 Fenghou Shi
Fenghou Shi Haoyi Feng
Haoyi Feng Xiaojing Li1,2†
Xiaojing Li1,2†- 1Collaborative Innovation Centre of Sustainable Forestry in Southern China, College of Forestry and Grassland, Nanjing Forestry University, Nanjing, China
- 2Yudu County Forestry Bureau, Ganzhou, China
- 3Shanghai Forestry General Station, Shanghai, China
- 4Shandong Provincial Center of Forest and Grass Germplasm Resources, Jinan, China
Introduction: As a native tree species in China, Machilus thunbergii is highly responsive to nitrogen fertilization. However, related studies are scarce. This research seeks to elucidate how different nitrogen fertilizers affect its growth and nitrogen metabolism across different growth stages, thereby determining the most suitable type and establishing a scientific foundation for its fertilization.
Methods: This study aimed to investigate the effects of topdressing with different nitrogen fertilizers on the growth and nitrogen metabolism of M. thunbergii seedlings, with the goal of providing a scientific basis for optimized nitrogen fertilization management in M. thunbergii cultivation. The study used 3-year-old M. thunbergii seedlings as the material, and the fertilization rate was 3 g per seedling. The experiment was conducted in a one-way randomized block design with four treatments, including the control treatment and three nitrogen fertilizer treatments: urea (amide nitrogen fertilizer), ammonium sulfate (ammonium nitrogen fertilizer), and sodium nitrate (nitrate nitrogen fertilizer). From the start of the experiment, the branch and leaf morphology, the height growth, and the basal diameter growth of the seedlings in each treatment were monitored periodically. The activities of the nitrogen-metabolizing enzymes, such as nitrate reductase, glutamine synthetase, glutamate synthetase, and glutamate dehydrogenase, in the leaves were also measured. At growth cessation, all treatments were evaluated for biomass production, root morphological characteristics, and total nitrogen content in different plant parts (i.e., roots, stems, and leaves).
Results: Due to the high nitrogen content in the cultivation substrate, the application of the different nitrogen fertilizers induced varying levels of fertilizer injury. Temporal analysis revealed that the growth inhibition was not uniform across stages. While all nitrogen treatments ultimately suppressed the overall height and diameter growth compared with the control, the timing and the intensity of these effects varied. For instance, the urea treatment initially showed less inhibition, whereas the sodium nitrate treatment consistently exhibited the strongest inhibitory effect throughout the experiment. Similarly, the promotion of nitrogen metabolism enzyme activity by the different fertilizers also displayed distinct temporal patterns, with peaks occurring at different measurement points. All nitrogen treatments increased the nitrogen content in the root, stem, and leaf parts, but decreased the nitrogen translocation efficiency of M. thunbergii seedlings. All nitrogen treatments increased the nitrogen accumulation in the roots and stems of seedlings. Urea treatment enhanced foliar nitrogen accumulation, whereas both the ammonium sulfate and sodium nitrate treatments reduced foliar nitrogen accumulation.
Conclusion: All three nitrogen treatments significantly influenced both the growth and physiological indices of M. thunbergii seedlings. While generally enhancing the nitrogen metabolism and accumulation, improper selection of fertilizer types or excessive application rates elevated the tissue nitrogen concentration, inducing phytotoxic effects that ultimately inhibited seedling growth. In this research, sodium nitrate had the greatest toxic effect on M. thunbergii seedlings, followed by ammonium sulfate and urea. Among the nitrogen fertilizers tested, urea proved superior at an application rate of 3 g per plant for 3-year-old M. thunbergii seedlings.
1 Introduction
As the predominant mineral nutrient for plants, nitrogen is an important component of various secondary metabolites in plants and is involved in the synthesis of proteins, nucleic acids, and phospholipids, among others. It also has significant impact on plant growth and nitrogen metabolism (Farhan et al., 2024; Gong et al., 2024; Langworthy et al., 2023; Xu et al., 2017). Given the naturally low nitrogen availability in the majority of soils, nitrogen fertilization is essential for seedling cultivation. Optimal nitrogen fertilizer application facilitates higher chlorophyll content (Qiu et al., 2024) and nutrient balance (Dong W.N, et al., 2025), thereby promoting overall plant development (Li Y.L, et al., 2023). However, inappropriate application can lead to a series of problems such as low nitrogen fertilizer utilization efficiency, plant growth inhibition, reduced yield, soil compaction, and environmental pollution (Saurav et al., 2024). Since the 1980s, with the objective of enhancing agricultural and forestry yields, the amount of nitrogen fertilizer applied in China has rapidly increased, causing a series of issues. Aiming to avoid unsuitable fertilization, numerous studies have been conducted on the types and the amounts of nitrogen fertilizers. The main types of nitrogen absorbed by plants from the soil are nitrate nitrogen (NO3−–N) and ammonium nitrogen (NH4+–N). These distinct forms of nitrogen have significant effects not only on the plant growth but also on its physiology (Yin and Zhang, 2025; Ge et al., 2024). Therefore, studying the response of plants to different forms of nitrogen is of great significance in agricultural and forestry applications. Nitrogen assimilation, the conversion of inorganic nitrogen to organic forms, can be evaluated through the activities of key enzymes [e.g., nitrate reductase (NR), glutamine synthetase (GS), glutamate synthetase (GOGAT), and glutamate dehydrogenase (GDH)] (Yu et al., 2024). The amino acids synthesized in this process are further converted into various substances, such as proteins, chlorophyll, vitamins, and nucleic acids, among others, to support plant life activities (Kusano et al., 2011).
Machilus thunbergii is a broad-leaved evergreen tree in the genus Machilus of the family Lauraceae, which is widely distributed in the provinces of Shandong, Zhejiang, and Guangdong in China (Huang et al., 2013). This species possesses significant ornamental, economic, and medicinal values, as well as ecological functions including soil/water conservation, salt tolerance, and wind resistance. M. thunbergii has broad potential for cultivation and utilization. Current studies on M. thunbergii have primarily concentrated on population distribution (Li et al., 2024; Zhang et al., 2023), growth and reproduction (Li et al., 2022; Qian, 2022), and landscape and medicinal value utilization (Ma and Huang, 2019), among others. These studies are of great significance for the protection of M. thunbergii. However, research on the fertilization strategies for M. thunbergii seedlings is limited, with no established standardized fertilization protocols. Fertilization experiments were conducted on Lindera megaphylla and Cinnamomum chekiangense, which belong to the family Lauraceae, similarly to M. thunbergii. The results showed that, among the elements nitrogen, phosphorus, and potassium, nitrogen had the greatest impact on the growth of seedlings (Dong et al., 2011; Song and Fei, 2013). Therefore, it is of practical significance to study the effects of different nitrogen treatments on the growth and nitrogen metabolism of M. thunbergii seedlings. However, the response of plants to nitrogen is dynamic, not static. Therefore, a fertilizer that provides a strong initial growth stimulus may not offer sustained benefits, and the effect on nitrogen metabolism can vary over time. To comprehensively evaluate the effects of different nitrogen forms, it is essential to monitor the key growth and physiological parameters at multiple stages throughout the growth cycle. This time series approach is crucial for the identification of nitrogen sources that provide balanced and persistent advantages.
Employing 3-year-old M. thunbergii container seedlings, this study evaluated the effects of different nitrogen types on the seedling growth, nitrogen transport, and assimilation through sequential measurements over 150 days. The aim was to identify the optimal nitrogen fertilizer at the specified application level, thereby establishing a scientific basis for optimal fertilization and facilitating protection. The response of M. thunbergii seedlings to nitrogen fertilization exhibits a dynamic nature, with the effects varying by nitrogen form, manifesting as heterogeneous patterns such as early-stage rapid promotion or long-term sustained effects. Therefore, this experiment was conducted with a measurement frequency of every 30 days to monitor the seedling height growth, the ground diameter growth, and the nitrogen metabolism enzyme activity. This approach was designed to obtain continuous multi-time point data, enabling a comprehensive analysis of the impacts of the three nitrogen fertilizers on the seedlings over both the short-term and entire experimental periods, thereby providing substantial and reliable data for subsequent research.
2 Materials and methods
2.1 Test site and experimental materials
The fertilization experiment with M. thunbergii seedlings was conducted at the teaching and research base of Nanjing Forestry University in Baima Town (119.181946° E, 31.605082° N, altitude of 10 m). A satellite image is provided in Figure 1. The site features a subtropical monsoon climate with favorable hydrothermal conditions: a mean annual temperature of 15.5°C (summer maximum of 38°C and winter minimum of −8°C), an annual precipitation of 1,053 mm, and sunshine duration 2,240 h. We selected 3-year-old M. thunbergii seedlings for the experiment. The nursery seeds were obtained from the natural population of M. thunbergii in Sheshan Island, Shanghai Municipality. M. thunbergii seedlings with nearly uniform growth were transplanted into pots for acclimatization on March 21, 2021 (pot size: height, 20.5 cm; upper aperture, 24.5 cm; lower aperture, 18 cm), one plant per pot. Each pot was filled with 4 kg seedling substrate (bulk density, 0.31 g cm−3; total porosity, 65%; pH 6.59; EC, 2.71; organic matter content, 37%; and total nitrogen, phosphorus, and potassium content, 2%), which was produced by Jiangsu Xingnong Substrate Science and Technology Co. Ltd. (Zhenjiang, China). In July, the experimental seedlings were transferred into a simple greenhouse for standard cultivation management during the adaptation period. On April 17, 2022, experimental seedlings (80–90 cm tall, with uniform growth) were selected for the formal trial.
Figure 1. Satellite image of the test site.
2.2 Experimental design
The experiment was conducted using a one-way randomized block design with four treatments: urea (amide nitrogen fertilizer, 46.7% N) treatment, ammonium sulfate (ammonium nitrogen fertilizer, 21.2% N) treatment, sodium nitrate (nitrate nitrogen fertilizer, 16.5% N) treatment, and a control (CK) treatment with no nitrogen fertilizer applied. The experiment employed three replicates per treatment, with 12 seedlings per replicate, giving a final sample size of 144 seedlings. The initial fertilization day was designated as 0 DAT (days after transplantation). Nitrogen was applied at a rate of 3 g per seedling. This dosage was determined based on the Jiangxi Provincial Administration for Market Regulatio, (2018), which recommends an appropriate nitrogen application rate of 2–3 kg per 667 m2. Calculated based on the substrate of each pot in this experiment, the application rate was set at 3 g per plant divided equally into three applications (0, 30, and 60 DAT), with each dose dissolved in 200 ml deionized water and applied to the substrate. The entire fertilizer experiment was maintained for 150 days, during which the seedlings were under standard irrigation management throughout. Detailed treatment specifications are presented in Table 1.

Table 1. Formulation and application rates of nitrogen fertilizer treatments.
2.3 Observation of the seedling morphology and determination of the growth indicators
The stem and leaf morphology was observed and recorded at 30-day intervals for all treatments throughout the experiment. The seedling height and the basal diameter were measured at 30-day intervals on 12 randomly selected seedlings per treatment. Height was measured using a steel ruler (0.1-cm precision), and diameter was measured with digital calipers (0.01-mm precision) (Chen et al., 2023). The growth increments for both parameters were calculated for each interval. At the end of the experiment, three seedlings per treatment were randomly sampled. These seedlings were extracted from the soil manually, divided into three parts (i.e., roots, stems, and leaves) according to the method described by Li et al. (2022), dried under 105°C until the weight remains constant, and then weighed individually with an electronic balance (0.001 g).
2.4 Determination of the seedling root metrics
At the end of the experiment, three seedlings per treatment were randomly selected. The roots were carefully excavated from the substrate, gently rinsed with water, and then scanned using an Epson scanner. The total root length, the root surface area, the root volume, the average root basal diameter, and the total root tip number were analyzed using the WinRHIZO PRO 2007 software (Ren et al., 2023).
2.5 Determination of the leaf nitrogen-metabolizing enzyme activity
A total of 12 seedlings were randomly selected for each treatment. Throughout the experiment, one or two intact mature leaves from the middle and the upper two to three layers of the seedlings were taken at 30-day intervals, frozen and stored in an ice box for transport back to the laboratory, and then cut and mixed. Subsequently, the activities of NR, GS, GOGAT, and GDH were measured for each treatment using test kits. The kits for the measurement of the enzyme activities were obtained from Suzhou Keming Biotechnology Co. (Suzhou, China).
2.6 Determination of the total nitrogen content and accumulation in each part and the transfer factor of the seedling
To investigate the nitrogen uptake and translocation in plants, the nitrogen content and its accumulation were determined at the end of the experiment, and the transfer factor (TF) was studied. To determine the nitrogen content in each part of the seedlings with the different treatments, the seedlings were divided into three parts: roots, stems, and leaves. Pooled samples from the same treatment and plant part were dried, ground, and digested with sulfuric acid–hydrogen peroxide. The total nitrogen content was determined using the micro-Kjeldahl method, adhering to the established standard for plant NPK determination (Ministry of Agriculture and Rural Affairs of the People’s Republic of China, 2011).
where NA is the nitrogen accumulation (in milligrams per seedling), Nc is the nitrogen content in a specific plant part, and DM is the dry matter of that part.
where TF is the nitrogen transfer factor, Nstem is the nitrogen content of the stems, Nleaf is the nitrogen content of the leaves, and Nroot is the nitrogen content of the roots (Mostafa et al., 2021).
2.7 Data processing and analysis
All statistical analyses were performed using SPSS 23.0. Significant differences were determined using one-way analysis of variance (ANOVA). Statistical significance was defined as p < 0.01 and p < 0.05. Graphs were generated using Origin 2024 software.
3 Results
3.1 Assessment of M. thunbergii seedling branch and leaf morphology
M. thunbergii seedlings received distinct nitrogen treatments, with the foliar morphology monitored throughout the experimental period. The morphological changes and photos of the seedlings across the experiment are shown in Table 2 and Figure 2, respectively.
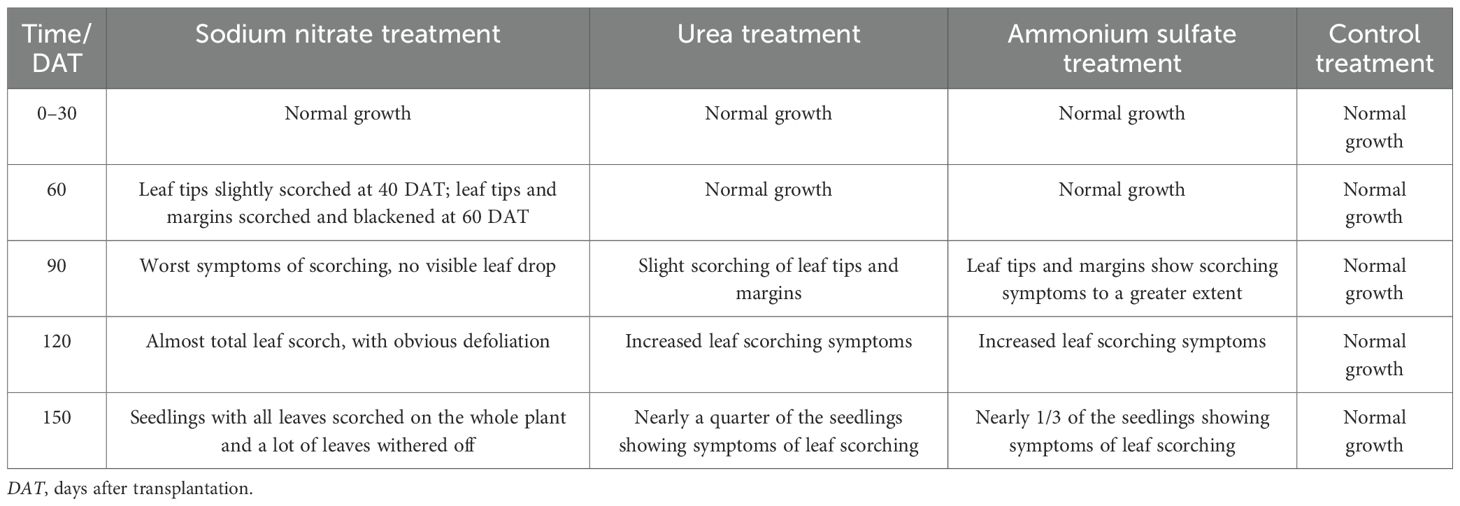
Table 2. Temporal changes in the foliar and branch morphology of Machilus thunbergii seedlings under different nitrogen treatments.

Figure 2. Key stages in the fertilization experiment of Machilus thunbergii seedlings. (A) Transplantation of seedlings into experimental pots. (B) Acclimatization period prior to fertilization. (C) Application of the nitrogen fertilizer treatments.
Table 2 demonstrates that all nitrogen treatments induced varying degrees of fertilizer injury in M. thunbergii seedlings, with the symptom severity progressively intensifying over time. This phytotoxicity occurred because the initially elevated nitrogen concentration in the substrate was exacerbated by supplemental fertilization. Sodium nitrate-treated seedlings elicited the earliest and most severe symptoms, followed by the ammonium sulfate-treated seedlings. The urea-treated seedlings exhibited the mildest phytotoxic effects. No seedling mortality occurred by 150 DAT. Subsequent observation of the seedlings showed that the majority of them gradually sprouted new shoots in the basal stem portion.
3.2 Measurement results of M. thunbergii seedling growth indicators
3.2.1 Determination of the height growth and the basal diameter growth of M. thunbergii seedlings in the different treatments
The height and diameter increments of M. thunbergii seedlings are presented in Table 3 and Figure 3. Throughout the experimental period, the performance of the different nitrogen fertilizer treatments exhibited significant stage specificity. In terms of height growth (Figure 3A), the CK group maintained a leading position across multiple stages (0–30 DAT and 90–150 DAT) and was significantly greater than all other fertilizer treatments during 90–150 DAT (0.01 < p < 0.05). However, the fertilizer treatments also demonstrated advantages during specific periods: the ammonium sulfate treatment achieved the maximum height increment at 30–60 DAT, while the urea treatment at 60–90 DAT was significantly higher than the other two nitrogen treatments (0.01 < p < 0.05), albeit not significantly different from the control (p > 0.05).
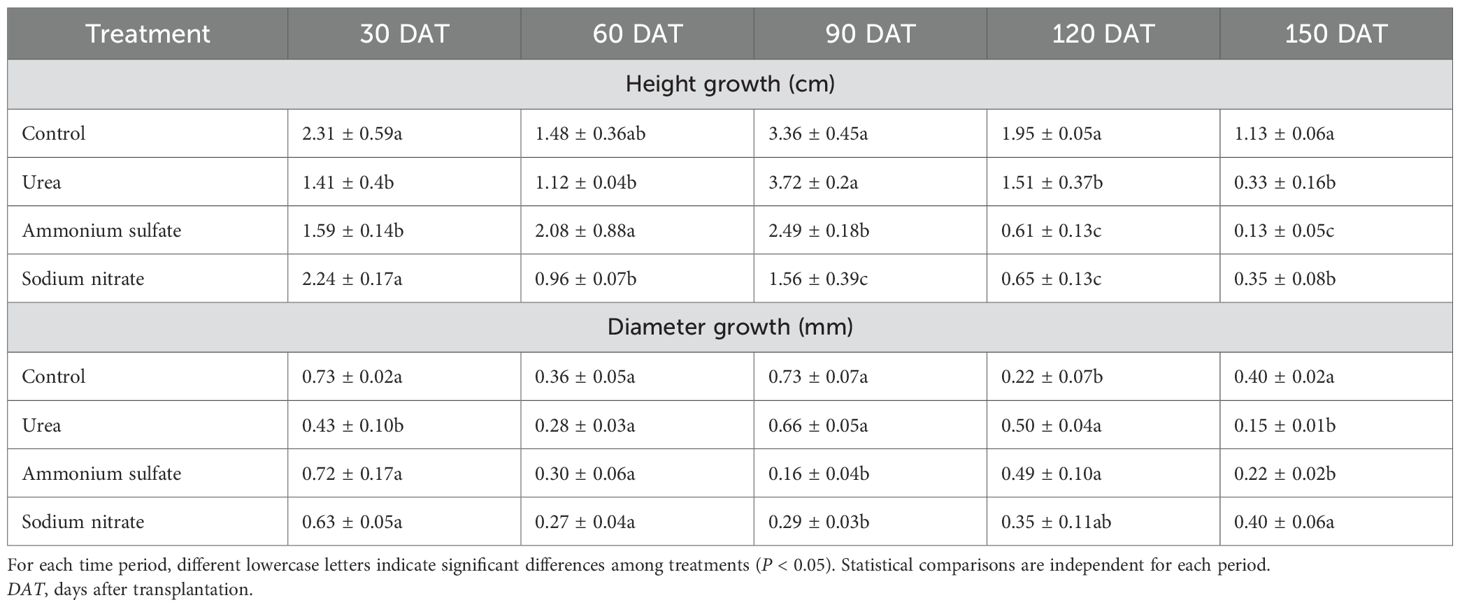
Table 3. Temporal patterns of the height and diameter growth of Machilus thunbergii seedlings across nitrogen treatments.
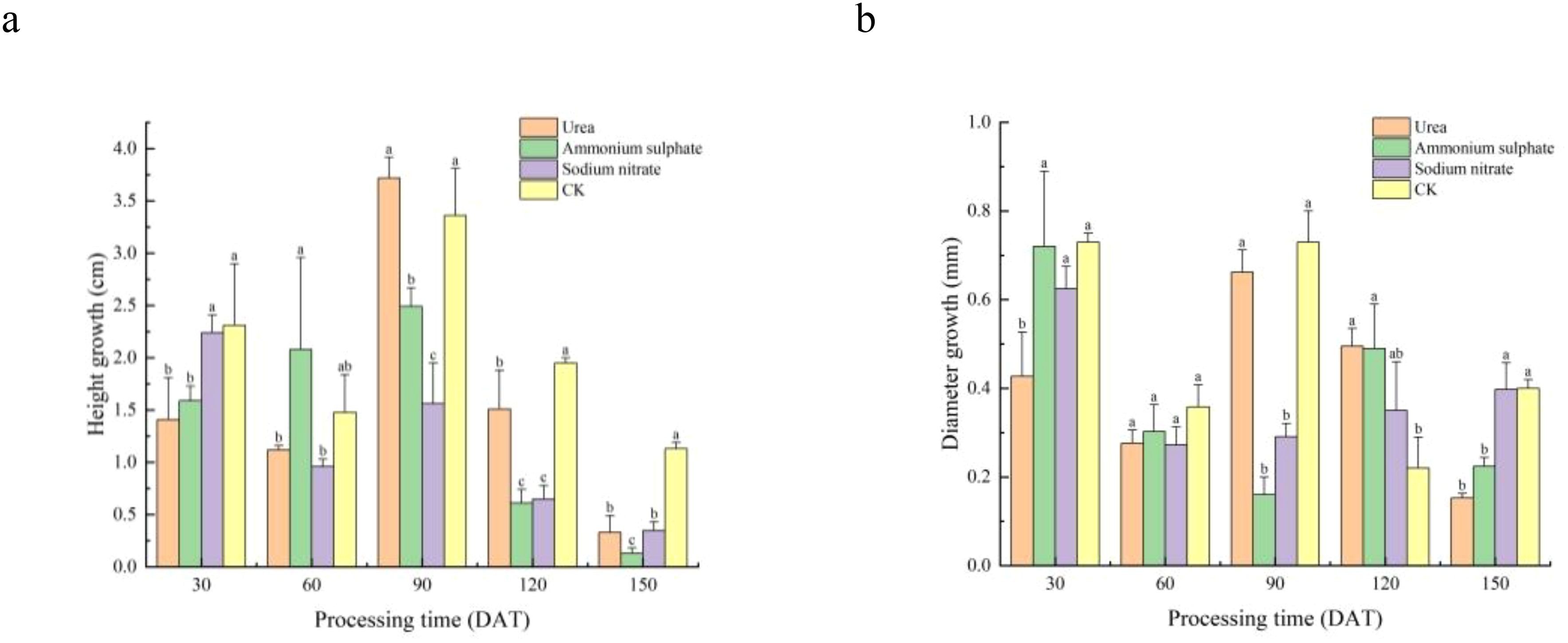
Figure 3. Temporal dynamics of Machilus thunbergii seedling growth under different nitrogen regimes. Increments in height growth (A) and diameter growth (B) across five measurement periods. For each time period, different lowercase letters above bars indicate significant differences among treatments (p < 0.05). Statistical comparisons are independent for each period.
In terms of diameter growth, the control group yielded the greatest increment during 0–90 DAT. In the later growth stages, the effects of the nitrogen fertilizers became apparent: at 90–120 DAT, the diameter increment in the urea treatment was significantly greater than that in the control (0.01 < p < 0.05). At 120–150 DAT, the diameter increment in the sodium nitrate treatment peaked and was significantly higher than that in both the urea and ammonium sulfate treatments (0.01 < p < 0.05).
Under the fertilization level set in this experiment, the promoting effects of the nitrogen fertilizers on the growth of M. thunbergii seedlings were stage-specific and did not demonstrate a significant and sustained overall advantage over the control group.
3.2.2 Determination of the biomass of M. thunbergii seedlings in the different treatments
The seedling biomass reflects the vegetative growth of seedlings and serves as a key quality indicator (Wei et al., 2018). The biomass measurements across treatments are shown in Table 4.
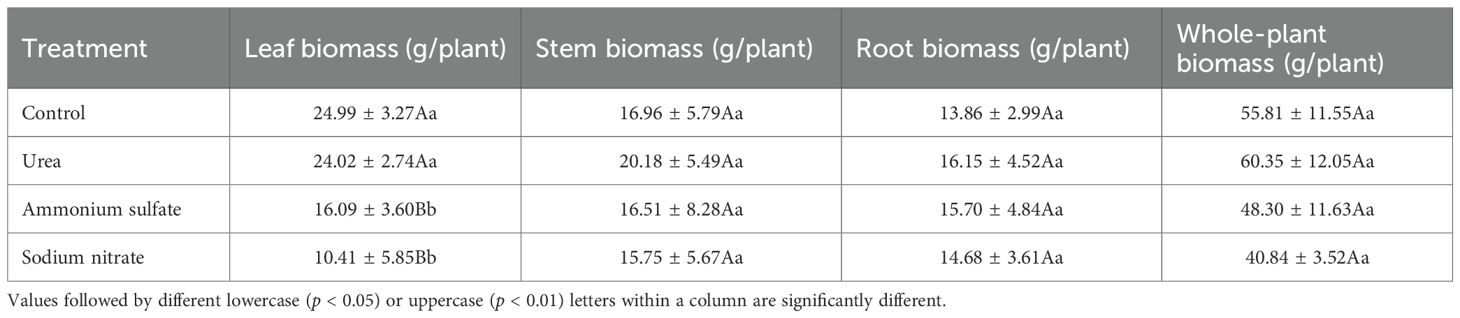
Table 4. Partitioning of the root, stem, and leaf biomass in response to nitrogen fertilizer treatments at the end of the experiment.
As shown in Table 4, the urea treatment showed maximum values for the stem, root, and total biomass, increasing by 18.99%, 20.85%, and 8.13%, respectively, compared with the control. The seedlings in the sodium nitrate treatment had the lowest values across all biomass parameters. The stem and root biomass of the control seedlings accounted for 30.89% and 24.83% of the whole plant biomass, respectively, which were lower than those of the other three nitrogen treatments. All three nitrogen treatments enhanced the stem and root biomass (which was most pronounced with urea); however, they reduced the leaf biomass.
3.3 Determination of the root indexes of M. thunbergii seedlings in the different treatments
The results of the seedling root index measurements for each treatment are shown in Table 5.
Table 5. Root system morphological traits of Machilus thunbergii seedlings under different nitrogen treatments.
As shown in Table 5, the urea-treated seedlings showed maximal root system values: the total root length (2,690.30 cm) exceeded those in the ammonium sulfate and sodium nitrate treatments by 17.57% and 28.27%, respectively, representing 1.18-fold of the control. The root surface area, the root volume, and the total number of root tips of the seedlings in this nitrogen treatment were significantly higher than those in the other nitrogen treatments (0.01 < p < 0.05), which were 1,293.74 cm2, 132.60 cm3, and 1,228, respectively, and showing 43.86%, 105.65%, and 66.40% increases over the control, respectively. The mean root basal diameter of the urea-treated seedlings was 1.99 mm, showing a 40.14% increase over the ammonium sulfate-treated seedlings, with the difference reaching a significant level (p < 0.05). The above analyses showed that urea could effectively promote the root growth of M. thunbergii seedlings at this level of nitrogen application, while the ammonium sulfate and sodium nitrate treatments showed negligible effects.
3.4 Determination of the nitrogen metabolism enzyme activity in the leaves of M. thunbergii seedlings with the different treatments
3.4.1 Determination of the leaf NR activity of M. thunbergii seedlings with the different treatments
The results of the leaf NR activity in the seedlings of each treatment are shown in Table 6. Figure 4A reveals that, from 30 to 90 DAT post-treatment, the leaf NR activity in each nitrogen fertilizer treatment was significantly higher than that of the control seedlings (p < 0.05), indicating that the nitrogen fertilizer had a significant promoting effect on the NR activity at this application level during this period. At 120 DAT, the seedlings treated with urea and ammonium sulfate had significantly higher leaf NR activity than those in the sodium nitrate and control treatments (0.01 < p < 0.05). By 150 DAT, the urea treatment maintained significantly superior NR activity compared with all the other groups (0.01 < p < 0.05). Throughout the experiment, all treatments exhibited bimodal fluctuations in the leaf NR activity over time. The first peak of NR activity in the seedlings in the sodium nitrate treatment appeared at 30 DAT, while those in the urea and ammonium sulfate treatments appeared at 60 DAT. The second peak of NR activity in each nitrogen application treatment appeared at 120 DAT. These results indicate that the leaf NR activity of M. thunbergii seedlings is co-regulated by the nitrogen source and the treatment duration. The promoting effect was phase-specific, with urea demonstrating a more pronounced and sustained induction of NR activity, particularly at the later stage of the experiment.
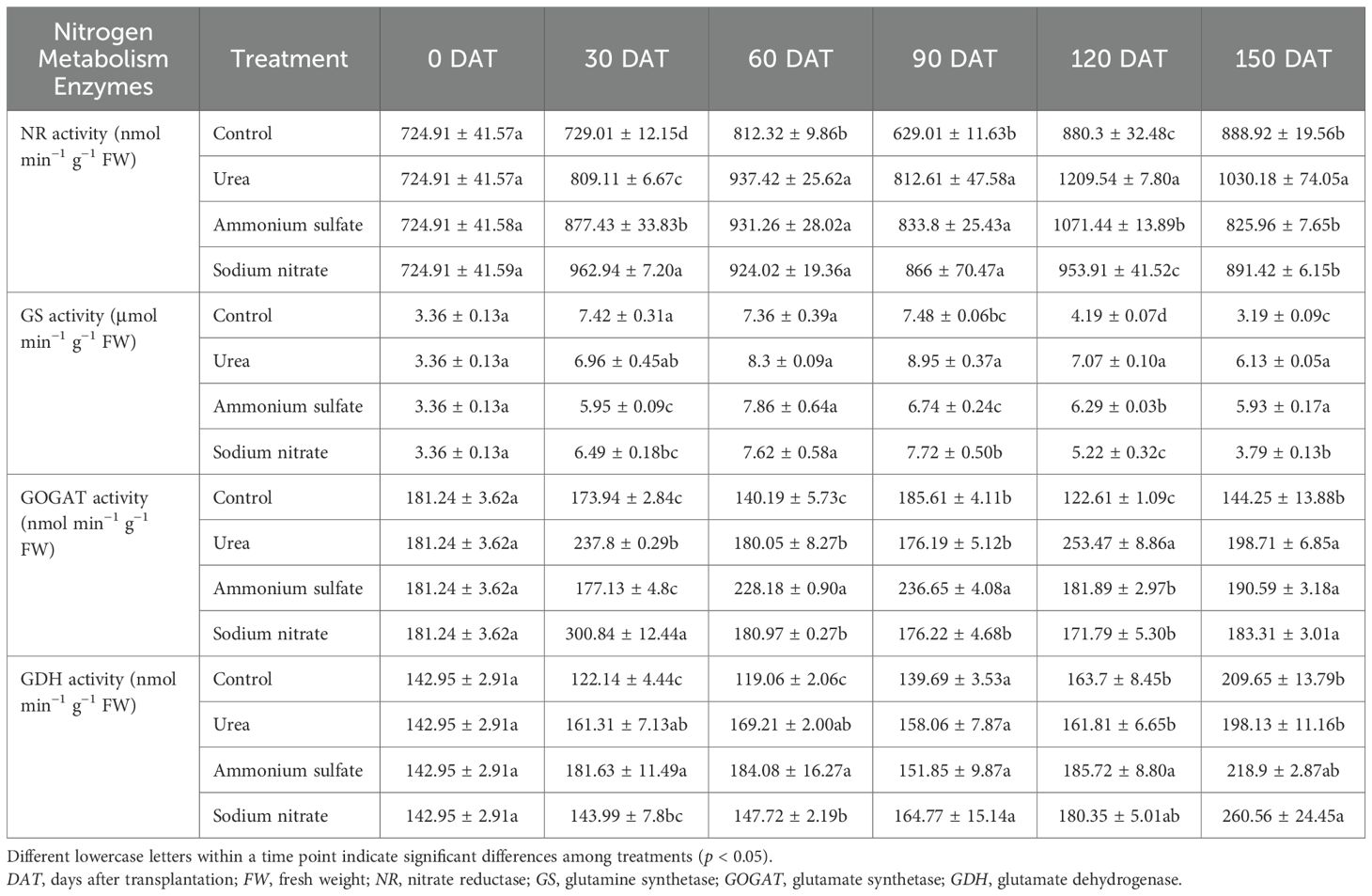
Table 6. Temporal dynamics of the activities of the key nitrogen metabolism enzymes in the leaves of Machilus thunbergii seedlings across nitrogen treatments.
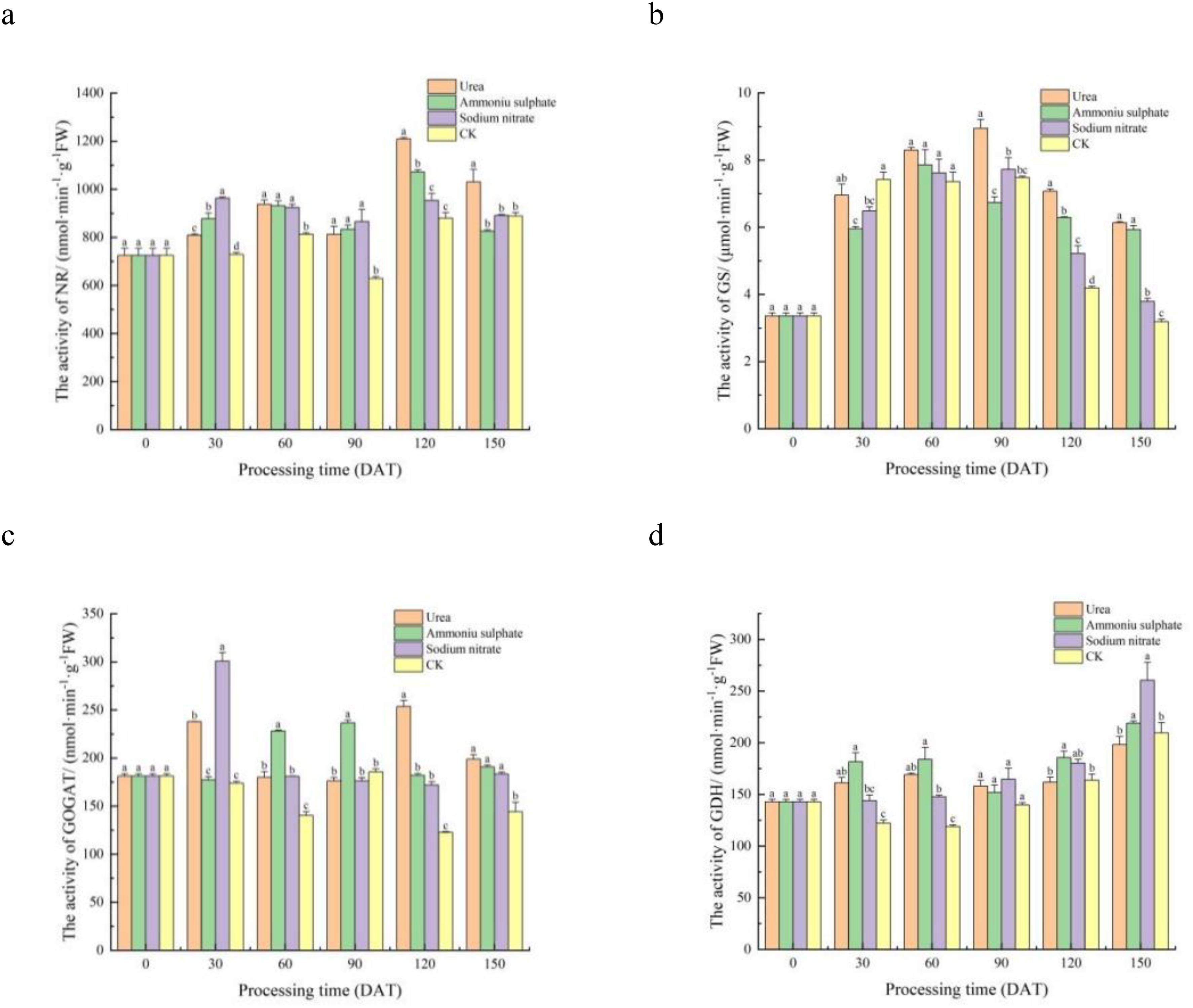
Figure 4. Time course changes in the leaf nitrogen metabolism enzyme activities of Machilus thunbergii seedlings under different nitrogen fertilizers. Nitrate reductase (NR) (A), glutamine synthetase (GS) (B), glutamate synthase (GOGAT) (C), and glutamate dehydrogenase (GDH) (D) were measured at five time points. Different lowercase letters within a time point indicate significant differences among treatments (p < 0.05).
3.4.2 Determination of the leaf GS activity of M. thunbergii seedlings with the different treatments
The results of the leaf GS activity in the seedlings of each treatment are shown in Table 6. Figure 4B shows that, at 30 DAT, the leaf GS activity of the seedlings in each nitrogen fertilizer treatment was observably lower than that of the control, although the difference was not statistically significant (p > 0.05). From 30 to 120 DAT, all treatments displayed a unimodal pattern in leaf GS activity, initially increasing and then decreasing. The leaf GS activity of the seedlings treated with ammonium sulfate reached its peak at 60 DAT, while those of the seedlings treated with urea and sodium nitrate reached their peaks at 90 DAT. At 150 DAT, the leaf GS activity of the seedlings in the three nitrogen treatments was significantly higher than that of the control (p < 0.05). These results demonstrate that the leaf GS activity in M. thunbergii seedlings was co-regulated by the nitrogen fertilizer type and the application duration. Specifically, while nitrogen fertilization initially suppressed the GS activity relative to the control, it ultimately induced a significant and sustained enhancement by the end of the experiment, with different nitrogen sources triggering distinct temporal patterns in peak activity.
3.4.3 Determination of the leaf GOGAT activity of M. thunbergii seedlings with the different treatments
The results of the leaf GOGAT activity in the seedlings of each treatment are shown in Table 6. Figure 4C illustrates distinct temporal dynamics in the GOGAT activity among the nitrogen treatments. Specifically, the activity in the ammonium sulfate- and sodium nitrate-treated seedlings followed a unimodal pattern, whereas a bimodal pattern was observed in the urea-treated seedlings. At 30 DAT, the leaf GOGAT activity in the seedlings of both the urea and sodium nitrate treatments reached the first peak and was significantly higher than that of the control (0.01 < p < 0.05). No significant difference was observed between the ammonium sulfate treatment and the control (p > 0.05). By 60 DAT, the ammonium sulfate treatment exhibited significantly higher GOGAT activity than all the other treatments (0.01 < p < 0.05), and this superiority persisted at its peak (90 DAT). A second pronounced peak occurred at 120 DAT in the urea-treated seedlings, which was significantly higher than all other nitrogen treatments (0.01 < p < 0.05). Throughout the experiment, the leaf GOGAT activity of the seedlings in the three nitrogen treatments showed higher values than that of the control. This result indicates that the changes in the GOGAT activity in M. thunbergii seedling leaves are co-regulated by the nitrogen fertilizer type and the application duration, exhibiting a complex temporal pattern. Among the fertilizers tested, urea was the most effective in enhancing the activity of GOGAT, inducing two significant peaks over the experimental period.
3.4.4 Determination of the leaf GDH activity of M. thunbergii seedlings with the different treatments
The results of the leaf GDH activity of the seedlings in each treatment are shown in Table 6. Statistical analysis of the temporal pattern (Figure 4D) revealed that the GDH activity was relatively stable during the early stage (0–60 DAT), with no consistent significant differences observed among the treatments. The ranking of GDH activity among treatments varied across time points: at 30 and 60 DAT, the order was ammonium sulfate > urea > sodium nitrate > the control, while by 90 DAT, it shifted to sodium nitrate > urea > ammonium sulfate > the control. At 120 DAT, the ammonium sulfate treatment reached its peak GDH activity, which was significantly higher than that of the control (0.01 < p < 0.05). By the end of the experiment (150 DAT), all nitrogen treatments had reached their peak leaf GDH activity. Seedlings with the sodium nitrate treatment showed the highest leaf GDH activity, differing significantly from those in the control and urea treatments (0.01 < p < 0.05). Compared with that in the control, the leaf GDH activity in the ammonium sulfate, sodium nitrate, and urea treatments was increased by 4.41%, 24.28%, and 5.49%, respectively.
This analysis demonstrates that the leaf GDH activity in M. thunbergii seedlings was influenced by both the nitrogen fertilizer type and the treatment duration, with the effect of fertilizer type being more pronounced in the early stages and the effect of duration becoming dominant later. Overall, all three nitrogen sources enhanced the GDH activity by the end of the experiment, with sodium nitrate showing the greatest relative increase.
3.5 Accumulation and transport of nitrogen by M. thunbergii seedlings under the different nitrogen fertilizer treatments
3.5.1 Determination of the nitrogen content and accumulation in different parts of M. thunbergii seedlings
Table 7 and Figure 5 present the results of the nitrogen content and accumulation (calculated with Equation 1) in the roots, stems, and leaves of M. thunbergii seedlings across treatments. As shown in Figure 5A, there were significant differences (p < 0.05) between the nitrogen contents in the leaf, stem, and root parts of M. thunbergii seedlings for the different nitrogen fertilizer treatments, and the total nitrogen content in the seedlings of each nitrogen treatment followed the order: sodium nitrate treatment > ammonium sulfate treatment > urea treatment > the control. Compared with the seedlings in the control group, those treated with urea, ammonium sulfate, and sodium nitrate increased their nitrogen content by 14.66%–56.86% in the leaves, 27.44%–99.04% in the stems, and 45.25%–87.43% in the roots, respectively. For nitrogen accumulation in each part of the seedlings, the leaf part of the urea-treated seedlings showed maximum accumulation, while that of the sodium nitrate-treated seedlings had the minimum. No significant difference (p > 0.05) was observed in the stem nitrogen accumulation among treatments (p > 0.05). The nitrogen accumulation in the root part of the seedlings in each nitrogen treatment was higher than that of the control, with the nitrogen accumulation in the root part of the seedlings treated with ammonium sulfate exhibiting the maximum value. In both the urea and control treatments, nitrogen accumulation exhibited the following pattern: leaf > stem > root. However, the ammonium sulfate and sodium nitrate treatments showed peak nitrogen accumulation in the roots and stems, respectively, which was due to the phytotoxicity from the two treatments: a large number of leaves fell off. These results indicate that the application of nitrogen fertilizers increases the nitrogen content in all parts of M. thunbergii seedlings, but that ammonium sulfate and sodium nitrate treatments adversely affect the leaf nitrogen accumulation.
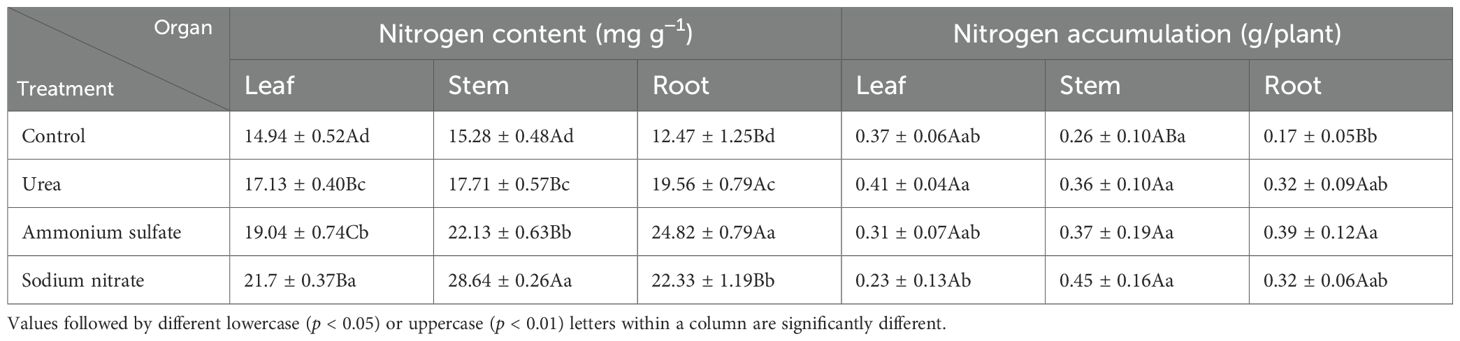
Table 7. Nitrogen content and accumulation in the different organs of Machilus thunbergii seedlings at the final harvest.
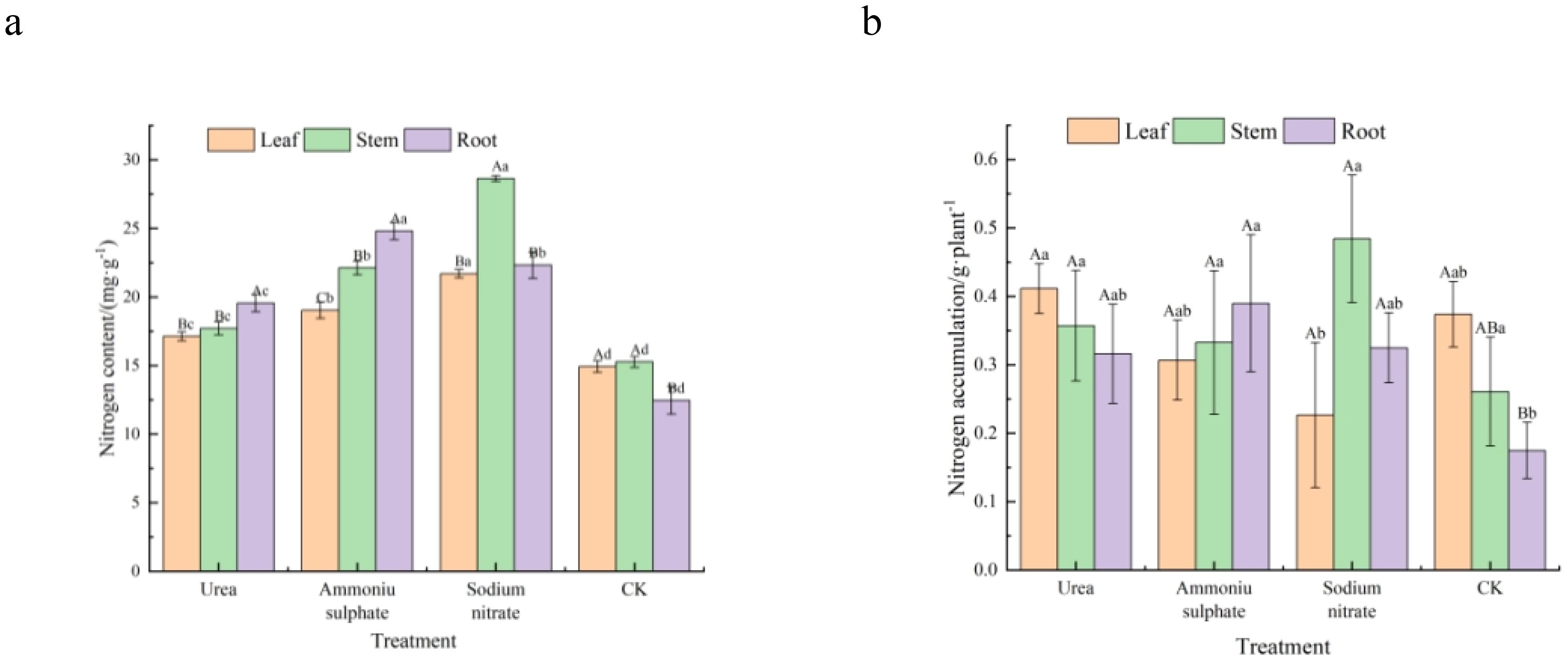
Figure 5. Organ-specific nitrogen allocation in Machilus thunbergii seedlings as affected by nitrogen fertilization at the final harvest. Nitrogen content (A) and nitrogen accumulation (B) in the roots, stems, and leaves. Different lowercase letters above bars for a given plant part indicate significant differences among treatments (p < 0.05).
3.5.2 Determination of the nitrogen translocation capacity of M. thunbergii seedlings
The nitrogen transport capacity of the seedlings in each nitrogen treatment was calculated with Equation 2, and the results are shown in Figure 6 and Table 8. All nitrogen treatments exhibited lower nitrogen transfer factor compared with the control. The sodium nitrate treatment showed no significant difference from the control (p > 0.05), while the urea and ammonium sulfate treatments showed significantly lower values than the control (p < 0.01). The ammonium sulfate treatment yielded the minimum nitrogen transfer factor of 1.66. These results indicate that the nitrogen treatments do not enhance the nitrogen transport from the roots to the shoots; however, they improve the root nitrogen uptake, with ammonium sulfate showing the strongest promoting effect.
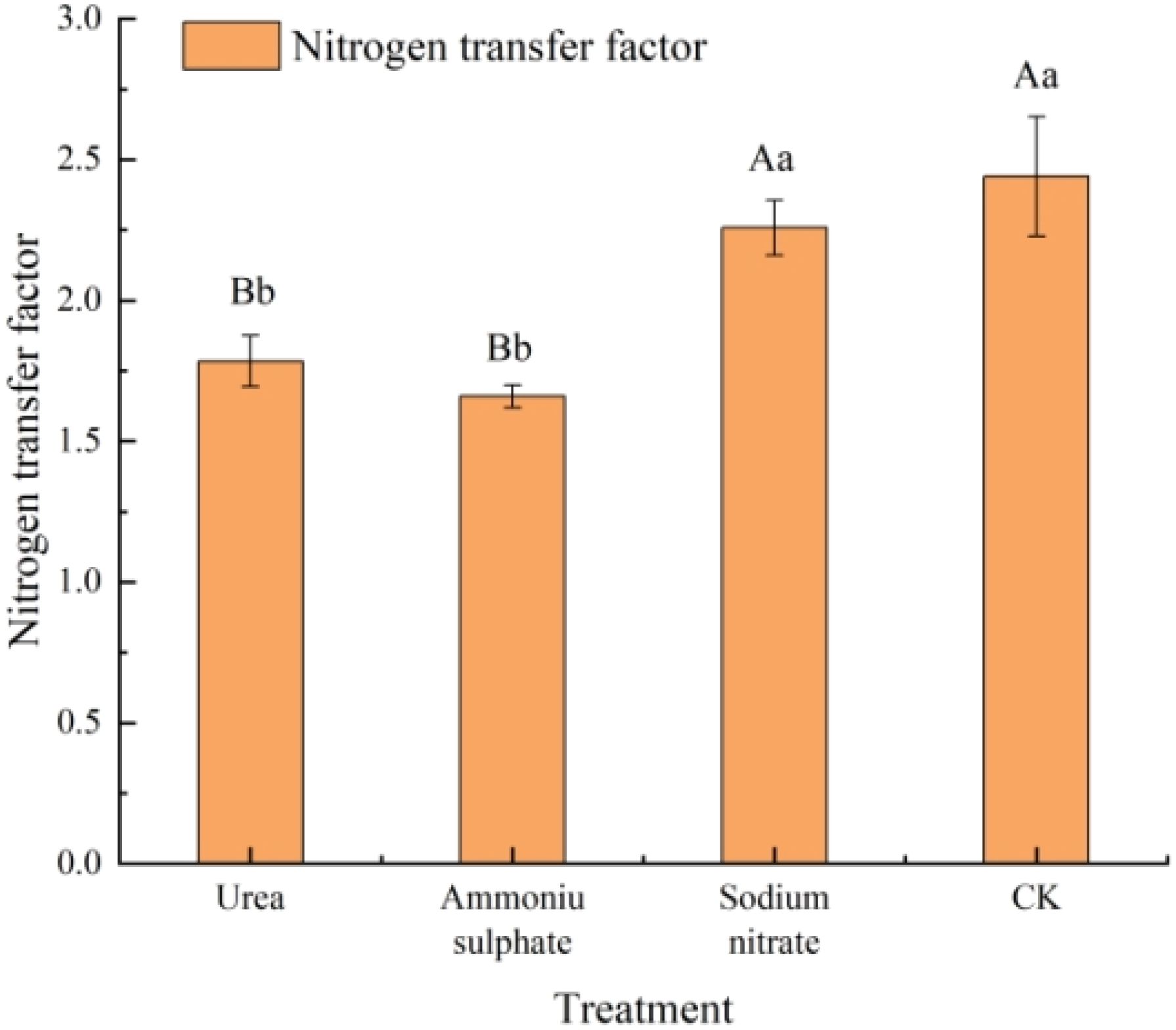
Figure 6. Nitrogen transfer factor of Machilus thunbergii seedlings in response to nitrogen fertilizer treatments. The transfer factor was calculated as the aboveground-to-root nitrogen content ratio. Different lowercase letters indicate significant differences among treatments (p < 0.05).
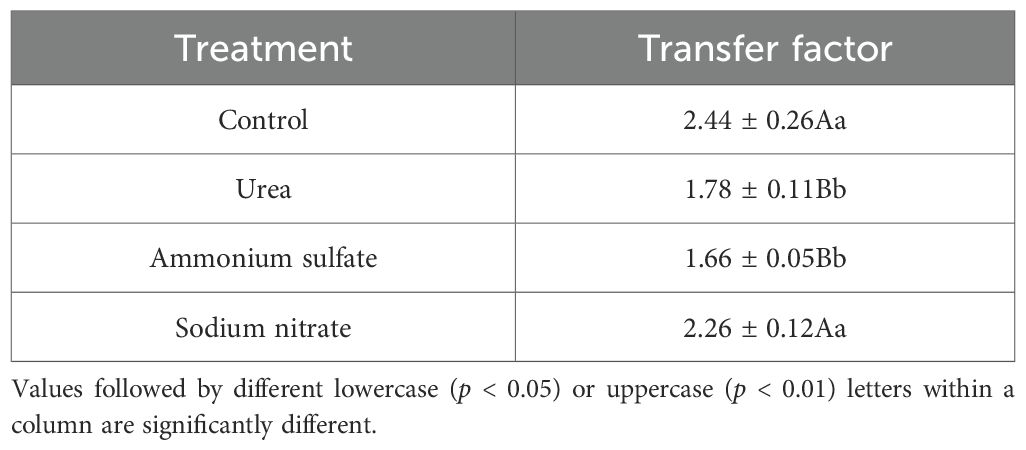
Table 8. Nitrogen transfer factor of Machilus thunbergii seedlings in response to nitrogen fertilizer treatments.
3.6 Correlation analysis of the indexes of M. thunbergii seedlings in the different treatments
To elucidate the macro-level associative patterns among traits, a correlation analysis was performed based on the treatment means of the various parameters for M. thunbergii seedlings under the different fertilization treatments. As shown in Figure 7, positive correlations were predominantly observed among several key clusters: between the biomass and root system parameters, in mutual correlations between the root parameters, between the biomass and enzyme activities, and between the nitrogen content and nitrogen accumulation. These interrelated associations collectively underpin the phenotypic expression of robust shoot and root growth, reflecting an integrated physiological foundation of a highly efficient nitrogen metabolism and a strategic resource allocation (Shipley and Meziane, 2002; Fan et al., 2025). The synergistic coordination between organ growth and nitrogen metabolism function is comprehensively embodied in these positive correlations.
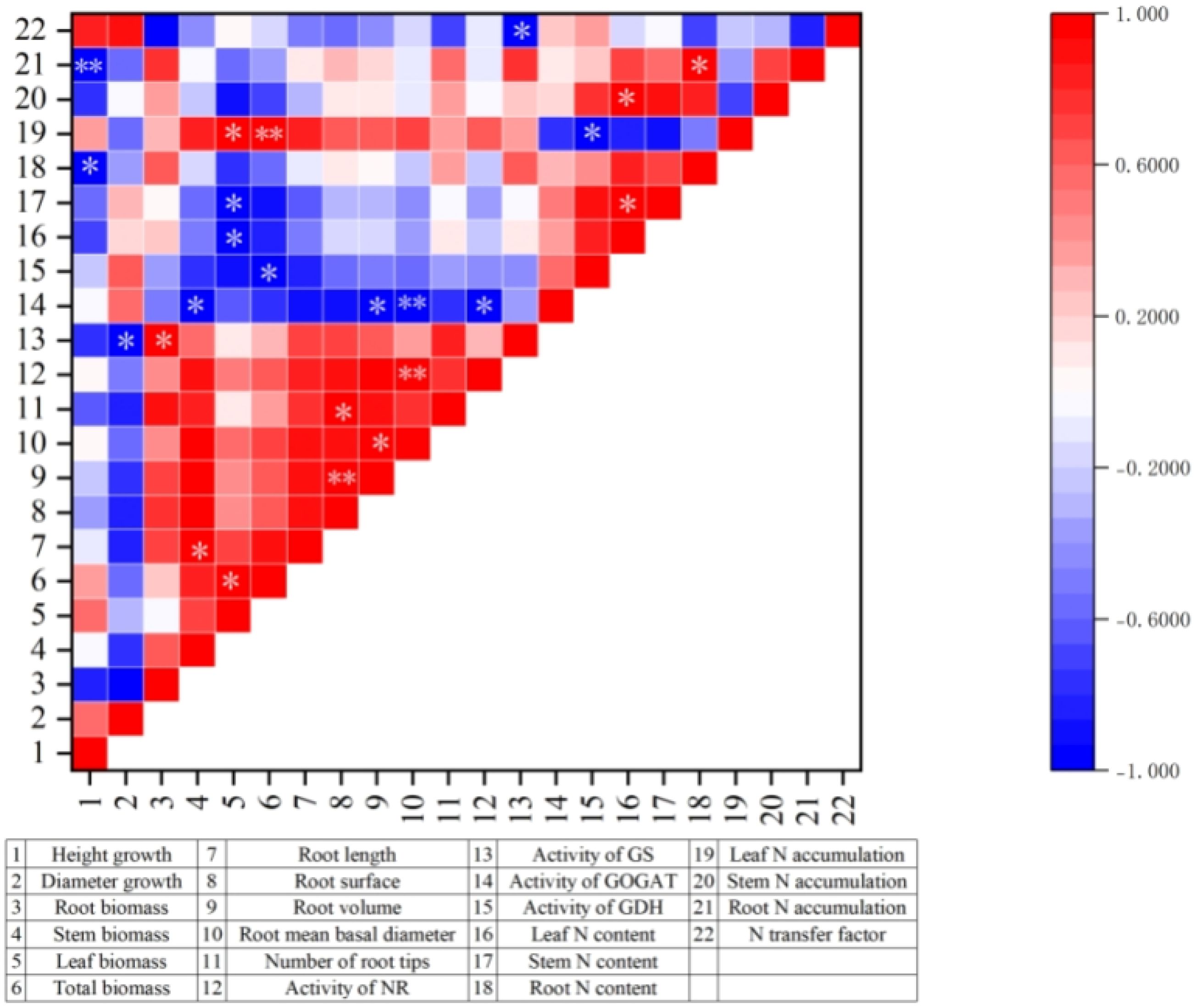
Figure 7. Correlations among final trait values in Machilus thunbergii seedlings across fertilization treatments. Asterisk indicates that the correlation is significant at the 0.05 level (two-tailed). Two asterisks represent significance at the 0.01 level (two-tailed).
In contrast, negative correlations were primarily concentrated between the growth indicators and the nitrogen metabolism components, including the key enzyme and nitrogen content/accumulation in various tissues. This suggests that, during periods of rapid growth, M. thunbergii seedlings tend to preferentially allocate assimilated resources to structural growth rather than to storage functions, reflecting a strategic resource allocation trade-off (Li et al., 2006).
Overall, the correlation analysis revealed tightly coupled synergistic and trade-off relationships among nutrient allocation, root system architecture, and the nitrogen metabolism system in response to the different nitrogen fertilizers, presenting a coherent physiological response profile. Consequently, the seedling response is a systematic process, representing the integrated outcome of an internal resource allocation and a balance between organ growth and nitrogen metabolic function.
4 Discussion
4.1 Effects of different nitrogen fertilizers on the growth and root morphology of M. thunbergii seedlings
Nitrogen fertilizer application has a significant impact on plant growth with species-specific requirements. Appropriate nitrogen application that meets plant needs promotes growth, whereas improper application restricts development (Su et al., 2010). Single nitrogen application significantly promotes plant growth at the early stage; however, this exhibits an inhibitory effect on plant growth with prolonged treatment (Sun et al., 2018). In this experiment, all three nitrogen fertilizers applied at 3 g per plant induced alterations in the leaves and roots of M. thunbergii seedlings, with the most severe symptoms observed under sodium nitrate treatment.Height and diameter growth are reliable indicators of the seedling growth status (Zhu et al., 2018). Studies on Cinnamomum camphora have demonstrated that an appropriate nitrogen application rate promotes growth in the seedling height and diameter (Dong E.Y, et al., 2025). Similarly, research on Machilus pingii reported a pattern of initial increase followed by a decrease in these growth metrics with increasing application of nitrogen (Deng et al., 2020). In contrast, the present study found that the height and diameter growth of M. thunbergii seedlings under the three different nitrogen fertilizer treatments were consistently lower than those in the control group. This indicates that the applied nitrogen concentrations are supra-optimal, leading to growth inhibition. The collective findings from these studies delineated a spectrum of growth responses to nitrogen fertilization. Furthermore, this experiment provided a basis for the selection of the most suitable fertilizer type for M. thunbergii under high-nitrogen stress conditions. The results of this experiment are in line with studies in Hovenia acerba (Zhang et al., 2018), Atropa belladonna (Wei et al., 2017), and Nicotiana tabacum (Xing and Ma, 2016).
The accumulation of biomass enhances plant tissue nutrient storage, demonstrating the influence of external conditions on plant growth (Yang et al., 2020). Research on C. camphora demonstrated that appropriate nitrogen application can enhance the biomass accumulation across plant parts and increase the proportional biomass allocation to stems and leaves (Dong et al., 2025). The study on M. pingii further revealed that the biomass in all components exhibited an initial increasing and then decreasing trend with increased nitrogen application (Deng et al., 2020). In the present experiment, however, none of the three nitrogen fertilizers significantly increased the total biomass accumulation of M. thunbergii seedlings compared with the control (p > 0.05). This indicates that the dosage of 3 g nitrogen per seedling is ineffective in altering biomass accumulation. Collectively, these findings outline a continuum of nitrogen response patterns: from a significant promotion in C. camphora, to a hump-shaped response in M. pingii, and finally to no significant response in M. thunbergii under the given conditions.
Notably, urea treatment resulted in increased biomass compared with the other nitrogen forms, which is likely linked to an optimized carbon–nitrogen balance. Yang et al. (2008) postulated that nitrogen fertilization enhances plant biomass primarily by boosting the photosynthetic capacity. The authors suggested that nitrogen application triggers a response mechanism in mesophyll cells, promoting chlorophyll synthesis and increasing its content, which in turn facilitates photosynthesis. In this experiment, the increased biomass of the M. thunbergii seedlings under urea treatment is likely linked to an optimized carbon–nitrogen balance. Urea presumably supplied ample nitrogenous compounds, such as chlorophyll and key photosynthetic enzymes, thereby directly enhancing the leaf photosynthetic capacity. The consequent increase in photosynthetic products (carbon skeletons) could then provide sustained energy and substrates for nitrogen uptake and assimilation (Yan et al., 2025). This positive feedback loop between carbon and nitrogen metabolism may represent the fundamental physiological basis for the significant biomass accumulation observed in the urea-treated seedlings.
In terms of root development, all three nitrogen treatments exhibited superior root morphology compared with the control, indicating that all three nitrogen fertilizers significantly enhanced root development, with the following efficacy ranking: urea treatment > ammonium sulfate treatment > sodium nitrate treatment. This is consistent with the findings for Paeonia lactiflora (Wang et al., 2025). In this experiment, the sodium nitrate treatment resulted in lower values for many of the seedling growth indicators compared with the control group. However, the same treatment led to greater values for root surface area, root volume, and total root tip number compared with the control, indicating a dichotomous effect: at the application rate of 3 g per seedling, sodium nitrate appears to exert phytotoxic effects on the overall growth, while its nitrate nitrogen component specifically promotes root system development. This phenomenon is consistent with the research by Zhang et al. (2024) on Ormosia henryi seedlings. Furthermore, the seedlings treated with urea exhibited superior root morphology compared with those treated with ammonium sulfate. This suggests that this fertilization level of amide nitrogen fertilizers enhances root development more effectively than ammonium nitrogen fertilizers. A similar pattern of differential growth responses to nitrogen forms was reported by Zhang et al. (2023) on Gossypium hirsutum. The final correlation analysis elucidated why the urea-treated seedlings exhibited superior height, diameter, and biomass compared with the other treatments. The significant positive correlation between biomass and the root system parameters indicates that the vigorous growth of the seedlings results from a positive feedback loop involving a highly efficient nitrogen assimilation and a well-developed root system.
4.2 Effects of the different nitrogen fertilizer treatments on the activity of nitrogen-metabolizing enzymes in the leaves of M. thunbergii seedlings
Numerous studies have shown that nitrogen fertilization affects the activity of the nitrogen metabolic enzymes in plants (Ning et al., 2019). Zhao et al. (2017) demonstrated that moderate nitrogen fertilization enhances the leaf NR and GS activities within optimal ranges; however, these activities decline when nitrogen exceeds threshold levels. This is reflected in this study. For instance, the leaf NR activity in the nitrogen-treated seedlings was generally higher than that in the control from 0 to 120 DAT; however, by 150 DAT, the activity in the ammonium sulfate treatment had fallen below that of the control. As for the leaf GS activity, all of the nitrogen treatments surpassed the control at 60 DAT, but with the ammonium sulfate treatment dropping below the control by 90 DAT, with a general declining trend in the GS activity observed over time across all nitrogen treatments. The results of this study indicate that the sodium nitrate treatment generally results in a higher NR activity compared with the ammonium sulfate treatment, suggesting that, at 3 g per plant, nitrate nitrogen has a better effect on the leaf NR activity than ammonium nitrogen. The experiment conducted by Zhang et al. (2024) yielded the same results. GS and GOGAT are the key enzymes in the plant nitrogen assimilation pathway, the “GS/GOGAT pathway,” and their activities directly reflect the nitrogen assimilation efficiency (Liu et al., 2011). Compared with other nitrogen fertilizers, an appropriate increase in ammonium nitrogen application can promote the plant GS activity at the same nitrogen level, in turn enhancing nitrogen metabolism (Yang et al., 2007). In the later stage of the experiment, the GS and GOGAT activities in the seedling leaves in the ammonium sulfate treatment were higher than that in the sodium nitrate treatment, indicating that, compared with nitrate nitrogen, ammonium nitrogen is beneficial for improving the leaf GOGAT activity. Zhang et al. (2017) believed that increasing the ratio of NH4+–N/NO3−–N in fertilizers can effectively enhance the GOGAT activity in seedling leaves. These two results mutually confirm each other. Throughout the experimental period, the urea-treated M. thunbergii seedlings exhibited the highest leaf GS enzyme activity, alongside superior growth performance (i.e., seedling height, ground diameter, and biomass) and the most developed root system architecture among the three nitrogen fertilizer treatments. This correlation suggests a synergistic relationship among these parameters. A superior root system architecture can significantly enhance the nitrogen uptake capacity of seedlings (Ren et al., 2021; Jing et al., 2023). To accommodate the increased nitrogen influx, the seedlings likely upregulated key assimilatory enzymes such as GS for efficient nitrogen assimilation (Tang et al., 2023). This process ultimately provided ample foundational materials for rapid plant growth, resulting in a significant growth advantage. It can be inferred that the promoting effect of urea treatment on M. thunbergii seedlings is systematic. Urea application enhances root development, and the resulting improved root system architecture not only ensures a stable nitrogen uptake but also supports the efficient and stable operation of the enzymatic systems for nitrogen metabolism in the leaves, thereby optimizing the overall growth performance of the plant. This systemic promoting mechanism is supported by the results of the correlation analysis in this study. The data revealed significantly positive correlations between the root biomass and GS activity, as well as between the root mean basal diameter and NR activity, indicating that modifications in the root morphology are closely linked to nutrient absorption and the overall nitrogen metabolism of the seedlings. Both the root mean diameter and the root volume showed significant negative correlations with the GOGAT activity, suggesting that specific changes in the root architecture may involve complex trade-offs with nitrogen assimilation. This aspect warrants further in-depth investigation.
4.3 Effects of the different nitrogen fertilizers on the nitrogen accumulation and translocation in M. thunbergii seedlings
The nitrogen absorption and translocation in plants can be characterized by indicators such as nitrogen content and nitrogen accumulation (Lian et al., 2022). Studies have shown that the application of nitrogen fertilizer has a significant impact on the nitrogen accumulation of plants, and increasing the application of nitrogen fertilizer can enhance the nitrogen accumulation in the aboveground part (Meng et al., 2025). In this study, nitrogen fertilizer application significantly increased both the total nitrogen content and the nitrogen accumulation in all tissues of M. thunbergii seedlings compared with the control. This indicates that all three nitrogen fertilizers effectively enhance the nitrogen uptake and accumulation at the 3-g per plant dosage, a finding consistent with previous research. The study by Han et al. (2025) on Avena sativa found that the nitrogen accumulation of plants with nitrogen applied at different concentrations all exceeded the control, which is similar to that in our study. The leaf nitrogen accumulation of the urea-treated seedlings was higher than that of the control, but that of the seedlings treated with ammonium sulfate and sodium nitrate was lower than that of the control. This could be due to the late stage of the experiment. Compared with the urea treatment, the ammonium sulfate and sodium nitrate treatments induced a more severe toxicity, leading to leaf wilting and a marked reduction in leaf biomass. This indicates that the application of urea is more effective for the uptake and accumulation of nitrogen in M. thunbergii seedlings at the 3-g per seedling application rate. The study by Dang et al. (2013) on Triticum aestivum found that, during the growth period, the nitrogen accumulation in the leaves of the plant was greater than that in the stems, which is consistent with the results of the experiment on M. thunbergii. These comparative findings are summarized in the table below. In this study, the nitrogen transfer factor of the M. thunbergii seedlings in the nitrogen treatments was all lower than that of the control, indicating that, at the 3-g per seedling application rate, the three nitrogen fertilizer types do not enhance the nitrogen transfer capacity of the seedlings, but improve their root nitrogen uptake. The seedlings treated with ammonium sulfate had the lowest transfer factor among the three nitrogen fertilizer treatments, indicating that, at this fertilization level, ammonium nitrogen has the greatest inhibitory effect on nitrogen transport in seedlings. This inhibitory effect represents one aspect of the regulatory role of nitrogen metabolism in plant growth. The correlation analysis in this study further elucidated the dual nature of this regulation. On the one hand, the positive correlation between GS activity and biomass indicates a driving effect of the nitrogen assimilation capacity on seedling growth. On the other hand, the significant negative correlation between height growth and root nitrogen content reveals a trade-off in resource allocation between supporting seedling growth and maintaining the nitrogen reserves. It is precisely this negative correlation pattern that explains why the height and the diameter growth of the seedlings in the three nitrogen treatment groups were lower than those in the control group, thereby providing insights into the resource allocation strategy of M. thunbergii in nitrogen-enriched environments.
5 Conclusion
At the 3-g per seedling application rate, all three nitrogen fertilizers produced a spectrum of toxicity in M. thunbergii seedlings. The effects of the different nitrogen fertilizer treatments on M. thunbergii seedlings were compared and summarized in Table 9. Although the application of nitrogen fertilizers inhibited the growth in the height and basal diameter of the seedlings, even triggering leaf scorching (with sodium nitrate being the most detrimental and urea the least), they conversely promoted root development. Consequently, the root morphology and biomass in all treatment groups surpassed those of the control. The seedlings treated with urea showed an increase in biomass, while those treated with ammonium sulfate and sodium nitrate did not exhibit this phenomenon. Therefore, sodium nitrate had the greatest toxic effect on M. thunbergii seedlings, followed by ammonium sulfate and urea. Among the nitrogen fertilizers tested, urea proved superior at the application rate of 3 g per plant for 3-year-old M. thunbergii seedlings. Nitrogen fertilization had dose-dependent effects: promoting growth at low levels, but causing inhibition or toxicity at higher concentrations with prolonged treatment. Nitrogen application enhanced the nitrogen-metabolizing enzyme activity in M. thunbergii leaves. The leaf NR, GS, and GOGAT activities showed initial increases, followed by declines with treatment duration, while the GDH activity exhibited sustained increases. All treatments increased the tissue nitrogen content and the nitrogen accumulation of the stems and roots, but decreased the transfer factor. While urea enhanced the foliar nitrogen accumulation, ammonium sulfate and sodium nitrate reduced it.
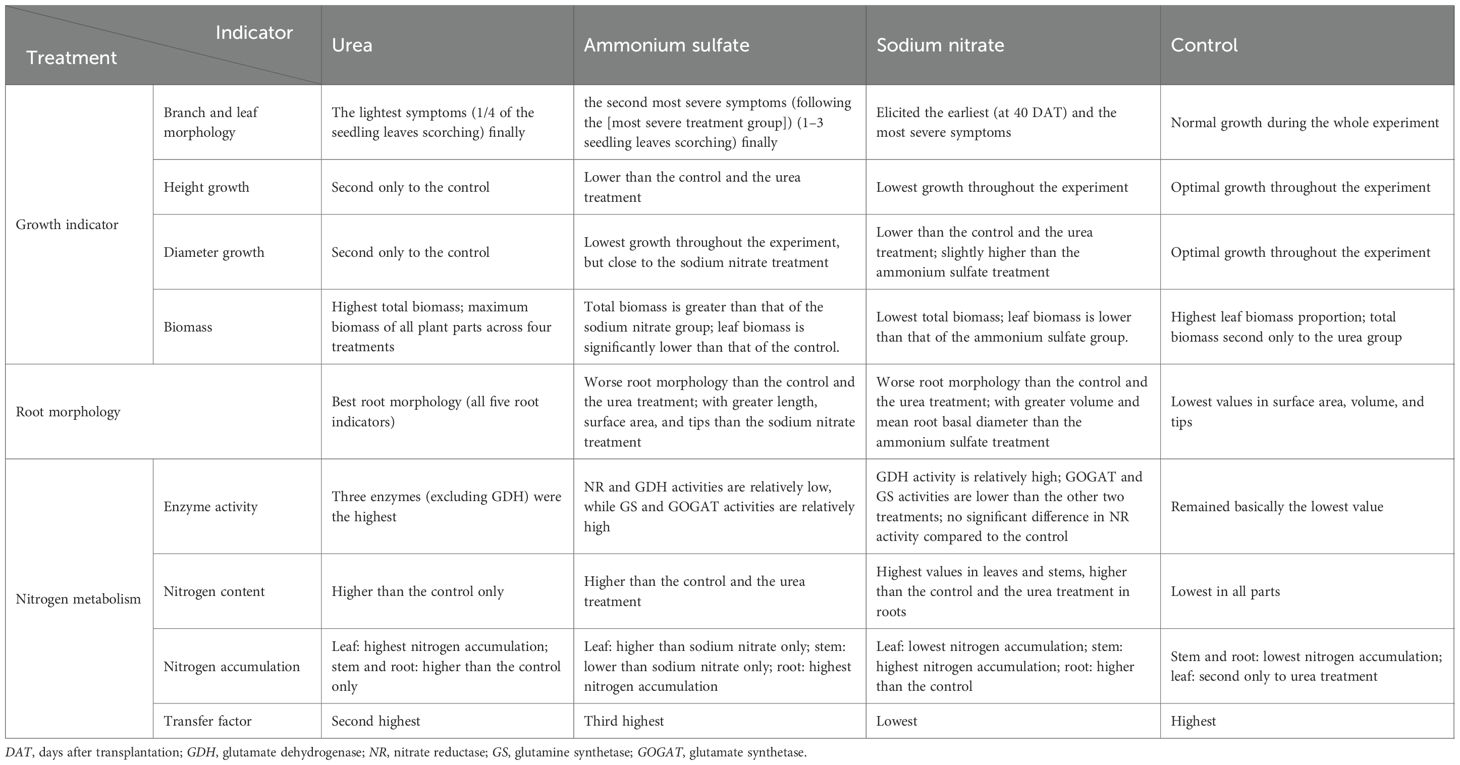
Table 9. Comparative summary of the effects of nitrogen fertilizer treatments on Machilus thunbergii seedlings.
This study demonstrated that, among the tested nitrogen fertilizers, urea exhibited significant superiority in promoting the growth of M. thunbergii seedlings and in enhancing the activity of the key nitrogen metabolism enzymes, establishing it as a preferred fertilizer source for the seedling stage. Notably, the different forms of nitrogen exerted inhibitory effects of varying intensities on the seedling nitrogen transport capacity. However, the underlying mechanisms remain unclear and warrant further investigation. Furthermore, this research did not encompass changes in the content of nutritional components such as proteins, soluble sugars, and starch within the seedlings, which are closely linked to nitrogen metabolism and carbon–nitrogen balance. This study revealed synergistic and trade-off effects between organ growth and nitrogen metabolism functions. Building upon this discovery, future research should focus on the following directions to advance the conservation of M. thunbergii species. Firstly, a systematic evaluation of integrated carbon–nitrogen metabolic responses is needed. On the basis of existing physiological and growth indicators, simultaneous analysis of the dynamic changes in nutrients such as proteins, soluble sugars, and starch in seedlings after fertilization should be conducted, which will enable a more comprehensive assessment of the effects of nitrogen fertilizers at the level of carbon–nitrogen balance, thereby revealing the chemical basis of resource allocation and trade-offs. Secondly, an in-depth analysis of the molecular mechanisms underlying the advantages of urea is essential. Moving beyond current phenotypic observations, research should delve into the key pathways through which urea regulates efficient nitrogen metabolism in M. thunbergii at the molecular level, including protein activity, enzyme kinetics, and related gene expression, which will provide a theoretical basis for precision fertilization management. Finally, the regulatory principles governing resource allocation trade-offs should be elucidated. In response to the negative correlation between seedling growth and nitrogen storage observed in this study, future investigations should employ methods such as controlling source–sink relationships or applying metabolic inhibitors. This approach will allow for an in-depth study of the regulatory signals governing resource allocation and nitrogen metabolism in M. thunbergii seedlings under nitrogen-enriched environments, thereby refining the theory of resource allocation in environmental adaptation.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author contributions
FS: Methodology, Writing – review & editing. HF: Formal analysis, Writing – original draft, Software. XL: Methodology, Writing – original draft. YB: Formal analysis, Writing – review & editing. YL: Data curation, Investigation, Writing – original draft. BT: Data curation, Investigation, Writing – original draft. YZ: Methodology, Writing – review & editing.
Funding
The author(s) declared that financial support was received for this work and/or its publication. This research was funded by Science and Technology Project of Shanghai Greening and City Appearance Management Bureau (G241209) and Key R&D Program of Shandong Province (2024LZGC003).
Conflict of interest
The authors declared that this work was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declared that generative AI was not used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1684502/full#supplementary-material
References
Chen, J. L., Xie, J., Sun, Y., Shi, F. H., Shen, Y. B., Zhang, Y. H., et al. (2023). Effects of substrate compositions on growth of container seedlings of ×Taxodiomera peizhongii. J. Northeast Forestry University. 51, 33–38. doi: 10.13759/j.cnki.dlxb.2023.07.021
Dang, H. K., Li, R. Q., Li, Y. M., Sun, Y. H., Zhang, X. W., and Liu, M. X. (2013). Absorption, accumulation and distribution of nitrogen in super-highly yielding winter wheat. J. Plant Nutr. Fertilizer 19, 1037–1047. doi: 10.11674/zwyf.2013.0502
Deng, W. D., Xu, D. H., and Li, F. W. (2020). The effect of nitrogen application rate on the growth and photosynthetic characteristics of Machilus Pingii seedlings. Cent. South For. Inventory Planning. 39, 53–57. doi: 10.16166/j.cnki.cn43-1095.2020.04.012
Dong, W. N., Lan, H. J., Hao, Q. Q., Qin, B. P., Zhang, M., Cai, R. G., et al. (2025). The effect of nitrogen sulfur interaction on dry matter accumulation and grain yield of winter wheat. J. Triticeae Crops. 45, 194–203. doi: 10.7606/j.issn.1009-1041.2025.02.07
Dong, E. Y., Zhang, T., Wu, Y. F., Zhang, Y. T., Wu, Q. H., Liu, X. L., et al. (2025). The effect of nitrogen fertilizer on the growth, photosynthetic characteristics, and leaf essential oil yield of Cinnamomum camphora seedlings. Shandong Agric. Sci. 57, 111–118. doi: 10.14083/j.issn.1001-4942.2025.05.014
Dong, L. J., Zhu, X. T., Lin, X. Z., and Xu, Z. D. (2011). The effect of fertilization on the growth of container seedlings of the family Lauraceae. Northern Horticulture. 13, 73–77.
Fan, X. T., Zhao, C. B., Li, M. Y., Hou, J. L., Xu, Y., and Meng, J. J. (2025). Response of root growth and nitrogen metabolism of cucumber with different nitrogen efficiencies to nitrogen. Jiangsu Agric. Sci. 53, 168–175. doi: 10.15889/j.issn.1002-1302.2025.10.022
Farhan, M., Sathish, M., Kiran, R., Mushtaq, A., Baazeem, A., Hasnain, A., et al. (2024). Plant nitrogen metabolism: Balancing resilience to nutritional stress and abiotic challenges. Phyton-International J. Exp. Bot. 93, 581–609. doi: 10.32604/phyton.2024.046857
Ge, J., Du, Y. R., Wang, Q., Xu, X., Li, J., Tao, J., et al. (2024). Effects of nitrogen fertilizer on the physicochemical, structural, functional, thermal, and rheological properties of mung bean (Vigna radiata) protein. Int. J. Biol. Macromolecules 260, 2. doi: 10.1016/j.ijbiomac.2024.129616
Gong, J., Wang, B., Zhang, Z., Song, L., Zhang, S., Zhang, W., et al. (2024). Temporal coupling of plant and microbial nitrogen uptake and plant nitrogen reallocation improve grassland nitrogen retention under nitrogen addition. J. Arid Environments 224. doi: 10.1016/j.jaridenv.2024.105195
Han, X., Wang, Y., Han, D. Y., Zhao, X. Y., Li, X. T., Zhang, H. J., et al. (2025). The effect of nitrogen application on the yield, nitrogen accumulation, and interspecific relationships of oat intercropping clover. Crops 1, 187–193. doi: 10.16035/j.issn.1001-7283.2025.01.023
Huang, J. R., Xie, J. L., Zhang, D. S., Fan, J. M., Wei, J. Q., Zhang, Z. H., et al. (2013). The value and technology of seedling nursing and afforestation of Machilus thunbergii. Guangdong Forests Technol. 29, 101–103.
Jiangxi Provincial Administration for Market Regulation. (2018). Technical Regulations for Seed Collecting and Seed Nursing in Machilus thunbergii (DB36/T 662-2018) [S]. (Nanchang: Jiangxi Standards Press).
Jing, B., Shi, W. J., and Ming, D. (2023). Nitrogen application levels based on critical nitrogen absorption regulate processing tomatoes productivity, nitrogen uptake, nitrate distributions, and root growth in Xinjiang, China. J. Arid Land 15, 1231–1244. doi: 10.1007/s40333-023-0108-2
Kusano, M., Fukushima, A., Redestig, H., and Saito, K. (2011). Metabolomic approaches toward understanding nitrogen metabolism in plants. Journal of experimental botany. 4, 1439–1456. doi: 10.1093/jxb/erq417
Langworthy, A. D., Corkrey, R., Rawnsley, R. P., and Hills, J. L. (2023). Shoot nitrogen concentrations required for optimal herbage growth of multiple perennial ryegrass (Lolium perenne L.) cultivars relative to modern summer-active cocksfoot (Dactylis glomerata L.) and tall fescue (Festuca arundinacea Schreb.) cultivars. Grass Forage Sci. 78, 525–535. doi: 10.1111/gfs.12627
Li, L. H., Chen, Y. Q., Wu, C. P., Yao, L. J., Wang, Z. G., Li, T. T., et al. (2024). Prediction of distribution area and analysis of major environmental variables of Machilus thunbergii in Zhejiang Province. J. Zhejiang Forestry Sci. Technol. 44, 9–15. doi: 10.11674/zwyf.2013.0502
Li, X., Li, X. J., Shi, F. H., and Shen, Y. B. (2023). Effects of low-release fertilizer amount on the growth of container seedlings of Phoebe sheareri. J. Northwest Forestry Univ. 37, 152–157. doi: 10.3969/j.issn.1001-7461.2022.04.20
Li, L., Wu, K., Qi, S. W., and Yan, X. F. (2022). Propagation technology of rare native tree species Machilus thunbergii south by cutting propagation in Yuntai Mountain, Jiangsu Province. Special Economic Anim. Plants. 25, 103–104.
Li, Y. L., Xu, Y. L., Tang, J. R., He, Y. N., Wang, D., Li, J. F., et al. (2023). The effect of nitrogen and phosphorus combined application on the growth and nutrient accumulation of Pinus yunnanensis seedlings. J. Zhejiang A&F University. 40, 115–125. doi: 10.11833/j.issn.2095-0756.20220223
Li, L. H., Chen, Y. Q., Wu, C. P., Yao, L. J., Wang, Z. G., Li, T. T., et al. (2024). Prediction of distribution area and analysis of major environmental variables of Machilus thunbergii in Zhejiang Province. Journal of Zhejiang Forestry Science and Technology, 44, 9–15. doi: 10.3969/j.issn.1001-3776.2024.05.002
Li, X., Yan, X. F., and Yu, T. (2006). Effect of color films shading on Phellodenron amurense seedlings biomass and primary assimilation enzyme activities. Chin. J. Appl. Ecol. 11, 2020–2023. doi: 10.13287/j.1001-9332.2006.0398
Lian, H. S., Zou, P. H., Li, X. X., Lin, L. J., and Liao, M. A. (2022). Effect of 2,4-epibrassinolide on the nitrogen uptake and translocation of grape seedlings and nitrogen content in soil. J. Yunnan Agric. Univ. (Natural Science). 37, 170–176. doi: 10.12101/j.issn.1004-390X(n)202105048
Liu, C. N., Cui, X. Y., Guo, Y. F., and Zheng, H. (2011). The effect of ammonium nitrogen and nitrate nitrogen ratio on the growth of Larix gmelinii seedlings. J. Northeast Forestry University. 39, 28–30. doi: 10.13759/j.cnki.dlxb.2011.01.025
Ma, L. Y. and Huang, J. J. (2019). Research progress and development and application prospects of native tree Machilus thunbergii. Prot. For. Sci. Technol. 6, 66–68. doi: 10.13601/j.issn.1005-5215.2019.06.026
Meng, Z. L., Ni, X. F., Zhu, Q., Wang, Q., Lv, X. L., Yan, X. Q., et al. (2025). The effect of nitrogen application rate and density interaction on nitrogen accumulation, transport and yield of nutritional organs in wheat after flowering. Jiangsu Agric. Sci. 53, 70–79. doi: 10.15889/j.issn.1002-1302.2025.12.010
Ministry of Agriculture and Rural Affairs of the People’s Republic of China.(2011). Determination of nitrogen, phosphorus and potassium in plants (NY/T 2017-2011)[S]. (Beijing: China Agriculture Press).
Mostafa, Y., Farhan, K. N., Hassan, A., and Yassi, B. (2021). The plant transfer factor of natural radionuclides and the soil radiation hazard of some crops. Environ. Monit. Assess. 193(6). doi: 10.1007/s10661-021-09061-7
Ning, G. X., Ma, Z. H., Mao, J., Li, W. F., Wang, Y., Hu, Z. J., et al. (2019). Effects of nitrogen application rates on the leaf qualityof‘Cabernet Gernischet’in desert areas. J. Fruit Sci. 36, 1150–1160. doi: 10.13925/j.cnki.gsxb.20190046
Qian, M. F. (2022). Seed cultivation and protective forest construction techniques for Machilus thunbergii in the mountainous areas of southern Anhui province. Rural Sci. Technol. 13, 1111–1114. doi: 10.19345/j.cnki.1674-7909.2022.05.038
Qiu, C. C., Hu, Y. T., Yan, H., Li, H. J., Yu, H. N., Zhao, T. T., et al. (2024). The effect of nitrogen application rate on chlorophyll content and photosynthetic characteristics of leymus chinensis. Hubei Agricultral Sci. 63, 141–146. doi: 10.14088/j.cnki.issn0439-8114.2024.11.024
Ren, H., Gao, G. Q., Ma, Y. Y., Li, Z. W., and Gu, J. C. (2021). Root nitrogen uptake and its relationship with root morphological and chemical traits in Pinus koraiensis at different ages. J. Beijing Forestry University. 43, 65–72. doi: 10.13759/j.cnki.dlxb.2023.05.019
Ren, W. L., Luo, S., Shi, F. H., Jin, X. C., Lu, Y. Z., and Han, B. (2023). Effects of bud-seedling root pruning of Ormosia hosiei Hemsl. et Wils. on the seedling growth. J. Northeast Forestry Univ. 51, 20–25+29. doi: 10.13759/j.cnki.dlxb.2023.05.019
Saurav, D., Snkita, M., Sahu, K., Panday, D., Ghimire, D., and Maharjan, B. (2024). Nitrogen dynamics as a function of soil types, compaction, and moisture. PloS One 19 (4). doi: 10.1371/journal.pone.0301296
Shipley, B. and Meziane, D. (2002). The balanced-growth hypothesis and the allometry of leaf and root biomass allocation. Funct. Ecol. 16, 326–331. doi: 10.1046/j.1365-2435.2002.00626.x
Song, G. and Fei, Y. J. (2013). The effects of different fertilization formulas on the growth and photosynthetic physiology of Lindera megaphylla. J. Yangtze University(Natural Sci. Edition) 10, 16–17+51+109-110. doi: 10.16772/j.cnki.1673-1409.2013.11.014
Su, J. Q., Li, X. R., He, H. Y., Jia, R. L., and Gao, Y. H. (2010). Responses of herbaceous layer vegetation to nitrogen fertilization in Shapotou area. Chin. J. Ecol. 29, 309–313. doi: 10.13292/j.1000-4890.2010.0010
Sun, M. H., Wu, L., and Xie, S. X. (2018). The effect of ammonium nitrate nutrition on glutamine synthetase activity and related gene expression in Citrus aurantium seedlings. Plant Physiol. J. 54, 1703–1710. doi: 10.13592/j.cnki.ppj.2018.0232
Tang, Q., Ma, Y., Zhao, L., Song, Z., Yin, Y., Wang, G., et al. (2023). Effects of water and nitrogen management on root morphology, nitrogen metabolism enzymes, and yield of rice under drip irrigation. Agronomy-Basel 13 (4). doi: 10.3390/agronomy13041118
Wang, X., Zhang, X. X., Mi, M., Li, S., Li, H. J., Ma, H. L., et al. (2025). Effects of one-time application of nitrogen fertilizer on growth and photosynthetic characteristics of Paeonia lactiflora. Chin. Agric. Sci. Bull. 41, 61–66.
Wei, D., Hu, R. X., Zhao, Q., and Tang, H. H. (2018). Effects of different concentration nitrogen treatments on the growth of two families of Bauhini apurpurea seedlings. J. Northwest Forestry Univ. 33, 116–122. doi: 10.3969/j.issn.1001-7461.2018.05.18
Wei, Y., Zhang, C. P., Guo, S., Lu, K. H., and Wu, N. B. (2017). Effect off nitrogen form on growth and nitrogen metabolism of Atropa belladonna. Pratacultral Sci. 34, 1669–1676. doi: 10.11829/j.issn.1001-0629.2016-0543
Xing, Y. and Ma, X. H. (2016). The effect of nitrogen forms on root growth and nitrogen utilization of tobacco seedlings. Acta Tabacaria Sin. 22, 52–61. doi: 10.16472/j.Chinatobacco.2015.557
Xu, W., Liu, S., You, X., Mei, Y. Y., Wang, D., and Wang, N. N. (2017). Over-expression of GmATG8c gene enhances low nitrogen tolerance and yield in soybean. Plant Physiol. J. 53, 241–247. doi: 10.13592/j.cnki.ppj.2016.1038
Yan, J. H., Weng, H., and He, T. (2025). Effects of replacing chemical fertilizers nitrogen with organic fertilizers nitrogenon growth characteristics,leaf carbonand nitrogen balance and nitrogen metabolism enzyme activities of hulless barley. J. Triticeae Crop 45, 121–129. doi: 10.7606/j.issn1009-1041.2025.01.14
Yang, Y., Shi, H. R., Ji, L., and Yang, L. X. (2020). Effects of exponential fertilization on growth and root morphology of Tilia amurensis seedlings. J. Nanjing Forestry University(Natural Sci. Edition) 44, 91–97. doi: 10.3969/j.issn.1000£2006.201811061
Yang, S. L., Zhang, F. L., Jia, X. L., Ma, R. K., and Yao, Y. R. (2008). Effects of water and nitrogen coupling on chlorophyll content and photosythetic rate of winter wheat leaves. Acta Agriculturae Boreali-Sinica 23, 161–164.
Yang, L., Zhao, H. W., and Liu, J. H. (2007). The effect of nitrogen application on glutamine synthetase activity and yield in functional leaves of spring maize. J. Northeast Forestry Univ. 3, 320–324. doi: 10.19720/j.cnki.issn.1005-9369.2007.03.007
Yin, D. S. and Zhang, J. Y. (2025). The effects of inorganic nitrogen forms and quality ratios on the growth and physiology of grafted Pinus koraiensis. J. Northeast Forestry University. 53, 65–69. doi: 10.13759/j.cnki.dlxb.2025.09.003
Yu, J. X., Cheng, C., Wang, L. D., Liang, H. Y., Wang, Y. R., and Yan, X. L. (2024). Effects of substrate ph and ratios of ammonium to nitrate on enzymatic activities of nitrogen metabolism and nitrogen content in Cunninghamia lanceolata and Schima superba seedlings. J. Sichuan Agric. Univ. 42, 815–827. doi: 10.16036/j.issn.1000-2650.202402391
Zhang, M. (2023). Study on the community succession characteristics of evergreen broad leaved forest in Xiangtou Mountain. Hunan Forestry Sci. Technology. 50, 87–91. doi: 10.6048/j.issn.1001-4330.2023.04.005
Zhang, D. S., Chen, D. S., and Liu H.N.and Luo, W. Y. (2018). The effect of nitrate to ammonium nitrogen ratio on the growth of Hovenia acerba seedlings. Forestry Environ. Sci. 34, 41–47.
Zhang, S. Y., Chu, G. X., and Liang, Y. C. (2017). The effect of increasing ammonium nutrition on nitrogen metabolism in cotton seedlings under low temperature stress. J. Plant Nutr. Fertilizers. 23, 983–990. doi: 10.11674/zwyf.16394
Zhang, C., Liang, Y., Yin, H., Zhang, Y. R., and Chen, B. L. (2023). The effects of nitrogen fertilizer forms and varieties on the root morphology and nitrogen accumulation of cotton. Xinjiang Agric. Sci. 60, 823–831. doi: 10.3969/j.issn.1003-5710.2023.01.011
Zhang, M. R., Wu, Q. X., Cao, J. W., Wu, X. L., Xiao, Y. Q., and Sun, M. H. (2024). The effect of different nitrogen form ratios on root growth and key enzyme activity of nitrogen metabolism in Ormosia henryi seedlings. Acta Agriculturae Boreali-occidentalis Sinica. 33, 1515–1522. doi: 10.7606/j.issn.1004-1389.2024.08.012
Zhao, C. B., Wang, C. N., Song, S. Y., Wang, Y., Chen, S. S., Mao, L. B., et al. (2017). Effects of nitrogen nutrition on change of key enzyme activity during nitrogen metabolism and nitrogenous compounds in cucumber. J. Jilin Agric. Univ. 39, 139–147. doi: 10.13327/j.jjlau.2017.2528
Keywords: Machilus thunbergii, nitrogen fertilizer, seedling growth, nitrogen metabolism, nitrogen accumulation, nitrogen metabolism enzyme
Citation: Shi F, Feng H, Li X, Zhang Y, Bai Y, Tong B and Lu Y (2025) Effects of different nitrogen treatments on the growth and nitrogen metabolism of Machilus thunbergii seedlings. Front. Plant Sci. 16:1684502. doi: 10.3389/fpls.2025.1684502
Received: 12 August 2025; Accepted: 25 November 2025; Revised: 13 November 2025;
Published: 17 December 2025.
Edited by:
Anoop Kumar Srivastava, Central Citrus Research Institute (ICAR), IndiaReviewed by:
Kamran Iqbal Shinwari, Nanjing Agricultural University, ChinaKhurram Shahzad, Lasbela University of Agriculture, Water and Marine Sciences, Pakistan
Copyright © 2025 Shi, Feng, Li, Zhang, Bai, Tong and Lu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fenghou Shi, c2hpZmVuZ2hvdUBuamZ1LmVkdS5jbg==; Yizeng Lu, bHV5aXplbmdAc2hhbmRvbmcuY24=; Yuhui Zhang, emhhbmd5dWh1aUBsaW55ZS5zaC5jbg==
†These authors have contributed equally to this work