changed. One miRNA ath-miR858b, was highly upregulated (more than eight-fold).
In roots, only one miRNA (ath-miR398b-5p) was significantly differentially expressed, but its downregulation in infected plants was very remarkable, more than 77-fold.
Predicted targets of miRNAs differentially expressed after B. cinerea infection are mainly involved in the regulation of transcription and protein synthesis and degradation. Putative target genes are also involved in development, stress responses, RNA processing, cell wall biogenesis and organization, regulation of cellular component biogenesis and phenylpropanoid metabolic process. Targets of ath-miR398b-5p, which is highly downregulated in roots, are mainly involved in RNA regulation of transcription, transport and signaling.
B. cinerea infection also affected miRNA expression in plants previously inoculated with PsJN (PBc vs PNI). Twenty miRNAs were differentially expressed in leaves, while there were no significantly changed miRNAs in roots (Supplementary Table S6). One of the upregulated miRNAs, ath-miR398b-5p was highly expressed, more than eight-fold, while the expression of ath-miR157a-3p, ath-miR161.2, ath-miR167c-3p, ath-miR169f-3p and ath-miR824-5p was decreased more than two-fold. Other miRNAs, including ath-miR5020b, ath-miR5634 and ath-miR8183 were also downregulated more than two-fold in the leaves of PsJN inoculated plants after B. cinerea infection.
Predicted targets of miRNAs altered in response to B. cinerea infection (PBc vs PNI), are involved in signaling, defense, transcription regulation, proteolysis and hemicellulose synthesis.
3.8 Co-expression analysis of miRNA-mRNA pairs
To investigate miRNA-mRNA regulatory mechanisms, we performed an integrative analysis of transcriptome and small RNA sequencing data. A total of 487 genes were predicted as targets of differentially expressed miRNAs. Most miRNAs were predicted to target multiple genes, with both positive and negative miRNA-mRNA interactions observed. For example, the upregulated ath-miR396b-5p was shown to downregulate five of its targets, while three targets were upregulated.
In leaves, of 108 miRNA-mRNA pairs examined, 30 exhibited opposite gene expression patterns, all of which were detected in response to B. cinerea (CBc vs CNI and PBc vs PNI treatment comparisons, Figures 7, 8). In both comparisons, higher expression of ath-miR157a-3p was associated with lower expression of two target genes: NET1A (KIP1-like protein MYB-related) and OHP2 (ONE-HELIX PROTEIN2). In the CBc vs CNI comparison (Figure 7), ath-miR168a-3p and its target gene BDG4 (lysophospholipase BODYGUARD 4) showed an inverse expression pattern. The integrated analysis also revealed possible interactions between miRNAs and transcription factors. For example, ath-miR858b was highly expressed in the CBc treatment, while its targets belonging to the MYB family of transcription factors were downregulated. Similarly, an inverse expression of ath-miR158b and its target, the transcription factor bHLH94 (basic Helix-Loop-Helix), was observed under the same treatment.
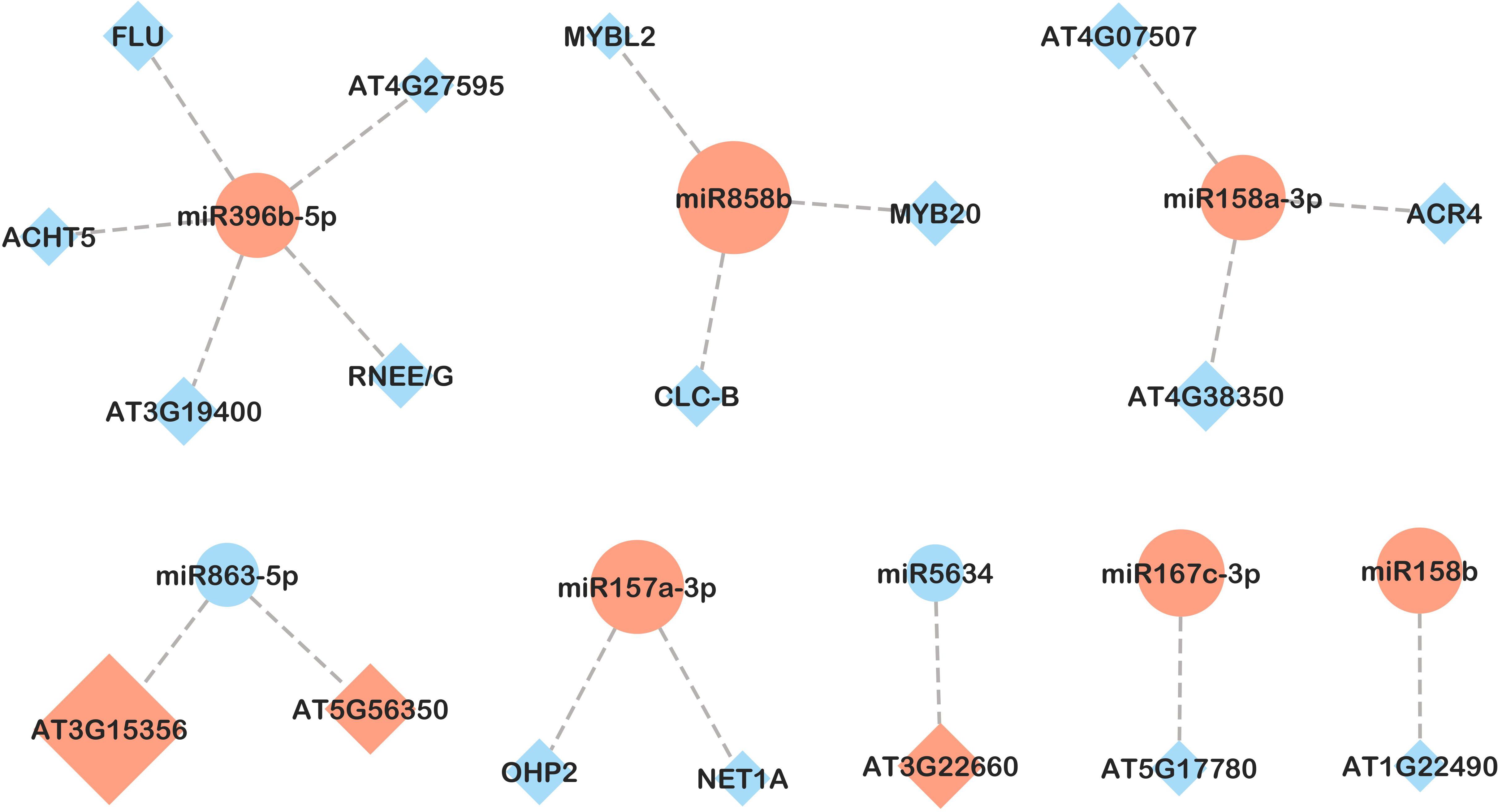
Figure 7. miRNA-mRNA network in leaves in response to B. cinerea infection (CBc vs CNI comparison). Candidate miRNA-mRNA target pairs including 8 differentially expressed miRNAs and 18 differentially expressed genes were integrated in the network. Diamonds represent mRNAs and circles represent miRNAs. Upregulated and downregulated genes are colored in red and blue, respectively. The size of the pattern represents the fold change in expression between CBc and CNI (miRNA-mRNA pairs with only inverse expression pattern are shown).
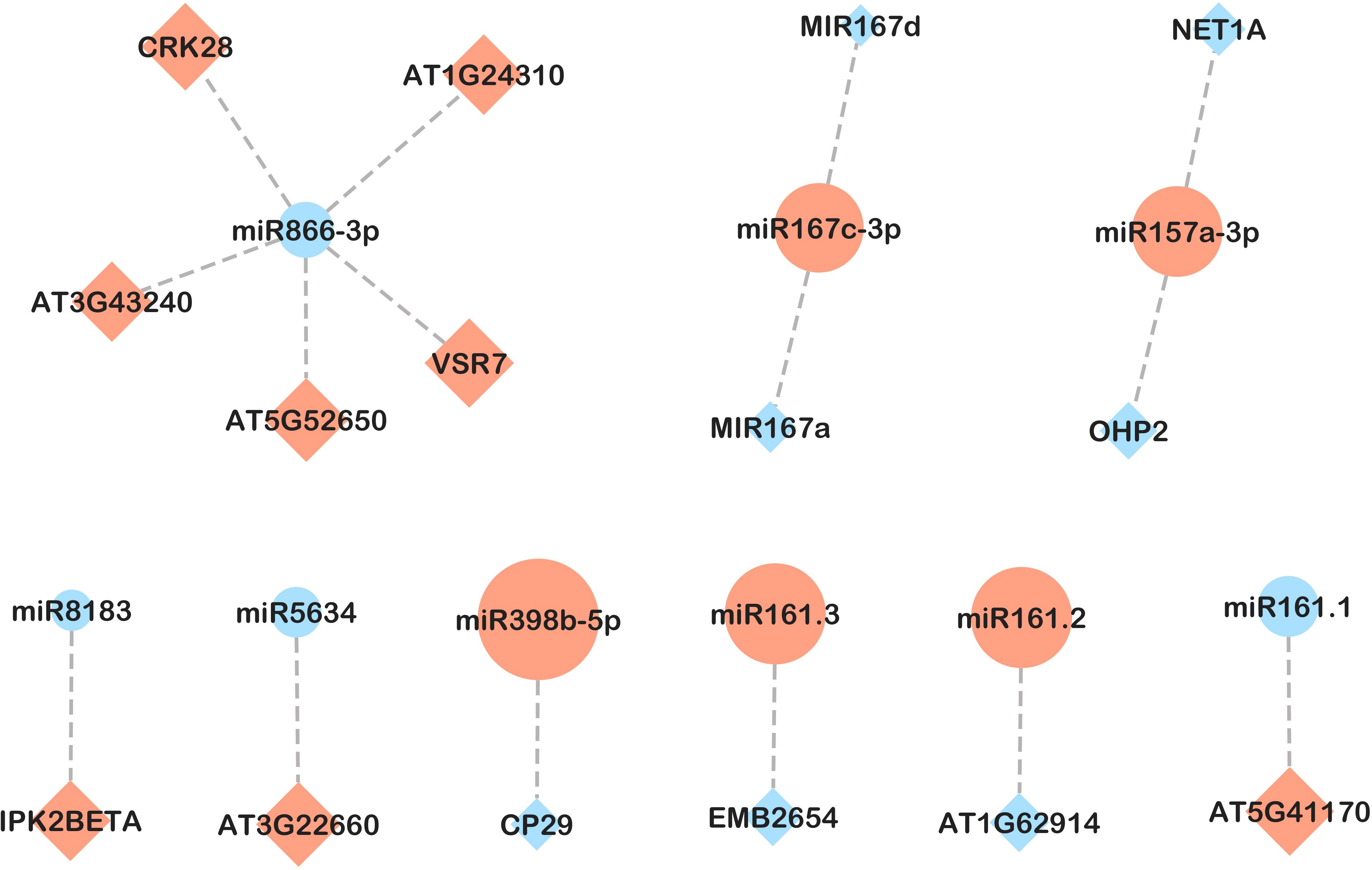
Figure 8. miRNA-mRNA network in leaves in response to B. cinerea infection in plants inoculated with PsJN (PBc vs PNI comparison). Candidate miRNA-mRNA target pairs including 9 differentially expressed miRNAs and 15 differentially expressed genes were integrated in the network. Diamonds represent mRNAs and circles represent miRNAs. Upregulated and downregulated genes are colored in red and blue, respectively. The size of the pattern represents the fold change in expression between PBc and PNI (miRNA-mRNA pairs with only inverse expression pattern are shown).
Regarding response to B. cinerea infection in plants inoculated with PsJN (PBc vs PNI, Figure 8), an inverse expression pattern was shown for ath-miR161.1 and ath-miR161.2 and members of the pentatricopeptide repeat (PPR-like) superfamily of proteins. Of the six predicted targets of ath-miR398b-5p, only CP29A (chloroplast RNA-binding protein 29) exhibited an inverse expression pattern. Additional miRNA-target pairs with inverse expression trends included ath-miR5634 and rRNA-processing protein EBP2, as well as ath-miR8183 and inositol polyphosphate kinase 2 beta (Ipk2β). Notably, we detected a negative correlation between the expression of ath-miR866-3p and its predicted target, an antifungal-related gene CRK28, a cysteine-rich receptor-like protein kinase (RLK).
4 Discussion
The beneficial bacteria belonging to genera Pseudomonas, Burkholderia and Paraburkholderia were previously shown as effective in controlling the necrotrophic fungus B. cinerea in some host plants, including Arabidopsis (Nie et al., 2017), cucumber (Zhu et al., 2021), and grapevine (Miotto-Vilanova et al., 2016). In this study, we examined the efficiency of P. phytofirmans PsJN in inducing systemic resistance to B. cinerea and promoting plant growth in A. thaliana. Our data also support the capacity of PsJN to reduce B. cinerea symptoms and improve plant growth when it was applied at the root level of A. thaliana grown both in soil and in hydroponics (Figure 1). This protective effect indicates that PsJN triggers systemic resistance against B. cinerea without compromising plant fitness. These beneficial effects could be related to a PsJN-induced priming state of the plants. However, insights about the mechanisms involved in this resistance state remain unclear so far. In this study, we used RNA-Seq sequencing and hydroponic system to investigate changes in gene expression in both roots and leaves, since most of the previous studies dealt only with processes occurring in leaves. We showed that PsJN induces transcriptional and miRNA reprogramming both locally (in roots) and systemically (leaves) before and after B. cinerea infection.
4.1 PsJN modulates the expression of abiotic stress response genes
The comparative transcriptome GO analysis of the PsJN-inoculated and control plants, revealed upregulation of pathways related to responses to abiotic stimuli, low oxygen levels, nutrient levels and low light intensity (Figure 2). Among the DEGs related to abiotic stresses we identified a significant number of genes related to hypoxia response. PsJN induced alterations in genes involved in oxygen sensing and signaling, hypoxia-related transcription regulators and some of the effector genes required for hypoxia response and tolerance (Figures 3, 4). This could be related to hydroponic plant growth conditions or to the pre-exposure of the plants to high humidity before B. cinerea infection. Ethylene has been shown to play a significant role in helping plants to cope with hypoxia, particularly during flooding stress (Liu et al., 2022). The expression patterns of genes associated with ethylene synthesis, sensing and signaling were modulated by PsJN inoculation in hydroponic conditions. PsJN affected the expression of numerous ERFs (Ethylene Responsive Factors), known as regulators of various ethylene-responsive genes and other important functions linked to abiotic stresses (Figure 3). Also, enzymes involved in plant adaptation to low oxygen conditions (PDC2, LDH1, AlaAT1, GLYI7, SRO4, FBA6, SUS1 and SUS4) were found upregulated, suggesting activation of alternative metabolic pathways for energy production and maintaining cellular functions (Figure 4).
Overall, PsJN inoculation positively influences various genes and pathways involved in response to hypoxia. It improves ethylene pathways and activates fermentation pathways to sustain energy production and optimize metabolic processes.
4.2 PsJN modulates expression of genes involved in microbe-plant interactions and ISR
Although most beneficial bacteria evade and suppress the local immune response to successfully colonize plant tissues, they modulate systemic plant defenses in a complex way to fight subsequent pathogens through induced systemic resistance (ISR). In this study, while the transcriptome was significantly altered in response to PsJN in non-infected plants, we detected only limited number of PsJN-modulated genes after 24 hours of B. cinerea infection (Figure 6; Supplementary Tables S4, S5). Although no necrotic lesions were visible at that time, gene expression had already been changed tremendously as a response to the pathogen challenge. Nevertheless, the increased expression of defense-related genes in the leaves prior to infection, combined with the protective effect of PsJN visible at later stages of infection, indicates that bacterial inoculation primes the plants for subsequent pathogen attack.
The most noticeable changes induced by PsJN in A. thaliana in our experiments, were in genes related to signaling, ethylene pathway, proteolysis and cell wall (Figure 5). Perception of microbes and signal transduction are crucial for a plant’s ability to shape its microbiome and to distinguish beneficial from pathogenic microorganisms. Among the genes involved in signaling, PsJN influenced a considerable number of receptor kinases and calcium binding proteins (Supplementary Tables S2, S3). Owing to their great diversity, receptor-like kinases (RLKs) regulate a wide range of vital processes in plants, including development and abiotic and biotic stress responses (Jose et al., 2020). We found both increased and decreased signaling genes in aboveground as well as in underground plant parts; however, they were predominantly down-regulated in roots, while mostly being upregulated in leaves. It is interesting to note that a loss of a plant receptor kinase recruits PGPB Pseudomonas fluorescens in Arabidopsis roots (Song et al., 2021). Although mutualists and pathogens share some of the microbe-associated molecular patterns (MAMPs) recognized by RLK, there are also epitopes specific to certain microbes. By combining the perception of different epitope variants, effectors, damage-associated molecular patterns (DAMPs) and other signals, plants can distinguish microorganisms and respond appropriately (Zhang and Kong, 2022). We can assume that the ability of a PGPB to modulate the expression of receptor kinases and other signaling genes, could facilitate colonization of the root, on the one hand, and alter the overall plant perception and response to microorganisms, on the other.
Effective plant immune reactions rely not only on de novo synthesis of defense proteins but also on their post-translational modifications and regulated degradation. Post-translational modifications enable rapid and dynamic responses to external stimuli and play an important role in plant immunity (Sharma et al., 2021). Increasing evidence supports a positive role of SUMOylation in plant immunity. This post-translational modification regulates the functioning of the plant immune response, since components of the defense signaling pathway are recognized as SUMO targets. On the other hand, pathogens target host SUMOylation to manipulate plant defense. A positive correlation between SUMOylation and fungal resistance has been found (Li et al., 2013a; Colignon et al., 2017). In our study, a remarkable increase in the expression of SAE1B, the SUMO-activating enzyme 1B (19-fold) was detected in leaves of PsJN-inoculated plants, suggesting a role of SUMOylation in the protective effect of PsJN against B. cinerea infection (Supplementary Table S1).
The ubiquitin–proteasome system (UPS) regulates all stages of plant immunity. On the other hand, pathogen effectors interfere with the plant UPS by hijacking its functions (Langin et al., 2023). Role of UPS in the establishment of mutualistic plant-microbe interactions is also proposed, however, studies addressing this question are very limited. It was shown that endophytic colonization of A. thaliana with a PGPB strain Kosakonia radicincitans interferes with ubiquitin-dependent protein degradation, while increasing the protein expression of 20S proteasome alpha-3 subunit (Witzel et al., 2017). Also, Dudnik et al. (2014) reported that endophyte Rhizobium sp. strain AP16 synthesizes a proteasome inhibitor syringolin A. Very complex interplay in plant-microbe interactions related to proteolytic pathway include microbe-induced targeting of immune proteins for degradation, as well as impairment of protein turnover required for adequate immune response (Langin et al., 2023). Transcriptome changes do not always reflect protein levels or their functionality; however, alterations in the expression of UPS and proteolysis related genes, provoked by PsJN inoculation, indicate that this strain modulates action of plant proteolytic pathway (Figure 5; Supplementary Tables S2, S3).
Among the defense-related genes, we detected a PsJN-dependent downregulation of pathogenesis-related (PR) proteins and beta glucanases in roots, while several PR genes were up or downregulated in leaves. These proteins are indispensable for plant defense and their expression increases when they encounter pathogenic invaders. Some studies reported an increased accumulation of PR proteins in PGPB-inoculated plants as part of the ISR mechanism of plant protection (van de Mortel et al., 2012; Leandro et al., 2019). However, others found PGPB-induced downregulation of PR proteins, and it has been suggested that suppression of some factors of the plant defense response may serve to facilitate colonization of plant tissues (Brusamarello-Santos et al., 2012; Thomas et al., 2019; Soares et al., 2024).
Both pathogenic and beneficial microorganisms have been found to influence cell wall composition, to facilitate their attachment and colonization. Downregulation of genes related to cell wall formation and modification has previously been observed in Arabidopsis and rice. It has been suggested that PGPB may suppress modifications of root cell walls, to support early stages of root colonization (Lakshmanan et al., 2013; Rekha et al., 2018). The effect of PGPB on root growth depends on bacterial strain and titer, plant species, developmental stage and growth conditions. In our experimental setup, most cell wall-related enzymes and proteins were downregulated in PsJN-inoculated roots. Transcriptome analysis was performed on plants several weeks after the first inoculation; however, it can be assumed that the hydroponic growth conditions and the lowered oxygen content that prevailed during the 48 hours before sampling also have an influence on the gene expression patterns.
While receptor kinases and calcium binding proteins, peroxidases, cell wall-related proteins and PR proteins, were mostly found downregulated in the PsJN-inoculated roots, their expression in leaves was predominantly upregulated (RLK, peroxidases and PR proteins), or differently regulated (for different cell wall proteins). Also, both increase and decrease of various transcription factors and hormone pathways related to biotic stimuli response were detected in PsJN-inoculated roots, while in leaves their upregulation is prevalent, especially regarding ethylene (ET) dependent pathway (Figure 5; Supplementary Tables S2, S3). These transcriptome alterations in leaves before occurrence of pathogen are in line with the PsJN-induced alleviation of disease symptoms after B. cinerea infection (Figure 1). They underline a priming state of plant for faster and stronger responses upon pathogen attack, i.e. ISR.
Both local and systemic changes in expression of ET pathway genes were detected in response to PsJN-inoculation (Figure 3). The altered expression of ET pathway genes in the roots during beneficial interactions suggests that ET signaling plays a role not only in defense against pathogens, but also in recognizing beneficial endophytic microbes, potentially regulating root colonization (Shekhawat et al., 2023). ET can act as both positive and negative regulator depending on the microbial strain and the stage of interaction. Some beneficial microbes require ET for plant growth promotion in stressful conditions (Shekhawat et al., 2021). On the other hand, some beneficial bacteria produce ACC deaminase, which lowers ET levels to alleviate plant stress and to promote plant growth by suppressing the ET-dependent plant defense system (Glick, 2014). This suggests that the effect of ET on plant-microbe interactions is concentration-dependent, and a finely regulated ET pathway is crucial for establishing beneficial interactions and enhancing protective effects (Shekhawat et al., 2023).
PsJN is found to both reduce ET level locally, through the activation of ACC deaminase (Sun et al., 2009), and to upregulate ET synthesis genes in both roots and shoots (Pinedo et al., 2015). It was also shown that plant growth promotion by PsJN is ET- dependent in Arabidopsis, since this effect was lost in ein2–1 mutants (Poupin et al., 2016). Thus, it is suggested that PsJN finely regulates ET to induce beneficial effects in plants. We found that in roots PsJN differently regulates different ET-related genes, while in leaves they were mostly upregulated. Taken together with the finding that PsJN mostly downregulated genes related to microbe recognition, signaling and PR in roots, we suppose that attenuation of immune response and modulation of ET pathway could serve to facilitate root colonization and the establishment of mutualistic relationship.
In leaves, the upregulation of ET pathway and signaling genes indicates PsJN-induced systemic response as a preparation of plant to the subsequent pathogen attack. It is also worth noting that after B. cinerea infection among a small number of DEGs observed in PsJN-inoculated plants we notice ET-related genes, all upregulated, indicating an important role of ET in the PsJN-mediated ISR in Arabidopsis. This is in accordance with the renowned importance of this phytohormone for plant resistance to necrotrophic pathogens (Ghozlan et al., 2020).
Along with ET, some PGPB were found to stimulate the JA pathway, which is required for defense-related gene expression in plants. We noticed that PsJN increases the expression of ERF1 which is known to integrate signals from both JA and ET pathways (Lorenzo et al., 2003). However, none of the enzymes involved in JA biosynthesis were found to be differentially expressed in PsJN-inoculated plants 24h after B. cinerea infection.
4.3 B. cinerea triggered transcriptome and sRNAome changes in Arabidopsis
Transcriptome analysis of Arabidopsis infected with B. cinerea provides valuable insights into the plant’s defense mechanisms and the pathogen’s infection strategies. As previously reported, the host plant undergoes extensive transcriptional reprogramming during B. cinerea infection (Wei et al., 2024). Our research confirmed transcriptional changes in metabolic pathways, signaling cascades, and transcriptional regulation in Arabidopsis plants after B. cinerea infection (Figure 6; Supplementary Tables S4, S5).
The understanding of how Arabidopsis roots respond to leaf B. cinerea infection is also of relevant importance for elucidating the complex signaling pathways that govern plant defense mechanisms, but was usually neglected. In our pathosystem almost 300 genes were significantly upregulated and about 600 downregulated in roots upon B. cinerea infection. They are involved in several important processes: ET biosynthesis and regulation, sugar metabolism and stress defense. Among the ET-related genes, the ACO3 (1-Aminocyclopropane-1-Carboxylate Oxidase 3) gene exhibited the most significant difference in expression compared to non-infected plants. This gene plays a crucial role in the final step of ET biosynthesis in plants. ET-responsive transcription factors such as ERF15, ERF35, ERF73, RAP2.9, and ORA59 were also upregulated in roots, suggesting a key role of ET in the response to pathogen attack in both affected plant organs and distant below-ground parts. We also identified other upregulated genes involved in sugar metabolism in response to B. cinerea infection, such as SUS3 (sucrose synthase), and SWEET11 and SWEET12 (sugar transporters). It has previously been reported that SWEET11 and SWEET12 transporters accumulate in the roots under stress conditions, facilitating the unloading of sucrose from the apoplast to the sink cells in the roots (Fatima et al., 2022). This process promotes root growth by reallocating more sucrose from the leaves to the roots (Durand et al., 2016). Additionally, SWEET transporters regulate sugar supply from shoots to roots and then deliver sugars to plant growth-promoting rhizobacteria (Desrut et al., 2021). Metallothionein MT2 gene was also upregulated and it is particularly important in stress responses, including oxidative stress and metal homeostasis (Liu et al., 2022).
In addition to transcriptome changes, the miRNAome is frequently altered under stress conditions. Our results also identified two mRNA-miRNA pairs that may be involved in biotic stress responses. In plants infected with B.cinerea, ath-miR168a-3p was upregulated, while its target, BDG4 (lysophospholipase BODYGUARD 4) was downregulated. Interestingly, ath-miR168a-3p was also found to be upregulated in Arabidopsis infected with another necrotrophic fungal pathogen, Sclerotinia sclerotiorum (Cao et al., 2020). BDG4 has been shown to be linked to cuticle-related resistance and plant defense. Arabidopsis plants with altered cuticular structures exhibited total immunity to B. cinerea (Bessire et al., 2007), while bdg4 mutants displayed increased resistance to B. cinerea (Chassot et al., 2007; Voisin et al., 2009; Aragón et al., 2021). Beyond its role as a physical barrier, the cuticle is likely involved in signaling, and BDG4 as an integral component might play a significant role in plant-pathogen interactions, particularly against B. cinerea, contributing to plant resistance (Aragón et al., 2021). Our findings suggest that ath-miR168a-3p may act as a signaling event that triggers a cascade-related immune response to enhance plant resistance to B. cinerea.
The effect of B. cinerea infection in PsJN-inoculated plants is marked by downregulation of ath-miR866-3p and a subsequent upregulation of the antifungal-related gene CRK28, a cysteine-rich receptor-like protein kinase. In Arabidopsis, CRK28 plays an essential role in integrating various environmental and developmental signals to regulate plant stress responses, particularly those involving abscisic acid signaling (Pelagio-Flores et al., 2019). CRK28 interacts with other components to form a pattern-recognition receptor (PRR) immune complex, which is crucial for triggering pattern-triggered immunity (PTI), including ROS production and programmed cell death. Constitutive overexpression of AtCRK28 enhanced resistance to the hemi-biotrophic bacterial pathogen Pseudomonas syringae pv. tomato DC3000 (Yadeta et al., 2017). In contrast, crk28 mutants showed severe disease symptoms after infection with Pst DC3000 and were more susceptible to biotrophic fungal pathogens like Golovinomyces orontii (Zeiner et al., 2023; Zhang et al., 2023). Our results suggest that ath-miR866-3p may be involved in pathogen responses, potentially enhanced by the beneficial bacterium PsJN.
Two miRNAs in B. cinerea-infected plants, ath-miR158b and ath-miR858b, target transcription factors involved in responses to hypoxic stress. Ath-miR158b downregulated bHLH94, a transcription factor belonging to the bHLH family, the second largest family of plant transcription factors after MYB. It has been shown that bHLH94 was downregulated under both waterlogging and osmotic stress (Pereira-Santana et al., 2017; Nuanlaong et al., 2021). The bHLH family is particularly important in JA signaling pathways and JA-dependent responses such as flavonoid production and defense against herbivores and pathogens (Goossens et al., 2017).
Moreover, our study found that ath-miR858b targets MYB family transcription factors, which regulate various secondary metabolic pathways. MYB-like 2 and MYB20 were downregulated, while MYB13 and MYB37 were upregulated in CB compared to CNI. miR858 is commonly associated with flavonoid-specific MYB transcription factors and is involved in resistance to necrotrophic and hemibiotrophic pathogens in Arabidopsis (Sharma et al., 2016; Camargo-Ramírez et al., 2018).
Several miRNAs identified in our study have been previously linked to drought or waterlogging responses. Ath-miR157a-3p was upregulated in both non-inoculated and PsJN-inoculated plants after B. cinerea infection (Supplementary Table S6). This miRNA was also reported under drought conditions in alfalfa (Li et al., 2017). In Arabidopsis, its target gene OHP2 (ONE-HELIX PROTEIN 2), which was downregulated in both CB and PB plants, is essential for light acclimation and photosynthesis (Beck et al., 2017).
In the PB vs. PNI comparison group, we found that ath-miR161.1 and ath-miR161.2 were altered, with their target genes belonging to the Pentatricopeptide Repeat (PPR) protein superfamily being both upregulated and downregulated. PPR proteins are involved in various aspects of RNA metabolism, while some PPRs play important roles in biotic and abiotic stress responses in Arabidopsis and other species (Laluk et al., 2011; Xing et al., 2018).
5 Concluding remarks
The capacity of PsJN in inducing ISR against B. cinerea was shown here on A. thaliana grown both in soil and in hydroponic conditions. Comparative transcriptome analysis revealed that PsJN induced differential expression of genes involved in abiotic and biotic stress responses, with the ET pathway-related genes being conspicuous in both cases. Prior to pathogen attack, PsJN modulated expression of genes involved in abiotic stress responses, microbe-plant interactions and ISR. In roots, PsJN predominantly downregulated the expression of genes related to microbe perception, signaling and immune response, indicating that PsJN locally provoked attenuation of defense responses to facilitate and support colonization and the maintenance of mutualistic relationship. In leaves, the increased expression of defense-related genes, including ET pathway genes, prior to infection, in combination with the protective effect of PsJN that is evident in later stages of infection, suggests that bacterial inoculation primes plants for enhanced systemic immune response after subsequent pathogen attack. Since it is important for the host to increase its combat readiness in advance of a disease outbreak, we hypothesize that the transcriptome changes in PsJN-treated Arabidopsis plants prior to infection enabled a more rapid response at the metabolic level when the pathogen emerged. Analyses of the proteome and post-translational modifications would provide more precise information on the role of the different defense-related genes and miRNA and their interplay in PsJN-induced resistance in A. thaliana against B. cinerea.
Data availability statement
The data discussed in this publication has been deposited in the SRA database and can be accessed through accession number PRJNA1177658 (https://www.ncbi.nlm.nih.gov/sra/PRJNA1177658).
Author contributions
SN: Conceptualization, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. DB: Formal analysis, Investigation, Writing – original draft. JS: Formal analysis, Writing – original draft. IN: Investigation, Writing – review & editing. AA: Methodology, Writing – review & editing. DN: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, and/or publication of this article. This research was supported by the Science Fund of the Republic of Serbia, 7744906 GRANT No, Exploring Bacterial OMV (Outer Membrane Vesicles)-sRNAs Mediated Interkingdom Communication with Plants and Fungi -ExplOMV and the Ministry of Science, Technological Development and Innovation of the Republic of Serbia (Contract 51-03-136/2025-03/200042).
Acknowledgments
The authors acknowledge Simon Duchateau and Sandra Villaume (RIBP) for providing the B. cinerea strain used in this study as well as sharing their experience and knowledge of working with this phytopathogen.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1554036/full#supplementary-material
References
Aragón, W., Formey, D., Aviles-Baltazar, N. Y., Torres, M., and Serrano, M. (2021). Arabidopsis thaliana Cuticle Composition Contributes to Differential Defense Response to Botrytis cinerea. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.738949
Axtell, M. J. and Bowman, J. L. (2008). Evolution of plant microRNAs and their targets. Trends Plant Sci. 13, 343–349. doi: 10.1016/j.tplants.2008.03.009
Aziz, A., Poinssot, B., Daire, X., Adrian, M., Bézier, A., Lambert, B., et al. (2003). Laminarin Elicits Defense Responses in Grapevine and Induces Protection Against Botrytis cinerea and Plasmopara viticola. Mol. Plant-Microbe Interact. 16, 1118–1128. doi: 10.1094/MPMI.2003.16.12.1118
Beck, J., Lohscheider, J. N., Albert, S., Andersson, U., Mendgen, K. W., Rojas-Stütz, M. C., et al. (2017). Small one-helix proteins are essential for photosynthesis in arabidopsis. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00007
Bessire, M., Chassot, C., Jacquat, A., Humphry, M., Borel, S., Petétot, J. M., et al. (2007). A permeable cuticle in Arabidopsis leads to a strong resistance to Botrytis cinerea. EMBO J. 26, 2158–2168. doi: 10.1038/sj.emboj.7601658
Bi, K., Liang, Y., Mengiste, T., and Sharon, A. (2023). Killing softly: a roadmap of Botrytis cinerea pathogenicity. Trends Plant Sci. 28, 211–222. doi: 10.1016/j.tplants.2022.08.024
Borges, F. and Martienssen, R. A. (2015). The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell Biol. 16, 727–741. doi: 10.1038/nrm4085
Brusamarello-Santos, L. C. C., Pacheco, F., Aljanabi, S. M. M., Monteiro, R. A., Cruz, L. M., Baura, V. A., et al. (2012). Differential gene expression of rice roots inoculated with the diazotroph Herbaspirillum seropedicae. Plant Soil 356, 113–125. doi: 10.1007/s11104-011-1044-z
Camargo-Ramírez, R., Val-Torregrosa, B., and San Segundo, B. (2018). MiR858-mediated regulation of flavonoid-specific MYB transcription factor genes controls resistance to pathogen infection in arabidopsis. Plant Cell Physiol. 59, 190–204. doi: 10.1093/pcp/pcx175
Cao, J.-Y., Xu, Y.-P., and Cai, X.-Z. (2020). Integrated miRNAome and Transcriptome Analysis Reveals Argonaute 2-Mediated Defense Responses Against the Devastating Phytopathogen Sclerotinia sclerotiorum. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00500
Cartieaux, F., Contesto, C., Gallou, A., Desbrosses, G., Kopka, J., Taconnat, L., et al. (2008). Simultaneous Interaction of Arabidopsis thaliana with Bradyrhizobium Sp. Strain ORS278 and Pseudomonas syringae pv. tomato DC3000 Leads to Complex Transcriptome Changes. Mol. Plant-Microbe Interact. 21, 244–259. doi: 10.1094/MPMI-21-2-0244
Chassot, C., Nawrath, C., and Métraux, J.-P. (2007). Cuticular defects lead to full immunity to a major plant pathogen. Plant J. 49, 972–980. doi: 10.1111/j.1365-313X.2006.03017.x
Colavolpe, M. B., Villarreal, N. M., Langer, S. E., Romero, F. M., Martínez, G. A., Saini, A., et al. (2021). Burkholderia sp. Strain AU4i promotes arabidopsis growth and increases plant defence response to necrotrophic pathogens. J. Plant Growth Regul. 40, 1939–1949. doi: 10.1007/s00344-020-10238-6
Coleman, N. V., Mattes, T. E., Gossett, J. M., and Spain, J. C. (2002). Biodegradation of cis-dichloroethene as the sole carbon source by a β-proteobacterium. Appl. Environ. Microbiol. 68, 2726–2730. doi: 10.1128/AEM.68.6.2726-2730.2002
Colignon, B., Dieu, M., Demazy, C., Delaive, E., Muhovski, Y., Raes, M., et al. (2017). Proteomic Study of SUMOylation During Solanum tuberosum–Phytophthora infestans Interactions. Mol. Plant-Microbe Interact. 30, 855–865. doi: 10.1094/MPMI-05-17-0104-R
Conn, S. J., Hocking, B., Dayod, M., Xu, B., Athman, A., Henderson, S., et al. (2013). Protocol: optimising hydroponic growth systems for nutritional and physiological analysis of Arabidopsis thaliana and other plants. Plant Methods 9, 4. doi: 10.1186/1746-4811-9-4
Conrath, U., Beckers, G. J. M., Langenbach, C. J. G., and Jaskiewicz, M. R. (2015). Priming for enhanced defense. Annu. Rev. Phytopathol. 53, 97–119. doi: 10.1146/annurev-phyto-080614-120132
Desrut, A., Thibault, F., Mercado-Blanco, J., Coutos-Thévenot, P., and Vriet, C. (2021). Transcriptional regulation of plant sugar transporter genes by beneficial rhizobacteria. J. Plant Interact. 16, 443–451. doi: 10.1080/17429145.2021.1974582
Dudnik, A., Bigler, L., and Dudler, R. (2014). Production of proteasome inhibitor syringolin A by the endophyte Rhizobium sp. strain AP16. Appl. Environ. Microbiol. 80, 3741–3748. doi: 10.1128/AEM.00395-14
Durand, M., Porcheron, B., Hennion, N., Maurousset, L., Lemoine, R., and Pourtau, N. (2016). Water deficit enhances C export to the roots in Arabidopsis thaliana plants with contribution of sucrose transporters in both shoot and roots. Plant Physiol. 170, 1460–1479. doi: 10.1104/pp.15.01926
Esmaeel, Q., Miotto, L., Rondeau, M., Leclère, V., Clément, C., Jacquard, C., et al. (2018). Paraburkholderia phytofirmans psJN-plants interaction: from perception to the induced mechanisms. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.02093
Fatima, U., Anjali, A., and Senthil-Kumar, M. (2022). AtSWEET11 and AtSWEET12: The twin traders of sucrose. Trends Plant Sci. 27, 958–960. doi: 10.1016/j.tplants.2022.06.009
Friedländer, M. R., Mackowiak, S. D., Li, N., Chen, W., and Rajewsky, N. (2012). miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 40, 37–52. doi: 10.1093/nar/gkr688
Ghozlan, M. H., Eman, E.-A., Tokgöz, S., Lakshman, D. K., and Mitra, A. (2020). Plant defense against necrotrophic pathogens. Am. J. Plant Sci. 11, 2122–2138. doi: 10.4236/ajps.2020.1112149
Gibbs, D. J., Lee, S. C., Isa, N. M., Gramuglia, S., Fukao, T., Bassel, G. W., et al. (2011). Homeostatic response to hypoxia is regulated by the N-end rule pathway in plants. Nature 479, 415–418. doi: 10.1038/nature10534
Glick, B. R. (2014). Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 169, 30–39. doi: 10.1016/j.micres.2013.09.009
Goossens, J., Mertens, J., and Goossens, A. (2017). Role and functioning of bHLH transcription factors in jasmonate signalling. J. Exp. Bot. 68, 1333–1347. doi: 10.1093/jxb/erw440
Gruau, C., Trotel-Aziz, P., Villaume, S., Rabenoelina, F., Clément, C., Baillieul, F., et al. (2015). Pseudomonas fluorescens PTA-CT2 Triggers Local and Systemic Immune Response Against Botrytis cinerea in Grapevine. Mol. Plant-Microbe Interact. 28, 1117–1129. doi: 10.1094/MPMI-04-15-0092-R
Issa, A., Esmaeel, Q., Sanchez, L., Courteaux, B., Guise, J.-F., Gibon, Y., et al. (2018). Impacts of Paraburkholderia phytofirmans Strain PsJN on Tomato (Lycopersicon esculentum L.) Under High Temperature. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01397
Iwakawa, H. and Tomari, Y. (2013). Molecular Insights into microRNA-Mediated Translational Repression in Plants. Mol. Cell 52, 591–601. doi: 10.1016/j.molcel.2013.10.033
Jose, J., Ghantasala, S., and Roy Choudhury, S. (2020). Arabidopsis transmembrane receptor-like kinases (RLKs): A bridge between extracellular signal and intracellular regulatory machinery. Int. J. Mol. Sci. 21, 4000. doi: 10.3390/ijms21114000
King, E., Wallner, A., Guigard, L., Rimbault, I., Parrinello, H., Klonowska, A., et al. (2023). Paraburkholderia phytofirmans PsJN colonization of rice endosphere triggers an atypical transcriptomic response compared to rice native Burkholderia s.l. endophytes. Sci. Rep. 13, 10696. doi: 10.1038/s41598-023-37314-7
Kong, X., Yang, M., Le, B. H., He, W., and Hou, Y. (2022). The master role of siRNAs in plant immunity. Mol. Plant Pathol. 23, 1565–1574. doi: 10.1111/mpp.13250
Lakshmanan, V., Castaneda, R., Rudrappa, T., and Bais, H. P. (2013). Root transcriptome analysis of Arabidopsis thaliana exposed to beneficial Bacillus subtilis FB17 rhizobacteria revealed genes for bacterial recruitment and plant defense independent of malate efflux. Planta 238, 657–668. doi: 10.1007/s00425-013-1920-2
Laluk, K., Abuqamar, S., and Mengiste, T. (2011). The Arabidopsis mitochondria-localized pentatricopeptide repeat protein PGN functions in defense against necrotrophic fungi and abiotic stress tolerance. Plant Physiol. 156, 2053–2068. doi: 10.1104/pp.111.177501
Langin, G., González-Fuente, M., and Üstün, S. (2023). The plant ubiquitin–proteasome system as a target for microbial manipulation. Annu. Rev. Phytopathol. 61, 351–375. doi: 10.1146/annurev-phyto-021622-110443
Langmead, B., Trapnell, C., Pop, M., and Salzberg, S. L. (2009). Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 10, R25. doi: 10.1186/gb-2009-10-3-r25
Leandro, M. R., Rangel, P. L., dos Santos, T. C., Andrade, L. F., de Souza Vespoli, L., Rangel, A. L. S., et al. (2019). Colonization of Arabidopsis thaliana by Herbaspirillum seropedicae promotes its growth and changes its proteomic profile. Plant Soil 443, 429–447. doi: 10.1007/s11104-019-04236-1
Li, S., Ji, R., Dudler, R., Yong, M., Deng, Q., Wang, Z., et al. (2013a). Wheat gene TaS3 contributes to powdery mildew susceptibility. Plant Cell Rep. 32, 1891–1901. doi: 10.1007/s00299-013-1501-7
Li, S., Liu, L., Zhuang, X., Yu, Y., Liu, X., Cui, X., et al. (2013b). MicroRNAs inhibit the translation of target mRNAs on the endoplasmic reticulum in Arabidopsis. Cell 153, 562–574. doi: 10.1016/j.cell.2013.04.005
Li, Y., Wan, L., Bi, S., Wan, X., Li, Z., Cao, J., et al. (2017). Identification of drought-responsive microRNAs from roots and leaves of alfalfa by high-throughput sequencing. Genes 8, 119. doi: 10.3390/genes8040119
Liao, Y., Smyth, G. K., and Shi, W. (2014). featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930. doi: 10.1093/bioinformatics/btt656
Licausi, F., van Dongen, J. T., Giuntoli, B., Novi, G., Santaniello, A., Geigenberger, P., et al. (2010). HRE1 and HRE2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant J. 62, 302–315. doi: 10.1111/j.1365-313X.2010.04149.x
Liu, Z., Hartman, S., van Veen, H., Zhang, H., Leeggangers, H. A. C. F., Martopawiro, S., et al. (2022). Ethylene augments root hypoxia tolerance via growth cessation and reactive oxygen species amelioration. Plant Physiol. 190, 1365–1383. doi: 10.1093/plphys/kiac245
Lorenzo, O., Piqueras, R., Sánchez-Serrano, J. J., and Solano, R. (2003). ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 15, 165–178. doi: 10.1105/tpc.007468
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. doi: 10.1186/s13059-014-0550-8
Lugtenberg, B. and Kamilova, F. (2009). Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 63, 541–556. doi: 10.1146/annurev.micro.62.081307.162918
Middleton, H., Yergeau, É., Monard, C., Combier, J.-P., and El Amrani, A. (2021). Rhizospheric plant-microbe interactions: miRNAs as a key mediator. Trends Plant Sci. 26, 132–141. doi: 10.1016/j.tplants.2020.09.005
Miotto-Vilanova, L., Jacquard, C., Courteaux, B., Wortham, L., Michel, J., Clément, C., et al. (2016). Burkholderia phytofirmans PsJN Confers Grapevine Resistance against Botrytis cinerea via a Direct Antimicrobial Effect Combined with a Better Resource Mobilization. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.01236
Mortazavi, A., Williams, B. A., McCue, K., Schaeffer, L., and Wold, B. (2008). Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 5, 621–628. doi: 10.1038/nmeth.1226
Nguyen, N. H., Trotel-Aziz, P., Villaume, S., Rabenoelina, F., Schwarzenberg, A., Nguema-Ona, E., et al. (2020). Bacillus subtilis and Pseudomonas fluorescens Trigger Common and Distinct Systemic Immune Responses in Arabidopsis thaliana Depending on the Pathogen Lifestyle. Vaccines 8, 503. doi: 10.3390/vaccines8030503
Nie, P., Li, X., Wang, S., Guo, J., Zhao, H., and Niu, D. (2017). Induced Systemic Resistance against Botrytis cinerea by Bacillus cereus AR156 through a JA/ET- and NPR1-Dependent Signaling Pathway and Activates PAMP-Triggered Immunity in Arabidopsis. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00238
Nikolić, I., Samardžić, J., Stevanović, S., Miljuš-Đukić, J., Milisavljević, M., and Timotijević, G. (2023). CRISPR/cas9-targeted disruption of two highly homologous arabidopsis thaliana DSS1 genes with roles in development and the oxidative stress response. Int. J. Mol. Sci. 24, 2442. doi: 10.3390/ijms24032442
Nuanlaong, S., Wuthisuthimathavee, S., and Suraninpong, P. (2021). Lysigenous aerenchyma formation: responsiveness to waterlogging in oil palm roots. Biol. Plant 65, 167–176. doi: 10.32615/bp.2021.002
Pelagio-Flores, R., Muñoz-Parra, E., Barrera-Ortiz, S., Ortiz-Castro, R., Saenz-Mata, J., Ortega-Amaro, M. A., et al. (2019). The cysteine-rich receptor-like protein kinase CRK28 modulates Arabidopsis growth and development and influences abscisic acid responses. Planta 251, 2. doi: 10.1007/s00425-019-03296-y
Pereira-Santana, A., Alvarado-Robledo, E. J., Zamora-Briseño, J. A., Ayala-Sumuano, J. T., Gonzalez-Mendoza, V. M., Espadas-Gil, F., et al. (2017). Transcriptional profiling of sugarcane leaves and roots under progressive osmotic stress reveals a regulated coordination of gene expression in a spatiotemporal manner. PloS One 12, 1–25. doi: 10.1371/journal.pone.0189271
Pertea, M., Pertea, G. M., Antonescu, C. M., Chang, T.-C., Mendell, J. T., and Salzberg, S. L. (2015). StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 33, 290–295. doi: 10.1038/nbt.3122
Pieterse, C. M. J., Zamioudis, C., Berendsen, R. L., Weller, D. M., Van Wees, S. C. M., and Bakker, P. A. H. M. (2014). Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 52, 347–375. doi: 10.1146/annurev-phyto-082712-102340
Pinedo, I., Ledger, T., Greve, M., and Poupin, M. J. (2015). Burkholderia phytofirmans PsJN induces long-term metabolic and transcriptional changes involved in Arabidopsis thaliana salt tolerance. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.00466
Poupin, M. J., Greve, M., Carmona, V., and Pinedo, I. (2016). A complex molecular interplay of auxin and ethylene signaling pathways is involved in arabidopsis growth promotion by burkholderia phytofirmans psJN. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.00492
Poupin, M. J., Timmermann, T., Vega, A., Zuñiga, A., and González, B. (2013). Effects of the Plant Growth-Promoting Bacterium Burkholderia phytofirmans PsJN throughout the Life Cycle of Arabidopsis thaliana. PloS One 8, null. doi: 10.1371/journal.pone.0069435
Rekha, K., Kumar, R. M., Ilango, K., Rex, A., and Usha, B. (2018). Transcriptome profiling of rice roots in early response to Bacillus subtilis (RR4) colonization. Botany 96, 749–765. doi: 10.1139/cjb-2018-0052
Schneider, C. A., Rasband, W. S., and Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Sharma, M., Fuertes, D., Perez-Gil, J., and Lois, L. M. (2021). SUMOylation in phytopathogen interactions: balancing invasion and resistance. Front. Cell Dev. Biol. 9. doi: 10.3389/fcell.2021.703795
Sharma, D., Tiwari, M., Pandey, A., Bhatia, C., Sharma, A., and Trivedi, P. K. (2016). MicroRNA858 is a potential regulator of phenylpropanoid pathway and plant development. Plant Physiol. 171, 944–959. doi: 10.1104/pp.15.01831
Shekhawat, K., Fröhlich, K., García-Ramírez, G. X., Trapp, M. A., and Hirt, H. (2023). Ethylene: A master regulator of plant–microbe interactions under abiotic stresses. Cells 12, 31. doi: 10.3390/cells12010031
Shekhawat, K., Saad, M. M., Sheikh, A., Mariappan, K., Al-Mahmoudi, H., Abdulhakim, F., et al. (2021). Root endophyte induced plant thermotolerance by constitutive chromatin modification at heat stress memory gene loci. EMBO Rep. 22, e51049. doi: 10.15252/embr.202051049
Soares, F. S., Rangel de Souza, A. L. S., de Souza, S. A., de Souza Vespoli, L., Pinto, V. B., Matiello, L., et al. (2024). Fine-Tuning of Arabidopsis thaliana Response to Endophytic Colonization by Gluconacetobacter diazotrophicus PAL5 Revealed by Transcriptomic Analysis. Plants 13, 1719. doi: 10.3390/plants13131719
Song, Y., Wilson, A. J., Zhang, X.-C., Thoms, D., Sohrabi, R., Song, S., et al. (2021). FERONIA restricts Pseudomonas in the rhizosphere microbiome via regulation of reactive oxygen species. Nat. Plants 7, 644–654. doi: 10.1038/s41477-021-00914-0
Sun, Y., Cheng, Z., and Glick, B. R. (2009). The presence of a 1-aminocyclopropane-1-carboxylate (ACC) deaminase deletion mutation alters the physiology of the endophytic plant growth-promoting bacterium Burkholderia phytofirmans PsJN. FEMS Microbiol. Lett. 296, 131–136. doi: 10.1111/j.1574-6968.2009.01625.x
Thimm, O., Bläsing, O., Gibon, Y., Nagel, A., Meyer, S., Krüger, P., et al. (2004). mapman: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 37, 914–939. doi: 10.1111/j.1365-313X.2004.02016.x
Thomas, J., Kim, H. R., Rahmatallah, Y., Wiggins, G., Yang, Q., Singh, R., et al. (2019). RNA-seq reveals differentially expressed genes in rice (Oryza sativa) roots during interactions with plant-growth promoting bacteria, Azospirillum brasilense. PloS One 14, 1–19. doi: 10.1371/journal.pone.0217309
Timmermann, T., Armijo, G., Donoso, R., Seguel, A., Holuigue, L., and González, B. (2017). Paraburkholderia phytofirmans PsJN Protects Arabidopsis thaliana Against a Virulent Strain of Pseudomonas syringae Through the Activation of Induced Resistance. Mol. Plant-Microbe Interact. 30, 215–230. doi: 10.1094/MPMI-09-16-0192-R
Timmermann, T., Poupin, M. J., Vega, A., Urrutia, C., Ruz, G. A., and González, B. (2019). Gene networks underlying the early regulation of Paraburkholderia phytofirmans PsJN induced systemic resistance in Arabidopsis. PloS One 14, 1–24. doi: 10.1371/journal.pone.0221358
van de Mortel, J. E., de Vos, R. C. H., Dekkers, E., Pineda, A., Guillod, L., Bouwmeester, K., et al. (2012). Metabolic and transcriptomic changes induced in arabidopsis by the rhizobacterium pseudomonas fluorescens SS101. Plant Physiol. 160, 2173–2188. doi: 10.1104/pp.112.207324
van Loon, L. C. (2007). Plant responses to plant growth-promoting rhizobacteria. Eur. J. Plant Pathol. 119, 243–254. doi: 10.1007/s10658-007-9165-1
Voisin, D., Nawrath, C., Kurdyukov, S., Franke, R. B., Reina-Pinto, J. J., Efremova, N., et al. (2009). Dissection of the complex phenotype in cuticular mutants of arabidopsis reveals a role of SERRATE as a mediator. PloS Genet. 5, 1–19. doi: 10.1371/journal.pgen.1000703
Waheed, S., Anwar, M., Saleem, M. A., Wu, J., Tayyab, M., and Hu, Z. (2021). The critical role of small RNAs in regulating plant innate immunity. Biomolecules 11, 184. doi: 10.3390/biom11020184
Wei, J., Zhou, Q., Zhang, J., Wu, M., Li, G., and Yang, L. (2024). Dual RNA-seq reveals distinct families of co-regulated and structurally conserved effectors in Botrytis cinerea infection of Arabidopsis thaliana. BMC Biol. 22, 239. doi: 10.1186/s12915-024-02043-4
Witzel, K., Üstün, S., Schreiner, M., Grosch, R., Börnke, F., and Ruppel, S. (2017). A proteomic approach suggests unbalanced proteasome functioning induced by the growth-promoting bacterium kosakonia radicincitans in arabidopsis. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00661
Wu, H.-J., Ma, Y.-K., Chen, T., Wang, M., and Wang, X.-J. (2012). PsRobot: a web-based plant small RNA meta-analysis toolbox. Nucleic Acids Res. 40, W22–W28. doi: 10.1093/nar/gks554
Xing, H., Fu, X., Yang, C., Tang, X., Guo, L., Li, C., et al. (2018). Genome-wide investigation of pentatricopeptide repeat gene family in poplar and their expression analysis in response to biotic and abiotic stresses. Sci. Rep. 8, 2817. doi: 10.1038/s41598-018-21269-1
Yadeta, K. A., Elmore, J. M., Creer, A. Y., Feng, B., Franco, J. Y., Rufian, J. S., et al. (2017). A cysteine-rich protein kinase associates with a membrane immune complex and the cysteine residues are required for cell death. Plant Physiol. 173, 771–787. doi: 10.1104/pp.16.01404
Young, M. D., Wakefield, M. J., Smyth, G. K., and Oshlack, A. (2010). Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 11, R14. doi: 10.1186/gb-2010-11-2-r14
Zeiner, A., Colina, F. J., Citterico, M., and Wrzaczek, M. (2023). CYSTEINE-RICH RECEPTOR-LIKE PROTEIN KINASES: their evolution, structure, and roles in stress response and development. J. Exp. Bot. 74, 4910–4927. doi: 10.1093/jxb/erad236
Zhang, M. and Kong, X. (2022). How plants discern friends from foes. Trends Plant Sci. 27, 107–109. doi: 10.1016/j.tplants.2021.11.001
Zhang, Y., Tian, H., Chen, D., Zhang, H., Sun, M., Chen, S., et al. (2023). Cysteine-rich receptor-like protein kinases: emerging regulators of plant stress responses. Trends Plant Sci. 28, 776–794. doi: 10.1016/j.tplants.2023.03.028
Zhou, L., Chen, J., Li, Z., Li, X., Hu, X., Huang, Y., et al. (2011). Integrated profiling of microRNAs and mRNAs: microRNAs located on xq27.3 associate with clear cell renal cell carcinoma. PloS One 5, 1–10. doi: 10.1371/journal.pone.0015224
Keywords: Arabidopsis thaliana, beneficial bacteria, Botrytis cinerea, roots, systemic response, transcriptome analysis, miRNA, priming
Citation: Nešić S, Bosnić D, Samardžić J, Nikolić I, Aziz A and Nikolić D (2025) Paraburkholderia phytofirmans PsJN triggers local and systemic transcriptional reprogramming in Arabidopsis thaliana and increases resistance against Botrytis cinerea. Front. Plant Sci. 16:1554036. doi: 10.3389/fpls.2025.1554036
Received: 31 December 2024; Accepted: 27 April 2025;
Published: 03 June 2025.
Edited by:
Raj Majumdar, Agricultural Research Service (USDA), United StatesReviewed by:
Srayan Ghosh, Durham University, United KingdomManivannan Paramasivan, Bharathidasan University, India
Copyright © 2025 Nešić, Bosnić, Samardžić, Nikolić, Aziz and Nikolić. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sofija Nešić, c29maWphLm5lc2ljQGltZ2dlLmJnLmFjLnJz