 Longhai Zhang1,2,3
Longhai Zhang1,2,3 Zhiyong Zhang1,2,3
Zhiyong Zhang1,2,3 Zihao Li1,2,3
Zihao Li1,2,3 Xinjun Chen4Shirui Pu1,2,3Qian Chen1Min Gong1
Xinjun Chen4Shirui Pu1,2,3Qian Chen1Min Gong1 Muhammad Anas Khan1,2,3
Muhammad Anas Khan1,2,3 Jinxing Zhou1,2,3*
Jinxing Zhou1,2,3*- 1Jianshui Research Station, School of Soil and Water Conservation, Beijing Forestry University, Beijing, China
- 2State Key Laboratory of Efficient Production of Forestry Resources, Beijing Forestry University, Beijing, China
- 3Engineering Research Center of Forestry Ecological Engineering, Ministry of Education, Beijing Forestry University, Beijing, China
- 4State Key Laboratory of Tree Genetics and Breeding, Institute of Ecological Protection and Restoration, Chinese Academy of Forestry, Beijing, China
Karst ecosystems, recognized as ecologically fragile systems, are characterized by vegetation-soil interaction mechanisms particularly vulnerable to wildfire disturbances. Understanding the post-fire coupling dynamics between vegetation and soil is crucial for guiding restoration in these vulnerable landscapes. This study investigated post-fire areas across five disturbance intensities (unburned, light, moderate, severe, extreme) in Jianshui County, Yunnan Province, China. We conducted a systematic analysis of soil physicochemical properties and herb diversity, and quantified the vegetation-soil coupling relationship using grey relational modeling. Key results reveal: (1) 21 herbaceous species were documented, with Asteraceae, Poaceae, and Fabaceae collectively constituting 76.2% of the flora. (2) Across the fire severity gradient, herbaceous diversity demonstrated an initial increase followed by a subsequent decline. (3) Grey correlation analysis identified soil pH, total potassium, and phosphatase activity as primary drivers of herb community variation. (4) Vegetation-soil coupling coordination followed a U-shaped trajectory, achieving optimal synergy (0.84, Higher coordination) under extreme-severity burns and minimal coordination (0.71, Medium coordination) in severe burns. These findings underscore that moderate fire regimes can play a positive role in enhancing the vegetation-soil coupling effect. Furthermore, the strategic regulation of soil pH and potassium availability during restoration emerges as a critical lever for optimizing ecosystem recovery and enhancing resilience. This study provides valuable insights for developing targeted post-fire management strategies in karst regions.
1 Introduction
Karst regions are recognized as one of the world’s most representative fragile ecosystems. Their unique geological structure and hydrological conditions—such as shallow soil layers and fractured bedrock—underlie a pronounced ecological vulnerability and complex recovery dynamics (Wang et al., 2004; Chen et al., 2021). Characterized by shallow soil layers, high bedrock exposure rates, and vegetation degradation, these areas form distinctive vegetation-soil-rock structures (Li et al., 2009; Zhang and Huisingh, 2018; Jiang et al., 2024), rendering them highly susceptible to ecological degradation under natural or anthropogenic disturbances (D’Ettorre et al., 2024; Jing et al., 2025). Among these disturbances, wildfire stands out as a particularly acute and transformative agent. In recent years, superimposed impacts of climate change and intensified human activities have maintained elevated forest fire frequencies in these regions, posing persistent ecological risks (Tian et al., 2014; Zhang et al., 2021). As a natural disturbance agent, wildfires rapidly alter biogeochemical cycles of carbon, nitrogen, and phosphorus, abruptly restructuring vegetation communities and soil properties (Keeley, 2009; Alcañiz et al., 2016; González-De Vega et al., 2018; Fernández-García et al., 2019), thereby disrupting pre-existing vegetation-soil equilibria (MacDougall et al., 2013; Toberman et al., 2014). Such perturbations are amplified in fragile karst ecosystems, where calcareous substrates and fissured bedrock may exacerbate post-fire ecological consequences (Keeley, 2009; Bradstock et al., 2010; Kardol et al., 2023; Luo et al., 2024), underscoring the critical need to investigate post-fire recovery mechanisms in these regions.
The soil and vegetation form a dynamic, interconnected system governed by intrinsic coupling mechanisms. Soil provides the essential material foundation for vegetation, directly shaping plant community structure and functionality (Lange et al., 2015; Wu et al., 2018; Hou et al., 2021). Plants modify soil physicochemical properties through processes such as nutrient absorption, stabilization, accumulation, and decomposition (Coradini et al., 2022; Chen et al., 2024; Da et al., 2024).Wildfire disturbance, mediated through thermal effects on surface soils and vegetation (Reinhart et al., 2016), induces spatiotemporally heterogeneous ecological responses. Flames alter soil organic carbon (SOC), pH, and microbial communities, thereby steering vegetation succession (Qiang et al., 2021; Agbeshie et al., 2022; Li X. et al., 2024), whereas plants adapt via nutrient resorption, serotinous cone strategies, and other regenerative mechanisms (Vallejo et al., 2012). These interconnected processes collectively regulate post-fire soil quality rehabilitation and vegetation community assembly (Moya et al., 2018a; Quigley et al., 2020). Nevertheless, systematic understanding of vegetation-soil interactions in burned karst ecosystems remains limited, particularly regarding feedback mechanisms governing ecological restoration (De Long et al., 2019).
The coupling relationship between soil and vegetation reflects the dynamic equilibrium between material cycling and energy flows within ecosystems. Current research has extensively documented fire impacts on isolated ecosystem components. For instance, Moya et al (Moya et al., 2018b). demonstrated that fire severity acutely affects soil phosphorus concentration, conductivity, and enzyme activity in Iberian burned areas, with gradual stabilization occurring over ≥15-year recovery periods. Li Z. et al. (2024) reported that Tibetan Plateau wildfires initially (<15 years) reduce soil pH while enriching nutrients to drive pioneer species succession, followed by synergistic improvements in enzyme activity and biodiversity enhancing community stability in later stages (>15 years). Mayor et al. (2016) identified positive correlations between vegetation cover and soil enzyme activities in fire-prone shrublands. However, prevailing studies predominantly focus on unidirectional fire effects on either soil or vegetation (Stancic and Repe, 2018; Wang et al., 2022; Cahojová et al., 2024; Rebi et al., 2025), with insufficient attention to their coupling dynamics. This knowledge gap is particularly acute in karst regions, where calcareous soils and fractured bedrock may engender unique post-fire feedback mechanisms.
We investigated understory herbaceous communities and soils across a fire severity gradient (light, moderate, severe, extreme) in Jianshui County, Yunnan Province. By characterizing herb composition and diversity patterns along this gradient and applying grey relational coupling modeling to quantify vegetation-soil interdependencies, this research aims to provide scientific guidance for post-fire ecological restoration in karst ecosystems.
2 Materials and methods
2.1 Overview of the study area
The research area is situated in a Pinus massoniana plantation (102°55′E, 23°40′N) at Yanbasi Village, Jianshui County, Yunnan Province, China (Figure 1). Key climatic parameters include an annual precipitation of 685 mm, mean annual air temperature of 19.8 °C, mean soil temperature of 20.8 °C, relative humidity of 72%, 2,322 annual sunshine hours, and a 307-day frost-free period. The region exhibits extensive karst topography dominated by limestone bedrock, with bedrock exposure rates ranging from 30% to 70%. The plantation comprises a tree layer dominated by Pinus massoniana, a shrub layer primarily consisting of Dodonaea viscosa and Indigofera tinctoria, and an herbaceous layer mainly including Panicum virgatum, Carex lanceolata, Eleusine indica, Bidens pilosa, Euphorbia esula, Cymbopogon citratus, and Arthraxon prionodes. The soil type of the studied site is calcareous soil (Calcaric Cambisols, more than 15% calcium carbonate (CaCO3) and are commonly found in arid, semi-arid, humid, and semi-humid regions.), developed from a limestone base and characterized by a red color (Pang et al., 2018).
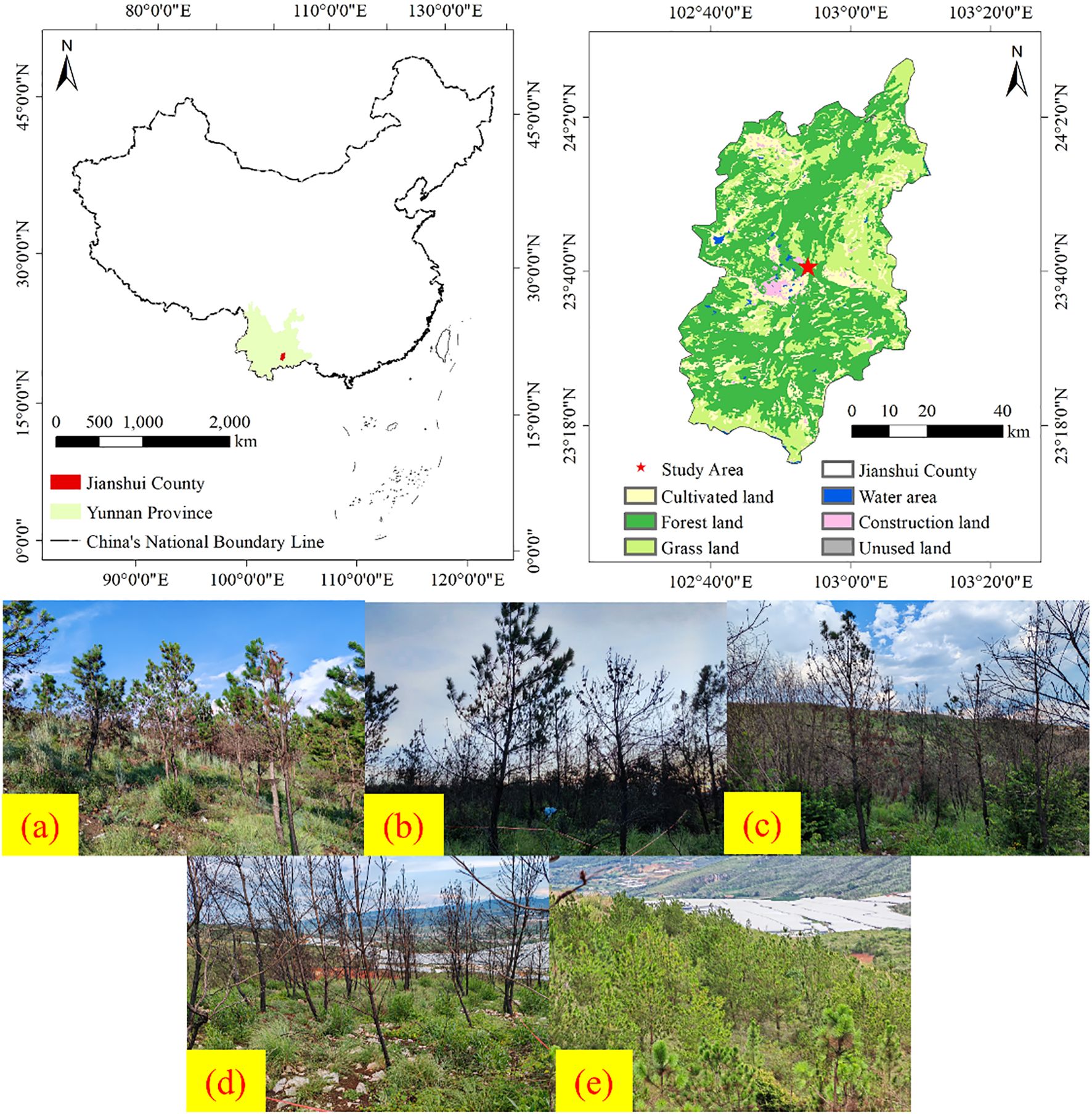
Figure 1. Research area overview. In Figure 1, (a) Light-severity fire, (b) Moderate-severity fire, (c) High-severity fire, (d) Extreme-severity fire, (e) Unburned control.
2.2 Plot setting and sample collection
A wildfire occurred in the study area in April 2024, with data collection and field surveys conducted in July 2024 following the event. Based on vegetation change parameters and fire characteristics (Xie et al., 2005; Eidenshink et al., 2007; Keeley, 2009), a total of five experimental plots were established, each representing one of the following fire severity categories: light burn (tree mortality ≤30%), moderate burn (30% < tree mortality ≤60%), severe burn (60% < tree mortality ≤90%), and complete burn (100% tree mortality), using adjacent unburned Pinus massoniana forest as control (Figure 1 and Table 1). To ensure pre-fire homogeneity, all plots shared comparable elevation, topography, soil type (calcareous soil), dominant vegetation composition, and vegetation coverage (Table 2). Within each of the five main plots, three 10 m × 10 m subplots were randomly established as spatial replicates for sampling the understory herbaceous community. Five 1 m × 1 m herbaceous quadrats were systematically positioned in each subplot using a five-point sampling method. Vegetation parameters, including species composition, abundance, coverage, and biomass, were recorded within the quadrats. After removing surface ash, triplicate soil samples were collected diagonally from each plot in both control and burned areas at a depth of 0–15 cm (Wang et al., 2022). The collected soils were stored in sealed bags as air-dried samples (for chemical analysis) and fresh samples (preserved at −4 °C) for subsequent laboratory assays. Detailed plot characteristics are provided in Table 1.
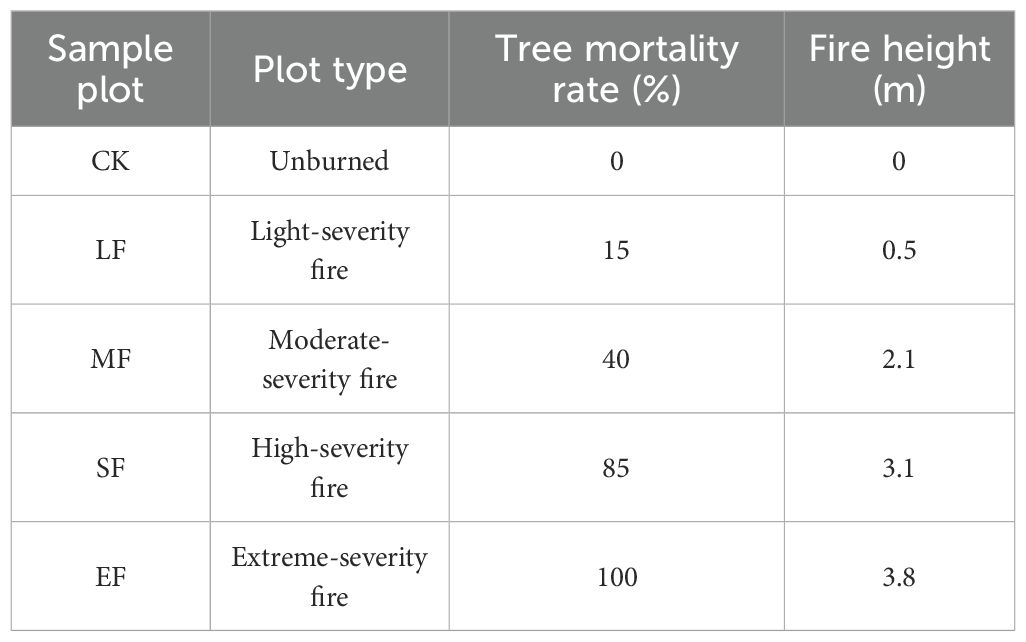
Table 1. Characteristics of the burned area.
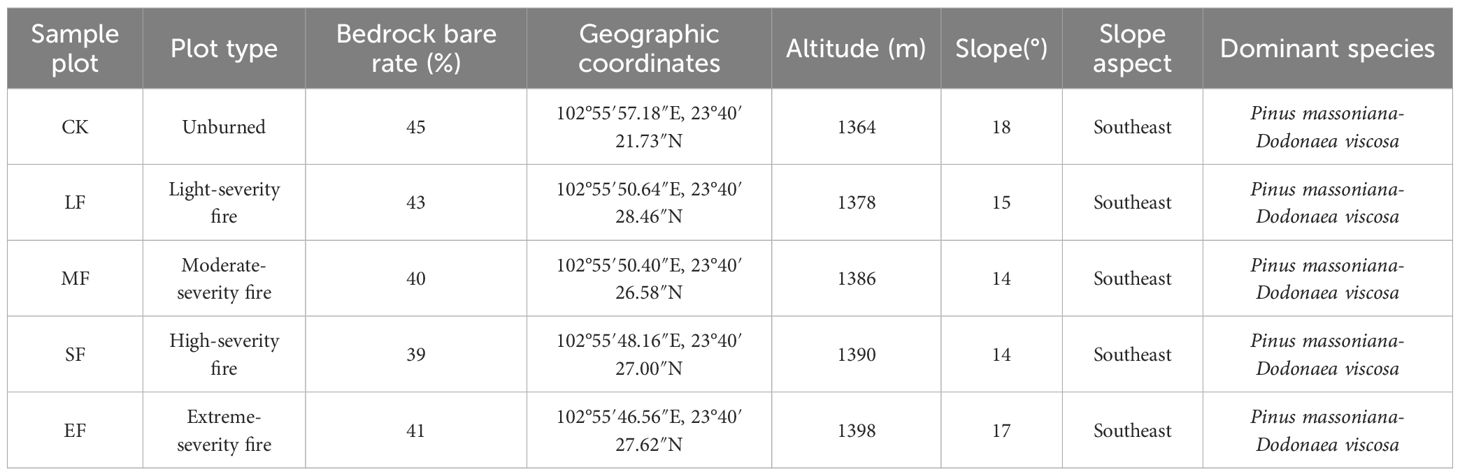
Table 2. Basic information of the sample plot.
2.3 Soil analysis
Soil water content (SWC) was determined by the drying method. Bulk density (BD) was determined using the core method (State Forestry Administration, 1999a). Soil pH was measured from a 1: 2.5 soil-to-water ratio using a pH meter (PB-10 pH meter) and electrical conductivity (5:1 water-soil ratio) using a conductivity meter(CT-3030). SOC was measured by the potassium dichromate external heating method (State Forestry Administration, 1999b). Available nitrogen (AN) was assessed via alkaline hydrolysis-diffusion (State Forestry Administration, 1999c), available phosphorus (AP) by molybdenum-antimony colorimetry (Olsen, 1954), total nitrogen (TN) by sulfuric acid digestion (Liu et al., 2013), total phosphorus (TP) by molybdenum-blue method (Sciences, C.A.O.A, 1988), and total potassium (TK) by sodium hydroxide fusion (State Forestry Administration, 1999d). Microbial biomass carbon (MBC) and nitrogen (MBN) were quantified using chloroform fumigation. Enzyme activities were determined via microplate fluorescence assays (Guan, 1986).
2.4 Vegetation diversity metrics
Herbaceous diversity was assessed using Equations 1–4
Shannon-Wiener index (H’)
Simpson index (D)
Margalef index (D’)
Pielou index (E)
where S is the number of species; Pi represents the relative abundance of the i species, calculated as Pi=Ni/N, with N denoting the total number of individuals of all species within the quadrat, and Ni indicating the number of individuals of the i species.
2.5 Grey relational coupling modeling
Owing to the interlaced complexity of vegetation-soil coupling dynamics and the inherent interdependencies between these systems, this study adopts the Grey Relational Analysis (GRA) (Equations 4–12). This approach facilitates a quantitative assessment of the coupling relationships and coordination levels between vegetation and soil across varying fire severity levels. Compared to traditional statistical methods (e.g., regression analysis, ANOVA, principal component analysis), the GRA method is applicable regardless of sample size or distribution patterns. Moreover, it requires minimal computational effort and does not suffer from inconsistencies between quantitative outcomes and qualitative interpretations, thereby addressing the limitations inherent in conventional statistical approaches for systemic analysis. We integrated four diversity indices with 17 soil variables through grey relational analysis. In this model, the composite vegetation system, represented by the four diversity indices, was defined as the parent sequence (reference sequence). The soil system, characterized by the 17 physicochemical and biological variables, was treated as the child sequences (comparison sequences). Data were normalized to eliminate unit effects prior to calculating relational coefficients (Liu et al., 2005). Utilizing the study by Gao et al. (2022), the coupling degree (C) was categorized into the following 7 levels (Table 3): 0 ≤ C < 0.4 (Serious incoordination), 0.4 ≤ C < 0.5 (Medium incoordination), 0.5 ≤ C < 0.6 (Light incoordination), 0.6 ≤ C < 0.7 (Light coordination), 0.7 ≤ C < 0.8 (Medium coordination), 0.8 ≤ C < 0.9 (Higher coordination), and 0.9 ≤ C ≤ 1.0 (Superior coordination).

Table 3. System of ecosystem coupling coordination.
1.Data standardization:
2. Incidence coefficient:
represents the grey relational coefficient of to , and is the resolution coefficient, the general value is ϵ= 0.5.
3. Calculate the Grey Relational Grade:
When 0 < ≤ 0.35, the correlation degree is weak; when 0.35 < ≤ 0.65, the correlation degree is medium; when 0.65 < ≤ 0.85, the correlation degree is strong; when 0.85 < ≤ 1.0, the correlation degree is significantly strong.
4. Coupling analysis:
m is the number of soil indicators, and l is the quantity of diversity indicators.
2.6 Data analysis
Species diversity indices, grey relational grade, and system coupling degree were calculated using Microsoft Excel 2019 (version 2019). Geospatial distribution maps of sampling areas were generated with ArcGIS 10.3. Statistical analyses, including one-way analysis of variance (ANOVA) with LSD post hoc test, were performed using IBM SPSS Statistics 27. Data visualization and graphical representations were conducted using Origin 2024.
3 Results
3.1 Variations in soil properties and understory herb composition with fire severity
Post-fire soil pH in the study area transitioned from alkaline to acidic(pH 7.24-6.31,Table 4). SWC and SOC increased with fire severity, while BD decreased. Soil electrical conductivity (EC), TN, TP, TK, AN, and AP showed an initial decline followed by subsequent increases. Soil enzyme activities displayed differential responses across fire intensity gradients: β-1,4-glucosidase activity significantly decreased (p < 0.05) but increased under extreme burning conditions. Cellobiohydrolase, leucine aminopeptidase, β-1,4-N-acetylglucosaminidase, and phosphatase activities showed significant variation(P<0.05) without distinct directional trends.
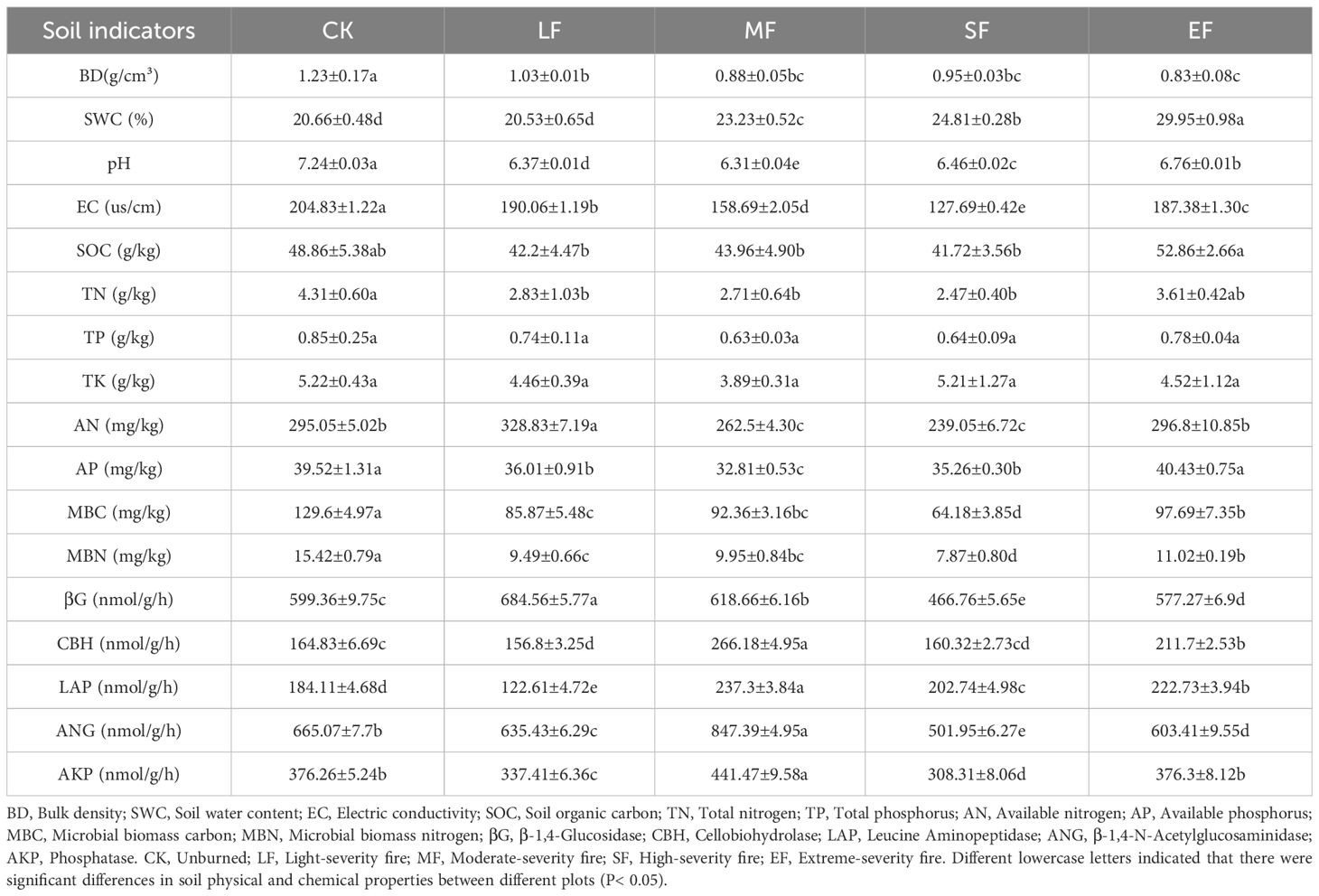
Table 4. Soil physical and chemical properties.
A total of 21 herbaceous plant species were recorded in the study area (Table 5), predominantly belonging to Asteraceae (33.3%), Poaceae (33.3%), Fabaceae (9.6%), Cyperaceae (4.8%), Euphorbiaceae (4.8%), Asparagaceae (4.8%), Orchidaceae (4.8%), and Liliaceae (4.8%). Compared to the unburned control areas, the plant community in fire-disturbed regions transitioned to pioneer species characterized by high adaptability and rapid growth, including typical taxa such as Eleusine indica (Poaceae), Cymbopogon citratus (Poaceae), Arthraxon prionodes (Poaceae), and Lespedeza cuneata (Fabaceae).
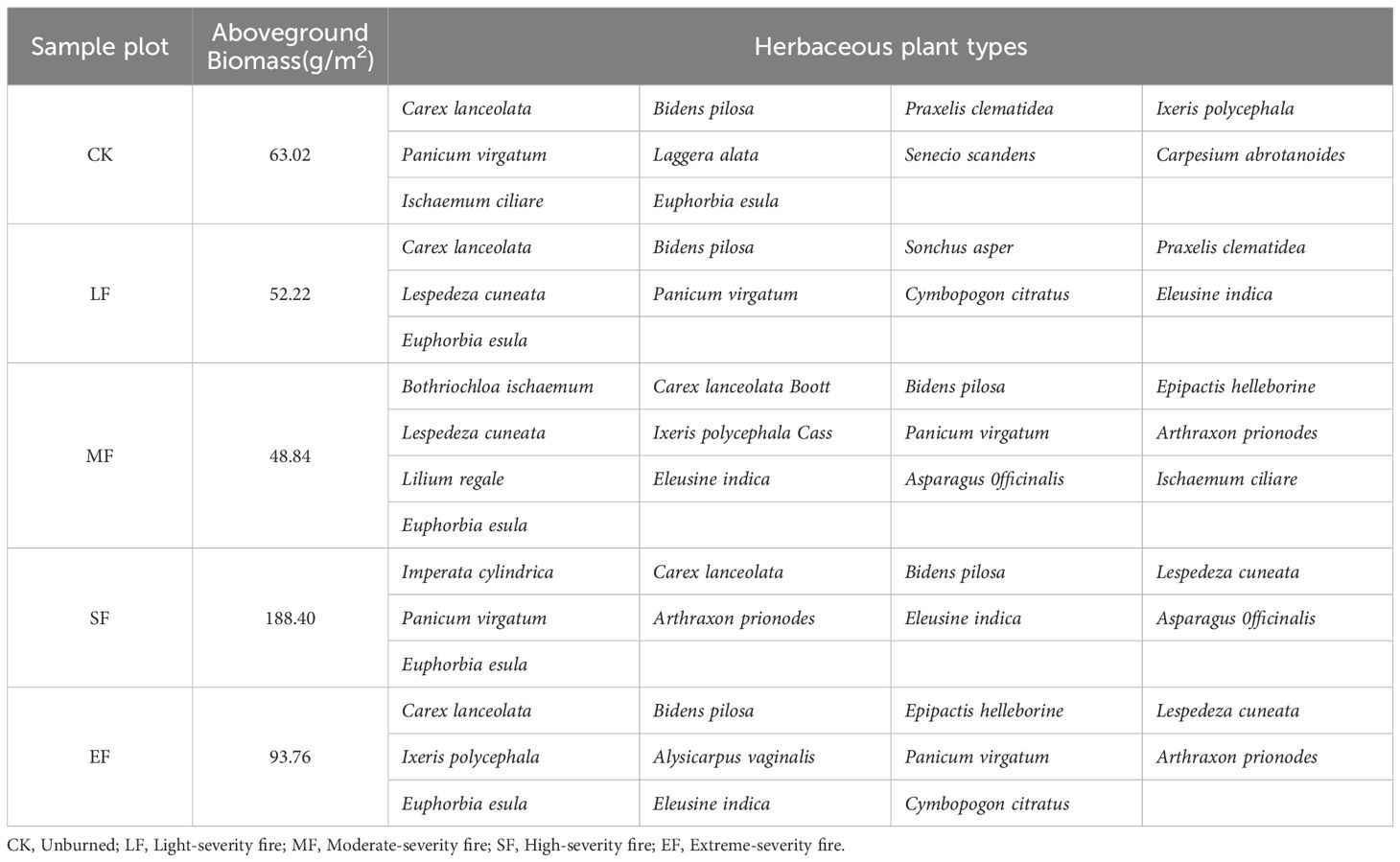
Table 5. Overview of herbaceous plant communities with different degrees of fire in the study area.
3.2 Diversity characteristics of herbaceous communities
With increasing fire intensity, the Shannon-Wiener diversity index (H’) and Margalef’s richness index (D’) of herbaceous communities exhibited a unimodal response (Figure 2). The Shannon-Wiener index peaked under high-severity fire conditions, while Margalef’s richness index reached its maximum under moderate-severity fire. Pielou’s evenness index (E) did not vary significantly across fire severity levels(ANOVA, P>0.05), with the lowest value (0.74) observed in unburned control plots. Simpson’s dominance index (D) displayed the most pronounced fire-induced changes, demonstrating an initial decrease followed by an increase with escalating fire severity, though all burned plots maintained lower values (D < 0.42) compared to unburned controls.
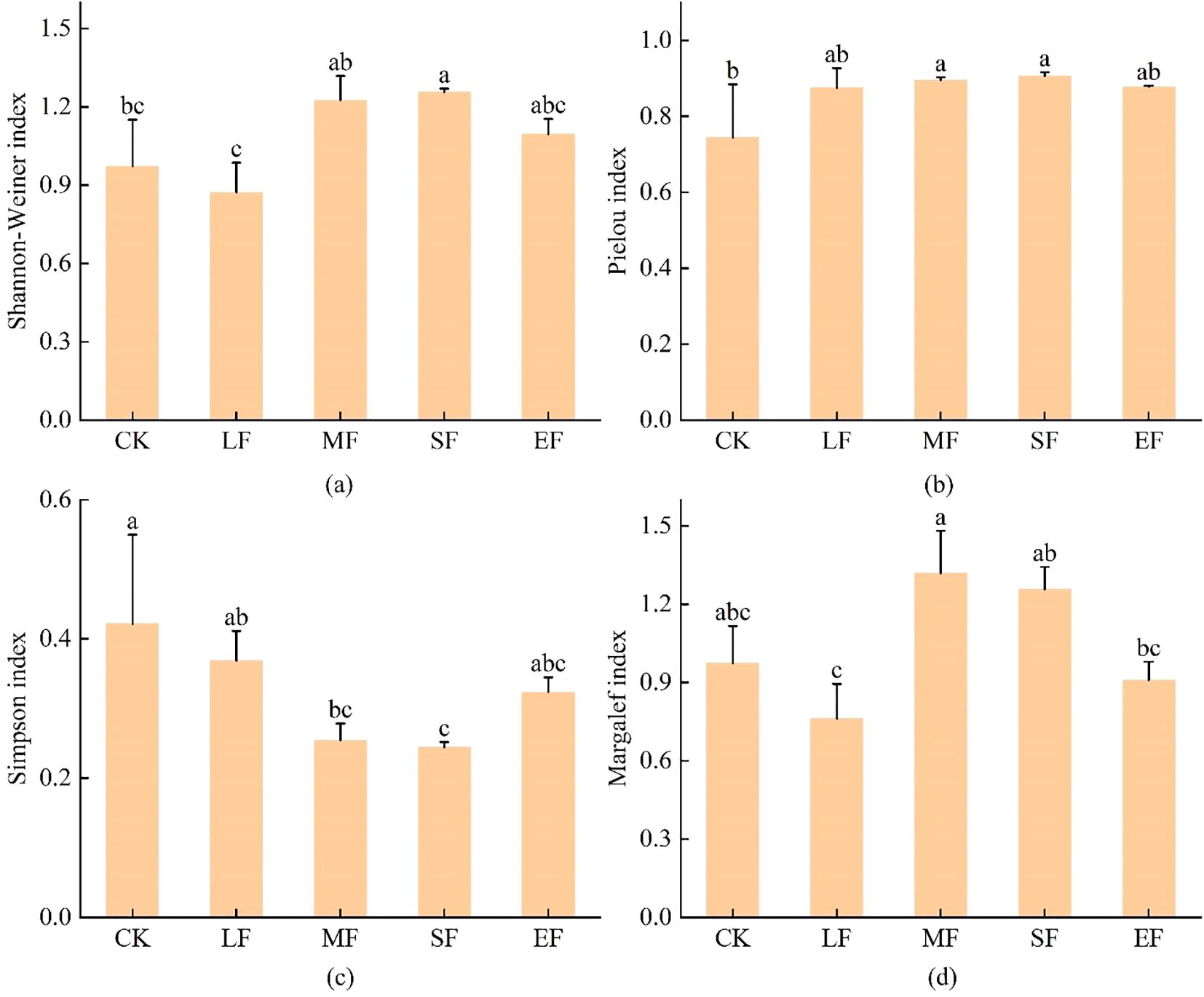
Figure 2. Diversity Indices of Herbaceous Communities. CK, Unburned; LF, Light-severity fire; MF, Moderate-severity fire; SF, High-severity fire; EF, Extreme-severity fire. Different lowercase letters in the figure indicate significant differences between groups (P< 0.05). (a) Shannon-Wiener index. (b) Pielou index. (c) Simpson index. (d) Margalef index.
3.3 Grey relational analysis of vegetation-soil system factors
Grey relational analysis indicated strong overall coupling between the plant and soil systems, with relational coefficients ranging from 0.61 to 0.87 (mean = 0.75) (Figure 3). Pielou’s evenness index (E) exhibited the strongest overall correlation with the soil factor suite (mean Γ = 0.79). Its dynamics were most closely linked to soil pH and soil water content. Shannon-Wiener index (H’) was primarily driven by extracellular enzyme activities, showing the highest correlations with phosphatase and leucine aminopeptidase. Simpson’s index (D) was most strongly associated with soil electrical conductivity, but had the weakest correlation with leucine aminopeptidase activity. Margalef’s richness index (D’) had the weakest overall coupling (mean Γ = 0.69), with leucine aminopeptidase activity being its most influential soil factor.
3.4 Coupling coordination analysis of the vegetation-soil system
As shown in Table 6, the coupling degree of the vegetation-soil system across fire severity gradients in the study area ranged from 0.71 to 0.84, exhibiting a U-shaped trajectory (initially decreasing then increasing) with escalating fire intensity. The maximum coupling degree (0.84, Higher coordination) was observed in extreme-severity burned plots, while the minimum value (0.71, Medium coordination) occurred in severe burns.
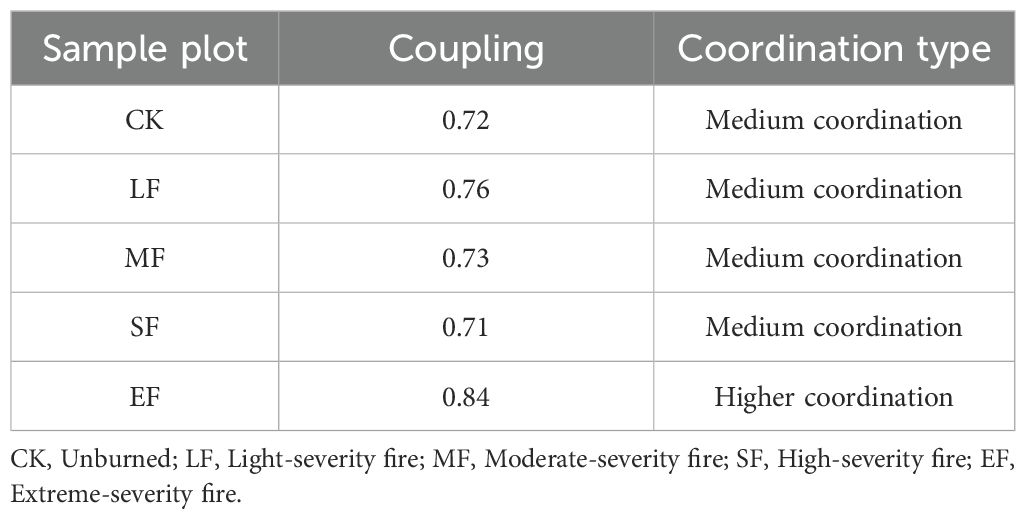
Table 6. The coupling and coordination degree between the diversity of herbaceous communities and soil systems.
4 Discussion
4.1 Effects of wildfire on soil properties
Our results demonstrate that forest fires exert a significant influence on soil pH. Several studies have documented marked post-fire pH elevation, which is primarily attributed to the release of alkaline elements from residual ash, the deacidifying effects of base cations on surface soil, and elevated soluble salt levels (SMITH, 1970; Xu et al., 2012; Bridges et al., 2019; Li et al., 2020). Contrary to the typical fire-induced increase in soil pH (Bárcenas-Moreno et al., 2022; Oliva et al., 2025), our study observed de-creased pH levels in the karst region. In calcareous soils (e.g., SW China karst), this acidification intensifies because wind and water erosion rapidly leach base cation-rich ash (K+, Ca²+, Mg²+), transporting soluble ions via runoff or percolation (Raison, 1979; Neary et al., 1999). Furthermore, fire-induced vegetation loss and soil structure degradation intensified erosion (Hamman et al., 2007), accelerating the leaching of base cations. This led to an increased relative proportion of H+ and Al³+ ions in the soil cation exchange capacity (CEC) (Neary et al., 1999; Salgado et al., 2024), driving the soil towards acidification.
Numerous studies confirm that fire disturbance alters soil physicochemical properties. Our investigation found that fires (low, moderate, and high severity) reduced SOC content, closely aligning with previous research (Li et al., 2020; Pellegrini et al., 2021; Pacaldo et al., 2025). Furthermore, fire reduced soil nitrogen (N) content. The decline in both organic carbon and total nitrogen is attributed to the combustion of surface soil organic matter, resulting in losses primarily via CO₂, nitrogenous gases, and particulate matter in smoke (Verma et al., 2019). Fire reduced total phosphorus (P) content, though not significantly—a finding attributable to phosphorus’s high volatilization threshold, resulting in non-significant differences (Neary et al., 2005).Generally, forest fires significantly impact TK. Some studies observed an initial increase in soil K immediately post-fire (Kennard and Gholz, 2001). However, high temperatures can volatilize total potassium, or its solubility, combined with leaching in specific karst regions with high drainage, may subsequently reduce its content (Verma and Jayakumar, 2012).
Fire not only modifies soil physicochemical properties but also profoundly influences soil enzyme activity. This study revealed that enzyme activities under moderate-severity fire were consistently higher than in control plots. While fire-induced high temperatures can directly denature enzymes or indirectly alter activity by modifying soil environmental conditions (Pei et al., 2023), moderate-severity fire enhanced plant diversity. This strengthened microbial community functionality, and increased root exudates provided energy for microbes, collectively improving the soil chemical environment and thereby regulating enzyme activity (López-Poma and Bautista, 2014).
It is important to note that our soil sampling strategy integrated the entire 0–15 cm depth layer. While this approach provides a standardized measure of the root zone environment relevant to herbaceous communities, it may also integrate contrasting signals from different horizons. Future studies employing horizon-specific sampling could yield more granular insights into the vertical redistribution of elements and post-fire biogeochemical cycling within the soil profile.
4.2 Analysis of herbaceous communities and diversity
The naturally regenerated herbaceous communities in post-fire areas of the study region were predominantly composed of species from Asteraceae, Poaceae, and Fabaceae (76.2% of total species). These taxa, characterized by broad ecological niches, drought tolerance, and adaptability to barren soils, exhibited a competitive advantage in early successional stages following fire (Gao et al., 2022; Geng et al., 2022). For instance, Holmes et al. (2000) documented a dramatic increase in Poaceae relative coverage following fire disturbance. Similarly, Kazanis and Arianoutsou (1996) observed absolute dominance of Fabaceae species in post-fire Pinus nigra forests in Greece, with Poaceae and Asteraceae serving as key indicator families. Georgiadis and Georgiadis (2002) further confirmed Asteraceae and Fabaceae as dominant annual herbs during initial post-fire recovery. These findings collectively demonstrate the cross-regional adaptability of pioneer species within these botanical families.
As a natural ecological factor in ecosystems, wildfire plays a significant role in maintaining biodiversity (He et al., 2019). Among nutrient limitations for vegetation growth, nitrogen (N) constitutes a key controlling element (2020). Karst ecosystems exhibit unique nitrogen cycling patterns: despite their calcareous soils demonstrating high inorganic N supply capacity and rapid N turnover (Garousi et al., 2021), post-fire reductions in total nitrogen content and N turnover rates inhibit vegetation recovery (Wang et al., 2022). This response contrasts sharply with certain non-karst ecosystems—for instance, Mediterranean pine forests show increased soil organic matter and nitrogen content after fire (Knicker et al., 2005), while fire disturbances in North American grasslands positively enhance nitrogen and phosphorus cycling (Reinhart et al., 2016). The distinctive post-disturbance response in karst regions primarily stems from their fragile geological context: shallow soil layers and fractured bedrock structures exacerbate post-fire soil-water loss, leading to nutrient depletion (D’Ettorre et al., 2024). This stressed environment favors drought-tolerant, oligotrophic species such as Asteraceae and Poaceae, which dominate herbaceous communities (66.6%). Notably, these taxa generally show lower proportions in post-fire successional communities of non-karst ecosystems (Abedi et al., 2023; Blinkova et al., 2024; Wieczorkowski et al., 2024).
Species diversity, a fundamental attribute of biological communities, underpins ecosystem stability (Nie et al., 2020). Following fire disturbance in the study area, both the Shannon-Wiener index (reflecting the combined effect of species richness and evenness) and the Margalef index (representing species richness) exhibited a unimodal pattern in response to fire intensity, indicating that species diversity peaked at moderate to high fire severity. This aligns with the Intermediate Disturbance Hypothesis (Connell, 1978), which posits that a certain level of disturbance promotes the development of species diversity. Specifically, the Margalef richness index reached its maximum under moderate fire severity, signifying the highest number of species present. Conversely, the Shannon-Wiener diversity index peaked under high fire severity, suggesting a species composition characterized by both relatively high richness and evenness at this level. This divergence may be linked to soil nitrogen dynamics, as excessive nitrogen availability can suppress species richness, particularly in the herb layer (Gilliam, 2006; McClean et al., 2011). The Pielou evenness index showed no significant differences across fire severity levels, indicating stable distribution uniformity of individuals within the community, a finding consistent with studies by Mahood and Balch (2019) and Neri et al. (2023). In contrast, the Simpson index, representing dominance, displayed an initial decrease followed by an increase, and was consistently lower in burned plots compared to the control. This suggests that fire suppressed the original dominant species and facilitated the establishment of new species. This finding provides complementary insights to the unchanged Pielou evenness, collectively indicating that fire primarily influences community structure by altering dominant species rather than overall evenness (Zhang et al., 2012; Wang et al., 2024).
4.3 Coupling analysis of the vegetation-soil system
Vegetation-soil interactions exhibit dynamic feedback mechanisms during ecosystem recovery. Vegetation growth and distribution significantly influence soil properties, while edaphic alterations reciprocally drive species-specific vegetation responses, ultimately shaping plant community structure and spatial patterns. Grey relational analysis identified soil pH, TK, and phosphatase activity as pivotal regulators of herbaceous diversity (Figure 3). Soil pH serves as a master variable influencing plant growth (Neina, 2019), while TK, as an essential element, is directly involved in key physiological processes such as osmoregulation, stomatal movement, and enzyme activation (Zhou et al., 2025). Phosphatase activity directly regulates the mineralization of organic phosphorus, determining plant access to phosphorus—a commonly limiting nutrient. During post-fire ecological recovery, soil pH modulates vegetation distribution by affecting root growth, metabolism, and enzyme activity. For instance, Zhang et al. (2012) noted that most understory plant roots grow optimally in slightly acidic environments (pH 5.5–6.5). A close functional relationship exists between phosphatase activity and soil pH, as confirmed by Acosta-Martinez and Tabatabai (2000). Post-fire changes in total potassium reveal the driving role of plants in nutrient cycling: although fire can cause potassium volatilization losses (Johnson et al., 2005), plants accelerate potassium cycle recovery by returning potassium to the soil through biomass turnover and litter decomposition.
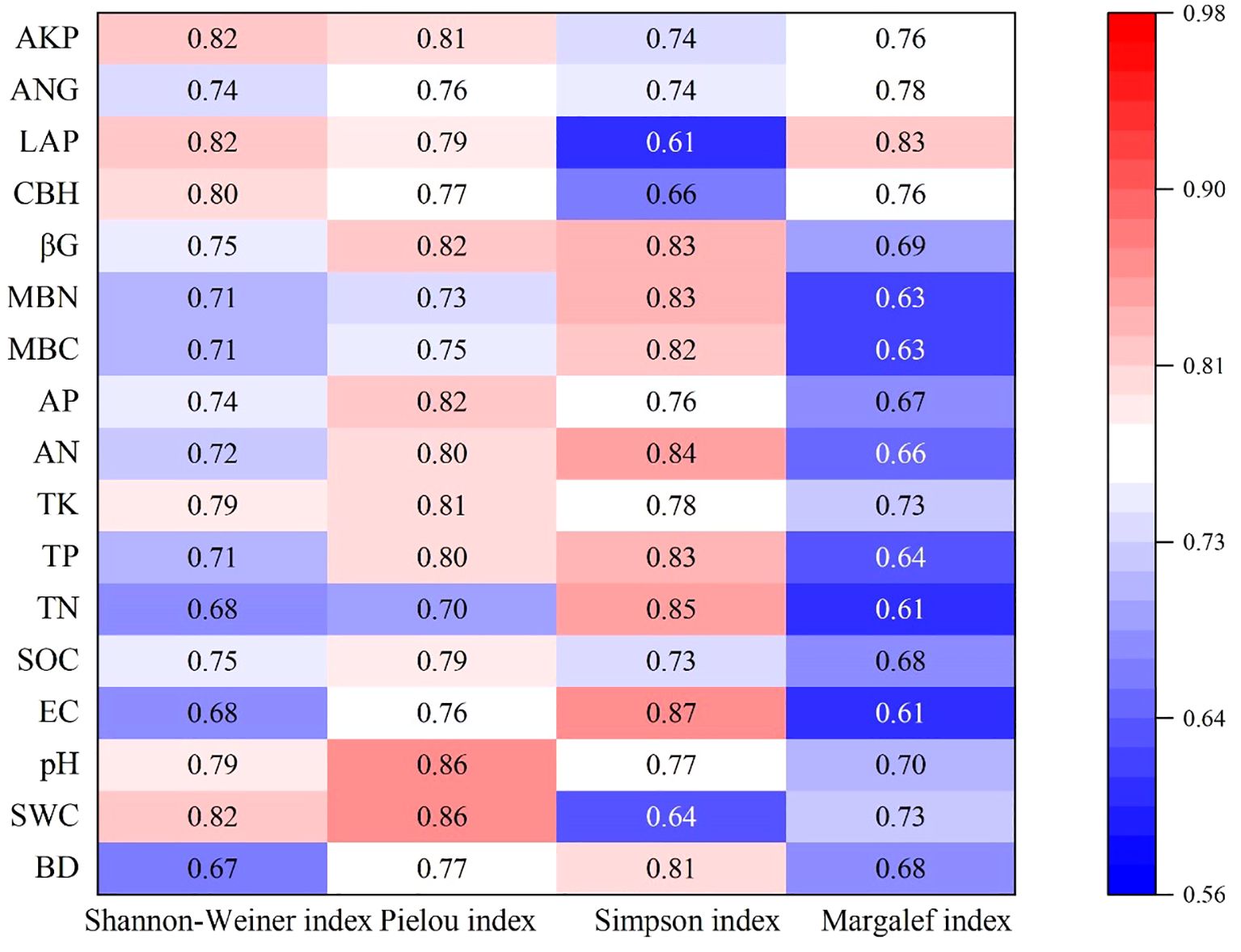
Figure 3. Coupled heat map of herbaceous community diversity and soil. BD, Bulk density; SWC, Soil water content; EC, Electric conductivity; SOC, Soil organic carbon; TN, Total nitrogen; TP, Total phosphorus; AN, Available nitrogen; AP, Available phosphorus; MBC, Microbial biomass carbon; MBN, Microbial biomass nitrogen; βG, β-1,4-Glucosidase; CBH, Cellobiohydrolase; LAP, Leucine Aminopeptidase; ANG, β-1,4-N-Acetylglucosaminidase; AKP, Phosphatase.
Herbaceous communities and soil systems in post-fire karst ecosystems of Southwest China exhibited limited coupling coordination variation, generally maintaining coordinated developmental patterns. Consistent with the Intermediate Disturbance Hypothesis (Waldrop and Brose, 1999), Disturbance of a certain intensity can promote the renewal of forest ecosystems, improve the undergrowth environment of the region and increase species diversity. However, intensified interspecific competition for spatial resources, soil moisture, and nutrients subsequently reduced vegetation-soil coupling. Additional factors including species elimination and mortality (Gao et al., 2022; Lin et al., 2022) further diminished system coordination. Critically, the highest level of coordination (0.84) was observed under extreme-severity burn conditions. This unsustainable transient state reflects a drastically simplified ecosystem condition immediately following complete vegetation removal, where the abrupt elimination of biological complexity and competition temporarily creates a state of low resistance to soil-driven influences. However, this short-lived synergy is a direct consequence of ecosystem degradation and is unlikely to persist as succession proceeds. In contrast, unburned herbaceous communities exhibited prolonged natural succession with intense competition, resulting in persistently low coupling degree. These findings suggest that optimal post-fire restoration in karst regions requires strategic vegetation density control and selection of stress-adapted species to balance competitive interactions. The temporary window of high coordination following extreme fires should be regarded as a critical, brief opportunity to initiate restoration through the introduction of pioneer species, before more competitive interactions become reestablished.
5 Conclusions
This study investigated short-term vegetation-soil coupling relationships in post-fire karst ecosystems of southwestern China. Results demonstrated that fire severity critically influenced herbaceous community composition and soil properties during the early recovery stage (three months post-fire). Pioneer species from Asteraceae, Poaceae, and Fabaceae dominated community regeneration across the fire severity gradient. The vegetation-soil system exhibited a U-shaped coupling coordination response across the fire severity gradient, peaking under extreme burns due to reduced competition, while minimal coordination in severe burns reflected intensified resource competition. Notably, regulating soil pH, potassium availability, and phosphatase activity emerged as critical levers for optimizing restoration, indirectly driving community reassembly through nutrient cycling and enzymatic activity.
This research provides practical guidance for post-fire management in karst regions:(1) Introduce pioneer species during periods of high vegetation-soil coupling following fire disturbance;(2) Implement targeted soil nutrient management, such as pH adjustment and potassium supplementation, to enhance ecosystem resilience. It should be noted that these findings and management implications are constrained by the single post-fire sampling event, which captured only a transient ecosystem state. Future studies should prioritize integrated long-term monitoring to elucidate vegetation-soil dynamics across full recovery trajectories—essential for developing robust, multi-scale ecological rehabilitation strategies.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
LZ: Conceptualization, Investigation, Methodology, Writing – original draft. ZZ: Conceptualization, Writing – review & editing. ZL: Formal Analysis, Writing – review & editing. XC: Investigation, Writing – review & editing. SP: Investigation, Writing – original draft. QC: Investigation, Writing – original draft. MG: Investigation, Writing – original draft. MK: Writing – review & editing. JZ: Project administration, Resources, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the National Key Research and Development Program of China (No: 2022YFF1302900), the National Natural Science Foundation of China (No: 42361144885,No: 42207065) and the 5·5 Engineering Research & Innovation Team Project of Beijing Forestry University (No: BLRC2023B09).
Acknowledgments
The authors gratefully acknowledge the support provided by the College of Soil and Water Conservation at Beijing Forestry University and the Jianshui Research Station. We also extend our thanks to our colleagues in the research group for their valuable assistance. Finally, we wish to express our sincere gratitude to the editors and reviewers for their insightful comments and constructive suggestions, which have greatly improved the quality of this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1709599/full#supplementary-material
References
Abedi, M., Omidipour, R., Hosseini, S. V., Bahalkeh, K., and Gross, N. (2023). Fire disturbance effects on plant taxonomic and functional β-diversity mediated by topographic exposure. Ecol. Evol. 13, e9841. doi: 10.1002/ece3.9841
Acosta-Martinez, V. and Tabatabai, M. A. (2000). Enzyme activities in a limed agricultural soil. Biol. Fertility Soils 31, 85–91. doi: 10.1007/s003740050628
State Forestry Administration (1999b). Determination of organic matter in forest soil and calculation carbon-nitrogen ratio. Forest soil analysis methods, ly/t 1237–1999 (Beijing: China Standard Press).
State Forestry Administration (1999c). Determination of hydrolysable nitrogen in forest soil. Forest soil analysis methods, ly/t 1229–1999 (Beijing: China Standard Press).
State Forestry Administration (1999d). Determination of total potassium in forest soil, ly/t 1234-1999 (Beijing: China Standard Press).
Agbeshie, A. A., Abugre, S., Atta-Darkwa, T., and Awuah, R. (2022). A review of the effects of forest fire on soil properties. J. Forestry Res. 33, 1419–1441. doi: 10.1007/s11676-022-01475-4
Alcañiz, M., Outeiro, L., Francos, M., Farguell, J., and Úbeda, X. (2016). Long-term dynamics of soil chemical properties after a prescribed fire in a mediterranean forest (montgrí massif, catalonia, Spain). Sci. Total Environ. 572, 1329–1335. doi: 10.1016/j.scitotenv.2016.01.115
Bárcenas-Moreno, G., Jiménez-Compán, E., San Emeterio, L. M., Jiménez-Morillo, N. T., and González-Pérez, J. A. (2022). Soil ph and soluble organic matter shifts exerted by heating affect microbial response. Int. J. Environ. Res. Public Health. 19, 15751. doi: 10.3390/ijerph192315751
Blinkova, O., Raichuk, L., and McDonald, I. (2024). First-year post-fire alterations of scots pine forest: composition, relationships, ecological consequences. Landscape Ecol. 40, 9. doi: 10.1007/s10980-024-02022-8
Bradstock, R. A., Hammill, K. A., Collins, L., and Price, O. (2010). Effects of weather, fuel and terrain on fire severity in topographically diverse landscapes of south-eastern Australia. Landscape Ecol. 25, 607–619. doi: 10.1007/s10980-009-9443-8
Bridges, J. M., Petropoulos, G. P., and Clerici, N. (2019). Immediate changes in organic matter and plant available nutrients of haplic luvisol soils following different experimental burning intensities in damak forest, Hungary. Forests 10, 453. doi: 10.3390/f10050453
Cahojová, L., Jakob, A., Valjavec, M. B., and Carni, A. (2024). Response of vulnerable karst forest ecosystems under different fire severities in the northern dinaric karst mountains (Slovenia). Fire Ecol. 20, 38. doi: 10.1186/s42408-024-00267-x
Chen, Y., Xiong, K., Ren, X., and Cheng, C. (2021). Vulnerability comparison between karst and non-karst nature reserves—with a special reference to guizhou province, China. Sustainability 13, 2442. doi: 10.3390/su13052442
Chen, J., Zhang, L., Cai, X., Luo, W., Lyu, Y., Cheng, A., et al. (2024). The impact of natural vegetation restoration on surface soil moisture of secondary forests and shrubs in the karst region of southwest China. Hydrological Processes 38, e15161. doi: 10.1002/hyp.15161
Connell, J. H. (1978). Diversity in tropical rain forests and coral reefs: high diversity of trees and corals is maintained only in a nonequilibrium state. Science 199, 1302–1310. doi: 10.1126/science.199.4335.1302
Coradini, K., Krejcová, J., and Frouz, J. (2022). Potential of vegetation and woodland cover recovery during primary and secondary succession, a global quantitative review. Land Degradation Dev. 33, 512–526. doi: 10.1002/ldr.4166
D’Ettorre, U. S., Liso, I. S., and Parise, M. (2024). Desertification in karst areas: a review. Earth-Science Rev. 253, 104786. doi: 10.1016/j.earscirev.2024.104786
Da, A., Han, H., Li, H., Li, H. Y., Wu, H. F., and Cheng, X. Q. (2024). Plant-litter-soil stoichiometric characteristics of typical plantation forests and their interaction in loess hilly and gully region. Sci. Soil Water Conserv. 22, 119–128. doi: 10.16843/j.sswc.2024069
De Long, J. R., Fry, E. L., Veen, G. F., and Kardol, P. (2019). Why are plant–soil feedbacks so unpredictable, and what to do about it? Funct. Ecol. 33, 118–128. doi: 10.1111/1365-2435.13232
Eidenshink, J., Schwind, B., Brewer, K., Zhu, Z., Quayle, B., and Howard, S. (2007). A project for monitoring trends in burn severity. Fire Ecol. 3, 3–21. doi: 10.4996/fireecology.0301003
Fernández-García, V., Miesel, J., Baeza, M. J., Marcos, E., and Calvo, L. (2019). Wildfire effects on soil properties in fire-prone pine ecosystems: indicators of burn severity legacy over the medium term after fire. Appl. Soil Ecol. 135, 147–156. doi: 10.1016/j.apsoil.2018.12.002
Gao, R., Ai, N., Liu, G., Liu, C., Qiang, F., Zhang, Z., et al. (2022). The coupling relationship between herb communities and soil in a coal mine reclamation area after different years of restoration. Forests 13, 1481. doi: 10.3390/f13091481
Garousi, F., Shan, Z. J., Ni, K., Yang, H., Shan, J., Cao, J. H., et al. (2021). Decreased inorganic n supply capacity and turnover in calcareous soil under degraded rubber plantation in the tropical karst region. Geoderma 381, 114754. doi: 10.1016/j.geoderma.2020.114754
Geng, B. J., Wang, S. F., Cao, Y. G., Guo, C. Y., Guo, D. G., Zhao, B. Q., et al. (2022). Comparative analysis of vegetation reconstruction characteristics of different years in the reclaimed land of the pingshuo opencast mining area, shanxi province. Acta Ecologica Sin. 42, 3400–3419. doi: 10.5846/stxb202102270548
Georgiadis, G. V. and Georgiadis, T. (2002). Post-fire vegetation succession: the case of aleppo pine (pinus halepensis miller) forests of northern achaia (Greece). Fresenius Environ. Bull. 11, 186–193.
Gilliam, F. S. (2006). Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. J. Ecol. 94, 1176–1191. doi: 10.1111/j.1365-2745.2006.01155.x
González-De Vega, S., de Las Heras, J., and Moya, D. (2018). Post-fire regeneration and diversity response to burn severity in pinus halepensis mill. Forests. Forests 9, 299. doi: 10.3390/f9060299
Hamman, S. T., Burke, I. C., and Stromberger, M. E. (2007). Relationships between microbial community structure and soil environmental conditions in a recently burned system. Soil Biol. Biochem. 39, 1703–1711. doi: 10.1016/j.soilbio.2007.01.018
He, T. H., Lamont, B. B., and Pausas, J. G. (2019). Fire as a key driver of earth’s biodiversity. Biol. Rev. 94, 1983–2010. doi: 10.1111/brv.12544
Holmes, P. M., Richardson, D. M., van Wilgen, B. W., and Gelderblom, C. (2000). Recovery of South African fynbos vegetation following alien woody plant clearing and fire: implications for restoration. Austral Ecol. 25, 631–639. doi: 10.1111/j.1442-9993.2000.tb00069.x
Hou, L., Zhang, Y., Li, Z., Shao, G., Song, L., and Sun, Q. (2021). Comparison of soil properties, understory vegetation species diversities and soil microbial diversities between chinese fir plantation and close-to-natural forest. Forests 12, 632. doi: 10.3390/f12050632
Jiang, Z. C., Li, Z. W., Luo, W. Q., Lan, F. N., and Wu, Z. Y. (2024). Research progress and prospect of soil and water leakage in karst areas of southwest China. Sci. Soil Water Conserv. 22, 1–11. doi: 10.16843/j.sswc.2024085
Jing, J., Li, R., Xiong, L., and Tang, Z. (2025). Binary response of runoff generation and erosion processes to karst vegetation-soil-rock structure and its hydrodynamic mechanisms. J. Hydrology 647, 132319. doi: 10.1016/j.jhydrol.2024.132319
Johnson, D. W., Murphy, J. F., Susfalk, R. B., Caldwell, T. G., Miller, W. W., Walker, R. F., et al. (2005). The effects of wildfire, salvage logging, and post-fire n-fixation on the nutrient budgets of a sierran forest. For. Ecol. Manage. 220, 155–165. doi: 10.1016/j.foreco.2005.08.011
Kardol, P., Yang, T., Arroyo, D. N., and Teste, F. P. (2023). Plant-soil feedback in the ‘real world’: how does fire fit into all of this? Plant Soil 485, 91–102. doi: 10.1007/s11104-022-05778-7
Kazanis, D. and Arianoutsou, M. (1996). Vegetation composition in a post-fire successional gradient of pinus halepensis forests in attica, Greece. Int. J. Wildland Fire 6, 83–91. doi: 10.1071/WF9960083
Keeley, J. E. (2009). Fire intensity, fire severity and burn severity: a brief review and suggested usage. Int. J. Wildland Fire 18, 116–126. doi: 10.1071/WF07049
Kennard, D. K. and Gholz, H. L. (2001). Effects of high- and low-intensity fires on soil properties and plant growth in a Bolivian dry forest. Plant Soil 234, 119–129. doi: 10.1023/A:1010507414994
Knicker, H., González-Vila, F. J., Polvillo, O., González, J. A., and Almendros, G. (2005). Fire-induced transformation of c- and n-forms in different organic soil fractions from a dystric cambisol under a mediterranean pine forest (pinus pinaster). Soil Biol. Biochem. 37, 701–718. doi: 10.1016/j.soilbio.2004.09.008
Lange, M., Eisenhauer, N., Sierra, C. A., Bessler, H., Engels, C., Griffiths, R. I., et al. (2015). Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 6, 6707. doi: 10.1038/ncomms7707
Li, X., Han, Y., Zhang, Y., Shao, Q., Dong, C., Li, J., et al. (2024). Effects of wildfire on soil microbial communities in karst forest ecosystems of southern guizhou province, China. Appl. Environ. Microbiol. 90, e01224–e01245. doi: 10.1128/aem.01245-24
Li, X. Y., Jin, H. J., Wang, H. W., Wu, X. D., Huang, Y. D., He, R. X., et al. (2020). Distributive features of soil carbon and nutrients in permafrost regions affected by forest fires in northern da xing’anling (hinggan) mountains, ne China. Catena 185, 104304. doi: 10.1016/j.catena.2019.104304
Li, Z., Wei, J., He, W., Cao, X., Zhou, X., and Tian, Q. (2024). Effect of plant-soil system on the restoration of community stability after wildfire in the northeast margin of qinghai-tibet plateau. Sci. Rep. 14, 10706. doi: 10.1038/s41598-024-61621-2
Li, S., Wei, X. H., Huang, J. G., Wang, X. Z., Lu, G. Y., and Li, H. X. (2009). The causes and processes responsible for rocky desertification in karst areas of southern China. Sci. Cold Arid Regions 1, 80–90.
Lin, X. Y., Zhang, M., Guo, Y. N., Li, D. D., and Lu, Z. H. (2022). Relationship between understory plant diversity and soil factors in coal mining subsidence area. J. Soil Water Conserv. 36, 268–276. doi: 10.13870/j.cnki.stbcxb.2022.01.035
Liu, Y. B., Li, R. D., and Song, X. F. (2005). Correlation analysis of the coupling between regional urbanization and ecological environment in China. Acta Geographica Sin. 2, 237–247. doi: 10.3321/j.issn:0375-5444.2005.02.007
Liu, W. J., Zeng, F. X., and Jiang, H. (2013). Determination of total nitrogen in solid samples by two-step digestionultraviolet spectrophotometry method. Commun. Soil Sci. Plant Anal. 44, 1080–1091. doi: 10.1080/00103624.2012.750330
López-Poma, R. and Bautista, S. (2014). Plant regeneration functional groups modulate the response to fire of soil enzyme activities in a mediterranean shrubland. Soil Biol. Biochem. 79, 5–13. doi: 10.1016/j.soilbio.2014.08.016
Luo, D., Zhou, Z., Zhang, L., Chen, Q., Huang, D., Feng, Q., et al. (2024). Evolution and driver analysis of forest carbon stocks in karst mountainous areas of southwest China in the context of rocky desertification management. Catena 246, 108335. doi: 10.1016/j.catena.2024.108335
MacDougall, A. S., McCann, K. S., Gellner, G., and Turkington, R. (2013). Diversity loss with persistent human disturbance increases vulnerability to ecosystem collapse. Nature 494, 86–89. doi: 10.1038/nature11869
Mahood, A. L. and Balch, J. K. (2019). Repeated fires reduce plant diversity in low-elevation wyoming big sagebrush ecosystems, (1984–2014). Ecosphere 10, e02591. doi: 10.1002/ECS2.2591
Mayor, A. G., Goirán, S. B., Vallejo, V. R., and Bautista, S. (2016). Variation in soil enzyme activity as a function of vegetation amount, type, and spatial structure in fire-prone mediterranean shrublands. Sci. Total Environ. 573, 1209–1216. doi: 10.1016/j.scitotenv.2016.03.139
McClean, C. J., van den Berg, L., Ashmore, M. R., and Preston, C. D. (2011). Atmospheric nitrogen deposition explains patterns of plant species loss. Global Change Biol. 17, 2882–2892. doi: 10.1111/j.1365-2486.2011.02462.x
Moya, D., González-De Vega, S., García-Orenes, F., Morugán-Coronado, A., Arcenegui, V., Mataix-Solera, J., et al. (2018a). Temporal characterisation of soil-plant natural recovery related to fire severity in burned pinus halepensis mill. Forests. Sci. Total Environ. 640, 42–51. doi: 10.1016/j.scitotenv.2018.05.212
Moya, D., González-De Vega, S., García-Orenes, F., Morugán-Coronado, A., Arcenegui, V., Mataix-Solera, J., et al. (2018b). Temporal characterisation of soil-plant natural recovery related to fire severity in burned pinus halepensis mill. Forests. Sci. Total Environ. 640-641, 42–51. doi: 10.1016/j.scitotenv.2018.05.212
Neary, D. G., Klopatek, C. C., DeBano, L. F., and Ffolliott, P. F. (1999). Fire effects on belowground sustainability: a review and synthesis. For. Ecol. Manage. 122, 51–71. doi: 10.1016/S0378-1127(99)00032-8
Neary, D. G., Ryan, K. C., and Debano, L. F. (2005). Wildland fire in ecosystems: effects of fire on soil and water. Gen. Tech. Rep. 4, 250. doi: 10.2737/RMRS-GTR-42-V4
Neina, D. (2019). The role of soil ph in plant nutrition and soil remediation. Appl. Environ. Soil Sci. 2019, 5794869. doi: 10.1155/2019/5794869
Neri, S. C. M., Bomfim, B., Pereira, R. S., Dos Santos, P. V., and Tetto, A. F. (2023). Decadal fire effects on the structure, composition, diversity, and aboveground carbon stocks of a neotropical savanna. Forests 14, 2294. doi: 10.3390/f14122294
Nie, Y. Y., Xu, L. J., Xin, X. P., Chen, B. R., and Zhang, B. H. (2020). Effects of fence enclosure on the plant community composition and niche characteristics in a temperate meadow steppe. Acta Prataculturae Sin. 29, 11–22. doi: 10.11686/cyxb2020239
Oldroyd, G. and Leyser, O. (2020). A plant’s diet, surviving in a variable nutrient environment. Science 368, 45. doi: 10.1126/science.aba0196
Oliva, M., Maffia, A., Marra, F., Canino, F., Battaglia, S., Mallamaci, C., et al. (2025). The complex impacts of fire on soil ecosystems: insights from the 2021 aspromonte national park wildfire. J. Forestry Res. 36, 68. doi: 10.1007/s11676-025-01864-5
Olsen, S. R. (1954). “Estimation of available phosphorus in soils by extraction with sodium bicarbonate,” in Miscellaneous paper institute for agricultural research samaru. (US: Department of Agriculture).
Pacaldo, R. S., Aydin, M., and Amarille, R. K. (2025). Forest fire and aspects showed no significant effects on most mineral soil properties of black pine forests. Catena 250, 108801. doi: 10.1016/j.catena.2025.108801
Pang, D. B., Cao, J. H., Dan, X. Q., Guan, Y. H., Peng, X. W., Cui, M., et al. (2018). Recovery approach affects soil quality in fragile karst ecosystems of southwest China: implications for vegetation restoration. Ecol. Eng. 123, 151–160. doi: 10.1016/j.ecoleng.2018.09.001
Pei, J. M., Wan, J. R., Wang, H., Fang, C. M., Nie, M., and Li, J. Q. (2023). Changes in the activity of soil enzymes after fire. Geoderma 437, 7. doi: 10.1016/j.geoderma.2023.116599
Pellegrini, A. F. A., Caprio, A. C., Georgiou, K., Finnegan, C., Hobbie, S. E., Hatten, J. A., et al. (2021). Low-intensity frequent fires in coniferous forests transform soil organic matter in ways that may offset ecosystem carbon losses. Global Change Biol. 27, 3810–3823. doi: 10.1111/gcb.15648
Qiang, W., He, L., Zhang, Y., Liu, B., Liu, Y., Liu, Q., et al. (2021). Aboveground vegetation and soil physicochemical properties jointly drive the shift of soil microbial community during subalpine secondary succession in southwest China. Catena 202, 105251. doi: 10.1016/j.catena.2021.105251
Quigley, K. M., Kolka, R., Sturtevant, B. R., Dickinson, M. B., Kern, C. C., Donner, D. M., et al. (2020). Prescribed burn frequency, vegetation cover, and management legacies influence soil fertility: implications for restoration of imperiled pine barrens habitat. For. Ecol. Manage. 470-471, 118163. doi: 10.1016/j.foreco.2020.118163
Raison, R. J. (1979). Modification of the soil environment by vegetation fires, with particular reference to nitrogen transformations: a review. Plant Soil 51, 73–108. doi: 10.1007/BF02205929
Rebi, A., Wang, G., Ejaz, I., Flynn, T., Kanomanyanga, J., Yang, T., et al. (2025). Fire and clipping drive microbial fixation pathways in soil phosphorus and sulfur cycling in China’s key karst region. Eur. J. Soil Biol. 124, 103707. doi: 10.1016/j.ejsobi.2024.103707
Reinhart, K. O., Dangi, S. R., and Vermeire, L. T. (2016). The effect of fire intensity, nutrients, soil microbes, and spatial distance on grassland productivity. Plant Soil 409, 203–216. doi: 10.1007/s11104-016-2957-3
Salgado, L., Alvarez, M. G., Díaz, A. M., Gallego, J. R., and Forján, R. (2024). Impact of wildfire recurrence on soil properties and organic carbon fractions. J. Environ. Manage. 354, 120293. doi: 10.1016/j.jenvman.2024.120293
Sciences, C.A.O.A (1988). Method for determination of soil total phosphorus, ny/t 88-1988 (Beijing: China Standard Press, Beijing).
Smith, D. W. (1970). Concentrations of soil nutrients before and after fire. Can. J. Soil Sci. 50, 17–29. doi: 10.4141/cjss70-003
Stancic, L. and Repe, B. (2018). Post-fire succession: selected examples from the karst region, southwest Slovenia. Acta Geographica Slovenica-Geografski Zbornik 58, 27–38. doi: 10.3986/AGS.1942
State Forestry Administration (1999a). Determination of forest soil water-related physical properties. Forest soil analysis methods, ly/t 1215–1999 (Beijing: China Standard Press).
Tian, X., Zhao, F., Shu, L., and Wang, M. (2014). Changes in forest fire danger for south-western China in the 21st century. Int. J. Wildland Fire 23, 185–195. doi: 10.1071/WF13014
Toberman, H., Chen, C. R., Lewis, T., and Elser, J. J. (2014). High-frequency fire alters c: n: p stoichiometry in forest litter. Global Change Biol. 20, 2321–2331. doi: 10.1111/gcb.12432
Vallejo, V. R., Arianoutsou, M., and Moreira, F. (2012). Fire ecology and post-fire restoration approaches in southern european forest types. Post-Fire Manage. Restor. South. Eur. Forests 24, 93–119. doi: 10.1007/978-94-007-2208-8_5
Verma, S. and Jayakumar, S. (2012). Impact of forest fire on physical, chemical and biological properties of soil: a review. Proc. Int. Acad. Ecol. Environ. Sci. 2, 168–176.
Verma, S., Singh, D., Singh, A. K., and Jayakumar, S. (2019). Post-fire soil nutrient dynamics in a tropical dry deciduous forest of western ghats, India. For. Ecosyst. 6, 6. doi: 10.1186/s40663-019-0168-0
Waldrop, T. A. and Brose, P. H. (1999). A comparison of fire intensity levels for stand replacement of table mountain pine (pinus pungens lamb.). For. Ecol. Manage. 113, 155–166. doi: 10.1016/S0378-1127(98)00422-8
Wang, M. H., Cui, F. K., Wu, Z. P., Xu, X. Y., Ou, S. K., Chen, J. H., et al. (2024). The pattern of understory plant diversity and soil physicochemical properties in the severely burned areas of forests. Forestry Environ. Sci. 40, 1–8. doi: 10.3969/j.issn.1006-4427.2024.04.001
Wang, S. J., Liu, Q. M., and Zhang, D. F. (2004). Karst rocky desertification in southwestern China: geomorphology, landuse, impact and rehabilitation. Land Degradation Dev. 15, 115–121. doi: 10.1002/ldr.592
Wang, G., Zhu, T., Zhou, J., Yu, Y., Petropoulos, E., and Müller, C. (2022). Slash-and-burn in karst regions lowers soil gross nitrogen (n) transformation rates and n-turnover. Geoderma 425, 116084. doi: 10.1016/j.geoderma.2022.116084
Wieczorkowski, J. D., Lehmann, C. E. R., Archibald, S., Banda, S., Goyder, D. J., Kaluwe, M., et al. (2024). Fire facilitates ground layer plant diversity in a miombo ecosystem. Ann. Bot. 133, 743–756. doi: 10.1093/aob/mcae035
Wu, S., Huang, B., Huang, C., Li, G., and Liao, P. (2018). The aboveground vegetation type and underground soil property mediate the divergence of soil microbiomes and the biological interactions. Microbial Ecol. 75, 434–446. doi: 10.1007/s00248-017-1050-7
Xie, F. J., Xiao, D. J., Li, X. J., Wang, X. G., and Xia, S. H. (2005). Forest landscape restoration assessment based on ndvi under different burn intensity in the burned blank of daxinganling mountains. Chin. J. Ecol. 4, 368–372. doi: 10.13292/j.1000-4890.2005.0270
Xu, Y., Sun, J., Lin, Q., Ma, J., Shi, Y., and Lou, K. (2012). Effects of a surface wildfire on soil nutrient and microbial functional diversity in a shrubbery. Acta Ecologica Sin. 32, 258–264. doi: 10.1016/j.chnaes.2012.07.007
Zhang, Y. L., Guo, Y., and Hu, H. Q. (2021). Characteristics of forest fire data in southwest China during 2001-2017. J. Northwest Forestry Univ. 36, 179–186. doi: 10.3969/j.issn.1001-7461.2021.01.26
Zhang, Z. and Huisingh, D. (2018). Combating desertification in China: monitoring, control, management and revegetation. J. Cleaner Production 182, 765–775. doi: 10.1016/j.jclepro.2018.01.233
Zhang, Y. H., Tan, B. X., Sun, M. L., and Zhou, Z. Q. (2012). Impact of forest fire on understory vegetation and soil in typical forest types of daxing’an mountains,northeastern China. J. Beijing Forestry University. 34, 7–13. doi: 10.13332/j.1000-1522.2012.02.009
Zhou, N., Wang, L., Mao, X. F., Wei, X. Y., Gao, Y. Y., Yu, H. Y., et al. (2025). Characteristics of plant community and soil and their relationship research in degraded alpine marsh wetlands in qinghai qilian mountain national park. Acta Agrestia Sin. 33, 1114–1126. doi: 10.11733/j.issn.1007-0435.2025.04.011
Keywords: karst ecosystems, post-fire disturbances, grey relational analysis, vegetationrestoration, soil drivers, coupling coordination, environmental stress
Citation: Zhang L, Zhang Z, Li Z, Chen X, Pu S, Chen Q, Gong M, Khan MA and Zhou J (2025) Herb-soil coupling in post-fire karst forests: a grey relational analysis in Yunnan, Southwest China. Front. Plant Sci. 16:1709599. doi: 10.3389/fpls.2025.1709599
Received: 20 September 2025; Accepted: 10 November 2025; Revised: 01 November 2025;
Published: 26 November 2025.
Edited by:
Peilei Hu, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Yongkuan Chi, Guizhou Normal University, ChinaChaoyi Luo, Northwest A&F University, China
Copyright © 2025 Zhang, Zhang, Li, Chen, Pu, Chen, Gong, Khan and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jinxing Zhou, YmpmdXpqeEAxMjYuY29t