 Chao Zhou1†Jia Wang2†Ting Xu1Kuan Pei3Baoxin Ma1Qingchao Li1Yang Liu1Xinying Ding4Yehui Han1*
Chao Zhou1†Jia Wang2†Ting Xu1Kuan Pei3Baoxin Ma1Qingchao Li1Yang Liu1Xinying Ding4Yehui Han1* Junqiang Wang1*
Junqiang Wang1*- 1Heilongjiang Academy of Agricultural Sciences, Qiqihar, China
- 2College of Agronomy, Heilongjiang Bayi Agricultural University, Daqing, Heilongjiang, China
- 3Tongliao Academy of Agricultural and Animal Husbandry Science, Tongliao, China
- 4Animal Husbandry and Veterinary Branch, Heilongjiang Academy of Agricultural Sciences, Qiqihar, China
Introduction: Manure substitution for chemical nitrogen has the potential to enhance crop yield, improve soil quality, and reduce environmental risks. Soil microorganisms perform critical functions in mediating soil nutrient cycling after the organic manure application. Nonetheless, how organic manure substitution regulates microbial communities to influence soil quality and crop yield remains unclear.
Methods: A one-year field experiment comprising four organic manure substitution rates (25%, 50%, 75%, and 100%) along with a no-substitution control was conducted.
Results and disscussion: Compared to CK, only 25% substitution rate maintained maize yield, while 50‒100% manure substitution decreased maize yield by 15.9‒67.2%. This is primarily due to the decrease in root biomass (13.5‒29.1%), length (14.8‒43.3%), surface area (14.1‒48.8%) and volume (17.9‒53.4%). Manure substitution only increased soil quality index by 44‒55% in the 0-20 cm depth, mainly as a result of increased contents of soil organic C, total nitrogen, microbial biomass C and N, and enzyme activities. Moreover, manure substitution significantly increased the abundance of Actinobacteriota, Acidobacteriota, Gemmatimonadota, and Methylomirabiliota, with increases ranging from 12% to 101%. The strong correlations between these bacterial taxa and soil nutrient and C/N acquiring enzyme activities highlight their pivotal roles in boosting soil nutrients and enhancing soil quality. Therefore, organic manure substitution can be a sustainable fertilizer regime to enhance soil quality and maintain maize yield in Northeast China, and the optimal substitution rate is 25%.
1 Introduction
The black soil zone in Northeast China constitutes the country’s most crucial grain production base, which contributes over 30% of the national maize output (Wang et al., 2022). However, the overapplication of chemical fertilizers has accelerated the degradation of soil organic matter, leading to reduced nutrient availability (Tian et al., 2022), the disruption of soil structure (Ali et al., 2024), and impaired soil multifunctionality (Wang et al., 2024a). These adverse alterations in soil conditions threaten the stability of agricultural ecosystems and undermine the long-term viability of crop production systems (De Corato et al., 2024). Confronted with these challenges, preventing the degradation of black soils and maintaining their capacity for high and stable crop yields are essential for achieving sustainable agricultural development.
The application of organic manure represents an environmentally sustainable solution to mitigate soil degradation and support long-term agricultural productivity (Wu et al., 2025). China possesses substantial livestock manure resources; however, an estimated 20%–45% of this organic material remains unmanaged, resulting in high risks of environmental contamination and nutrient loss (Zhang et al., 2023). Integrating manure into agricultural systems presents a dual benefit: it enables efficient recycling of organic resources to enhance soil fertility while simultaneously mitigating environmental pollution caused by unmanaged agricultural waste (Yin et al., 2025). Nevertheless, existing studies indicate that the exclusive application of organic manure may fail to meet crop nutrient demands owing to its relatively low nutrient use efficiency (Li et al., 2022). Consequently, the partial substitution of chemical fertilizers with organic amendments has been proposed as a strategy to mitigate the limitations associated with sole manure application (Zhai et al., 2022). Niu et al. (2024) demonstrated that replacing 50% of synthetic fertilizer with manure was a sound option for achieving high crop yield and yield stability in the Guanzhong Plain. However, the effect of organic manure substitution to reduce synthetic nitrogen inputs in Northeast China remains inadequately evaluated.
Soil quality is a key indicator for assessing the capacity of soil to sustain ecosystem services (Salimani et al., 2025). Soil quality was assessed using an area-based method that integrated a set of biotic and abiotic indicators and was significantly influenced by fertilization management (Jia et al., 2022). Previous studies have indicated that incorporating organic amendments can improve soil quality and increase crop yields in various agricultural systems (Li et al., 2025). These benefits are largely attributed to improvements in soil nutrient availability, physical properties, and microbial ecological environment (Ju et al., 2022; Zhu et al., 2025). Sihi et al. (2017) observed that organic fertilizer applied in Sierozem soil can enhance soil water content and reduce soil bulk density compared to conventional mineral fertilization practices. Organic fertilizer can also increase soil quality by enhancing soil organic C, total N, and total P (Tian et al., 2022). At the same time, organic amendments can mitigate inorganic nitrogen leaching—specifically ammonium and nitrate—by enhancing the soil’s nutrient retention capacity (Guo et al., 2025). Soil enzyme activities are widely recognized as sensitive indicators of soil quality (Trasar-Cepeda et al., 2008). Organic amendments have been directly linked to increased activities of soil C- and N-related enzymes, including urease, invertase, catalase, and various hydrolytic enzymes (Feng et al., 2025). While most studies have integrated these indices to evaluate soil quality under organic fertilization applied (Jiang et al., 2022; Zhou et al., 2022), few have focused on the organic manure substitution for chemical N, particularly in the severely soil-degraded regions of Northeast China.
The soil microbiome serves as a fundamental driver of soil functionality and fertility, with its compositional structure and diversity being strongly influenced by fertilization management practices (Li et al., 2022; Yang et al., 2023). The partial application of organic amendments can effectively improve microbial habitat conditions and alter microbial community structure (Zhou et al., 2024). For example, long-term manure application has been proven to significantly increase the abundance of Proteobacteria and Chloroflexi in reddish paddy soil (Cui et al., 2018). Additionally, Liu et al. (2024) demonstrated that the combined application of manure and chemical fertilizers enhanced the abundance of bacterial taxa associated with nutrient cycling and utilization efficiency, particularly Firmicutes, Actinobacteria, and Planctomycetes. Shifts in soil microbial community structure and function are primarily driven by fertilizer-induced alterations in soil physicochemical properties (Ma et al., 2025). Consequently, the high sensitivity of microbial community properties to soil nutrient dynamics makes them reliable biomarkers for evaluating soil quality (Li et al., 2023). However, the integration of soil microbial indicators into soil quality assessments has rarely been evaluated under organic fertilizer substitution for chemical fertilizers.
Therefore, we carried out a one-year field experiment with different manure substitution rates for chemical fertilizer in the Northeast China Mollisol region to 1) evaluate the influence of organic manure substitution on soil physicochemical properties, enzyme activities, and soil quality; 2) identify the response of key soil microbial taxa to varying manure substitution rates; and 3) elucidate the microbial mechanisms through which organic fertilizer substitution influences maize yield and soil quality. We hypothesized that organic manure substitution would improve soil quality and crop yield by enhancing soil physicochemical properties and enzyme activities, as well as by enriching specific soil microbial taxa. To elucidate the microbial mechanisms by which the partial substitution of chemical fertilizers with organic manure influences soil quality and crop productivity, we conducted a comprehensive analysis that integrated soil physicochemical properties, enzyme activities, bacterial communities, and key root architecture traits, with ultimate crop yield. The goal of this experiment was to determine the most effective rate of organic manure replacement that both improves soil quality and maintains maize yield in the Northeast China Mollisol zone.
2 Materials and methods
2.1 Study site
The study was carried out at the Qiqihar Experimental Station, Heilongjiang Academy of Agricultural Sciences (47°36′ N, 123°65′ E; elevation 127 m), situated in Qiqihar City, Heilongjiang Province, China. The experimental site experiences a mid-temperate continental monsoon climate, with a mean annual precipitation of 434.5 mm and an average temperature of 4.7 °C. The soil type was a Chernozem Mollisol (IUSS Working Group WRB 2015). The topsoil layer properties were soil organic carbon (SOC) 21.9 g kg−1, alkaline hydrolyzed nitrogen (AHN) 99.8 mg kg−1, pH 8.4, and bulk density (BD) 1.58 g cm−3.
2.2 Experimental design
A randomized complete block design with three replicates (n = 3) was implemented in 2024. The experiment comprised five treatments: CK, 100% chemical fertilizer N; 25%, 25% dairy manure N substitution; 50%, 50% dairy manure N substitution; 75%, 75% dairy manure N substitution; and 100%, 100% dairy manure N substitution. The control (CK) treatment received fertilizer applications at rates of 187.5 kg N, 42.6 kg P, and 62.2 kg K per hectare, following local agronomic recommendations. Nutrient application rates for all treatments are detailed in Table 1. Maize (cv. ‘Nendan 22’) was planted at a density of 67,500 plants ha−1, with row spacing of 65 cm and plant spacing of 22 cm within rows. The crop residues were incorporated to a depth of 0–20 cm each year. To ensure seed germination, all plots were irrigated with 30 mm of water following maize sowing. Neither pesticides nor herbicides were applied throughout the experiment. All other field management practices aligned with those commonly employed in the region.

Table 1. Details of fertilizer treatments and fertilizer rates (kg ha−1 year−1).
2.3 Soil sampling and analysis
Soil samples were obtained from depth intervals of 0–20 and 20–40 cm following maize harvest on 5 October 2024. Each soil sample was separated into three subsamples. One subsample was stored for analysis of SOC, total nitrogen (TN), available nitrogen (AHN), available phosphorus (AP), and available potassium (AK), following the analytical procedures described by Bao (2000). Soil BD was determined using the ring-knife sampler (volume of 100 cm3) method. Dissolved organic carbon (DOC) and nitrogen (DON) in the potassium sulfate extracts were measured using a carbon–nitrogen analyzer. The second subsample was kept at 4 °C to measure microbial biomass carbon (MBC)/microbial biomass nitrogen (MBN) and six hydrolyzing enzyme activities. The third subsample was stored at −20 °C for DNA extraction. Soil MBC and MBN were quantified using chloroform fumigation extraction (Vance et al., 1987). The activities of six hydrolyzing enzyme activities were identified using the fluorescence methods (Marx et al., 2001). Microbial metabolic limitations were also evaluated, and the calculation methods are according to (Equations 1–3):
Soil quality index (SQI) was calculated by comparing the area on a radar graph comprising all soil indicators (Feng et al., 2024).
where n represents the number of soil indicators used for SQI calculation, and ““ represents the linear score of the ith soil indicator.
2.4 Crop yield and root traits
Maize grain yield was quantified by harvesting within a 6-m2 area per plot, followed by air-drying and mechanical threshing to determine dry grain weight. Root traits were analyzed using an Epson Perfection V750 Pro scanner, and image analysis using the WinRHIZO software quantified root architectural parameters, including length, surface area, and volume. Root biomass was determined following oven-drying at 80°C.
2.5 DNA extraction and sequencing
DNA was extracted using the OMEGA Soil DNA Kit (M5635-02) (Omega Bio-Tek, Norcross, GA, USA) and kept at −20°C before further analysis. Samples were sequenced via Illumina® MiSeq (Genesky Biotechnologies Inc., Shanghai, China). The V4–V5 hypervariable regions of the 16S rRNA gene were amplified with the primers 907R (5′-CCGTCAATTCMTTTRAGTTT-3′) and 515F (5′-GTGCCAGCMGCCGCGG-3′). DNA quality was assessed using a NanoDrop NC2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) for concentration and purity, supplemented by agarose gel electrophoresis for integrity verification.
2.6 Statistical analysis
The normality of all datasets was verified using the Shapiro–Wilk test. The influences of organic manure substitution on maize yield, root biomass, length, surface area, and volume were analyzed using one-way ANOVA. Two-way ANOVA was employed to analyze the effects of organic manure substitution and soil depth (0–20 and 20–40 cm), along with their interaction, on soil physicochemical properties, hydrolase activities, soil quality index, microbial diversity, and dominant bacterial taxa. The correlations between soil physicochemical properties, hydrolase activities, maize yield, and soil quality were also assessed using Mantel tests implemented with the “linkET” package. Meanwhile, key predictors of maize yield and soil quality were evaluated using a random forest model, executed using the “rfPermuta” package. All statistical analyses were performed, and data visualizations were generated using the R software (v.4.1.3; R Core Team, 2022).
3 Results
3.1 Maize yield and root traits
Compared to CK, only 25% manure substitution maintained maize yield, while 50%–100% substitution ratios significantly reduced maize yields by 15.9%–67.2% (Table 2; p < 0.05). Moreover, manure substitution treatments decreased root biomass, root length, root surface area, and volume by 13.5%–29.1%, 14.8%–43.3%, 14.1%–48.8%, and 17.9%–53.4% respectively (Table 2; p < 0.05).
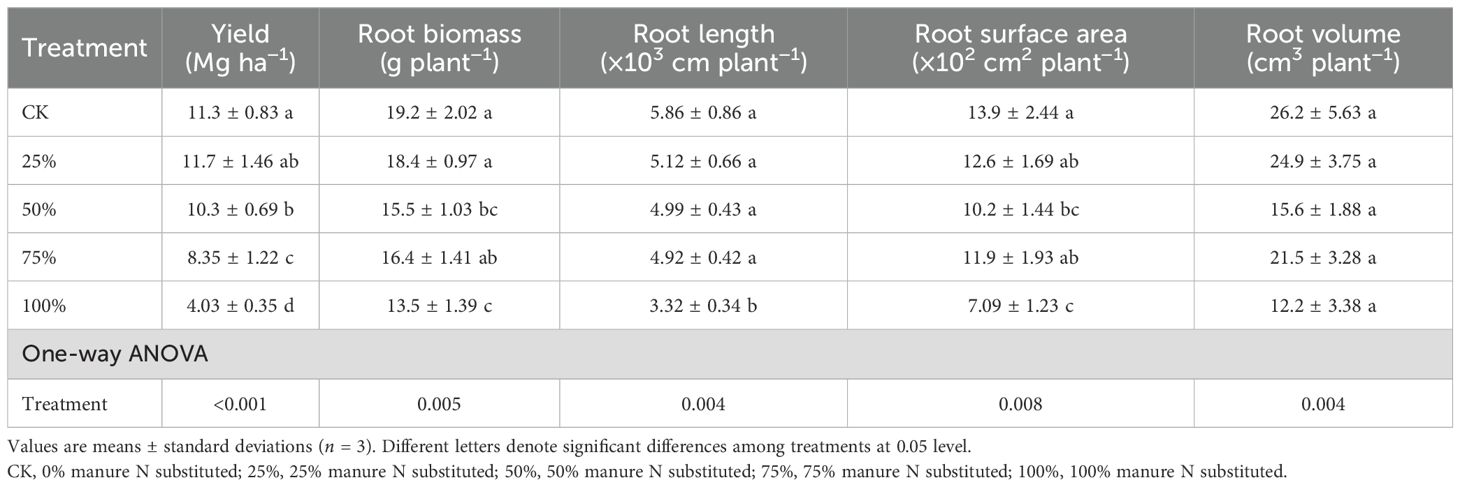
Table 2. Effects of substituting chemical nitrogen with manure on maize yield and root traits.
3.2 Soil indices and quality index
Compared to CK, manure substitution increased the contents of SOC (6%–10%), MBC (28%–42%), MBN (32%–112%), AP (22%–59%), and SW (5%–22%), but decreased DON (23%–31%) in the 0–20-cm depth (Figure 1A; p < 0.05). In the 20–40-cm depth, MBC (92%–112%), MBN (28%–59%), and SW (6%–22%) were increased by manure substitution treatments, whereas TN (2%–17%), DON (45%–52%), AP (42%–60%), AK (17%–35%), and BD (8%–14%) declined compared to CK (Figure 1B; p < 0.05).

Figure 1. Effects of substituting chemical nitrogen with manure on soil physicochemical properties at depths of 0–20 (A) and 20–40 cm (B). CK, 0% manure Nsubstituted; 25%, 25% manure N substituted; 50%, 50% manure N substituted; 75%, 75% manure N substituted; 100%, 100% manure N substituted.SOC, soil organic carbon; TN, total nitrogen; MBC, microbial biomass C; MBN, microbial biomass N; DOC, dissolved organic C; DON, dissolvedorganic N; AHN, alkaline nitrogen; AP, available phosphorus; AK, available potassium; BD, bulk density; SW, soil water content. Values are means ±standard deviations (n = 3). Different letters denote significant differences among treatments at 0.05 level.
Manure substitution increased the activities of β-1,4-glucosidase (BG), β-d-cellobiosidase (CB), β-d-xylopyranoside (XYL), β-1,4-N-acetylglucosaminidase (NAG), acid phosphatase (ALP), and l-leucine aminopeptidase (LAP) by 15%–67%, 138%–239%, 57.9%–117%, 52%–74%, 78%–125%, and 112%–185% in the 0–20-cm depth, respectively (Figure 2; p < 0.05). In contrast, manure substitution decreased the activities of BG (46%–68%), CB (12%–40%), XYL (28%–59%), and NAG (40%–53%), but increased LAP (67%–155%) and ALP (27%–118%) activities in the 20–40-cm depth (Figure 2; p < 0.05). Moreover, the SQI score increased by 44%–55% with manure substitution, but decreased by 13%–33% in the 20–40-cm depth (Figure 3; p < 0.05).
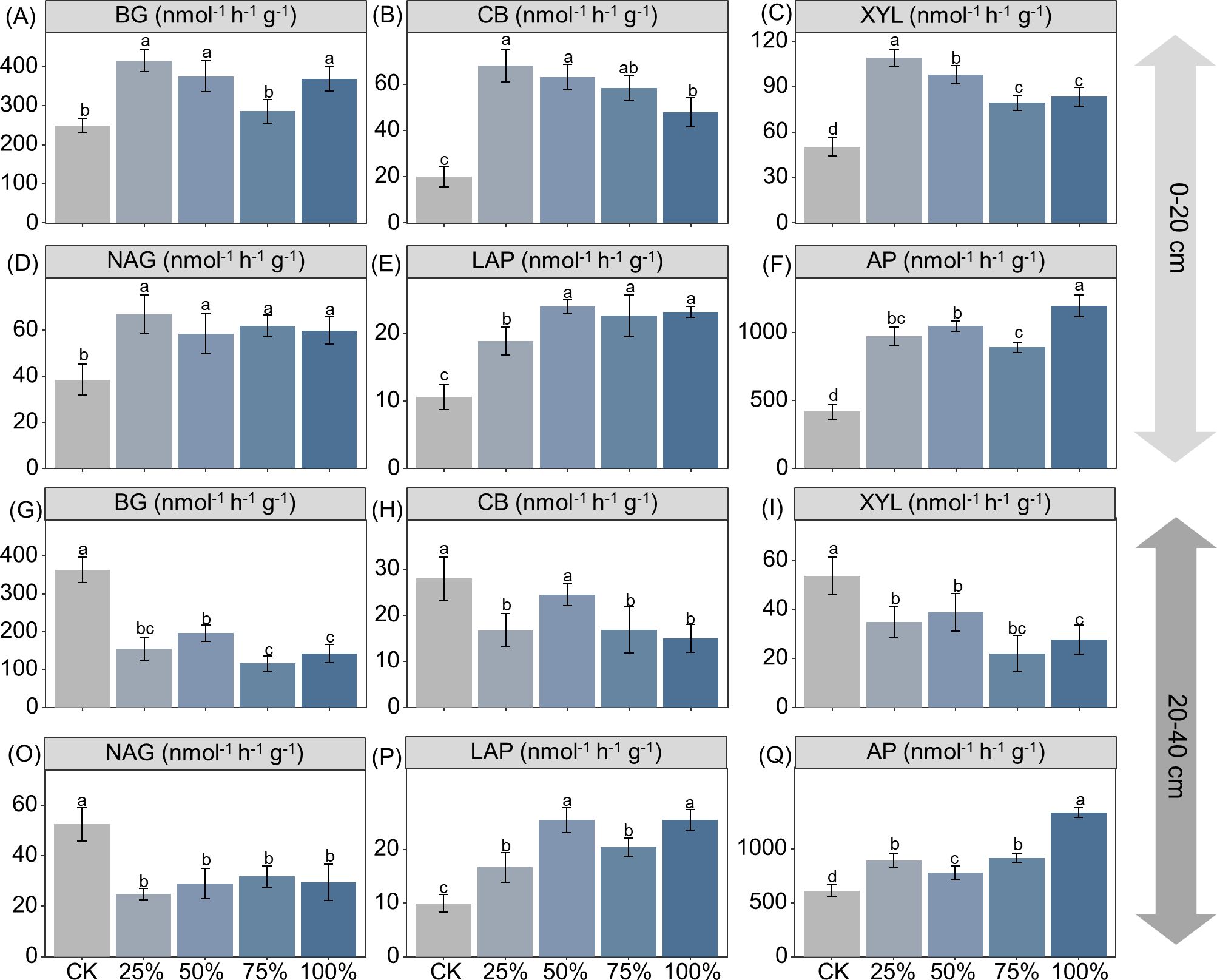
Figure 2. Effects of substituting chemical nitrogen with manure on soil C/N/P acquiring enzyme activities at depths of 0–20 (A–F) and 20–40 cm (G–Q). CK, 0% manure N substituted; 25%, 25% manure N substituted; 50%, 50% manure Nsubstituted; 75%, 75% manure N substituted; 100%, 100% manure N substituted. BG, b-1,4-glucosidase; XYL, b-D-xylopyranoside; CB, b-D-cellobiosidase; NAG, b-1,4-N-acetylglucosaminidase; LAP, L-leucineaminopeptidase; ALP, acid phosphatase. Values are means ± standard deviations (n = 3). Different letters denote significant differences among treatments at 0.05 level.
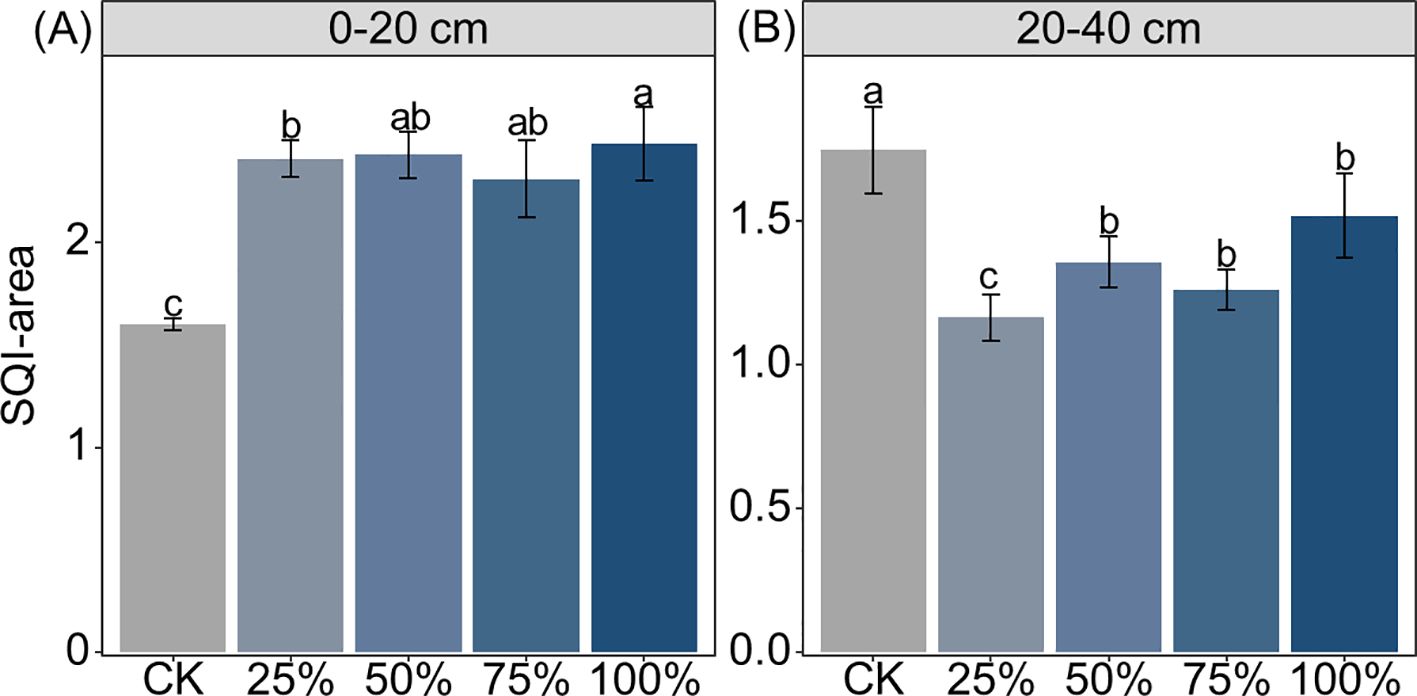
Figure 3. Effects of substituting chemical nitrogen with manure on soil quality index (SQI) area at depths of 0–20 (A) and 20–40 cm (B). CK, 0% manure Nsubstituted; 25%, 25% manure N substituted; 50%, 50% manure N substituted; 75%, 75% manure N substituted; 100%, 100% manure N substituted.Different letters denote significant differences among treatments at 0.05 level.
3.3 Soil bacterial community structure and composition
Compared to CK, manure substitution significantly increased the Simpson index of bacteria by 0.08%–0.14% in the 0–20-cm depth and 0.03%–0.06% in the 20–40-cm depth (Figures 4A, D; p < 0.05). Bacterial community structure was significantly altered by manure substitution treatments (Figures 4B, E).
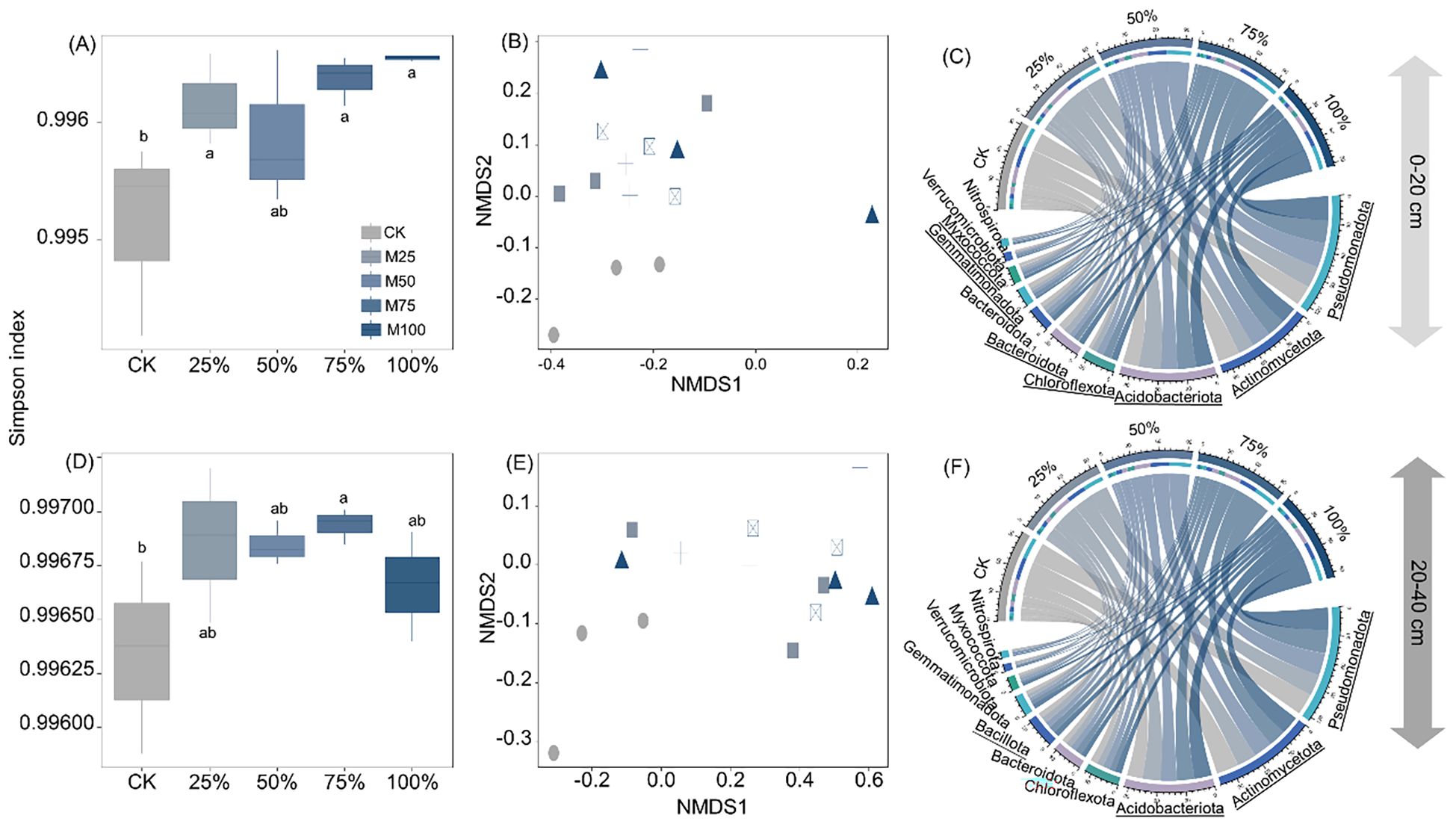
Figure 4. Effects of substituting chemical nitrogen with manure on bacterial diversity and community composition at depths of 0–20 (A–C) and 20–40 cm (D–F). CK, 0% manure N substituted; 25%, 25% manure N substituted; 50%, 50% manure N substituted; 75%, 75% manure N substituted; 100%, 100% manure N substituted. Different letters and underlined notations denote significant differences among treatments at 0.05 level.
In the 0–20-cm depth, manure substitution increased the relative abundance of Actinomycetota (14%–28%), Acidobacteriota (12%–35%), Gemmatimonadota (64%–88%), and Methylomirabilota (55%–101%) while decreasing Bacteroidota (21%–57%), Chloroflexota (15%–24%), and Pseudomonadota (13%–29%) (Figure 4C, p < 0.05). In the 20–40-cm depth, manure substitution increased the relative abundance of Pseudomonadota (5–6%), Acidobacteriota (11%–18%), and Bacillota (12%–21%), but decreased Actinomycetota (12%–20%) compared to CK (Figure 4F, p < 0.05).
3.4 Drivers of maize yield and soil quality index
Maize yield correlated with MBN, SW, ALP, LAP, and vector length at both depths of 0–20 and 20–40 cm, as well as with AP and DON in the 0–20-cm depth and with MBC, BG, and XYL in the 20–40-cm depth (Figures 5A, C; p < 0.05). Soil water content, ALP, and vector length were identified as the main factors influencing maize yield (Figures 5B, D; p < 0.05). These key factors are also linked to the relative abundance of Pseudomonadota, Actinomycetota, Gemmatimonadota, and Methylomirabilota (Figure 6; p < 0.05).
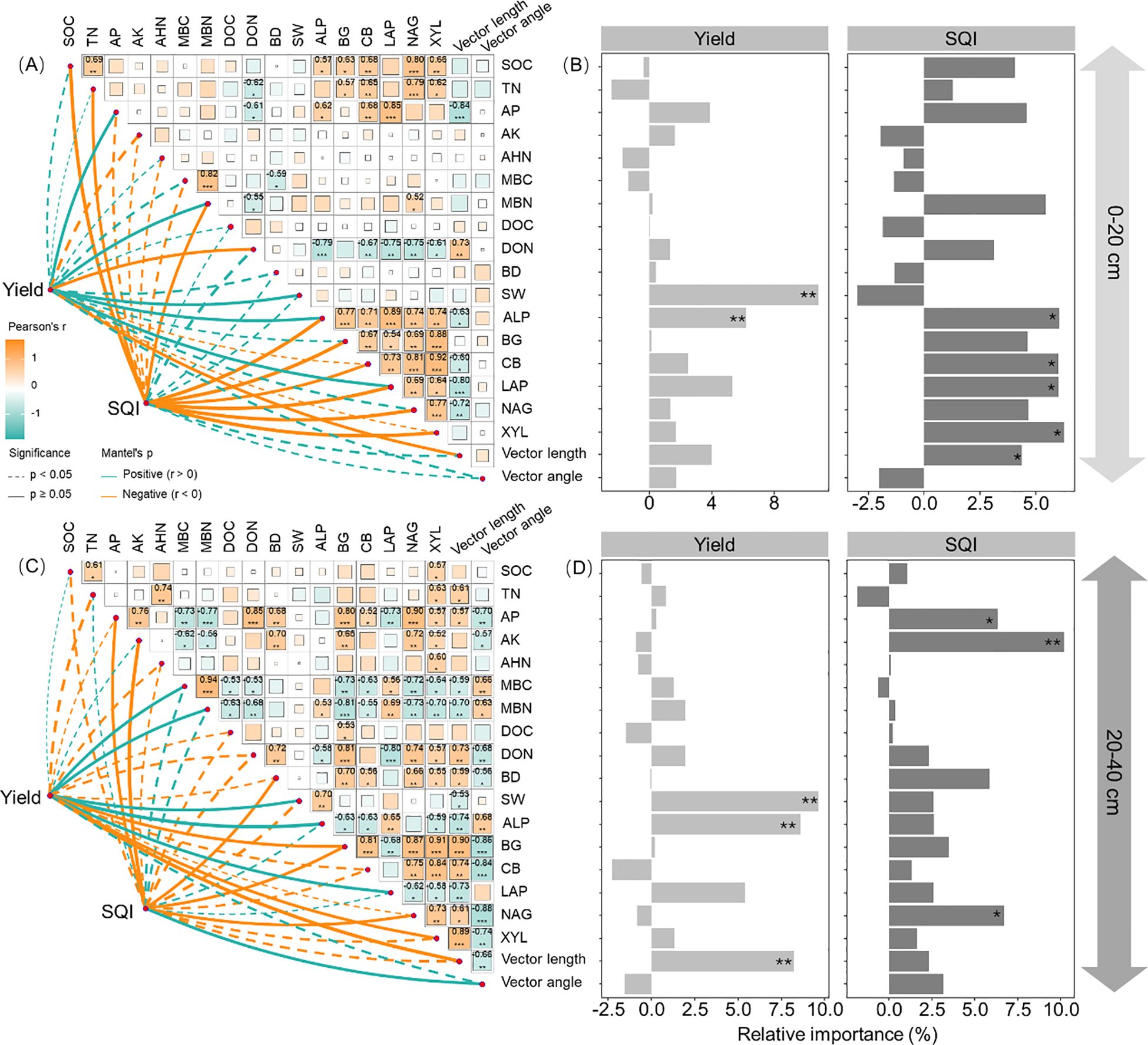
Figure 5. Correlations between soil physicochemical properties, enzyme activities, vector length, and angle affecting maize yield and soil quality index atdepths of 0–20 (A, B) and 20–40 cm (C, D). SOC, soil organic C; TN, total nitrogen; AP, available phosphorus; AK, available potassium; AHN, alkaline hydrolyzedN; MBC, microbial biomass C; MBN, microbial biomass N; DOC, dissolved organic C; DON, dissolved organic N; BD, bulk density; SW, soil watercontent; BG, b-1,4-glucosidase; XYL, b-D-xylopyranoside; CB, b-D-cellobiosidase; NAG, b-1,4-N-acetylglucosaminidase; LAP, L-leucineaminopeptidase; ALP, acid phosphatase. ***p < 0.001; **p < 0.01; *p < 0.05.
Soil quality index was correlated with SOC, TN, MBN, ALP, BG, CB, LAP, NAG, and XYL in the 0–20-cm depth and with AP, AK, BD, BG, NAG, and vector angle in the 20–40-cm depth (Figures 5A, C; p < 0.05). SQI was primarily affected by ALP, CB, LAP, XYL, and vector length in the 0–20-cm depth, while AP, AK, and NAG were more influential in the 20–40-cm depth (Figures 5B,D; p < 0.05). These key factors were also associated with the relative abundance of Actinomycetota, Acidobacteriota, Chloroflexota, and Methylomirabilota in the 0–20-cm depth and Actinomycetota in the 20–40-cm depth (Figure 6; p < 0.05).
4 Discussion
4.1 Effects of organic manure substitution with chemical N on maize yield
Contrary to our hypothesis, organic manure substitution negatively affected maize yield, with greater reductions observed at higher substitution rates (Table 2). These results were consistent with the results of Niu et al. (2024), who found that 75% substitution and above markedly dropped the annual yield of winter wheat and summer maize. This may be due to the excessive proportion of manure substitution causing insufficient N supply, as its slow-release nature requires time to mineralize (Figure 1; Wang et al., 2020). The lower DON in the 0–20-cm and 20–40-cm layers and its positive correlation with maize yield could support this point (Figures 1, 5A). Inhibited maize root growth under a high manure substitution rate served as another crucial factor impairing yield formation (Table 2; Wang et al., 2023). In this study, 50%–100% substitution rates significantly decreased root biomass, length, surface area, and volume (Table 2). This will undoubtedly restrict the nutrient uptake capacity of maize, consequently compromising its development and reducing yield (Feng et al., 2024). Interestingly, we observed that the 25% substitution rate could maintain the maize root growth and yield formation compared with the CK treatment. This can be attributed to the effective synchronization of nitrogen supply and crop demand: the 75% inorganic fertilizer readily satisfied the early peak demand, while the subsequent slow mineralization of the 25% organic manure complemented nitrogen availability during the later reproductive growth stages, thus ensuring stable yield (Table 2, Wang et al., 2020). Collectively, substitution chemical fertilizers with organic manure failed to enhance maize yield in northeast China, but a 25% substitution rate could serve as a sustainable fertilization strategy to reduce chemical fertilizer without a yield penalty.
4.2 Effects of organic manure substitution with chemical N on soil quality
Consistent with our hypothesis, the substitution of chemical fertilizer with organic manure significantly affected soil quality, but exerted distinct effects between depths of 0–20 and 20–40 cm. For the 0–20-cm depth, manure substitution increased soil quality by effectively improving soil chemical properties (e.g., SOC, TN, MBC, MBN, AP, and soil water content). The results may be because 1) the application of organic manure drives the soil carbon cycle by facilitating the transformation of organic matter, thereby increasing soil quality (Feng et al., 2024). 2) Manure substitution treatments promote nutrient release from organic manure and reduce nutrient losses (Cui et al., 2020). For example, organic manure can reduce ammonia volatilization and nitrate leaching through adsorption and, thereby, indirectly increase nutrient levels in the upper layer and contribute to soil quality improvement (Wang et al., 2024b). 3) The application of organic manure can accelerate soil C and N cycling via microbial-mediated priming effect, which may increase soil nutrient availability and improve soil quality (Zhou et al., 2025). These results can be further supported by the high content of microbial biomass C/N and its related enzyme activities (Figures 1, 2). It should be noted that the manure substitution treatments supplied substantial inputs of AP and AK relative to CK, which was also a significant contributor to topsoil quality.
However, the soil quality in the 20–40-cm depth under manure substitution treatments exhibited an opposite trend, being significantly reduced compared to the CK. This can be attributed to three main reasons: first, since both organic and chemical fertilizers were primarily incorporated into the topsoil through rotary tillage, manure substitution treatments had limited influence on the deeper soil layers (He et al., 2022). Second, organic manure input reduced the leaching of available nitrogen through physical adsorption (Wei et al., 2021). This can also be supported by the lower contents of TN, DON, and AHN in the 20–40-cm depth (Figure 1B). Third, high rates of manure-substituted chemical fertilizer inhibited maize root growth due to insufficient N supply. This likely led to diminished root residue incorporation into deeper soil layers, consequently reducing nutrient content and being detrimental to soil quality (Feng et al., 2024). Interestingly, we did not detect a significant association between the maize yield and soil quality. This is because a significant relationship between crop yield and soil quality in previous studies often emerges over longer time scales (He et al., 2022; Nie et al., 2025). In the short term, yield is predominantly influenced by immediate nutrient availability, while the benefits of soil quality improvements take longer to manifest and affect crop yield (Wang et al., 2020).
4.3 Soil microorganism response to organic manure substitution with chemical N
Soil bacteria serve as critical mediators in soil nutrient cycling, and their diversity and community structure are linked to the soil quality and crop performance (Philippot et al., 2024). Our study found that manure substitution treatments significantly increased the Simpson index of soil bacteria at depths of 0–20 and 20–40 cm (Figure 4). In line with Han et al. (2021), the organic manure substitution with chemical N fertilizer enhanced soil bacterial α-diversity. This increase in bacterial diversity may be attributed to two reasons: first, the organic manure application supplied a broader spectrum of substrates that support the diverse bacterial community growth (Lin et al., 2019). Second, organic manure enables the direct introduction of its inherent microbial community into the soil, thereby contributing to an increase in bacterial diversity (Li et al., 2022). Interestingly, although the majority of organic fertilizer was concentrated in the 0–20-cm depth, bacterial diversity also increased significantly in the deeper layer. This phenomenon can be primarily attributed to the leaching of soluble organic compounds derived from organic fertilizer decomposition, which may activate a more dormant microbial community in the deeper layer (Chen et al., 2024). In addition, the vertical transport of bacteria from the upper layer may have contributed substantially to the enhanced bacterial diversity observed in the 20–40-cm depth (He et al., 2023). This process was likely facilitated by the increased soil water content resulting from organic manure application, which improved hydrological connectivity and enhanced bacterial mobility through the soil profile.
Manure substitution shifted the soil microbial community structure and boosted the prevalence of beneficial taxa (Figure 4). For example, our study observed that manure substitution treatments significantly enriched the relative abundance of Actinomycetota, Acidobacteriota, Gemmatimonadota, and Methylomirabilota compared to CK (Figure 4). These findings are consistent with the previous literature on the effects of organic manure application on soil microbial communities (Ji et al., 2018; Lu et al., 2023). Acidobacteriota play a pivotal role in nutrient cycling by secreting enzymes that hydrolyze recalcitrant organic matter into bioavailable forms (Feng et al., 2024). This point was evidenced by the significant correlation between Acidobacteriota and soil C/N-acq enzyme activities (Figure 6). Actinobacteria are recognized for their significant contribution to soil carbon cycling, primarily due to their pronounced ability to decompose recalcitrant polymeric compounds (Nie et al., 2025). The strong correlation between Actinobacteria and soil C- and N-related enzyme activities further evidenced this point. Collectively, the enrichment of key soil microorganisms driven by organic manure substitution accelerated soil nutrient turnover, thereby enhancing soil quality.
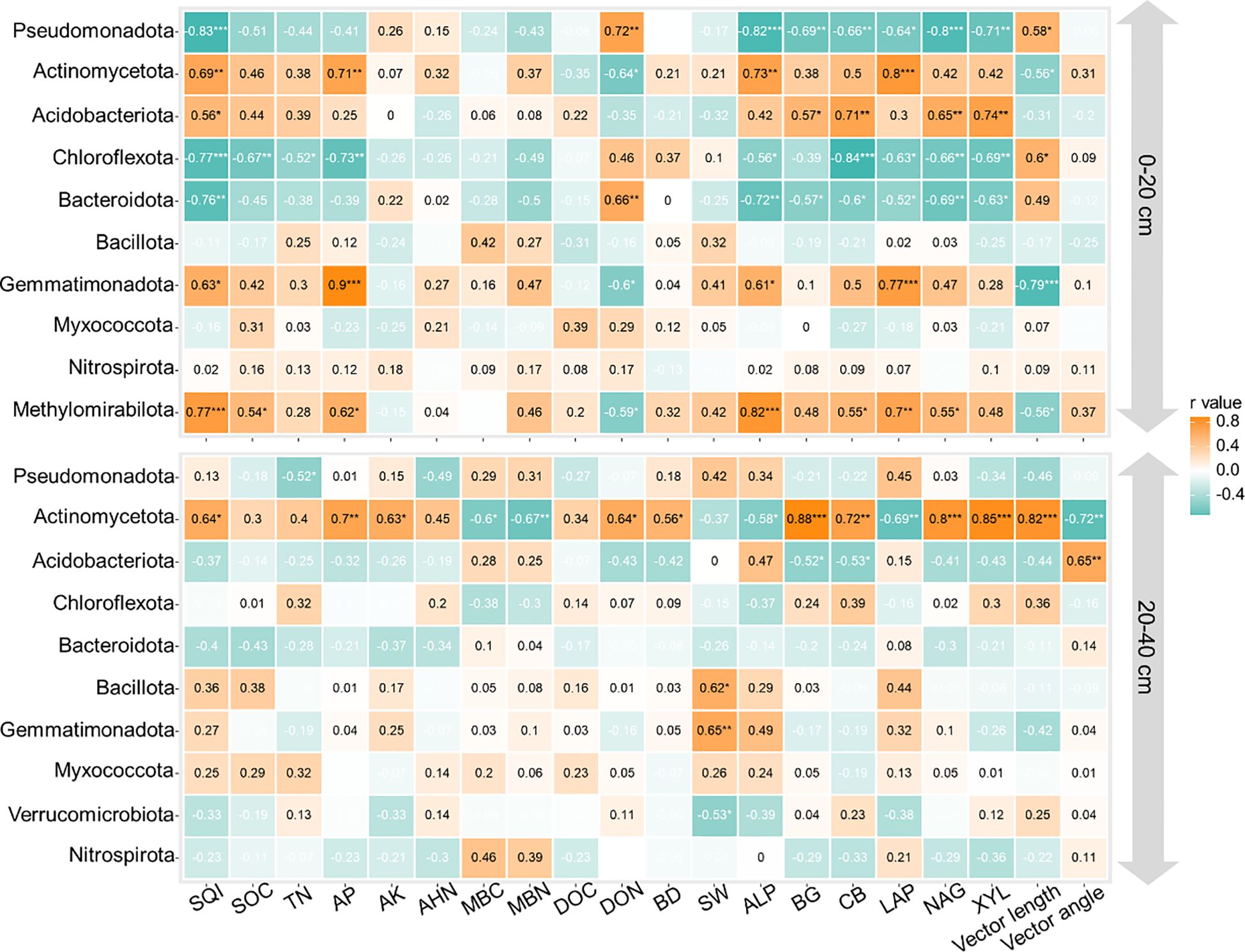
Figure 6. Correlations between the top 10 abundance of bacterial phyla and soil physicochemical properties, enzyme activities, vector length, and angle at depths of 0–20 and 20–40 cm. SQI, soil quality index; SOC, soil organic C; TN, total nitrogen; AP, available phosphorus; AK, available potassium; AHN, alkaline hydrolyzed N; MBC, microbial biomass C; MBN, microbial biomass N; DOC, dissolved organic C; DON, dissolved organic N; BD, bulk density; SW, soil water content; BG, β-1,4-glucosidase; XYL, β-d-xylopyranoside; CB, β-d-cellobiosidase; NAG, β-1,4-N-acetylglucosaminidase; LAP, l-leucine aminopeptidase; ALP, acid phosphatase. ***p < 0.001; ** p < 0.01; *p < 0.05.
5 Conclusion
In summary, this study demonstrates that short-term organic manure substitution generally decreased maize yield, which can be largely attributed to suppressed root growth caused by limited nitrogen availability. Only 25% substitution rates maintained maize yield compared to 100% chemical fertilizer applied. In contrast, organic manure application significantly enhanced soil quality within the 0–20-cm layer. Random forest analysis identified the key factors affecting soil quality, including alkaline phosphatase, cellobiohydrolase, leucine aminopeptidase, xylosidase, and enzyme vector length. The increased abundance of Actinomycetota, Acidobacteriota, and Methylomirabilota, along with their positive correlations with C- and N-acquiring enzyme activities, suggests that organic manure promotes soil organic carbon accumulation and nutrient availability through the enrichment of beneficial microbial taxa. However, a decline in soil quality was observed in the 20–40-cm layer, likely associated with reduced available phosphorus, available potassium, and N-acetylglucosaminidase activity. Collectively, these findings suggest that 25% organic manure substitution can serve as a sustainable fertilization strategy in Northeast China, capable of maintaining crop productivity while enhancing topsoil quality.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
CZ: Conceptualization, Data curation, Investigation, Writing – original draft. JW: Formal analysis, Investigation, Methodology, Software, Writing – original draft, Writing – review & editing. TX: Data curation, Methodology, Conceptualization, Validation, Writing – original draft. KP: Data curation, Investigation, Methodology, Software, Supervision, Writing – original draft. BM: Investigation, Data curation, Methodology, Software, Writing – original draft. QL: Software, Investigation, Writing – original draft. YL: Investigation, Methodology, Software, Conceptualization, Formal analysis, Funding acquisition, Project administration, Resources, Validation, Visualization, Writing – original draft. XD: Investigation, Software, Validation, Writing – original draft. YH: Funding acquisition, Investigation, Methodology, Project administration, Supervision, Validation, Visualization, Writing – review & editing, Writing – original draft. JQW: Funding acquisition, Software, Supervision, Writing – review & editing, Writing – original draft.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was financially supported by the Heilongjiang Provincial Scientific Research Operating Funds for Provincial Research Institutes (Grant No. CZKYF2025-1-C016) and the earmarked fund for China Agriculture Research System (CARS-02-46).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1694608/full#supplementary-material
References
Ali, A., Liu, X., Yang, W., Li, W., Chen, J., Qiao, Y., et al. (2024). Impact of bio-organic fertilizer incorporation on soil nutrients, enzymatic activity, and microbial community in wheat–maize rotation system. Agronomy 14, 1942. doi: 10.3390/agronomy14091942
Bao, S. (2000). Soil and agricultural chemistry analysis (Beijing, 178-200: China Agriculture Press China Agriculture Press).
Chen, L., Zhou, S., Zhang, Q., Zou, M., Yin, Q., Qiu, Y., et al. (2024). Effect of organic material addition on active soil organic carbon and microbial diversity: A meta-analysis. Soil Tillage Res. 241, 106128. doi: 10.1016/j.still.2024.106128
Cui, N., Cai, M., Zhang, X., Abdelhafez, A. A., Zhou, L., Sun, H., et al. (2020). Runoff loss of nitrogen and phosphorus from a rice paddy field in the east of China: Effects of long-term chemical N fertilizer and organic manure applications. Global Ecol. Conserv. 22, e01011. doi: 10.1016/j.gecco.2020.e01011
Cui, X., Zhang, Y., Gao, J., Peng, F., and Gao, P. (2018). Long-term combined application of manure and chemical fertilizer sustained higher nutrient status and rhizospheric bacterial diversity in reddish paddy soil of Central South China. Sci. Rep. 8, 16554. doi: 10.1038/s41598-018-34685-0
De Corato, U., Viola, E., Keswani, C., and Minkina, T. (2024). Impact of the sustainable agricultural practices for governing soil health from the perspective of a rising agri-based circular bioeconomy. Appl. Soil Ecol. 194, 105199. doi: 10.1016/j.apsoil.2023.105199
Feng, W., Ai, J., Sánchez-Rodríguez, A. R., Li, S., Zhang, W., Yang, H., et al. (2025). Depth-dependent patterns in soil organic C, enzymatic stochiometric ratio, and soil quality under conventional tillage and reduced tillage after 55-years. Agriculture Ecosyst. Environ. 385, 109584. doi: 10.1016/j.agee.2025.109584
Feng, W., Sánchez-Rodríguez, A. R., Bilyera, N., Wang, J., Wang, X., Han, Y., et al. (2024). Mechanisms of biochar-based organic fertilizers enhancing maize yield on a Chinese Chernozem: Root traits, soil quality and soil microorganisms. Environ. Technol. Innovation 36, 103756. doi: 10.1016/j.eti.2024.103756
Guo, R., Gong, W., Qi, S., Xu, J., Shang, Z., and Joseph, S. (2025). Biochar-based urea enhances nitrogen use efficiency and mitigates nitrogen leaching in greenhouse vegetable production. Environ. Technol. Innovation 38, 104104. doi: 10.1016/j.eti.2025.104104
He, H., Peng, M., Lu, W., Hou, Z., and Li, J. (2022). Commercial organic fertilizer substitution increases wheat yield by improving soil quality. Sci. Total Environ. 851, 158132. doi: 10.1016/j.scitotenv.2022.158132
He, L., Sun, X., Li, S., Zhou, W., Chen, Z., and Bai, X. (2023). The vertical distribution and control factor of microbial biomass and bacterial community at macroecological scales. Sci. Total Environ. 869, 161754. doi: 10.1016/j.scitotenv.2023.161754
Ji, L., Wu, Z., You, Z., Yi, X., Ni, K., Guo, S., et al. (2018). Effects of organic substitution for synthetic N fertilizer on soil bacterial diversity and community composition: A 10-year field trial in a tea plantation. Agriculture Ecosyst. Environ. 268, 124–132. doi: 10.1016/j.agee.2018.09.008
Jia, R., Zhou, J., Chu, J., Shahbaz, M., Yang, Y., Jones, D. L., et al. (2022). Insights into the associations between soil quality and ecosystem multifunctionality driven by fertilization management: A case study from the North China Plain. J. Cleaner Production 362, 132265. doi: 10.1016/j.jclepro.2022.132265
Jiang, Y., Zhang, R., Zhang, C., Su, J., Cong, W.-F., and Deng, X. (2022). Long-term organic fertilizer additions elevate soil extracellular enzyme activities and tobacco quality in a tobacco-maize rotation. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.973639
Ju, J., Gu, Q., Zhou, H., Zhang, H., Mao, W., Yang, H., et al. (2022). Effects of organic fertilizer combined with chemical fertilizer on nutrients, enzyme activities, and rice yield in reclaimed soil. Commun. Soil Sci. Plant Anal. 53, 3060–3071. doi: 10.1080/00103624.2022.2101661
Li, H., Lehmann, A., Rongstock, R., Xu, Y., Kunze, E., Meidl, P., et al. (2025). Diversity of organic amendments increases soil functions and plant growth. PLANTS PEOPLE PLANET 7, 80–86. doi: 10.1002/ppp3.10588
Li, X., Li, B., Chen, L., Liang, J., Huang, R., Tang, X., et al. (2022). Partial substitution of chemical fertilizer with organic fertilizer over seven years increases yields and restores soil bacterial community diversity in wheat–rice rotation. Eur. J. Agron. 133, 126445. doi: 10.1016/j.eja.2021.126445
Li, G., Zhu, S., Long, J., Mao, H., Dong, Y., and Hou, Y. (2023). Differences in microbial community structure and metabolic activity among tea plantation soils under different management strategies. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1219491
Lin, Y., Ye, G., Kuzyakov, Y., Liu, D., Fan, J., and Ding, W. (2019). Long-term manure application increases soil organic matter and aggregation, and alters microbial community structure and keystone taxa. Soil Biol. Biochem. 134, 187–196. doi: 10.1016/j.soilbio.2019.03.030
Liu, L., Gao, Y., Yang, W., Liu, J., and Wang, Z. (2024). Community metagenomics reveals the processes of nutrient cycling regulated by microbial functions in soils with P fertilizer input. Plant Soil 499, 139–154. doi: 10.1007/s11104-023-05875-1
Lu, Z., Zhou, Y., Li, Y., Li, C., Lu, M., Sun, X., et al. (2023). Effects of partial substitution of chemical fertilizer with organic manure on the activity of enzyme and soil bacterial communities in the mountain red soil. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1234904
Ma, X., Wang, J., Zhang, K., Yang, Z., Gao, Y., Wu, A., et al. (2025). Enhanced multifunctionality of sandy loam soil with co-application of biochar and organic manure is driven by microbial network complexity rather than community diversity. Agriculture Ecosyst. Environ. 393, 109812. doi: 10.1016/j.agee.2025.109812
Marx, M.-C., Wood, M., and Jarvis, S. C. (2001). A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol. Biochem. 33, 1633–1640. doi: 10.1016/S0038-0717(01)00079-7
Nie, J., Xie, Q., Zhou, Y., He, F., Yousaf, M., Zhu, B., et al. (2025). Long-term legume green manure residue incorporation is more beneficial to improving bacterial richness, soil quality and rice yield than mowing under double-rice cropping system in Dongting Lake Plain, China. Front. Plant Sci. 16. doi: 10.3389/fpls.2025.1603434
Niu, J., Saeed, Q., Wang, W., Zhang, R., Liu, L., Lv, F., et al. (2024). Manure replacing synthetic fertilizer improves crop yield sustainability and reduces carbon footprint under winter wheat–summer maize cropping system. J. Environ. Manage. 358, 120936. doi: 10.1016/j.jenvman.2024.120936
Philippot, L., Chenu, C., Kappler, A., Rillig, M. C., and Fierer, N. (2024). The interplay between microbial communities and soil properties. Nat. Rev. Microbiol. 22, 226–239. doi: 10.1038/s41579-023-00980-5
Salimani, S., Verma, R., Kumar, K. B., and Mehta, S. (2025). Impact of biofertilizers and integrated nutrient management on transplanted rice for improving the growth, yield and soil quality: A review. Int. J. Plant Soil Sci. 37, 271–281. doi: 10.9734/ijpss/2025/v37i55451
Sihi, D., Dari, B., Sharma, D. K., Pathak, H., Nain, L., and Sharma, O. P. (2017). Evaluation of soil health in organic vs. conventional farming of basmati rice in North India. J. Plant Nutr. Soil Sci. 180, 389–406. doi: 10.1002/jpln.201700128
Tian, S., Zhu, B., Yin, R., Wang, M., Jiang, Y., Zhang, C., et al. (2022). Organic fertilization promotes crop productivity through changes in soil aggregation. Soil Biol. Biochem. 165, 108533. doi: 10.1016/j.soilbio.2021.108533
Trasar-Cepeda, C., Leiros, M., and Gil-Sotres, F. (2008). Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol. Biochem. 40, 2146–2155. doi: 10.1016/jsoilbio.2008.03.015
Vance, E. D., Brookes, P. C., and Jenkinson, D. S. (1987). An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 19, 703–707. doi: 10.1016/0038-0717(87)90052-6
Wang, X., Liu, M., Ciampitti, I. A., Cui, J., Fang, K., Zhao, S., et al. (2024b). Benefits and trade-offs of replacing inorganic fertilizer by organic substrate in crop production: A global meta-analysis. Sci. Total Environ. 925, 171781. doi: 10.1016/j.scitotenv.2024.171781
Wang, J., Liu, Z., Shan, X., and Zhang, Y. (2023). Partial replacement of chemical fertilizer by manure affects maize root traits in North China Plain. Soil Use Manage. 39, 1545–1556. doi: 10.1111/sum.12933
Wang, C., Ma, Y., He, W., Kuzyakov, Y., Bol, R., Chen, H., et al. (2024a). Soil quality and ecosystem multifunctionality after 13-year of organic and nitrogen fertilization. Sci. Total Environ. 931, 172789. doi: 10.1016/j.scitotenv.2024.172789
Wang, H., Yang, S., Wang, Y., Gu, Z., Xiong, S., Huang, X., et al. (2022). Rates and causes of black soil erosion in Northeast China. CATENA 214, 106250. doi: 10.1016/j.catena.2022.106250
Wang, X., Yang, Y., Zhao, J., Nie, J., Zang, H., Zeng, Z., et al. (2020). Yield benefits from replacing chemical fertilizers with manure under water deficient conditions of the winter wheat – summer maize system in the North China Plain. Eur. J. Agron. 119, 126118. doi: 10.1016/j.eja.2020.126118
Wei, Z., Hoffland, E., Zhuang, M., Hellegers, P., and Cui, Z. (2021). Organic inputs to reduce nitrogen export via leaching and runoff: A global meta-analysis. Environ. pollut. 291, 118176. doi: 10.1016/j.envpol.2021.118176
Wu, Z., Chen, X., Lu, X., Zhu, Y., Han, X., Yan, J., et al. (2025). Impact of combined organic amendments and chemical fertilizers on soil microbial limitations, soil quality, and soybean yield. Plant Soil 507, 317–334. doi: 10.1007/s11104-024-06733-4
Yang, S., Xiao, J., Liang, T., and Tan, H. (2023). Response of bacterial compositions to the use of slow-release fertilizers with long-acting agents and synergists. Appl. Soil Ecol. 182, 104699. doi: 10.1016/j.apsoil.2022.104699
Yin, Y., Guo, S., Xu, Q., Liu, J., Wang, H., Zhuang, Y., et al. (2025). Co-benefits for cropland yield, nitrogen emissions, and climate impact through multi-objective optimization agricultural manure solutions. Nat. Commun. 16, 6415. doi: 10.1038/s41467-025-61885-w
Zhai, L., Wang, Z., Zhai, Y., Zhang, L., Zheng, M., Yao, H., et al. (2022). Partial substitution of chemical fertilizer by organic fertilizer benefits grain yield, water use efficiency, and economic return of summer maize. Soil Tillage Res. 217, 105287. doi: 10.1016/j.still.2021.105287
Zhang, Q., Chu, Y., Yin, Y., Ying, H., Zhang, F., and Cui, Z. (2023). Comprehensive assessment of the utilization of manure in China’s croplands based on national farmer survey data. Sci. Data 10, 223. doi: 10.1038/s41597-023-02154-7
Zhou, S., Chang, T., Zhang, Y., Shaghaleh, H., Zhang, J., Yang, X., et al. (2024). Organic fertilizer compost alters the microbial composition and network structure in strongly acidic soil. Appl. Soil Ecol. 195, 105263. doi: 10.1016/j.apsoil.2023.105263
Zhou, X., Feng, Z., Yao, Y., Liu, R., Shao, J., Jia, S., et al. (2025). Nitrogen input alleviates the priming effects of biochar addition on soil organic carbon decomposition. Soil Biol. Biochem. 202, 109689. doi: 10.1016/j.soilbio.2024.109689
Zhou, Z., Zhang, S., Jiang, N., Xiu, W., Zhao, J., and Yang, D. (2022). Effects of organic fertilizer incorporation practices on crops yield, soil quality, and soil fauna feeding activity in the wheat-maize rotation system. Front. Environ. Sci. 10. doi: 10.3389/fenvs.2022.1058071
Keywords: organic fertilizer application, maize yield, root traits, soil enzyme activity, soil bacterial community
Citation: Zhou C, Wang J, Xu T, Pei K, Ma B, Li Q, Liu Y, Ding X, Han Y and Wang J (2025) Manure substitution for chemical nitrogen enhances soil quality without compromising maize yield: a short-term field experiment in Northeast China. Front. Plant Sci. 16:1694608. doi: 10.3389/fpls.2025.1694608
Received: 28 August 2025; Accepted: 17 October 2025;
Published: 17 November 2025.
Edited by:
Jie Zhou, Nanjing Agricultural University, ChinaReviewed by:
Yang Lei, Anhui Academy of Agricultural Sciences AAAS, ChinaQiqi Gao, Institute of Agricultural Resources and Regional Planning (CAAS), China
Copyright © 2025 Zhou, Wang, Xu, Pei, Ma, Li, Liu, Ding, Han and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yehui Han, aGFueWVodWkxOTgyQDE2My5jb20=; Junqiang Wang, QXVndXN0LXdqcUAxNjMuY29t
†These authors have contributed equally to this work